Грибок - Fungus
| Грибы | |
|---|---|
 | |
По часовой стрелке сверху слева:
| |
| Научная классификация | |
| (без рейтинга): | Опистоконта |
| (без рейтинга): | Голомикота |
| (без рейтинга): | Зооспория |
| Королевство: | Грибы (Л. ) Р. Т. Мур[1] |
| Подцарства / Phyla | |
| |
А грибок (множественное число: грибы[2] или же грибки[3]) является любым членом группы эукариотический организмов, которые включают микроорганизмы, такие как дрожжи и формы, а также более знакомые грибы. Эти организмы классифицируются как Королевство, который отделен от других эукариотических жизненных царств растения и животные.
Характеристика, которая помещает грибы в царство, отличное от растений, бактерий и некоторых простейших, - это хитин в их клеточные стенки. Как и животные, грибы гетеротрофы; они получают пищу, поглощая растворенные молекулы, обычно выделяя пищеварительные ферменты в их среду. Грибы не фотосинтезировать. Рост - их средство мобильность, за исключением спор (некоторые из которых жгутиковые), которые могут перемещаться по воздуху или воде. Грибы - главное разлагатели в экологических системах. Эти и другие различия помещают грибы в одну группу родственных организмов, названных Eumycota (настоящие грибы или же Eumycetes), которые разделяют общий предок (из монофилетический группа), интерпретация, которая также решительно поддерживается молекулярная филогенетика. Эта группа грибов отличается от сходных по строению миксомицеты (слизевики) и оомицеты (водяные формы). Дисциплина биология посвященный изучению грибов, известен как микология (от Греческий μύκης Mykes, гриб). В прошлом микология считалась отраслью ботаника, хотя теперь известно, что грибы генетически более близки к животным, чем к растениям.
Большинство грибов, распространенных во всем мире, незаметны из-за небольшого размера их структур и загадочный образ жизни в почве или на мертвом теле. Грибы включают симбионты растений, животных или других грибов, а также паразиты. Они могут стать заметными, когда плодоношение в виде грибов или плесени. Грибы играют важную роль в разложении органических веществ и играют важную роль в питательных веществах. кататься на велосипеде и обмен в окружающей среде. Они издавна использовались в качестве прямого источника пищи для человека в виде грибов и трюфели; как закваска средство для хлеба; и в ферментация различных пищевых продуктов, таких как вино, пиво, и соевый соус. С 1940-х годов грибы использовались для производства антибиотики, а в последнее время - различные ферменты производимые грибами используются промышленно И в моющие средства. Грибы также используются как биологические пестициды для борьбы с сорняками, болезнями растений и насекомыми-вредителями. Многие виды производят биоактивный соединения, называемые микотоксины, Такие как алкалоиды и поликетиды, которые токсичны для животных, включая человека. Плодовые структуры некоторых видов содержат психотропный соединения и потребляются развлекательно или в традиционных духовные церемонии. Грибы могут разрушать производимые материалы и здания и становиться значительными патогены людей и других животных. Потери посевов из-за грибковых заболеваний (например, рисовая болезнь ) или еда порча может иметь большое влияние на человека запасы еды и местная экономика.
Царство грибов включает в себя огромное разнообразие таксоны с разнообразной экологией, жизненный цикл стратегии и морфологии начиная от одноклеточных водных хитриды к крупным грибам. Однако мало что известно об истинном биоразнообразие of Kingdom Fungi, численность которого оценивается в 2,2–3,8 миллиона видов.[4] Из них описано только около 120 000 видов, из которых известно, что более 8 000 видов вредны для растений, и по меньшей мере 300 видов могут быть патогенными для человека.[5] С начала 18-19 веков таксономический произведения Карл Линней, Кристиан Хендрик Персун, и Элиас Магнус Фрис, грибы были классифицированный в соответствии с их морфологией (например, такими характеристиками, как цвет спор или микроскопические особенности) или физиология. Достижения в молекулярная генетика открыли путь для Анализ ДНК для включения в таксономию, которая иногда бросает вызов историческим группировкам, основанным на морфологии и других признаках. Филогенетический исследования, опубликованные в первом десятилетии XXI века, помогли изменить классификацию Kingdom Fungi, которая разделена на одну субцарство, Семь тип, и десять подтипов.
