Натриевый канал - Sodium channel
Натриевые каналы находятся интегральные мембранные белки эта форма ионные каналы, проводящие ионы натрия (Na+ ) через клетки плазматическая мембрана.[1][2] Они принадлежат к суперсемейство катионных каналов и может быть классифицирован в соответствии с триггером, который открывает канал для таких ионов, то есть либо по изменению напряжения («регулируемый по напряжению», «чувствительный к напряжению» или «зависимый от напряжения» натриевый канал; также называемый «VGSC») или «Навигационный канал») или связывание вещества ( лиганд ) в канал (лиганд-зависимые натриевые каналы).
В возбудимых клетках, таких как нейроны, миоциты, и некоторые типы глия, натриевые каналы отвечают за фазу нарастания потенциалы действия. Эти каналы проходят через три различных состояния: состояние покоя, активное и неактивное состояние. Даже несмотря на то, что состояние покоя и неактивное состояние не позволяют ионам проходить через каналы, существует разница в их структурной конформации.
Селективность
Натриевые каналы очень селективны для транспорта ионов через клеточные мембраны. Высокая селективность по иону натрия достигается многими различными способами. Все они включают инкапсуляцию иона натрия в полости определенного размера внутри более крупной молекулы.[3]
Напряжение-управляемые натриевые каналы
Структура

Натриевые каналы состоят из крупных α-субъединиц, которые связаны с белками, например β-субъединиц. Субъединица α образует ядро канала и функционирует сама по себе. Когда белок α-субъединицы экспрессируется клеткой, он может образовывать каналы, которые проводят Na+ потенциалзависимым способом, даже если β-субъединицы или другие известные модулирующие белки не экспрессируются. Когда дополнительные белки собираются с α-субъединицами, полученный комплекс может демонстрировать измененную зависимость от напряжения и клеточную локализацию.
Α-субъединица имеет четыре повторяющихся домена, обозначенных от I до IV, каждый из которых содержит шесть трансмембранных сегментов, обозначенных с S1 по S6. Очень консервированный Сегмент S4 действует как датчик напряжения канала. Чувствительность к напряжению этого канала обусловлена положительными аминокислотами, расположенными в каждой третьей позиции.[5] Когда стимулируется изменением трансмембранное напряжение этот сегмент перемещается к внеклеточной стороне клеточной мембраны, позволяя каналу становиться проницаемым для ионов. Ионы проходят через поры, которые можно разделить на две части. Более внешняя (то есть более внеклеточная) часть поры образована «Р-петлями» (область между S5 и S6) четырех доменов. Эта область является наиболее узкой частью поры и отвечает за ее ионную селективность. Внутренняя часть (т.е. более цитоплазматическая) поры образована комбинированными сегментами S5 и S6 четырех доменов. Область, связывающая домены III и IV, также важна для функции канала. Эта область закрывает канал после продолжительной активации, деактивируя его.
Ворота
Напряжение-gated Na+ каналы имеют три основных конформационных состояния: закрытый, открытый и неактивный. Прямые / обратные переходы между этими состояниями соответственно называются активацией / деактивацией (между открытым и закрытым, соответственно), инактивацией / реактивацией (между инактивированным и открытым, соответственно) и восстановлением после инактивации / инактивации в закрытом состоянии (между инактивированным и закрытым. , соответственно). Закрытое и инактивированное состояния не проницаемы для ионов.
До возникновения потенциала действия аксональная мембрана находится в нормальном состоянии. потенциал покоя, около -70 мВ в большинстве нейронов человека и Na+ каналы находятся в деактивированном состоянии, заблокированы на внеклеточной стороне своими ворота активации. В ответ на повышение мембранного потенциала примерно до -55 мВ (в данном случае, вызванного потенциалом действия), ворота активации открываются, позволяя положительно заряженному Na+ ионы поступают в нейрон через каналы, вызывая повышение напряжения на мембране нейрона до +30 мВ в нейронах человека. Поскольку напряжение на мембране изначально отрицательное, по мере увеличения ее напряжения к и прошлый ноль (от -70 мВ в состоянии покоя до максимального значения +30 мВ), говорят, что он деполяризуется. Это увеличение напряжения составляет фазу нарастания потенциала действия.
| Потенциал действия | Мембранный потенциал | Целевой потенциал | Целевое состояние ворот | Целевое состояние нейрона |
|---|---|---|---|---|
| Отдыхает | −70 мВ | −55 мВ | Деактивировано → Активировано | Поляризованный |
| Рост | −55 мВ | 0 мВ | Активирован | Поляризованные → Деполяризованные |
| Рост | 0 мВ | +30 мВ | Активировано → Деактивировано | Деполяризованный |
| Падение | +30 мВ | 0 мВ | Инактивировано | Деполяризованный → Реполяризованный |
| Падение | 0 мВ | −70 мВ | Инактивировано | Реполяризованный |
| Недокус | −70 мВ | −75 мВ | Деактивировано → Деактивировано | Реполяризованный → Гиперполяризованный |
| Отскок | −75 мВ | −70 мВ | Деактивировано | Гиперполяризованный → Поляризованный |
На пике потенциала действия, когда достаточно Na+ попал в нейрон, и потенциал мембраны стал достаточно высоким, Na+ каналы деактивируют себя, закрывая свои ворота инактивации. Ворота инактивации можно рассматривать как «пробку», привязанную к доменам III и IV внутриклеточной альфа-субъединицы канала. Закрытие ворот инактивации вызывает Na+ поток через канал останавливается, что, в свою очередь, приводит к прекращению роста мембранного потенциала. Закрытие ворот инактивации создает рефрактерный период в каждом отдельном Na+ канал. Этот рефрактерный период исключает возможность движения потенциала действия в противоположном направлении обратно к соме. Когда его ворота инактивации закрыты, канал считается неактивным. С Na+ канал больше не вносит вклад в мембранный потенциал, потенциал уменьшается обратно до своего потенциала покоя, поскольку нейрон реполяризуется и впоследствии гиперполяризуется, и это составляет фазу падения потенциала действия. Поэтому рефрактерный период каждого канала жизненно важен для однонаправленного распространения потенциала действия вниз по аксону для правильной связи между нейронами.
