IRF8 - IRF8





Регуляторный фактор интерферона 8 (IRF8), также известный как белок, связывающий консенсусную последовательность интерферона (ICSBP), является белок что у людей кодируется IRF8 ген.[5][6][7] IRF8 - это фактор транскрипции который играет решающую роль в регулировании приверженности к родословной и в миелоидный ячейка созревание, включая решение о том, что общий миелоидный предшественник (CMP) различать в моноцит клетка-предшественник.
Функция
Белок, связывающий консенсусную последовательность интерферона (ICSBP) - это фактор транскрипции из интерферон регуляторный фактор (IRF ) семья. Белки этого семейства состоят из консервативного ДНК-связывающий домен в N-концевой регион и расходящиеся C-терминал регион, который служит регулирующим доменом. Белки семейства IRF связываются с IFN-стимулированным элемент ответа (ISRE) и регулируют экспрессию генов, стимулируемых IFN типа I, а именно IFN-α и IFN-β. Белки семейства IRF также контролируют экспрессию IFN-α и IFN-β-регулируемых генов, которые индуцируются вирусной инфекцией.[5]
Нокаут-исследования
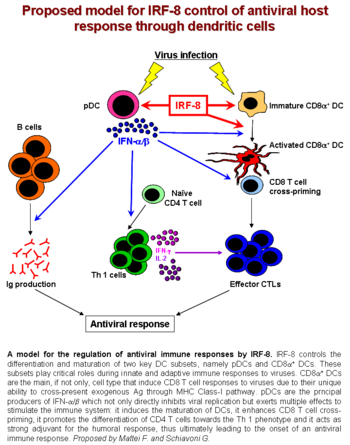
IFN-продуцирующие клетки (mIPC) отсутствовали во всех лимфоидных органах от ICSBP. выбить (KO) мышей, о чем свидетельствует отсутствие CD11cнизкийB220+Ly6C+CD11b− клетки. Параллельно CD11c+ клетки, выделенные из селезенки ICSBP KO, были неспособны продуцировать IFN типа I в ответ на вирусную стимуляцию. Мыши ICSBP KO также продемонстрировали заметное снижение подмножества DC, экспрессирующего маркер CD8alpha (CD8alpha+ ДК) в селезенке, лимфатических узлах и тимусе. Более того, ICSBP-дефицитный CD8alpha+ ДК проявляли заметно нарушенный фенотип по сравнению с ДК дикого типа. Они экспрессировали очень низкие уровни костимулирующих молекул (молекула межклеточной адгезии ICAM1, CD40, CD80, CD86 ) и хемокинового рецептора Т-клетки CCR7.[8]
Клиническое значение
В миелоидных клетках IRF8 регулирует экспрессию Bax и Фас регулировать апоптоз.[9] В хронический миелолейкоз (CML), IRF8 регулирует кислотность керамидаза опосредовать апоптоз ХМЛ.[10]
IRF8 высоко экспрессируется в миелоидных клетках и первоначально был идентифицирован как критический клон-специфический фактор транскрипции для дифференцировки миелоидных клеток,[11] Однако недавние исследования показали, что IRF8 также конститутивно экспрессируется в не-кроветворный раковые клетки, хотя и на более низком уровне. Кроме того, IRF8 также может активироваться IFN-γ в негемопоэтических клетках. IRF8 опосредует экспрессию Fas, Bax, КУВЫРОК, Jak1 и STAT1 опосредовать апоптоз негемотопоэтических раковых клеток.[12][13][14]
Анализ базы данных геномики рака человека показал, что IRF8 не сильно очагово амплифицируется по всему набору данных из 3131 опухоли, но значительно удален во всем наборе данных из 3131 опухоли, что позволяет предположить, что IRF8 потенциально является подавитель опухолей в людях.[15] Молекулярный анализ показал, что промотор гена IRF8 гиперметилирован у человека. карцинома толстой кишки клетки[14][16] предполагая, что эти клетки могут использовать метилирование ДНК, чтобы подавить экспрессию IRF8, чтобы продвинуть болезнь.
