Fas рецептор - Fas receptor
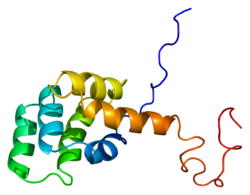




В Fas рецептор, также известный как Фас, FasR, антиген апоптоза 1 (АПО-1 или APT), кластер дифференциации 95 (CD95) или суперсемейство рецепторов фактора некроза опухолей член 6 (TNFRSF6), это белок что у людей кодируется ФАС ген.[5][6] Fas был впервые идентифицирован с помощью моноклональное антитело генерируется иммунизацией мышей клеточной линией FS-7. Таким образом, название Fas происходит от FS-7-асвязанный surface антиген.[7]
Рецептор Fas - это рецептор смерти на поверхности клеток, что приводит к запрограммированной гибели клеток (апоптоз ) если он связывает свой лиганд, Fas лиганд (FasL). Это один из двух путей апоптоза, другой - митохондриальный.[8]
Ген
Ген рецептора FAS расположен на длинном плече хромосома 10 (10q24.1) у людей и на хромосоме 19 у мышей. Ген лежит на плюсе (Уотсон нити ) и имеет длину 25 255 оснований, организованных в девять белков, кодирующих экзоны. Подобные последовательности, связанные эволюцией (ортологи )[9] находятся в большинстве млекопитающие.
Протеин
Предыдущие отчеты идентифицировали целых восемь вариантов сварки, которые переведены в семь изоформы белка. Рецептор Fas, вызывающий апоптоз, получил название изоформы 1 и относится к типу 1. трансмембранный белок. Многие другие изоформы встречаются редко. гаплотипы которые обычно связаны с болезненным состоянием. Однако две изоформы, индуцирующая апоптоз мембраносвязанная форма и растворимая форма, являются нормальными продуктами, производство которых через альтернативное сращивание регулируется цитотоксическим РНК-связывающим белком TIA1.[10]
Зрелый белок Fas состоит из 319 аминокислот, имеет прогнозируемую молекулярную массу 48 килодальтон и разделен на 3 домена: внеклеточный домен, трансмембранный домен и цитоплазматический домен. Внеклеточный домен состоит из 157 аминокислот и богат остатками цистеина. Трансмембранный и цитоплазматический домены содержат 17 и 145 аминокислот соответственно. Экзоны с 1 по 5 кодируют внеклеточную область. Экзон 6 кодирует трансмембранную область. Экзоны 7-9 кодируют внутриклеточную область.
Функция
Fas формирует сигнальный комплекс, вызывающий смерть (DISC) при связывании лиганда. С мембраной Fas лиганд тример на поверхности соседней клетки вызывает олигомеризацию Fas. Недавние исследования показали, что тримеризация Fas не может быть подтверждена. Другие модели предполагали олигомеризацию до 5-7 молекул Fas в DISC.[11]Это событие также имитируется связыванием агонистического антитела Fas, хотя некоторые данные свидетельствуют о том, что апоптотический сигнал, индуцированный антителом, ненадежен при изучении передачи сигналов Fas. С этой целью было использовано несколько умных способов тримеризации антитела для исследования in vitro.
После последующей агрегации домена смерти (DD) рецепторный комплекс интернализуется через клеточную эндосомный машины. Это позволяет адаптерная молекула FADD связать домен смерти Фас через его собственный домен смерти.[12]
FADD также содержит эффекторный домен смерти (DED) рядом с его амино-концом,[13] который облегчает связывание с DED FADD-подобного бета-превращающего фермента интерлейкина-1 (FLICE), более часто называемого каспаза-8. Затем FLICE может автоматически активироваться через протеолитическое расщепление на субъединицы p10 и p18, каждая из которых образует активный гетеротетрамерный фермент. Затем активная каспаза-8 высвобождается из DISC в цитозоль, где она расщепляет другие эффекторные каспазы, что в конечном итоге приводит к деградации ДНК, образованию пузырей на мембранах и другим признакам апоптоза.
Недавно было показано, что Fas способствует росту опухоли, поскольку во время прогрессирования опухоли он часто подавляется или клетки становятся устойчивыми к апоптозу. Раковые клетки в целом, независимо от их чувствительности к апоптозу Fas, зависят от конститутивной активности Fas. Это стимулируется производимым раком лигандом Fas для оптимального роста.[14]
Хотя было показано, что Fas способствует росту опухоли на вышеупомянутых моделях мышей, анализ базы данных геномики рака человека показал, что FAS не подвергается значительной фокусной амплификации в наборе данных из 3131 опухоли (FAS не является онкоген ), но значительно удален из всего набора данных 3131 опухолей,[15] предполагая, что ФАС функционирует как подавитель опухолей в людях.