Когда напряжение мембраны становится достаточно низким, ворота инактивации снова открываются, а ворота активации закрываются в процессе, называемом деинактивация. При закрытых воротах активации и открытых воротах инактивации Na+ канал снова находится в деактивированном состоянии и готов участвовать в другом потенциале действия.
Когда какой-либо ионный канал не деактивируется, он считается постоянно (или тонически) активным. Некоторые виды ионных каналов по своей природе постоянно активны. Однако генетические мутации, вызывающие постоянную активность в других каналах, могут вызывать заболевание, создавая чрезмерную активность определенных типов нейронов. Мутации, мешающие Na+ инактивация каналов может способствовать сердечно-сосудистым заболеваниям или эпилептическим припадкам за счет оконные токи, что может вызвать чрезмерное возбуждение мышечных и / или нервных клеток.
Моделирование поведения ворот
Временное поведение Na+ каналы могут быть смоделированы Марковский схема или Ходжкина – Хаксли формализм. В первой схеме каждый канал занимает отдельный государственный с дифференциальные уравнения описание переходов между состояниями; во втором - каналы рассматриваются как численность населения на которые влияют три независимых стробирующих переменных. Каждая из этих переменных может принимать значение от 1 (полностью проницаемая для ионов) до 0 (полностью непроницаемая), произведение этих переменных дает процент проводящих каналов. Можно показать, что модель Ходжкина – Хаксли эквивалентна марковской модели.
Непроницаемость для других ионов
Пора натриевых каналов содержит фильтр селективности из отрицательно заряженного аминокислота остатки, которые притягивают положительный Na+ ion и не допускать попадания отрицательно заряженных ионов, таких как хлористый. Катионы попадают в более узкую часть поры размером 0,3 на 0,5. нм широкий, который достаточно велик, чтобы позволить одному Na+ ион с водой молекула связаны, чтобы пройти. Чем больше K+ ion не может пройти через эту область. Ионы разного размера также не могут взаимодействовать с отрицательно заряженными глютаминовая кислота остатки, выстилающие поры.[нужна цитата ]
Разнообразие
Управляемые напряжением натриевые каналы обычно состоят из альфа-субъединицы, которая формирует поры для ионной проводимости, и одной-двух бета-субъединиц, которые выполняют несколько функций, включая модуляцию блокировки канала.[6] Экспрессии одной альфа-субъединицы достаточно для создания функционального канала.
Альфа-субъединицы
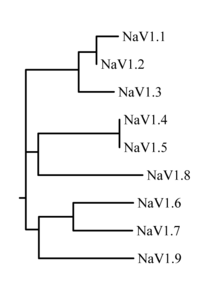
Семейство натриевых каналов состоит из девяти известных членов с аминокислотной идентичностью> 50% в трансмембранных сегментах и областях внеклеточной петли. В настоящее время используется стандартизованная номенклатура натриевых каналов, которую поддерживает ИУФАР.[7][8]
Белки этих каналов получили название Nav1.1 через Nav1.9. Названия генов обозначаются от SCN1A до SCN11A (ген SCN6 / 7A является частью NaИкс подсемейства и имеет неопределенную функцию). Вероятная эволюционная взаимосвязь между этими каналами, основанная на сходстве их аминокислотных последовательностей, показана на рисунке 1. Индивидуальные натриевые каналы отличаются не только различиями в их последовательности, но также их кинетикой и профилями экспрессии. Некоторые из этих данных обобщены в таблице 1 ниже.
