Экзон - Exon

An экзон является частью ген который закодирует часть финального зрелого РНК произведенный этим геном после интроны были удалены Сплайсинг РНК. Период, термин экзон относится как к последовательности ДНК в гене, так и к соответствующей последовательности в транскриптах РНК. При сплайсинге РНК интроны удаляются, а экзоны ковалентно соединяются друг с другом как часть образования зрелой информационная РНК. Так же, как и весь набор генов разновидность составляет геном, весь набор экзонов составляет экзом.
История
Период, термин экзон происходит от выраженного региона и был придуман американцами биохимик Уолтер Гилберт в 1978 году: "Понятие цистрон … Должен быть заменен единицей транскрипции, содержащей регионы, которые будут потеряны из зрелого мессенджера - которые я предлагаю назвать интронами (для внутригенных регионов) - чередующиеся с регионами, которые будут экспрессироваться - экзонами ».[1]
Это определение изначально было сделано для кодирующих белки транскриптов, которые сплайсируются перед трансляцией. Позднее этот термин стал включать последовательности, удаленные из рРНК[2] и тРНК,[3] и позже он был использован для молекул РНК, происходящих из разных частей генома, которые затем перевязанный транс-сплайсингом.[4]
Вклад в геномы и распределение по размерам
Хотя одноклеточный эукариоты например, у дрожжей интронов либо нет, либо очень мало, многоклеточные животные и особенно позвоночное животное геномы имеют большую долю некодирующая ДНК. Например, в человеческий геном только 1,1% генома занято экзонами, тогда как 24% находятся в интронах, причем 75% генома составляют межгенная ДНК.[5] Это может дать практическое преимущество в омики -помощь здравоохранение (Такие как точная медицина ), потому что это делает коммерциализированные секвенирование всего экзома меньшая и менее дорогая задача, чем коммерческая секвенирование всего генома. Большой разброс в размер генома и C-значение через жизнь форм поставили интересную задачу, названную Загадка значения C.
Во всех эукариотических генах в GenBank было (в 2002 г.) в среднем 5,48 экзонов на ген. В среднем экзон кодируется 30-36 аминокислоты.[6] В то время как самый длинный экзон в геноме человека - 11555 бп long, было обнаружено, что несколько экзонов имеют длину всего 2 п.н.[7] Сообщается об однонуклеотидном экзоне из Арабидопсис геном.[8]
Структура и функции
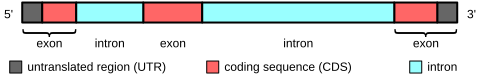
В генах, кодирующих белок, экзоны включают как последовательность, кодирующую белок, так и 5'- и 3'-непереведенные регионы (UTR). Часто первый экзон включает как 5'-UTR, так и первую часть кодирующей последовательности, но экзоны, содержащие только области 5'-UTR или (реже) 3'-UTR, встречаются в некоторых генах, то есть UTR могут содержать интроны. .[9] Немного некодирующая РНК транскрипты также имеют экзоны и интроны.
Зрелые мРНК, происходящие из одного и того же гена, не обязательно должны включать одни и те же экзоны, поскольку разные интроны в пре-мРНК могут быть удалены в процессе альтернативное сращивание.
Экзонизация - это создание нового экзона в результате мутаций в интроны.[10]
Экспериментальные подходы с использованием экзонов
Захват экзонов или же 'захват генов ' это молекулярная биология техника, которая использует существование интрона-экзона сращивание найти новые гены.[11] Первый экзон «захваченного» гена встраивается в экзон, содержащийся в инсерционная ДНК. Этот новый экзон содержит ORF для репортерный ген что теперь можно выразить с помощью усилители которые контролируют целевой ген. Ученый знает, что новый ген был захвачен, когда репортерный ген экспрессируется.
Сплайсинг может быть экспериментально модифицирован таким образом, чтобы экзоны-мишени были исключены из транскриптов зрелой мРНК, путем блокирования доступа управляющих сплайсингом малых ядерных рибонуклеопротеиновых частиц (snRNP) к пре-мРНК с использованием Морфолино-антисмысловые олигонуклеотиды.[12] Это стало стандартной техникой в биология развития. Морфолино-олигонуклеотиды также могут быть нацелены на предотвращение связывания молекул, регулирующих сплайсинг (например, энхансеров сплайсинга, супрессоров сплайсинга) с пре-мРНК, изменяя характер сплайсинга.