В культивируемых клетках FasL вызывает различные типы апоптоза раковых клеток через рецептор Fas. На моделях мышей с AOM-DSS-индуцированной карциномой толстой кишки и MCA-индуцированной саркомой было показано, что Fas действует как опухолевый супрессор.[16] Кроме того, рецептор Fas также опосредует противоопухолевую цитотоксичность опухолеспецифических цитотоксических Т-лимфоцитов (CTL).[17]
Роль в апоптозе
В некоторых сообщениях предполагается, что внешнего пути Fas достаточно для индукции полного апоптоз в определенных типах клеток посредством сборки DISC и последующей активации каспазы-8. Эти клетки называются клетками типа 1 и характеризуются неспособностью антиапоптотических членов семейства Bcl-2 (а именно Bcl-2 и Bcl-xL) защищать от Fas-опосредованного апоптоза. Характерные клетки типа 1 включают H9, CH1, SKW6.4 и SW480, все из которых являются линиями лимфоцитов, за исключением последней, которая является линией аденокарциномы толстой кишки. Однако доказательства в пользу перекрестные помехи Между внешним и внутренним путями существует сигнальный каскад Fas.
В большинстве типов клеток каспаза-8 катализирует расщепление проапоптотического белка BH3. Ставка в усеченном виде, tBid. Только BH-3 члены семейства Bcl-2 задействуют исключительно антиапоптотических членов семейства (Bcl-2, Bcl-xL ), позволяя Бак и Bax перемещаться на внешнюю митохондриальную мембрану, тем самым повышая ее проницаемость и облегчая высвобождение проапоптотических белков, таких как цитохром с и Smac / DIABLO, антагонист ингибиторы апоптоза белки (ИПБ).
Взаимодействия
Было показано, что рецептор Fas взаимодействовать с:
- Каспаза 8,[18][19][20]
- Каспаза 10,[21]
- CFLAR,[19][20]
- FADD,[18][19][22][23][24][25]
- Fas лиганд,[18][26][27][28]
- PDCD6,[29] и
- Малый модификатор, связанный с убиквитином 1.[30][31]
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000026103 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000024778 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ссылка на Mouse PubMed:». Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Lichter P, Walczak H, Weitz S, Behrmann I, Krammer PH (сентябрь 1992 г.). «Антиген APO-1 человека (APT) отображается на 10q23, область, которая синтенична хромосоме 19 мыши». Геномика. 14 (1): 179–80. Дои:10.1016 / S0888-7543 (05) 80302-7. PMID 1385299.
- ^ Инадзава Дж., Ито Н., Абэ Т., Нагата С. (ноябрь 1992 г.). «Отнесение гена антигена Fas человека (Fas) к 10q24.1». Геномика. 14 (3): 821–2. Дои:10.1016 / S0888-7543 (05) 80200-9. PMID 1385309.
- ^ Нагата S (июль 2004 г.). «Ранняя работа над функцией CD95, интервью с Шиге Нагата». Гибель клеток и дифференциация. 11 Дополнение 1 (Дополнение 1): S23-7. Дои:10.1038 / sj.cdd.4401453. PMID 15143352.
- ^ Ваджант Х (2002). «Путь передачи сигналов Fas: больше, чем парадигма». Наука. 296 (5573): 1635–6. Дои:10.1126 / science.1071553. PMID 12040174. S2CID 29449108.
- ^ «Филогенетический маркер OrthoMaM: кодирующая последовательность FAS». Архивировано из оригинал на 2016-03-03. Получено 2009-12-02.
- ^ Искьердо Дж. М., Майос Н., Боннал С., Мартинес К., Каштелу Р., Гиго Р., Бильбао Д., Валькарсель Дж. (Август 2005 г.). «Регулирование альтернативного сплайсинга Fas за счет антагонистических эффектов TIA-1 и PTB на определение экзона». Мол. Клетка. 19 (4): 475–84. Дои:10.1016 / j.molcel.2005.06.015. PMID 16109372.
- ^ Ванга; и другие. (2010). «Комплексная структура домена смерти Fas – FADD раскрывает основу сборки DISC и болезненных мутаций». Нат Структ Мол Биол. 17 (11): 1324–29. Дои:10.1038 / нсмб.1920. ЧВК 2988912. PMID 20935634.
- ^ Хуанг Б; и другие. (1996). «Структура ЯМР и мутагенез домена смерти Fas (APO-1 / CD95)». Природа. 384 (6610): 638–41. Дои:10.1038 / 384638a0. PMID 8967952. S2CID 2492303.
- ^ Eberstadt M; и другие. (1998). «Структура ЯМР и мутагенез эффекторного домена смерти FADD (Mort1)». Природа. 392 (6679): 941–5. Дои:10.1038/31972. PMID 9582077. S2CID 4370202.