Этимология
Английское слово грибок напрямую заимствован из латинский грибок (гриб), использованный в трудах Гораций и Плиний.[6] Это, в свою очередь, происходит из Греческий слово сфонги (σφόγγος «губка»), что относится к макроскопический строение и морфология грибов и плесени;[7] корень также используется в других языках, таких как немецкий Schwamm («губка») и Шиммель ("плесень").[8]
Слово микология происходит от греческого Mykes (μύκης «гриб») и логотипы (λόγος «дискурс»).[9] Это означает научное изучение грибов. Латинская форма прилагательного слова «микология» (микологическийæ) появился еще в 1796 году в книге по этому поводу автора Кристиан Хендрик Персун.[10] Это слово появилось в английском языке еще в 1824 году в книге А. Роберт Кэй Гревиль.[11] В 1836 г. английский натуралист Майлз Джозеф Беркли публикация Английская флора сэра Джеймса Эдварда Смита, Vol. 5. также относится к микологии как к изучению грибов.[7][12]
Группа всех грибов, присутствующих в определенной области или географическом регионе, известна как микобиота (существительное во множественном числе, без единственного числа), например, «микобиота Ирландии».[13]
Характеристики


До введения молекулярные методы для филогенетического анализа, систематики считается, что грибы являются членами царство растений из-за сходства в образе жизни: и грибы, и растения в основном неподвижен, и имеют сходство в общей морфологии и среде обитания. Как и растения, грибы часто растут в почве, а в случае грибы, образуют заметные плодовые тела, которые иногда напоминают растения, такие как мхи. Грибы теперь считаются отдельным царством, отличным от растений и животных, от которых они, по-видимому, имеют расходился около миллиарда лет назад (примерно в начале Неопротерозойский Эра).[14][15] Некоторые морфологические, биохимические и генетические особенности являются общими с другими организмами, в то время как другие уникальны для грибов, что четко отделяет их от других царств:
Общие функции:
- С другим эукариоты: Грибковые клетки содержат мембраносвязанный ядра с хромосомы которые содержат ДНК с некодирующие области называется интроны и кодирующие области, называемые экзоны. У грибов есть мембраносвязанная цитоплазматическая органеллы Такие как митохондрии, стерол -содержащие мембраны, и рибосомы из 80-е годы тип.[16] У них есть характерный диапазон растворимых углеводов и запасных соединений, в том числе сахарные спирты (например., маннитол ), дисахариды, (например, трегалоза ), и полисахариды (например., гликоген, который также встречается у животных[17]).
- С животными: отсутствие грибов хлоропласты и есть гетеротрофный организмов и поэтому требуют предварительно сформированных органические соединения как источники энергии.[18]
- С растениями: у грибов есть клеточная стенка[19] и вакуоли.[20] Они размножаются как половым, так и бесполым путем и любят базальный группы растений (например, папоротники и мхи ) производить споры. Подобно мхам и водорослям, грибы обычно имеют гаплоидный ядра.[21]
- С эвгленоиды и бактерии: высшие грибы, эвгленоиды и некоторые бактерии производят аминокислота L-лизин в частности биосинтез шаги, называемые α-аминоадипатный путь.[22][23]
- Клетки большинства грибов растут в виде трубчатых, удлиненных и нитевидных (нитчатых) структур, называемых гифы, которые могут содержать несколько ядер и расширяться за счет роста на концах. Каждая подсказка содержит набор агрегированных пузырьки - клеточные структуры, состоящие из белки, липиды, и другие органические молекулы, называемые Spitzenkörper.[24] И грибы, и оомицеты растут как нитчатые клетки гиф.[25] Напротив, похожие на вид организмы, такие как нитчатые зеленые водоросли, растут путем многократного деления клеток в цепочке клеток.[17] Есть еще одноклеточные грибы (дрожжи ), которые не образуют гиф, а некоторые грибы имеют как гифы, так и дрожжевые формы.[26]
- Как и некоторые виды растений и животных, более 70 видов грибов отображать биолюминесценция.[27]
Уникальные черты:
- Некоторые виды растут как одноклеточные дрожжи, которые размножаются подающий надежды или же деление. Диморфные грибы может переключаться между дрожжевой фазой и фазой гиф в зависимости от условий окружающей среды.[26]
- Клеточная стенка грибка состоит из глюканы и хитин; в то время как глюканы также содержатся в растениях, а хитин - в экзоскелет из членистоногие,[28][29] грибы - единственные организмы, которые объединяют эти две структурные молекулы в своей клеточной стенке. В отличие от растений и оомицетов, клеточные стенки грибов не содержат целлюлозы.[30]

У большинства грибов отсутствует эффективная система для переноса воды и питательных веществ на большие расстояния, например ксилема и флоэма у многих растений. Чтобы преодолеть это ограничение, некоторые грибы, такие как Армиллярия, форма ризоморфы,[31] которые напоминают и выполняют функции, аналогичные корни растений. Грибы, как эукариоты, обладают биосинтетический путь для производства терпены который использует мевалоновая кислота и пирофосфат в качестве химические строительные блоки.[32] У растений и некоторых других организмов есть дополнительный путь биосинтеза терпена в своих хлоропластах, структура которого отсутствует у грибов и животных.[33] Грибы производят несколько вторичные метаболиты которые похожи или идентичны по структуре тем, которые производятся растениями.[32] Многие из растительных и грибковых ферментов, которые делают эти соединения, отличаются друг от друга последовательность и другие характеристики, которые указывают на раздельное происхождение и конвергентную эволюцию этих ферментов в грибах и растениях.[32][34]