Взаимодействия
IRF8 был показан взаимодействовать с участием IRF1[17][18] и COPS2.[19]
Смотрите также
использованная литература
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000140968 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000041515 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б «Энтрез Ген: фактор 8 регуляции интерферона IRF8».
- ^ Weisz A, Marx P, Sharf R, Appella E, Driggers PH, Ozato K, Levi BZ (декабрь 1992 г.). «Белок, связывающий консенсусную последовательность человеческого интерферона, является негативным регулятором энхансерных элементов, общих для генов, индуцируемых интерфероном». J. Biol. Chem. 267 (35): 25589–96. PMID 1460054.
- ^ Nehyba J, Hrdlicková R, Burnside J, Bose HR (июнь 2002 г.). «Новый регуляторный фактор интерферона (IRF), IRF-10, играет уникальную роль в иммунной защите и индуцируется онкопротеином v-Rel». Мол. Cell. Биол. 22 (11): 3942–57. Дои:10.1128 / MCB.22.11.3942-3957.2002. ЧВК 133824. PMID 11997525.
- ^ Тамура Т., Озато К. (январь 2002 г.). «ICSBP / IRF-8: его регуляторные роли в развитии миелоидных клеток». J. Интерферон цитокин Res. 22 (1): 145–52. Дои:10.1089/107999002753452755. PMID 11846985.
- ^ Ян Дж., Ху X, Циммерман М., Торрес С.М., Ян Д., Смит С.Б., Лю К. (ноябрь 2011 г.). «Передний край: IRF8 регулирует транскрипцию Bax in vivo в первичных миелоидных клетках». J. Immunol. 187 (9): 4426–30. Дои:10.4049 / jimmunol.1101034. ЧВК 3197864. PMID 21949018.
- ^ Ху X, Ян Д., Циммерман М., Лю Ф., Ян Дж., Каннан С., Бурхерт А., Шульц З., Белавска А., Озато К., Бхалла К., Лю К. (апрель 2011 г.). «IRF8 регулирует экспрессию кислой церамидазы, опосредуя апоптоз и подавляя миелогенный лейкоз». Рак Res. 71 (8): 2882–91. Дои:10.1158 / 0008-5472.CAN-10-2493. ЧВК 3078194. PMID 21487040.
- ^ Holtschke T, Löhler J, Kanno Y, Fehr T., Giese N, Rosenbauer F, Lou J, Knobeloch KP, Gabriele L, Waring JF, Bachmann MF, Zinkernagel RM, Morse HC, Ozato K, Horak I (октябрь 1996). «Иммунодефицит и хронический миелолейкозоподобный синдром у мышей с целевой мутацией гена ICSBP». Ячейка. 87 (2): 307–17. Дои:10.1016 / S0092-8674 (00) 81348-3. PMID 8861914. S2CID 18065448.
- ^ Ян Д., Ван С., Брукс С., Донг З., Шенлейн П.В., Кумар В., Оуян Х, Сюн Х., Лахат Г., Хейс-Джордан А., Лазар А., Поллок Р., Лев Д., Лю К. (февраль 2009 г.). «Регуляторный фактор 8 IFN сенсибилизирует клетки саркомы мягких тканей к апоптозу, инициированному рецептором смерти, посредством подавления экспрессии FLICE-подобного белка». Рак Res. 69 (3): 1080–8. Дои:10.1158 / 0008-5472.CAN-08-2520. ЧВК 2633427. PMID 19155307.
- ^ Ян Д., Тхангараджу М., Браунинг Д. Д., Донг З., Корчин Б., Лев Д. К., Ганапати В., Лю К. (октябрь 2007 г.). «Регуляторный фактор 8 IFN опосредует апоптоз негемопоэтических опухолевых клеток посредством регуляции экспрессии Fas». J. Immunol. 179 (7): 4775–82. Дои:10.4049 / jimmunol.179.7.4775. PMID 17878376.