| Название протеина | Ген | Профиль выражения | Связанный человек каннелопатии |
|---|---|---|---|
| Nav1.1 | SCN1A | Центральные нейроны, [периферические нейроны] и сердечные миоциты | фебрильный эпилепсия, ГЭФС +, Синдром Драве (также известен как тяжелая миклоническая эпилепсия младенчества или SMEI), пограничный SMEI (SMEB), синдром Веста (также известный как инфантильные спазмы), Синдром Дуза (также известный как миоклоническая астатическая эпилепсия), трудноизлечимая детская эпилепсия с генерализованными тонико-клоническими припадками (ICEGTC), синдром Панайотопулоса, семейная гемиплегическая мигрень (FHM), семейный аутизм, энцефалит Расмуссенса и синдром Леннокса-Гасто.[9] |
| Nav1.2 | SCN2A | Центральные нейроны, периферические нейроны | унаследованный фебрильные судороги, эпилепсия, и расстройство аутистического спектра |
| Nav1.3 | SCN3A | Центральные нейроны, периферические нейроны и сердечные миоциты | эпилепсия, боль, пороки развития головного мозга[10][11] |
| Nav1.4 | SCN4A | Скелетные мышцы | гиперкалиемический периодический паралич, врожденная парамиотония, и миотония с обострением калия |
| Nav1.5 | SCN5A | Сердечные миоциты, неиннервируемые скелетные мышцы, центральные нейроны, гладкомышечные клетки желудочно-кишечного тракта и интерстициальные клетки Кахаля | Сердечный: Синдром удлиненного интервала QT Тип 3, Синдром Бругада, прогрессирующее заболевание сердечной проводимости, семейный мерцательная аритмия и идиопатический мерцание желудочков;[12] Желудочно-кишечный тракт: Синдром раздраженного кишечника;[13] |
| Nav1.6 | SCN8A | Центральные нейроны, ганглии задних корешков, периферические нейроны, сердце, клетки глии | Эпилепсия,[14] атаксия, дистония, тремор [15] |
| Nav1.7 | SCN9A | Ганглии задних корней, симпатические нейроны, Шванновские клетки, и нейроэндокринные клетки | эритромелалгия, PEPD, нечувствительность к боли, связанная с каннелопатией[10] и недавно обнаружил инвалидизирующую форму фибромиалгия (полиморфизм rs6754031)[16] |
| Nav1.8 | SCN10A | Ганглии задних корней | боль,[10] нервно-психические расстройства |
| Nav1.9 | SCN11A | Ганглии задних корней | боль[10] |
| NaИкс | SCN7A | сердце, матка, скелетные мышцы, астроциты, ганглиозные клетки дорзального корня | никто не известен |
Бета-субъединицы
Бета-субъединицы натриевого канала представляют собой трансмембранные гликопротеины типа 1 с внеклеточным N-концом и цитоплазматическим С-концом. Как члены суперсемейства Ig, бета-субъединицы содержат прототипическую петлю Ig V-набора во внеклеточном домене. Они не имеют какой-либо гомологии со своими аналогами кальциевых и калиевых каналов.[17] Вместо этого они гомологичны молекулам адгезии нервных клеток (CAM) и большому семейству L1 CAM. Существует четыре различных бета-версии, названных в порядке обнаружения: SCN1B, SCN2B, SCN3B, SCN4B (таблица 2). Бета 1 и бета 3 взаимодействуют с альфа-субъединицей нековалентно, тогда как бета 2 и бета 4 связываются с альфа через дисульфидную связь.[18] Натриевые каналы с большей вероятностью будут оставаться открытыми при подпороговом мембранном потенциале при взаимодействии с бета-токсинами, что, в свою очередь, вызывает немедленное ощущение боли.[19]
Роль бета-субъединиц как молекул клеточной адгезии
Помимо регуляции стробирования каналов, бета-субъединицы натриевых каналов также модулируют экспрессию каналов и образуют связи с внутриклеточный цитоскелет через анкирин и спектрин.[6][20][21]Управляемые напряжением натриевые каналы также собираются с множеством других белков, таких как белки FHF (гомологичный фактор фактора роста фибробластов), кальмодулин, цитоскелет или регуляторные киназы,[22][6][23][24][25] которые образуют комплекс с натриевыми каналами, влияя на его экспрессию и / или функцию. Несколько бета-субъединиц взаимодействуют с одним или несколькими внеклеточный матрикс (ЕСМ) молекулы. Контактин, также известный как F3 или F11, связывается с бета-1, как показано посредством коиммунопреципитации.[26] Фибронектин -подобные (FN-подобные) повторы Tenascin -C и Tenascin -R связывать с бета 2 в отличие от Фактор эпидермального роста -подобные (EGF-подобные) повторы, которые отталкивают beta2.[27] Дезинтегрин и металлопротеиназа (ADAM) 10 сбрасывают бета-2 эктодомен возможно индуцирование роста нейритов.[28] Бета 3 и бета 1 связываются с нейрофасцином в Узлах Ранвье в развивающихся нейронах.[29]
| Название протеина | Генная ссылка | Собирается с | Профиль выражения | Связанный человек каннелопатии |
|---|---|---|---|---|
| Navβ1 | SCN1B | Nav1,1 к Nav1.7 | Центральные нейроны, периферические нейроны, скелетные мышцы, сердце, глия | эпилепсия (GEFS +), Синдром Бругада[30] |
| Navβ2 | SCN2B | Nav1.1, Nav1.2, Nav1,5 к Nav1.7 | Центральные нейроны, периферические нейроны, сердце, глия | Синдром Бругада[30] |
| Navβ3 | SCN3B | Nav1,1 к Nav1.3, Nav1.5 | центральные нейроны, надпочечники, почки, периферические нейроны | Синдром Бругада[30] |
| Navβ4 | SCN4B | Nav1.1, Nav1.2, Nav1.5 | сердце, скелетные мышцы, центральные и периферические нейроны | никто не известен |
Натриевые каналы, управляемые лигандами
Лиганд-закрытый натриевые каналы активируются путем связывания лиганд вместо изменения мембранного потенциала.