Разнообразие

Грибы распространены по всему миру и растут в самых разных средах обитания, включая экстремальные условия, такие как пустыни или в районах с высокой концентрацией соли[35] или же ионизирующего излучения,[36] а также в глубокое море отложения.[37] Некоторые могут пережить интенсивные УФ и космическое излучение встречаются во время космических путешествий.[38] Большинство из них растут в наземных средах, хотя некоторые виды живут частично или исключительно в водных средах, например, Читрид грибок Batrachochytrium dendrobatidis, а паразит что привело к снижению во всем мире амфибия населения. Этот организм проводит часть своего жизненного цикла в качестве подвижного зооспора, позволяя ему продвигаться по воде и входить в своего хозяина-амфибии.[39] Другие примеры водных грибов включают те, которые живут в гидротермальный районы океана.[40]
Было обнаружено около 120000 видов грибов. описанный к систематики,[41] но глобальное биоразнообразие царства грибов до конца не изучено.[41] По оценкам 2017 года, может быть от 2,2 до 3,8 миллиона видов.[4] В микологии виды исторически различались с помощью множества методов и концепций. Классификация на основе морфологический Такие характеристики, как размер и форма спор или плодовых структур, традиционно преобладали в таксономии грибов.[42] Виды также можно различать по их биохимический и физиологический характеристики, такие как их способность метаболизировать определенные биохимические вещества или их реакция на химические тесты. В концепция биологических видов различает виды на основе их способности приятель. Применение молекулярный инструменты, такие как Секвенирование ДНК и филогенетический анализ для изучения разнообразия значительно повысил разрешающую способность и повысил надежность оценок генетическое разнообразие внутри различных таксономических групп.[43]
Микология
Микология это филиал биология связаны с систематическим изучением грибов, включая их генетические и биохимические свойства, их таксономию, и их использование для людей в качестве источника лекарств, продуктов питания и психотропные вещества употребляемые в религиозных целях, а также их опасности, такие как отравление или инфекция. Поле фитопатология, изучение болезней растений, тесно связано, потому что многие патогены растений являются грибами.[44]

Использование грибов людьми восходит к предыстории; Эци-ледяной человек, хорошо сохранившаяся мумия 5300-летнего Неолит человек, найденный замороженным в австрийских Альпах, нес два вида полипор грибы, которые, возможно, использовались как Трут (Fomes fomentarius ) или в лечебных целях (Piptoporus betulinus ).[45] Древние народы тысячелетиями использовали грибы в качестве источников пищи - часто неосознанно - при приготовлении квасного хлеба и сброженных соков. Некоторые из самых старых письменных источников содержат упоминания об уничтожении посевов, которое, вероятно, было вызвано патогенными грибами.[46]
История
Микология - относительно новая наука, которая стала систематической после развития микроскоп в 17 веке. Хотя споры грибов впервые были обнаружены Джамбаттиста делла Порта в 1588 году основополагающим трудом в развитии микологии считается публикация Пьер Антонио Микели работа 1729 г. Nova plantarum родов.[47] Микели не только наблюдал за спорами, но и показал, что при определенных условиях они могут быть индуцированы к росту в те же виды грибов, от которых они произошли.[48] Расширение использования биномиальная система номенклатуры представлен Карл Линней в его Виды plantarum (1753), голландцы Кристиан Хендрик Персун (1761–1836) создал первую классификацию грибов с таким мастерством, что считается основоположником современной микологии. Потом, Элиас Магнус Фрис (1794–1878) далее разработал классификация грибов, используя цвет спор и микроскопические характеристики, методы, которые до сих пор используются систематиками. Другие известные ранние участники микологии в 17-19 и начале 20 веков включают: Майлз Джозеф Беркли, Август Карл Джозеф Корда, Антон де Бари, братья Луи Рене и Чарльз Туласне, Артур Х. Р. Буллер, Кертис Дж. Ллойд, и Пьер Андреа Саккардо. В 20-м и 21-м веках развитие биохимия, генетика, молекулярная биология, биотехнология, Секвенирование ДНК филогенетический анализ позволил по-новому взглянуть на взаимоотношения грибов и биоразнообразие, и бросил вызов традиционным группировкам на основе морфологии грибковых таксономия.[49]
Морфология
Микроскопические структуры
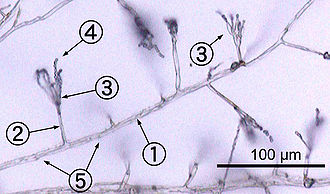
Большинство грибов растут как гифы, представляющие собой цилиндрические, нитевидные конструкции 2–10 мкм в диаметре и до нескольких сантиметров в длину. Гифы растут на концах (вершинах); новые гифы обычно образуются в результате появления новых кончиков вдоль существующих гиф в результате процесса, называемого разветвление, или иногда растущие вилки кончиков гиф, дающие начало двум параллельно растущим гифам.[50] Гифы также иногда сливаются при соприкосновении, этот процесс называется слиянием гиф (или анастомоз ). Эти процессы роста приводят к развитию мицелий, взаимосвязанная сеть гиф.[26] Гифы могут быть либо перегородка или же ценоцитарный. Септатные гифы разделены на отсеки, разделенные поперечными стенками (внутренними клеточными стенками, называемыми перегородками, которые образуются на прямые углы клеточной стенке, придающей гифу ее форму), причем каждый отсек содержит одно или несколько ядер; ценоцитарные гифы не разделены.[51] Септы есть поры что позволяет цитоплазма, органеллы, а иногда ядра, чтобы пройти; Примером может служить перегородка долипора у грибов типа Basidiomycota.[52] Ценоцитарные гифы по существу многоядерный суперячейки.[53]