- ^ Чен Л., Парк С. М., Туманов А. В., Хау А., Савада К., Фейг С., Тернер Дж. Р., Фу YX, Ромеро ИЛ, Лендьель Е., Питер М.Э. (май 2010 г.). «CD95 способствует росту опухоли». Природа. 465 (7297): 492–6. Дои:10.1038 / природа09075. ЧВК 2879093. PMID 20505730.
- ^ «Туморскейп». Институт Броуда. Архивировано из оригинал на 2012-04-14. Получено 2012-07-05.
- ^ Лю Ф, Бардхан К., Ян Д., Тхангараджу М., Ганапати В., Лайлс Дж., Ли Дж., Лю К. (июнь 2012 г.). «NF-κB напрямую регулирует транскрипцию Fas, чтобы модулировать Fas-опосредованный апоптоз и подавление опухоли». J Biol Chem. 287 (30): 25530–40. Дои:10.1074 / jbc.M112.356279. ЧВК 3408167. PMID 22669972.
- ^ Ян Д., Торрес К.М., Бардхан К., Циммерман М., МакГаха Т.Л., Лю К. (май 2012 г.). «Децитабин и вориностат взаимодействуют для сенсибилизации клеток карциномы толстой кишки к апоптозу, индуцированному лигандом Fas, in vitro и подавлению опухоли in vivo». J. Immunol. 188 (9): 4441–9. Дои:10.4049 / jimmunol.1103035. ЧВК 3398838. PMID 22461695.
- ^ а б c Гаджате С., Моллинедо Ф. (март 2005 г.). «Цитоскелет-опосредованный рецептор смерти и концентрация лиганда в липидных рафтах образуют кластеры, способствующие апоптозу при химиотерапии рака». J. Biol. Chem. 280 (12): 11641–7. Дои:10.1074 / jbc.M411781200. PMID 15659383.
- ^ а б c Макфарлейн М., Ахмад М., Сринивасула С.М., Фернандес-Алнемри Т., Коэн Г.М., Алнемри Е.С. (октябрь 1997 г.). «Идентификация и молекулярное клонирование двух новых рецепторов цитотоксического лиганда TRAIL». J. Biol. Chem. 272 (41): 25417–20. Дои:10.1074 / jbc.272.41.25417. PMID 9325248.
- ^ а б Шу Х. Б., Халпин Д. Р., Геддел Д. В. (июнь 1997 г.). «Каспер является индуктором апоптоза, связанным с FADD и каспазой». Иммунитет. 6 (6): 751–63. Дои:10.1016 / S1074-7613 (00) 80450-1. PMID 9208847.
- ^ Винченц К., Диксит В.М. (март 1997 г.). «Связанный с Fas белок домена смерти интерлейкин-1-бета-превращающий фермент 2 (FLICE2), гомолог ICE / Ced-3, проксимально участвует в передаче сигналов смерти, опосредованной CD95 и p55». J. Biol. Chem. 272 (10): 6578–83. Дои:10.1074 / jbc.272.10.6578. PMID 9045686.
- ^ Пан Дж., О'Рурк К., Чиннайян А.М., Генц Р., Эбнер Р., Ни Дж., Диксит В.М. (апрель 1997 г.). «Рецептор цитотоксического лиганда TRAIL». Наука. 276 (5309): 111–3. Дои:10.1126 / science.276.5309.111. PMID 9082980. S2CID 19984057.
- ^ Хуанг Б., Эберштадт М., Олейничак Е.Т., Медоуз Р.П., Фесик С.В. (1996). «Структура ЯМР и мутагенез домена смерти Fas (APO-1 / CD95)». Природа. 384 (6610): 638–41. Дои:10.1038 / 384638a0. PMID 8967952. S2CID 2492303.
- ^ Чиннайян А.М., О'Рурк К., Тевари М., Диксит В.М. (май 1995 г.). «FADD, новый белок, содержащий домен смерти, взаимодействует с доменом смерти Fas и инициирует апоптоз». Клетка. 81 (4): 505–12. Дои:10.1016/0092-8674(95)90071-3. PMID 7538907. S2CID 16906755.
- ^ Томас Л.Р., Стиллман ди-джей, Торберн А. (сентябрь 2002 г.). «Регулирование взаимодействий Fas-ассоциированного домена смерти с помощью эффекторного домена смерти, идентифицированного с помощью модифицированного обратного двухгибридного скрининга». J. Biol. Chem. 277 (37): 34343–8. Дои:10.1074 / jbc.M204169200. PMID 12107169.
- ^ Мишо О., Чопп Дж. (Июль 2003 г.). «Индукция апоптоза, опосредованного рецептором TNF I, посредством двух последовательных сигнальных комплексов» (PDF). Клетка. 114 (2): 181–90. Дои:10.1016 / S0092-8674 (03) 00521-X. PMID 12887920. S2CID 17145731.