Они находятся, например, в нервномышечное соединение в качестве никотиновые рецепторы, где лиганды ацетилхолин молекулы. Большинство каналов этого типа в некоторой степени проницаемы как для калия, так и для натрия.
Роль в потенциале действия
Напряжение управляемых натриевых каналов играет важную роль в потенциалы действия. Если достаточно каналов открывается при изменении в ячейке мембранный потенциал, небольшое, но значительное количество Na+ ионы будут двигаться в клетку вниз по их электрохимический градиент, дальше деполяризующий клетка. Таким образом, чем больше Na+ каналов, локализованных в области клеточной мембраны, тем быстрее будет распространяться потенциал действия и тем больше возбудимый эта область ячейки будет. Это пример петля положительной обратной связи. Способность этих каналов переходить в закрытое-неактивное состояние вызывает период отражения и имеет решающее значение для распространения потенциалов действия вниз по аксон.
Na+ каналы открываются и закрываются быстрее, чем K+ каналы, производя приток положительного заряда (Na+) к началу потенциал действия и истечение (K+) ближе к концу.
С другой стороны, управляемые лигандами натриевые каналы создают изменение мембранного потенциала в первую очередь в ответ на связывание с ним лиганда.
Фармакологическая модуляция
Блокираторы
Активаторы
Следующие естественные вещества постоянно активируют (открывают) натриевые каналы:
- Алкалоид -основанные токсины
- аконитин
- батрахотоксин
- бреветоксин
- сигуатоксин
- дельфинин
- немного серыйанотоксины, например, грейанотоксин I (другие гранотоксины неактивны или закрывают натриевые каналы)
- вератридин
Модификаторы стробирования
Следующие токсины изменяют закрытие натриевых каналов:
- Пептид -основанные токсины
модуляция pH
Изменения рН крови и тканей сопровождают физиологические и патофизиологические состояния, такие как физические упражнения, ишемия сердца, ишемический инсульт и прием кокаина. Известно, что эти состояния вызывают симптомы электрических заболеваний у пациентов с мутациями натриевых каналов. Протоны вызывают различные изменения в стробировании натриевых каналов, которые обычно приводят к уменьшению амплитуды переходного натриевого тока и увеличению доли не инактивирующих каналов, по которым проходят постоянные токи. Эти эффекты характерны для болезнетворных мутантов в нейрональных, скелетных мышцах и сердечной ткани и могут быть объединены с мутантами, которые придают большую протонную чувствительность к натриевым каналам, что предполагает роль протонов в запуске острых симптомов электрического заболевания.[33]
Молекулярные механизмы протонного блока
Одноканальные данные кардиомиоцитов показали, что протоны могут снижать проводимость отдельных натриевых каналов.[34] Фильтр селективности натриевого канала состоит из одного остатка в каждой из четырех петель пор четырех функциональных доменов. Эти четыре остатка известны как мотив DEKA.[35] Скорость проникновения натрия через натриевые каналы определяется четырьмя карбоксилатными остатками, мотивом EEDD, которые составляют внешнее заряженное кольцо.[35] Протонирование этих карбоксилатов является одним из основных факторов блокирования протонов в натриевых каналах, хотя есть и другие остатки, которые также влияют на чувствительность к pH.[36] Одним из таких остатков является C373 в сердечном натриевом канале, что делает его наиболее чувствительным к pH натриевым каналом среди натриевых каналов, которые были изучены на сегодняшний день.[37]
Модуляция pH стробирования натриевых каналов
Так как натриевой канал сердца является наиболее чувствительным к pH натриевым каналом, большая часть того, что известно, основано на этом канале. Было показано, что снижение внеклеточного pH деполяризует зависимость активации и инактивации от напряжения до более положительных потенциалов. Это указывает на то, что во время действий, снижающих pH крови, таких как упражнения, вероятность активации и деактивации каналов выше, чем положительные мембранные потенциалы, что может привести к потенциальным побочным эффектам.[38] Натриевые каналы, экспрессируемые в волокнах скелетных мышц, превратились в каналы, относительно нечувствительные к pH. Было высказано предположение, что это защитный механизм против потенциальной чрезмерной или недостаточной возбудимости в скелетных мышцах, поскольку уровни pH крови очень чувствительны к изменению во время движения.[39][40] Недавно было показано, что мутация смешанного синдрома, которая вызывает периодический паралич и миотонию в скелетном натриевом канале, придает этому каналу чувствительность к pH, что делает закрытие этого канала подобным таковому у сердечного подтипа.[41]
Модуляция pH среди изученных на данный момент подтипов