Многие виды развили специализированные структуры гиф для поглощения питательных веществ от живых хозяев; примеры включают гаустория у видов растений-паразитов большинства типов грибов, и арбускулы из нескольких микоризный грибы, которые проникают в клетки-хозяева для потребления питательных веществ.[54]
Хотя грибы опистоконцы - группа эволюционно связанных организмов, в целом характеризующихся одним задним жгутик - все типы, кроме хитриды потеряли задние жгутики.[55] Грибы необычны среди эукариот тем, что имеют клеточную стенку, которая, помимо глюканы (например, β-1,3-глюкан) и другие типичные компоненты, также содержат биополимер хитин.[56]
Макроскопические структуры

Мицелий грибов может стать видимым невооруженным глазом, например, на различных поверхностях и субстраты, например, сырые стены и испорченная еда, где их обычно называют формы. Мицелий вырос на твердом агар СМИ в лаборатории чашки Петри обычно упоминаются как колонии. Эти колонии могут иметь форму роста и окраску (из-за спор или пигментация ), которые могут использоваться в качестве диагностических признаков при идентификации видов или групп.[57] Некоторые отдельные грибковые колонии могут достигать необычных размеров и возраста, как в случае клональный колония Армиллярия солидипес, который простирается на площади более 900 ха (3,5 квадратных миль), с предполагаемым возрастом около 9000 лет. годы.[58]
В апотеций - специализированная структура, важная в половое размножение у аскомицетов - плодовое тело чашеобразной формы, часто макроскопическое и удерживающее гимений, слой ткани, содержащий споровые клетки.[59] Плодовые тела базидиомицетов (базидиокарпы ), а некоторые аскомицеты иногда могут вырасти очень большими, и многие из них хорошо известны как грибы.
Рост и физиология

Рост грибов в виде гиф на твердых субстратах или в них или в виде отдельных клеток в водной среде адаптирован для эффективного извлечения питательных веществ, поскольку эти формы роста имеют высокий отношение площади поверхности к объему.[60] Гифы специально адаптированы для роста на твердых поверхностях и вторжения субстраты и ткани.[61] Они могут проявлять большие проникающие механические силы; например, многие патогены растений, включая Magnaporthe grisea, образуют структуру, называемую аппрессориум которые эволюционировали, чтобы прокалывать ткани растений.[62] Давление, создаваемое аппрессорием, направленное против растения. эпидермис, может превышать 8 мегапаскали (1200 фунтов на квадратный дюйм).[62] Нитчатый гриб Paecilomyces lilacinus использует аналогичную структуру для проникновения в яйца нематоды.[63]
Механическое давление, оказываемое аппрессорием, создается физиологическими процессами, которые увеличивают внутриклеточное тургор производя осмолиты Такие как глицерин.[64] Подобные адаптации дополняются гидролитические ферменты выделяется в окружающую среду для переваривания крупных органических молекул, таких как полисахариды, белки, и липиды - в более мелкие молекулы, которые затем могут быть поглощены как питательные вещества.[65][66][67] Подавляющее большинство нитчатых грибов растут полярным образом (распространяясь в одном направлении) за счет удлинения на кончике (вершине) гифы.[68] Другие формы роста грибов включают интеркалярное расширение (продольное расширение гифальных компартментов, расположенных ниже верхушки), как в случае некоторых эндофитный грибы,[69] или рост за счет увеличения объема во время развития грибов ножки и другие крупные органы.[70] Рост грибов как многоклеточные структуры состоящий из соматический и репродуктивные клетки - особенность, независимо развившаяся у животных и растений.[71]- выполняет несколько функций, включая развитие плодовых тел для распространения половых спор (см. Выше) и биопленки для колонизации субстрата и межклеточная связь.[72]
Грибы традиционно считаются гетеротрофы, организмы, которые полагаются исключительно на фиксированный углерод другими организмами для метаболизм. Грибы имеют развился высокая степень метаболической универсальности, которая позволяет им использовать широкий спектр органических субстратов для роста, включая простые соединения, такие как нитрат, аммиак, ацетат, или же этиловый спирт.[73][74] У некоторых видов пигмент меланин может играть роль в извлечении энергии из ионизирующего излучения, Такие как гамма-излучение. Эта форма "радиотрофный" рост был описан только для нескольких видов, влияние на скорость роста невелико, а лежащие в основе биофизический и биохимические процессы малоизвестны.[36] Этот процесс может иметь сходство с CO2 фиксация через видимый свет, но вместо этого использует ионизирующее излучение в качестве источника энергии.[75]
Размножение

Размножение грибов сложное, оно отражает различия в образе жизни и генетическом составе этого разнообразного царства организмов.[76] По оценкам, треть всех грибов размножается с использованием более чем одного метода размножения; например, воспроизводство может происходить в два четко дифференцированных этапа в пределах жизненный цикл вида, телеоморф и анаморф.[77] Условия окружающей среды запускают генетически детерминированные состояния развития, которые приводят к созданию специализированных структур для полового или бесполого размножения. Эти структуры способствуют размножению, эффективно рассеивая споры или содержащие споры. пропагулы.