Смотрите также
Рекомендации
- ^ Гилберт В. (февраль 1978 г.). «Почему гены по частям?». Природа. 271 (5645): 501. Дои:10.1038 / 271501a0. PMID 622185.
- ^ Кистер К.П., Эккерт WA (март 1987 г.). «Характеристика аутентичного интермедиата в процессе самосплайсинга РНК-предшественника рибосом в макронуклеарах Tetrahymena thermophila». Исследования нуклеиновых кислот. 15 (5): 1905–20. Дои:10.1093 / nar / 15.5.1905. ЧВК 340607. PMID 3645543.
- ^ Валенсуэла П., Венегас А., Вайнберг Ф., Епископ Р., Раттер В. Дж. (Январь 1978 г.). «Структура генов фенилаланин-тРНК дрожжей: промежуточный сегмент ДНК в области, кодирующей тРНК». Труды Национальной академии наук Соединенных Штатов Америки. 75 (1): 190–4. Дои:10.1073 / пнас.75.1.190. ЧВК 411211. PMID 343104.
- ^ Лю А.Ю., Ван дер Плоег Л.Х., Рейсевик Ф.А., Борст П. (июнь 1983 г.). «Единица транспозиции варианта гена 118 поверхностного гликопротеина Trypanosoma brucei. Наличие повторяющихся элементов на его границе и отсутствие последовательностей, связанных с промотором». Журнал молекулярной биологии. 167 (1): 57–75. Дои:10.1016 / S0022-2836 (83) 80034-5. PMID 6306255.
- ^ Venter J.C .; и другие. (2000). «Последовательность генома человека». Наука. 291 (5507): 1304–51. Дои:10.1126 / science.1058040. PMID 11181995.
- ^ Сахаркар М., Пассетти Ф, де Соуза Дж. Э., Лонг М., де Соуза С. Дж. (2002). «ExInt: база данных экзонов интронов». Нуклеиновые кислоты Res. 30 (1): 191–4. Дои:10.1093 / nar / 30.1.191. ЧВК 99089. PMID 11752290.
- ^ Сахаркар М.К .; Chow VT; Кангуане П. (2004). «Распределение экзонов и интронов в геноме человека». В Силико Биол. 4 (4): 387–93. PMID 15217358.
- ^ Го Лэй, Лю Чунь-Мин (2015). "Однонуклеотидный экзон, обнаруженный в Арабидопсис ». Научные отчеты. 5: 18087. Дои:10.1038 / srep18087. ЧВК 4674806. PMID 26657562.
- ^ Бикнелл, AA (декабрь 2012 г.). «Интроны в UTR: почему мы должны перестать их игнорировать». BioEssays. 34 (12): 1025–1034. Дои:10.1002 / bies.201200073. PMID 23108796. S2CID 5808466.
- ^ Сорек Р. (октябрь 2007 г.). «Рождение новых экзонов: механизмы и эволюционные последствия». РНК. 13 (10): 1603–8. Дои:10.1261 / rna.682507. ЧВК 1986822. PMID 17709368.
- ^ Дуйк Г. М; Kim S. W .; Майерс Р. М; Кокс Д. Р. (1990). «Улавливание экзонов: генетический скрининг для идентификации последовательностей-кандидатов в транскрибируемые последовательности в клонированной геномной ДНК млекопитающих». Труды Национальной академии наук. 87 (22): 8995–8999. Дои:10.1073 / pnas.87.22.8995. ЧВК 55087. PMID 2247475.
- ^ Morcos PA (июнь 2007 г.). «Достижение целевого и поддающегося количественной оценке изменения сплайсинга мРНК с морфолино-олигонуклеотидами». Сообщения о биохимических и биофизических исследованиях. 358 (2): 521–7. Дои:10.1016 / j.bbrc.2007.04.172. PMID 17493584.
Общие ссылки
- Zhang MQ (май 1998 г.). «Статистические особенности экзонов человека и их фланкирующих областей». Молекулярная генетика человека. 7 (5): 919–32. Дои:10,1093 / чмг / 7,5,919. PMID 9536098.
- Танарадж Т.А., Робинсон А.Дж. (ноябрь 2000 г.). «Предсказание точных границ экзонов». Краткий. Биоинформ. 1 (4): 343–56. Дои:10.1093 / bib / 1.4.343. PMID 11465052.