Эффекты протонирования описаны в Nav1.1-Nav1.5. Среди этих каналов Nav1.1-Nav1.3 и Nav1.5 демонстрируют деполяризованную зависимость активации от напряжения, тогда как активация в Nav1.4 остается нечувствительной к ацидозу. Зависимость от напряжения установившейся быстрой инактивации в Nav1.1-Nav1.4 не изменилась, но установившаяся быстрая инактивация в Nav1.5 деполяризована. Следовательно, среди натриевых каналов, которые были изучены до сих пор, Nav1.4 является наименьшим, а Nav1.5 - наиболее чувствительными к протонам подтипами.[42]
Эволюция
Напряжениезависимый натриевый канал присутствует в членах хоанофлагелляты, считается ближайшим из ныне живущих одноклеточных родственников животных.[43][44] Это говорит о том, что предковая форма животного канала была среди многих белков, которые играют центральную роль в жизни животных, но которые, как полагают, эволюционировали до многоклеточности.[45] Четырехдоменный потенциал-управляемый натриевый канал животных, вероятно, произошел от ионного канала с одной субъединицей, который, вероятно, был проницаемым для ионов калия, посредством последовательности двух событий дупликации.[46] Эта модель опирается на тот факт, что субъединицы I и III (а также II и IV) группируются по сходству, предполагая, что двухканальный промежуточный продукт, образованный в результате первой дупликации, существовал достаточно долго, чтобы произошло расхождение между двумя его субъединицами. После второго дублирования на канале осталось два набора похожих доменов.[46] Полагают, что полученный четырехдоменный канал был проницаем в первую очередь для кальция и несколько раз достигал селективности по натрию независимо.[47][48] После расхождения с беспозвоночными линия позвоночных претерпела два полногеномные дупликации (WGDs), давая набор из четырех прологов гена натриевых каналов у предковых позвоночных, все из которых были сохранены.[49][50] После разделения четвероногих и костистых костистые кости, вероятно, подверглись третьей WGD, что привело к появлению восьми прологов натриевого канала, выраженных у многих современных рыб.[49] Считается, что современный набор генов натрия из десяти паралогов у млекопитающих возник в результате серии параллельных и вложенных дупликаций с участием двух из четырех паралогов, присутствующих в предках всех четвероногих.[50]
Смотрите также
Рекомендации
- ^ Джесселл TM, Кандел ER, Шварц JH (2000). Принципы нейронологии (4-е изд.). Нью-Йорк: Макгроу-Хилл. стр.154–69. ISBN 978-0-8385-7701-1.CS1 maint: несколько имен: список авторов (связь)
- ^ Бертил Гиллель (2001). Ионные каналы возбудимых мембран. (3-е изд.). Сандерленд, Массачусетс: Синауэр. С. 73–7. ISBN 978-0-87893-321-1.
- ^ Лим С, Дудев Т (2016). «Глава 10. Селективность калия и натрия в фильтрах селективности моновалентных ионных каналов». В Astrid S, Helmut S, Roland KO S (ред.). Ионы щелочных металлов: их роль в жизни. Ионы металлов в науках о жизни. 16. Springer. С. 325–347. Дои:10.1007/978-4-319-21756-7_9 (неактивно 10.09.2020).CS1 maint: DOI неактивен по состоянию на сентябрь 2020 г. (связь)
- ^ Ю. Ф. Х., Каттералл В. А. (2003). «Обзор семейства натриевых каналов с регулируемым напряжением». Геномная биология. 4 (3): 207. Дои:10.1186 / gb-2003-4-3-207. ЧВК 153452. PMID 12620097.
- ^ Николлс, Мартин, Фукс, Браун, Даймонд, Вайсблат. (2012) «От нейрона к мозгу», 5-е изд. стр. 86
- ^ а б c Isom LL (февраль 2001 г.). «Бета-субъединицы натриевого канала: все, кроме вспомогательных». Нейробиолог. 7 (1): 42–54. Дои:10.1177/107385840100700108. PMID 11486343. S2CID 86422657.
- ^ IUPHAR - Международный союз фундаментальной и клинической фармакологии
- ^ Каттералл, В. А., Голдин А. Л., Ваксман С. Г. (декабрь 2005 г.). "Международный союз фармакологии. XLVII. Номенклатура и взаимосвязь между структурой и функцией потенциалзависимых натриевых каналов". Фармакологические обзоры. 57 (4): 397–409. Дои:10.1124 / пр.57.4.4. PMID 16382098. S2CID 7332624.
- ^ Лоссин К. «Информационная база SCN1A». В архиве из оригинала 2011-07-21. Получено 2009-10-30.
компиляция генетических вариаций гена SCN1A, которые изменяют экспрессию или функцию Nav1.1
- ^ а б c d Беннетт Д.Л., Кларк А.Дж., Хуанг Дж., Ваксман С.Г., Диб-Хадж С.Д. (апрель 2019 г.). «Роль управляемых напряжением натриевых каналов в передаче болевых сигналов». Физиологические обзоры. 99 (2): 1079–1151. Дои:10.1152 / Physrev.00052.2017. PMID 30672368.
- ^ Смит Р.С., Кенни С.Дж., Ганеш В., Янг А., Борхес-Монрой Р., Партлоу Дж. Н. и др. (Сентябрь 2018 г.). «V1.3) Регулирование складок коры головного мозга человека и развития оральной моторики». Нейрон. 99 (5): 905–913.e7. Дои:10.1016 / j.neuron.2018.07.052. ЧВК 6226006. PMID 30146301.
- ^ Чоккалингам П., Уайльд А. (сентябрь 2012 г.). «Многогранный натриевой канал сердца и его клинические последствия». Сердце. 98 (17): 1318–24. Дои:10.1136 / heartjnl-2012-301784. PMID 22875823. S2CID 44433455.