- ^ Starling GC, Bajorath J, Emswiler J, Ledbetter JA, Aruffo A, Kiener PA (апрель 1997 г.). «Идентификация аминокислотных остатков, важных для связывания лиганда с Fas». J. Exp. Med. 185 (8): 1487–92. Дои:10.1084 / jem.185.8.1487. ЧВК 2196280. PMID 9126929.
- ^ Schneider P, Bodmer JL, Holler N, Mattmann C, Scuderi P, Terskikh A, Peitsch MC, Tschopp J (июль 1997 г.). «Характеристика взаимодействия Fas (Apo-1, CD95) -Fas лиганд». J. Biol. Chem. 272 (30): 18827–33. Дои:10.1074 / jbc.272.30.18827. PMID 9228058.
- ^ Юнг Ю.С., Ким К.С., Ким К.Д., Лим Дж.С., Ким Дж.В., Ким Э. (октябрь 2001 г.). «Ген 2, связанный с апоптозом, связывается с доменом гибели Fas и диссоциирует от Fas во время Fas-опосредованного апоптоза в клетках Jurkat». Biochem. Биофиз. Res. Сообщество. 288 (2): 420–6. Дои:10.1006 / bbrc.2001.5769. PMID 11606059.
- ^ Окура Т., Гонг Л., Камитани Т., Вада Т., Окура И., Вэй С.Ф., Чанг Х.М., Йе Э.Т. (ноябрь 1996 г.). «Защита от гибели клеток, опосредованной Fas / APO-1 и фактором некроза опухоли, с помощью нового белка, сентрина». J. Immunol. 157 (10): 4277–81. PMID 8906799.
- ^ Ryu SW, Chae SK, Kim E (декабрь 2000 г.). «Взаимодействие Daxx, белка, связывающего Fas, с сентрином и Ubc9». Biochem. Биофиз. Res. Сообщество. 279 (1): 6–10. Дои:10.1006 / bbrc.2000.3882. PMID 11112409.
дальнейшее чтение
- Нагата С (1997). «Апоптоз по фактору смерти». Клетка. 88 (3): 355–65. Дои:10.1016 / S0092-8674 (00) 81874-7. PMID 9039262. S2CID 494841.
- Cascino I, Papoff G, Eramo A, Ruberti G (2004). «Растворимые варианты сплайсинга Fas / Apo-1 и апоптоз». Фронт. Biosci. 1 (4): d12–8. Дои:10.2741 / A112. PMID 9159204.
- Uckun FM (1998). «Тирозинкиназа Брутона (BTK) как регулятор двойной функции апоптоза». Biochem. Pharmacol. 56 (6): 683–91. Дои:10.1016 / S0006-2952 (98) 00122-1. PMID 9751072.
- Краммер PH (2000). «Смертельная миссия CD95 в иммунной системе». Природа. 407 (6805): 789–95. Дои:10.1038/35037728. PMID 11048730. S2CID 4328897.
- Сигел Р.М., Чан Ф.К., Чун Х.Дж., Ленардо MJ (2001). «Многогранная роль передачи сигналов Fas в гомеостазе иммунных клеток и аутоиммунитет». Nat. Иммунол. 1 (6): 469–74. Дои:10.1038/82712. PMID 11101867. S2CID 345769.
- Йонехара С (2003). «Рецептор смерти Fas и аутоиммунное заболевание: от первоначального поколения до терапевтического применения агонистических моноклональных антител против Fas». Фактор роста цитокинов Rev. 13 (4–5): 393–402. Дои:10.1016 / S1359-6101 (02) 00024-2. PMID 12220552.
- Чой С., Бенвенист Э. Н. (2004). «Fas-лиганд / Fas-система в головном мозге: регулятор иммунных и апоптотических реакций». Brain Res. Brain Res. Rev. 44 (1): 65–81. Дои:10.1016 / j.brainresrev.2003.08.007. PMID 14739003. S2CID 46587211.
- Poppema S, Maggio E, van den Berg A (2004). «Развитие лимфомы при аутоиммунном лимфопролиферативном синдроме (ALPS) и его связь с мутациями гена Fas». Лейк. Лимфома. 45 (3): 423–31. Дои:10.1080/10428190310001593166. PMID 15160902. S2CID 35128360.
внешняя ссылка
- FAS + рецептор в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Обзор всей структурной информации, доступной в PDB за UniProt: P25445 (Член суперсемейства рецепторов фактора некроза опухолей человека 6) в PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: P25446 (Член суперсемейства рецепторов фактора некроза опухолей мышей 6) на PDBe-KB.
Галерея PDB | |
|---|---|
|