- ^ а б Ян Д., Тангараджу М., Гринелч К., Браунинг Д. Д., Шенлейн П. В., Тамура Т., Озато К., Ганапати В., Абрамс С. И., Лю К. (апрель 2007 г.). «Подавление регуляторного фактора 8 IFN метилированием ДНК является молекулярной детерминантой устойчивости к апоптозу и метастатического фенотипа в метастатических опухолевых клетках». Рак Res. 67 (7): 3301–9. Дои:10.1158 / 0008-5472.CAN-06-4068. PMID 17409439.
- ^ «Туморскейп». Институт Броуда. Архивировано из оригинал на 2012-04-14. Получено 2012-07-05.
- ^ McGough JM, Yang D, Huang S, Georgi D, Hewitt SM, Röcken C, Tänzer M, Ebert MP, Liu K (декабрь 2008 г.). «Метилирование ДНК подавляет индуцированную IFN-гамма и сигнальный преобразователь и активатор активации регулирующего фактора 8 IFN, опосредованной транскрипцией 1, в клетках карциномы толстой кишки». Мол. Рак Res. 6 (12): 1841–51. Дои:10.1158 / 1541-7786.MCR-08-0280. ЧВК 2605678. PMID 19074829.
- ^ Шапер Ф, Кирхгоф С., Позерн Г., Кёстер М., Умар А., Шарф Р., Леви Б. З., Хаузер Х (октябрь 1998 г.). «Функциональные домены фактора регуляции интерферона I (IRF-1)». Biochem. J. 335 (1): 147–57. Дои:10.1042 / bj3350147. ЧВК 1219763. PMID 9742224.
- ^ Шарф Р., Азриэль А., Лейбкович Ф., Виноград С.С., Эрлих Р., Леви Б.З. (июнь 1995 г.). «Анализ функционального домена белка, связывающего консенсусную последовательность интерферона (ICSBP), и его связь с факторами регуляции интерферона». J. Biol. Chem. 270 (22): 13063–9. Дои:10.1074 / jbc.270.22.13063. PMID 7768900.
- ^ Коэн Х., Азриэль А., Коэн Т., Мераро Д., Хашмуэли С., Бех-Отсчир Д., Крафт Р., Дубиль В., Леви Б.З. (декабрь 2000 г.). «Взаимодействие между связывающим консенсусную последовательность интерферона белком и COP9 / субъединицей CSN2 (Trip15). Возможная связь между передачей сигналов фактора регуляции интерферона и COP9 / сигнаносомой». J. Biol. Chem. 275 (50): 39081–9. Дои:10.1074 / jbc.M004900200. PMID 10991940.
Иллюстрации
дальнейшее чтение
- Weisz A, Marx P, Sharf R, Appella E, Driggers PH, Ozato K, Levi BZ (1993). «Связывающий консенсусную последовательность человеческого интерферона белок является негативным регулятором энхансерных элементов, общих для генов, индуцируемых интерфероном». J. Biol. Chem. 267 (35): 25589–96. PMID 1460054.
- Шарф Р., Азриэль А., Лейбкович Ф., Виноград С.С., Эрлих Р., Леви Б.З. (1995). «Анализ функционального домена белка, связывающего консенсусную последовательность интерферона (ICSBP), и его связь с факторами регуляции интерферона». J. Biol. Chem. 270 (22): 13063–9. Дои:10.1074 / jbc.270.22.13063. PMID 7768900.
- Holtschke T, Löhler J, Kanno Y, Fehr T., Giese N, Rosenbauer F, Lou J, Knobeloch KP, Gabriele L, Waring JF, Bachmann MF, Zinkernagel RM, Morse HC, Ozato K, Horak I. (1996). «Иммунодефицит и хронический миелолейкозоподобный синдром у мышей с целевой мутацией гена ICSBP». Ячейка. 87 (2): 307–17. Дои:10.1016 / S0092-8674 (00) 81348-3. PMID 8861914. S2CID 18065448.
- Шапер Ф, Кирхгоф С., Позерн Г., Кёстер М., Умар А., Шарф Р., Леви Б. З., Хаузер Х (1998). «Функциональные домены фактора регуляции интерферона I (IRF-1)». Biochem. J. 335 (1): 147–57. Дои:10.1042 / bj3350147. ЧВК 1219763. PMID 9742224.