- ^ Beyder A, Mazzone A, Strege PR, Tester DJ, Saito YA, Bernard CE, Enders FT, Ek WE, Schmidt PT, Dlugosz A, Lindberg G, Karling P, Ohlsson B, Gazouli M, Nardone G, Cuomo R, Usai- Satta P, Galeazzi F, Neri M, Portincasa P, Bellini M, Barbara G, Camilleri M, Locke GR, Talley NJ, D'Amato M, Ackerman MJ, Farrugia G (июнь 2014 г.). «Потеря функции потенциалзависимого натриевого канала NaV1.5 (каннелопатии) у пациентов с синдромом раздраженного кишечника». Гастроэнтерология. 146 (7): 1659–1668. Дои:10.1053 / j.gastro.2014.02.054. ЧВК 4096335. PMID 24613995.
- ^ Батлер К.М., да Силва С., Шафир Ю., Вайсфельд-Адамс Д.Д., Александр Дж.Дж., Хегде М., Эскайг А. (январь 2017 г.). «De novo и унаследованные мутации эпилепсии SCN8A, обнаруженные с помощью анализа генной панели». Исследования эпилепсии. 129: 17–25. Дои:10.1016 / j.eplepsyres.2016.11.002. ЧВК 5321682. PMID 27875746.
- ^ Мейслер MH, Кирни JA (август 2005 г.). «Мутации натриевых каналов при эпилепсии и других неврологических расстройствах». Журнал клинических исследований. 115 (8): 2010–7. Дои:10.1172 / JCI25466. ЧВК 1180547. PMID 16075041.
- ^ Варгас-Аларкон Г., Альварес-Леон Э., Фрагосо Дж. М., Варгас А., Мартинес А., Вальехо М., Мартинес-Лавин М. (февраль 2012 г.). «Кодируемый геном SCN9A полиморфизм натриевых каналов ганглиев дорсальных корешков, связанный с тяжелой фибромиалгией». BMC скелетно-мышечные заболевания. 13: 23. Дои:10.1186/1471-2474-13-23. ЧВК 3310736. PMID 22348792.
- ^ Catterall WA (апрель 2000 г.). «От ионных токов к молекулярным механизмам: структура и функция потенциалзависимых натриевых каналов». Нейрон. 26 (1): 13–25. Дои:10.1016 / S0896-6273 (00) 81133-2. PMID 10798388. S2CID 17928749.
- ^ Isom LL, De Jongh KS, Patton DE, Reber BF, Offord J, Charbonneau H, Walsh K, Goldin AL, Catterall WA (май 1992 г.). «Первичная структура и функциональная экспрессия бета-1 субъединицы натриевого канала мозга крысы». Наука. 256 (5058): 839–42. Bibcode:1992Наука ... 256..839I. Дои:10.1126 / наука.1375395. PMID 1375395.
- ^ blackboard.jhu.edu (PDF) https://blackboard.jhu.edu/courses/1/AS.410.603.83.SU20/db/_10095565_1/Isolation%20and%20Characterization%20of%20CvIV4%20A%20Pain%20Inducing%20a-scorpion%20Toxin.pdf. Получено 2020-07-16. Отсутствует или пусто
| название =(помощь) - ^ Малхотра Дж. Д., Казен-Гиллеспи К., Хорч М., Исом Л. Л. (апрель 2000 г.). «Бета-субъединицы натриевого канала опосредуют адгезию гомофильных клеток и привлекают анкирин к точкам межклеточного контакта». Журнал биологической химии. 275 (15): 11383–8. Дои:10.1074 / jbc.275.15.11383. PMID 10753953.
- ^ Малхотра Дж. Д., Купманн М. С., Казен-Гиллеспи К. А., Феттман Н., Хорч М., Исом Л. Л. (июль 2002 г.). «Структурные требования для взаимодействия субъединиц бета 1 натриевого канала с анкирином». Журнал биологической химии. 277 (29): 26681–8. Дои:10.1074 / jbc.M202354200. PMID 11997395.
- ^ Cantrell AR, Catterall WA (июнь 2001 г.). «Нейромодуляция Na + -каналов: неожиданная форма клеточной пластичности». Обзоры природы. Неврология. 2 (6): 397–407. Дои:10.1038/35077553. PMID 11389473. S2CID 22885909.
- ^ Шах Б.С., Раш А.М., Лю С., Тиррелл Л., Блэк Дж. А., Диб-Хадж С. Д., Ваксман С. Г. (август 2004 г.). «Контактин связывается с натриевым каналом Nav1.3 в естественных тканях и увеличивает плотность каналов на поверхности клетки». Журнал неврологии. 24 (33): 7387–99. Дои:10.1523 / JNEUROSCI.0322-04.2004. ЧВК 6729770. PMID 15317864.
- ^ Виттмак Е.К., Раш А.М., Кранер М.Дж., Гольдфарб М., Ваксман С.Г., Диб-Хадж С.Д. (июль 2004 г.). «Фактор 2B, гомологичный фактору роста фибробластов: ассоциация с Nav1.6 и избирательная совместная локализация в узлах Ранвье аксонов дорсального корешка». Журнал неврологии. 24 (30): 6765–75. Дои:10.1523 / JNEUROSCI.1628-04.2004. ЧВК 6729706. PMID 15282281.