Бесполое размножение
Бесполое размножение происходит через вегетативные споры (конидии ) или через мицелиальная фрагментация. Фрагментация мицелия происходит, когда мицелий гриба разделяется на части, и каждый компонент превращается в отдельный мицелий. Фрагментация мицелия и вегетативные споры поддерживают клональный население адаптировано к конкретному ниша, и допускают более быстрое распространение, чем половое размножение.[78] «Грибки несовершенные» (грибки, не имеющие идеальной или половой стадии) или Deuteromycota включают все виды, у которых отсутствует наблюдаемый половой цикл.[79] Deuteromycota не является общепринятой таксономической кладой, и теперь под ним понимаются просто грибы, у которых отсутствует известная половая стадия.
Половое размножение
Половое размножение с помощью мейоз непосредственно наблюдается во всех типах грибов, кроме Гломеромикота[80] (генетический анализ также предполагает мейоз у Glomeromycota). Он во многом отличается от полового размножения животных или растений. Различия также существуют между группами грибов и могут использоваться для различения видов по морфологическим различиям в половой структуре и репродуктивных стратегиях.[81][82] Эксперименты по спариванию грибковых изолятов могут определять виды на основе представлений о биологических видах.[82] Первоначально основные группы грибов были выделены на основании морфологии их половых структур и спор; например, споросодержащие структуры, asci и базидия, могут быть использованы для идентификации аскомицетов и базидиомицетов соответственно. Грибы используют двух системы спаривания: гетероталлический виды допускают спаривание только между особями противоположного тип вязки, в то время как гомоталлический виды могут спариваться и размножаться половым путем с любым другим человеком или с самим собой.[83]
Большинство грибов имеют как гаплоидный и диплоид стадии в их жизненных циклах. В грибах, размножающихся половым путем, совместимые особи могут объединяться, сливая свои гифы во взаимосвязанную сеть; этот процесс, анастомоз, требуется для начала полового цикла. Многие аскомицеты и базидиомицеты проходят через дикариотический стадия, на которой ядра, унаследованные от двух родителей, не объединяются сразу после слияния клеток, а остаются отдельными в клетках гиф (см. гетерокариоз ).[84]
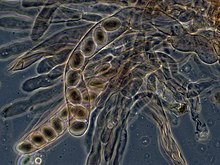
У аскомицетов дикариотические гифы гимений (слой споровой ткани) образуют характерный крюк у перегородки гифы. В течение деление клеток образование крючка обеспечивает правильное распределение вновь разделенных ядер на апикальный и базальный компартменты гиф. Аскус (множественное число asci), в котором кариогамия (ядерный синтез) происходит. Asci встроены в аскокарпий, или плодовое тело. Кариогамия в асках немедленно сопровождается мейозом и образованием аскоспоры. После распространения аскоспоры могут прорасти и образовать новый гаплоидный мицелий.[85]
Половое размножение у базидиомицетов похоже на половое размножение аскомицетов. Совместимые гаплоидные гифы сливаются с образованием дикариотического мицелия. Однако дикариотическая фаза более обширна у базидиомицетов, часто также присутствуя в вегетативно растущем мицелии. Специализированная анатомическая структура, называемая зажимное соединение, образуется на каждой перегородке гифы. Как и в случае со сходным по структуре крючком у аскомицетов, зажимное соединение у базидиомицетов требуется для контролируемого переноса ядер во время деления клеток, для поддержания дикариотической стадии с двумя генетически разными ядрами в каждом компартменте гиф.[86] А базидиокарпий формируется клубная структура, известная как базидия генерировать гаплоид базидиоспоры после кариогамии и мейоза.[87] Наиболее известными базидиокарпами являются грибы, но они могут принимать и другие формы (см. Морфология раздел).