- Ли В., Нагинени С.Н., Ге Х., Эфиок Б., Чепелинский А.Б., Эгвуагу С.Э. (1999). «Белок, связывающий консенсусную последовательность интерферона, конститутивно экспрессируется и дифференциально регулируется в хрусталике глаза». J. Biol. Chem. 274 (14): 9686–91. Дои:10.1074 / jbc.274.14.9686. PMID 10092656.
- Ли В., Нагинени К.Н., Хукс Дж. Дж., Чепелинский А.Б., Эгвуагу К.Э. (1999). «Передача сигналов гамма-интерферона в пигментных эпителиальных клетках сетчатки человека, опосредованная факторами транскрипции STAT1, ICSBP и IRF-1». Вкладывать деньги. Офтальмол. Vis. Наука. 40 (5): 976–82. PMID 10102295.
- Rehli M, Poltorak A, Schwarzfischer L, Krause SW, Andreesen R, Beutler B (2000). «PU.1 и белок, связывающий консенсусную последовательность интерферона, регулируют миелоидную экспрессию гена человеческого Toll-подобного рецептора 4». J. Biol. Chem. 275 (13): 9773–81. Дои:10.1074 / jbc.275.13.9773. PMID 10734131.
- Коэн Х., Азриэль А., Коэн Т., Мераро Д., Хашмуэли С., Бех-Отсчир Д., Крафт Р., Дубиль В., Леви Б.З. (2001). «Взаимодействие между связывающим консенсусную последовательность интерферона белком и COP9 / субъединицей CSN2 (Trip15). Возможная связь между передачей сигналов фактора регуляции интерферона и COP9 / сигнаносомой». J. Biol. Chem. 275 (50): 39081–9. Дои:10.1074 / jbc.M004900200. PMID 10991940.
- Nehyba J, Hrdlicková R, Burnside J, Bose HR (2002). «Новый регуляторный фактор интерферона (IRF), IRF-10, играет уникальную роль в иммунной защите и индуцируется онкопротеином v-Rel». Мол. Cell. Биол. 22 (11): 3942–57. Дои:10.1128 / MCB.22.11.3942-3957.2002. ЧВК 133824. PMID 11997525.
- Чжу Ц., Сабервал Г, Лу И, Платаниас Л.С., Эклунд Э.А. (2005). «Белок, связывающий консенсусную последовательность интерферона, активирует транскрипцию гена, кодирующего нейрофибромин 1». J. Biol. Chem. 279 (49): 50874–85. Дои:10.1074 / jbc.M405736200. PMID 15371411.
- Лю Дж, Гуань Х, Тамура Т., Озато К., Ма Х (2005). «Синергетическая активация транскрипции гена интерлейкина-12 p35 с помощью фактора регуляции интерферона-1 и белка, связывающего консенсусную последовательность интерферона». J. Biol. Chem. 279 (53): 55609–17. Дои:10.1074 / jbc.M406565200. PMID 15489234.
- Барриос-Родилес М., Браун К.Р., Оздамар Б., Бозе Р., Лю З., Донован Р.С., Синджо Ф., Лю Ю., Дембовы Дж., Тейлор И. В., Луга В., Прзуль Н., Робинсон М., Сузуки Н., Хаяшизаки Ю., Юрисика И., Врана JL (2005). «Высокопроизводительное отображение динамической сигнальной сети в клетках млекопитающих». Наука. 307 (5715): 1621–5. Bibcode:2005Научный ... 307.1621B. Дои:10.1126 / science.1105776. PMID 15761153. S2CID 39457788.
- Xiong H, Li H, Kong HJ, Chen Y, Zhao J, Xiong S, Huang B, Gu H, Mayer L, Ozato K, Unkeless JC (2005). «Убиквитин-зависимая деградация фактора регуляции интерферона-8, опосредованная Cbl, подавляет экспрессию интерлейкина-12». J. Biol. Chem. 280 (25): 23531–9. Дои:10.1074 / jbc.M414296200. PMID 15837792.