- ^ Раш А.М., Виттмак Е.К., Тиррелл Л., Блэк Дж. А., Диб-Хадж С. Д., Ваксман С. Г. (май 2006 г.). «Дифференциальная модуляция натриевого канала Na (v) 1,6 двумя членами подсемейства фактора 2 гомологичного фактора роста фибробластов». Европейский журнал нейробиологии. 23 (10): 2551–62. Дои:10.1111 / j.1460-9568.2006.04789.x. PMID 16817858.
- ^ Казаринова-Нойес К., Малхотра Д.Д., МакИвен Д.П., Маттей Л.Н., Берглунд Э.О., Ранчт Б., Левинсон С.Р., Шахнер М., Шрагер П., Изом Л.Л., Сяо З.С. (октябрь 2001 г.). «Контактин связывается с Na + каналами и увеличивает их функциональную экспрессию». Журнал неврологии. 21 (19): 7517–25. Дои:10.1523 / JNEUROSCI.21-19-07517.2001. ЧВК 6762905. PMID 11567041.
- ^ Сринивасан Дж., Шахнер М., Каттералл В.А. (декабрь 1998 г.). «Взаимодействие потенциалзависимых натриевых каналов с молекулами внеклеточного матрикса тенасцин-C и тенасцин-R». Труды Национальной академии наук Соединенных Штатов Америки. 95 (26): 15753–7. Bibcode:1998PNAS ... 9515753S. Дои:10.1073 / пнас.95.26.15753. ЧВК 28116. PMID 9861042.
- ^ Ким Д.Й., Ингано Л.А., Кэри Б.В., Петтингелл У.Х., Ковач Д.М. (июнь 2005 г.). «Опосредованное пресенилином / гамма-секретазой расщепление бета2-субъединицы потенциал-зависимого натриевого канала регулирует клеточную адгезию и миграцию». Журнал биологической химии. 280 (24): 23251–61. Дои:10.1074 / jbc.M412938200. PMID 15833746.
- ^ Ratcliffe CF, Westenbroek RE, Curtis R, Catterall WA (июль 2001 г.). «Субъединицы бета1 и бета3 натриевого канала связаны с нейрофасцином через их внеклеточный иммуноглобулиноподобный домен». Журнал клеточной биологии. 154 (2): 427–34. Дои:10.1083 / jcb.200102086. ЧВК 2150779. PMID 11470829.
- ^ а б c Анцелевич С., Патоцкай Б. (январь 2016 г.). «Синдром Бругада: клинические, генетические, молекулярные, клеточные и ионные аспекты». Актуальные проблемы кардиологии. 41 (1): 7–57. Дои:10.1016 / j.cpcardiol.2015.06.002. ЧВК 4737702. PMID 26671757.
- ^ Grolleau F, Stankiewicz M, Birinyi-Strachan L, Wang XH, Nicholson GM, Pelhate M, Lapied B (февраль 2001 г.). «Электрофизиологический анализ нейротоксического действия токсина воронкообразного паука, дельта-атракотоксина-HV1a, на потенциалзависимые Na + каналы насекомых». Журнал экспериментальной биологии. 204 (Пт 4): 711–21. PMID 11171353.
- ^ Поссани Л.Д., Бесеррил Б., Делепьер М., Титгат Дж. (Сентябрь 1999 г.). «Токсины скорпиона, специфичные для Na + -каналов». Европейский журнал биохимии. 264 (2): 287–300. Дои:10.1046 / j.1432-1327.1999.00625.x. PMID 10491073.
- ^ Петерс CH, Ghovanloo MR, Gershome C, Ruben PC (февраль 2018 г.). "pH-модуляция натриевых каналов, управляемых напряжением". Напряжение-управляемые натриевые каналы: структура, функции и каннелопатии. Справочник по экспериментальной фармакологии. 246. С. 147–160. Дои:10.1007/164_2018_99. ISBN 978-3-319-90283-8. PMID 29460150.
- ^ Чжан Дж. Ф., Зигельбаум С. А. (декабрь 1991 г.). «Влияние внешних протонов на одиночные сердечные натриевые каналы миоцитов желудочков морской свинки». Журнал общей физиологии. 98 (6): 1065–83. Дои:10.1085 / jgp.98.6.1065. ЧВК 2229074. PMID 1664454.
- ^ а б Sun YM, Favre I, Schild L, Moczydlowski E (декабрь 1997 г.). «На структурной основе избирательного проникновения органических катионов через потенциал-управляемый натриевый канал. Влияние мутаций аланина в локусе DEKA на селективность, ингибирование Са2 + и Н + и молекулярное просеивание». Журнал общей физиологии. 110 (6): 693–715. Дои:10.1085 / jgp.110.6.693. ЧВК 2229404. PMID 9382897.
- ^ Хан А., Романцева Л., Лам А., Липкинд Г., Фоззард Х.А. (август 2002 г.). «Роль карбоксилатов внешнего кольца поры натриевого канала скелетных мышц крысы в протонном блоке». Журнал физиологии. 543 (Pt 1): 71–84. Дои:10.1113 / jphysiol.2002.021014. ЧВК 2290475. PMID 12181282.