У грибов, ранее классифицированных как Zygomycota гаплоидные гифы двух особей сливаются, образуя гаметангий, специализированная клеточная структура, которая становится плодородной гамета -продуцирующая клетка. Гаметангий превращается в зигоспора, толстостенная спора, образованная слиянием гамет. Когда зигоспора прорастает, она подвергается мейоз, генерируя новые гаплоидные гифы, которые затем могут образовывать бесполые спорангиоспоры. Эти спорангиоспоры позволяют грибку быстро распространяться и прорастать в новые генетически идентичные гаплоидные грибковые мицелии.[88]
Распространение спор
Как бесполые, так и половые споры или спорангиоспоры часто активно распространяются путем насильственного изгнания из их репродуктивных структур. Этот выброс обеспечивает выход спор из репродуктивных структур, а также их перемещение по воздуху на большие расстояния.

Специализированные механические и физиологические механизмы, а также структуры поверхности спор (например, гидрофобины ), обеспечивают эффективное удаление спор.[89] Например, структура споровые клетки у некоторых видов аскомицетов такова, что накопление вещества влияя на объем клеток и баланс жидкости, вызывает взрывной выброс спор в воздух.[90] Насильственное удаление отдельных спор, называемое баллистоспоры включает образование маленькой капли воды (капля Буллера), которая при контакте со спорой приводит к высвобождению ее снаряда с начальным ускорением более 10 000 грамм;[91] в результате спора выбрасывается 0,01–0,02 см, достаточное расстояние, чтобы он провалился через жабры или же поры в воздух внизу.[92] Другие грибы, такие как клубнички, полагаться на альтернативные механизмы высвобождения спор, такие как внешние механические силы. В гидноидные грибы (зубные грибы) образуют споры на висячих, зубных или шиповидных выступах.[93] В грибы птичьего гнезда используйте силу падающих капель воды, чтобы высвободить споры из чашевидных плодовых тел.[94] Другая стратегия видна в вонючие рога, группа грибов с яркими цветами и гнилостным запахом, которые привлекают насекомых для распространения их спор.[95]
Большинство исследованных видов грибов переносятся ветром.[96][97] Такие виды часто образуют сухие или гидрофобные споры, которые не поглощают воду и легко рассеиваются, например, каплями дождя.[96][98][99]
Гомоталлизм
В гомоталлический половое размножение, два гаплоидный ядра, полученные из одного и того же индивидуального предохранителя, чтобы сформировать зигота что затем может пройти мейоз. Гомоталлические грибы включают виды с аспергиллоподобной бесполой стадией (анаморфы), встречающиеся во множестве различных родов,[100] несколько видов аскомицет род Cochliobolus,[101] и аскомицет Pneumocystis jiroveccii.[102] Heitman[103] рассмотрели доказательства, касающиеся эволюции полового размножения у грибов, и пришли к выводу, что самым ранним способом полового размножения среди эукариот, вероятно, был гомоталлизм, то есть самооплодотворение однополое размножение.
Другие половые процессы
Помимо регулярного полового размножения с мейозом, некоторые грибы, например, из этих родов Пенициллий и Аспергиллы, может обмениваться генетическим материалом через парасексуальный процессы, инициированные анастомозом между гифами и плазмогамия грибковых клеток.[104] Частота и относительная важность парасексуальных событий неясна и может быть ниже, чем у других половых процессов. Известно, что он играет роль во внутривидовой гибридизации.[105] и, вероятно, требуется для гибридизации между видами, которая была связана с основными событиями в эволюции грибов.[106]
Эволюция

В отличие от растения и животные, ранняя летопись окаменелостей грибов скудна. Факторы, которые, вероятно, способствуют недостаточной представленности видов грибов среди ископаемых, включают природу грибов. плодовые тела, которые представляют собой мягкие, мясистые и легко разлагаемые ткани, а также микроскопические размеры большинства грибковых структур, которые, следовательно, не так очевидны. Окаменелости грибов трудно отличить от окаменелостей других микробов, и их легче всего идентифицировать, если они напоминают сохранившийся грибы.[107] Часто восстанавливается после перминерализованный растения или животные-хозяева, эти образцы обычно изучаются путем изготовления препаратов тонких срезов, которые можно исследовать с помощью световая микроскопия или же просвечивающая электронная микроскопия.[108] Исследователи изучают окаменелости сжатия растворяя окружающую матрицу кислотой, а затем используя свет или сканирующая электронная микроскопия изучить детали поверхности.[109]
Самые ранние окаменелости, обладающие характеристиками грибов, датируются Палеопротерозой эра, некоторые 2,400 миллион лет назад (Ма ); эти многоклеточные бентосный у организмов были нитчатые структуры, способные к анастомоз.[110] Другие исследования (2009 г.) оценивают прибытие грибковых организмов примерно в 760–1060 человек. Млн лет назад на основе сравнений скорости эволюции близкородственных групп.[111] Для большей части Палеозой Эра (542–251 гг. Ma), грибы, по-видимому, были водными и состояли из организмов, похожих на современные хитриды наличие спор, несущих жгутик.[112] Эволюционная адаптация от водного к наземному образу жизни потребовала диверсификации экологических стратегий получения питательных веществ, в том числе паразитизм, сапробизм, и развитие мутуалистический такие отношения как микориза и лихенизация.[113] Недавние (2009 г.) исследования показывают, что экологическое состояние предков Аскомикота был сапробизм, и это независимое лихенизация события произошли несколько раз.[114]