- Чжао Дж., Конг Х.Дж., Ли Х., Хуан Б., Ян М., Чжу С., Богунович М., Чжэн Ф., Майер Л., Озато К., Ункелесс Дж., Сюн Х. (2006). «Консенсусный белок IRF-8 / интерферон (IFN), связывающий консенсусную последовательность, участвует в передаче сигналов Toll-подобного рецептора (TLR) и способствует перекрестной связи между сигнальными путями TLR и IFN-gamma». J. Biol. Chem. 281 (15): 10073–80. Дои:10.1074 / jbc.M507788200. PMID 16484229.
- Хуанг В., Сабервал Г., Хорват Е., Чжу С., Линдси С., Эклунд Е.А. (2006). «Связанные с лейкемией, конститутивно активные мутанты тирозинфосфатазы белка SHP2 ингибируют активацию транскрипции NF1 белком, связывающим консенсусную последовательность интерферона». Мол. Cell. Биол. 26 (17): 6311–32. Дои:10.1128 / MCB.00036-06. ЧВК 1592828. PMID 16914719.
- Димберг А., Карихед К., Нильссон К., Оберг Ф (2006). «Ингибирование дифференцировки моноцитов с помощью Stat1 с дефицитом фосфорилирования связано с нарушением экспрессии Stat2, ICSBP / IRF8 и C / EBPepsilon». Сканд. J. Immunol. 64 (3): 271–9. Дои:10.1111 / j.1365-3083.2006.01827.x. PMID 16918696.
- Леонард Д., Свенунгссон Э., Сэндлинг Дж. К., Берггрен О., Йонсен А., Бенгтссон К., Ван С., Йенсен-Урстад К., Гранстам С. О., Бенгтссон А. А., Густафссон Ю. Т., Гуннарссон И., Рантапя-Дальквист С., Нордмарк Г., Элоранта М. Л., Сивянта AC, Рённблом Л. (2013). "Ишемическая болезнь сердца при системной красной волчанке связана с вариантами гена фактора регуляции 8 интерферона". Circ. Кардиоваск. Genet. 6 (3): 255–63. Дои:10.1161 / CIRCGENETICS.113.000044. PMID 23661672.
- Пэн Ю.Б., Йерле М., Лю Б. (2007). «Картирование девяти генов фактора регуляции интерферона свиней». Anim. Genet. 37 (6): 600–1. Дои:10.1111 / j.1365-2052.2006.01525.x. PMID 17121613.
- Хуанг В., Хорват Э., Эклунд Э.А. (2007). «PU.1, фактор регуляции интерферона (IRF) 2 и белок, связывающий консенсусную последовательность интерферона (ICSBP / IRF8), взаимодействуют для активации транскрипции NF1 в дифференцирующихся миелоидных клетках». J. Biol. Chem. 282 (9): 6629–43. Дои:10.1074 / jbc.M607760200. PMID 17200120.
- Маттей Ф., Скьявони Дж., Борги П., Вендитти М., Канини I, Сестили П., Пьетрафорте I, Морс ХК, Рамони С., Беларделли Ф., Габриэле Л. (2006). «ICSBP / IRF-8 по-разному регулирует захват антигена во время развития дендритных клеток и влияет на презентацию антигена CD4 + Т-клеткам». Кровь. 108 (2): 609–617. Дои:10.1182 / кровь-2005-11-4490. PMID 16569763.
- Скьявони Г., Маттей Ф., Сестили П., Борги П., Вендитти М., Морс ХК, Беларделли Ф., Габриэле Л. (2002). «ICSBP необходим для развития клеток, продуцирующих интерферон I типа мыши, а также для генерации и активации дендритных клеток CD8alpha (+)». J. Exp. Med. 196 (11): 1415–1425. Дои:10.1084 / jem.20021263. ЧВК 2194263. PMID 12461077.
внешние ссылки
- IRF8 + белок, + человек в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
Эта статья включает текст из Национальная медицинская библиотека США, который находится в всеобщее достояние.