- ^ Вилин Ю.Ю., Петерс СН, Рубен П.К. (2012). «Ацидоз по-разному модулирует инактивацию в каналах na (v) 1,2, na (v) 1,4 и na (v) 1,5». Границы фармакологии. 3: 109. Дои:10.3389 / fphar.2012.00109. ЧВК 3372088. PMID 22701426.
- ^ Джонс Д.К., Петерс СН, Аллард С.Р., Клейдон Т.В., Рубен П.К. (февраль 2013 г.). «Протонные датчики в поровой области сердечного потенциалзависимого натриевого канала». Журнал биологической химии. 288 (7): 4782–91. Дои:10.1074 / jbc.M112.434266. ЧВК 3576083. PMID 23283979.
- ^ Хан А., Кайл Дж. У., Ханк Д. А., Липкинд Г. М., Фоззард Н. А. (октябрь 2006 г.). «Изоформ-зависимое взаимодействие потенциалзависимых натриевых каналов с протонами». Журнал физиологии. 576 (Pt 2): 493–501. Дои:10.1113 / jphysiol.2006.115659. ЧВК 1890365. PMID 16873405.
- ^ Хермансен Л., Оснес Дж. Б. (март 1972 г.). «PH крови и мышц после максимальной нагрузки у человека». Журнал прикладной физиологии. 32 (3): 304–8. Дои:10.1152 / jappl.1972.32.3.304. PMID 5010039.
- ^ Ghovanloo MR, Abdelsayed M, Peters CH, Ruben PC (апрель 2018 г.). «Смешанный периодический мутант паралича и миотонии, P1158S, придает чувствительность к pH в натриевых каналах скелетных мышц, управляемых напряжением». Научные отчеты. 8 (1): 6304. Bibcode:2018НатСР ... 8.6304G. Дои:10.1038 / с41598-018-24719-у. ЧВК 5908869. PMID 29674667.
- ^ Ghovanloo MR, Peters CH, Ruben PC (октябрь 2018 г.). «Влияние ацидоза на нейронные потенциалзависимые натриевые каналы: Nav1.1 и Nav1.3». каналы. 12 (1): 367–377. Дои:10.1080/19336950.2018.1539611. ЧВК 6284583. PMID 30362397.
- ^ Моран Ю., Барзилай М.Г., Либескинд Б.Дж., Закон Х.Х. (февраль 2015 г.). «Эволюция потенциалзависимых ионных каналов при появлении Metazoa». Журнал экспериментальной биологии. 218 (Pt 4): 515–25. Дои:10.1242 / jeb.110270. PMID 25696815.
- ^ Liebeskind BJ, Hillis DM, Zakon HH (май 2011 г.). «Развитие натриевых каналов предшествовало возникновению нервной системы у животных». Труды Национальной академии наук Соединенных Штатов Америки. 108 (22): 9154–9. Bibcode:2011ПНАС..108.9154Л. Дои:10.1073 / pnas.1106363108. ЧВК 3107268. PMID 21576472.
- ^ Кинг Н., Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. И др. (Февраль 2008 г.). «Геном хоанофлагеллаты Monosiga brevicollis и происхождение многоклеточных животных». Природа. 451 (7180): 783–8. Bibcode:2008Натура.451..783K. Дои:10.1038 / природа06617. ЧВК 2562698. PMID 18273011.
- ^ а б Стронг М., Чанди К.Г., Гутман Г.А. (январь 1993 г.). «Молекулярная эволюция генов потенциалочувствительных ионных каналов: о происхождении электрической возбудимости». Молекулярная биология и эволюция. 10 (1): 221–42. Дои:10.1093 / oxfordjournals.molbev.a039986. PMID 7680747.
- ^ Liebeskind BJ, Hillis DM, Zakon HH (ноябрь 2013 г.). «Независимое определение селективности натрия в натриевых каналах бактерий и животных». Текущая биология. 23 (21): R948–9. Дои:10.1016 / j.cub.2013.09.025. PMID 24200318.
- ^ Касимова М.А., Граната Д., Карневале В. (2016). Натриевые каналы, управляемые напряжением: история эволюции и отличительные особенности последовательности. Актуальные темы в мембранах. 78. С. 261–86. Дои:10.1016 / bs.ctm.2016.05.002. ISBN 9780128053867. PMID 27586287.
- ^ а б Видмарк Дж., Сундстрём Дж., Окампо Даса Д., Лархаммар Д. (январь 2011 г.). «Дифференциальная эволюция потенциалозависимых натриевых каналов у четвероногих и костистых рыб». Молекулярная биология и эволюция. 28 (1): 859–71. Дои:10.1093 / molbev / msq257. PMID 20924084.
- ^ а б Zakon HH, Jost MC, Lu Y (апрель 2011 г.). «Расширение семейства генов потенциал-зависимых Na + -каналов у ранних четвероногих совпало с появлением наземных форм и повышенной сложности мозга». Молекулярная биология и эволюция. 28 (4): 1415–24. Дои:10.1093 / molbev / msq325. ЧВК 3058772. PMID 21148285.
внешняя ссылка
- Натрий + каналы в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- "Натриевые каналы, управляемые напряжением". База данных рецепторов и ионных каналов IUPHAR. Международный союз фундаментальной и клинической фармакологии.