В мае 2019 года ученые сообщили об открытии окаменелый гриб, названный Ourasphaira giraldae, в Канадская Арктика, которые, возможно, выросли на суше миллиард лет назад, задолго до растения жили на суше.[115][116][117] Ранее предполагалось, что грибы заселили землю во время Кембрийский (542–488.3 Ма), также задолго до наземных растений.[118] Ископаемые гифы и споры, извлеченные из Ордовик штата Висконсин (460 Ма) напоминают современные Гломералы, и существовала в то время, когда наземная флора, вероятно, состояла только из несосудистых мохообразный -подобные растения.[119] Прототакситы, который, вероятно, был грибком или лишайником, был бы самым высоким организмом последнего Силурийский и рано Девонский. Окаменелости грибов не стали обычным явлением и не вызывали споров до начала Девонский (416–359.2 Млн лет назад), когда они в большом количестве встречаются в Рини черт, в основном как Zygomycota и Chytridiomycota.[118][120][121] Примерно в это же время примерно 400 Ma, Ascomycota и Basidiomycota разошлись,[122] и все современные классы грибов присутствовали поздним Каменноугольный (Пенсильванский, 318.1–299 Ма).[123]
Лишайник -подобные окаменелости были обнаружены в Формация Доушантуо на юге Китая датируется 635–551 гг. Ма.[124] Лишайники являлись компонентом ранних наземных экосистем, и предполагаемый возраст самого старого ископаемого наземного лишайника составляет 400 лет. Ма;[125] эта дата соответствует возрасту самого старого из известных спорокарпий ископаемое, а Палеопиреномициты виды, найденные в Rhynie Chert.[126] Самая старая окаменелость с микроскопическими чертами, напоминающими современных базидиомицетов, - это Палеоанциструс, найденный перминерализованным с папоротник из Пенсильвании.[127] Редко в летописи окаменелостей встречаются Homobasidiomycetes (a таксон примерно эквивалентно производящим грибы видам Агарикомицеты ). Два Янтарь -сохраненные образцы свидетельствуют о том, что самые ранние известные грибообразующие грибы (вымершие виды Археомаразмиус леггетти ) появились в конце Меловой, 90 Ма.[128][129]
Через некоторое время после Пермско-триасовое вымирание (251.4 Ma), грибковый шип (первоначально предполагалось, что это необычайное изобилие грибковых спор в отложения ), предполагая, что грибы были доминирующей формой жизни в то время, представляя почти 100% доступного Окаменелости на этот период.[130] Однако относительная доля спор грибов по отношению к спорам, образованным водоросль species is difficult to assess,[131] the spike did not appear worldwide,[132][133] and in many places it did not fall on the Permian–Triassic boundary.[134]
65 million years ago, immediately after the Cretaceous–Paleogene extinction event that famously killed off most dinosaurs, there is a dramatic increase in evidence of fungi, apparently the death of most plant and animal species leading to a huge fungal bloom like "a massive compost heap".[135]
Таксономия
Although commonly included in botany curricula and textbooks, fungi are more closely related to животные than to plants and are placed with the animals in the монофилетический group of opisthokonts.[136] Analyses using молекулярная филогенетика support a монофилетический origin of fungi.[43] В таксономия of fungi is in a state of constant flux, especially due to recent research based on DNA comparisons. These current phylogenetic analyses often overturn classifications based on older and sometimes less discriminative methods based on morphological features and biological species concepts obtained from experimental matings.[137]
There is no unique generally accepted system at the higher taxonomic levels and there are frequent name changes at every level, from species upwards. Efforts among researchers are now underway to establish and encourage usage of a unified and more consistent nomenclature.[43][138] Fungal species can also have multiple scientific names depending on their life cycle and mode (sexual or asexual) of reproduction. Web sites such as Index Fungorum и ЭТО list current names of fungal species (with cross-references to older synonyms).
The 2007 classification of Kingdom Fungi is the result of a large-scale collaborative research effort involving dozens of mycologists and other scientists working on fungal taxonomy.[43] It recognizes seven phyla, two of which—the Ascomycota and the Basidiomycota—are contained within a branch representing subkingdom Dikarya, the most species rich and familiar group, including all the mushrooms, most food-spoilage molds, most plant pathogenic fungi, and the beer, wine, and bread yeasts. The accompanying кладограмма depicts the major fungal таксоны and their relationship to opisthokont and unikont organisms, based on the work of Philippe Silar,[139] "The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research"[140] and Tedersoo et al. 2018.[141] The lengths of the branches are not proportional to эволюционный distances.
| Zoosporia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Базидиомицеты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Аскомикота |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxonomic groups

The major phyla (sometimes called divisions) of fungi have been classified mainly on the basis of characteristics of their sexual reproductive structures. Currently, seven phyla are proposed: Microsporidia, Chytridiomycota, Blastocladiomycota, Neocallimastigomycota, Glomeromycota, Ascomycota, and Basidiomycota.[43]
Phylogenetic analysis has demonstrated that the Microsporidia, unicellular parasites of animals and protists, are fairly recent and highly derived endobiotic fungi (living within the tissue of another species).[112][142] One 2006 study concludes that the Microsporidia are a sister group to the true fungi; that is, they are each other's closest evolutionary relative.[143] Hibbett and colleagues suggest that this analysis does not clash with their classification of the Fungi, and although the Microsporidia are elevated to phylum status, it is acknowledged that further analysis is required to clarify evolutionary relationships within this group.[43]
В Chytridiomycota are commonly known as chytrids. These fungi are distributed worldwide. Chytrids and their close relatives Neocallimastigomycota и Blastocladiomycota (below) are the only fungi with active motility, producing zoospores that are capable of active movement through aqueous phases with a single flagellum, leading early taxonomists to classify them as protists. Molecular phylogenies, inferred from rRNA sequences in рибосомы, suggest that the Chytrids are a базальный group divergent from the other fungal phyla, consisting of four major clades with suggestive evidence for paraphyly or possibly polyphyly.[144]
В Blastocladiomycota were previously considered a taxonomic clade within the Chytridiomycota. Recent molecular data and ultrastructural characteristics, however, place the Blastocladiomycota as a sister clade to the Zygomycota, Glomeromycota, and Dikarya (Ascomycota and Basidiomycota). The blastocladiomycetes are saprotrophs, feeding on decomposing organic matter, and they are parasites of all eukaryotic groups. Unlike their close relatives, the chytrids, most of which exhibit zygotic meiosis, the blastocladiomycetes undergo sporic meiosis.[112]
В Neocallimastigomycota were earlier placed in the phylum Chytridomycota. Members of this small phylum are anaerobic organisms, living in the digestive system of larger herbivorous mammals and in other terrestrial and aquatic environments enriched in cellulose (e.g., domestic waste landfill sites).[145] They lack mitochondria but contain hydrogenosomes of mitochondrial origin. As in the related chrytrids, neocallimastigomycetes form zoospores that are posteriorly uniflagellate or polyflagellate.[43]


Члены Гломеромикота form arbuscular mycorrhizae, a form of mutualist симбиоз wherein fungal hyphae invade plant root cells and both species benefit from the resulting increased supply of nutrients. All known Glomeromycota species reproduce asexually.[80] The symbiotic association between the Glomeromycota and plants is ancient, with evidence dating to 400 million years ago.[146] Formerly part of the Zygomycota (commonly known as 'sugar' and 'pin' molds), the Glomeromycota were elevated to phylum status in 2001 and now replace the older phylum Zygomycota.[147] Fungi that were placed in the Zygomycota are now being reassigned to the Glomeromycota, or the subphyla incertae sedis Mucoromycotina, Kickxellomycotina, то Zoopagomycotina и Entomophthoromycotina.[43] Some well-known examples of fungi formerly in the Zygomycota include black bread mold (Rhizopus stolonifer ), и Pilobolus species, capable of ejecting споры several meters through the air.[148] Medically relevant genera include Mucor, Rhizomucor, и Rhizopus.
В Аскомикота, commonly known as sac fungi or ascomycetes, constitute the largest taxonomic group within the Eumycota.[42] These fungi form meiotic spores called ascospores, which are enclosed in a special sac-like structure called an ascus. This phylum includes morels, a few mushrooms и truffles, unicellular yeasts (e.g., of the genera Saccharomyces, Kluyveromyces, Пичиа, и Candida ), and many filamentous fungi living as saprotrophs, parasites, and mutualistic symbionts (e.g. lichens). Prominent and important genera of filamentous ascomycetes include Aspergillus, Penicillium, Fusarium, и Claviceps. Many ascomycete species have only been observed undergoing asexual reproduction (called anamorphic species), but analysis of molecular data has often been able to identify their closest teleomorphs in the Ascomycota.[149] Because the products of meiosis are retained within the sac-like ascus, ascomycetes have been used for elucidating principles of genetics and heredity (e.g., Neurospora crassa ).[150]
Члены Базидиомицеты, commonly known as the club fungi or basidiomycetes, produce meiospores called basidiospores on club-like stalks called basidia. Most common mushrooms belong to this group, as well as ржавчина и головневые грибы, which are major pathogens of grains. Other important basidiomycetes include the кукуруза pathogen Ustilago maydis,[151] human commensal species of the genus Malassezia,[152] и opportunistic human pathogen, Cryptococcus neoformans.[153]
Fungus-like organisms
Because of similarities in morphology and lifestyle, the slime molds (mycetozoans, plasmodiophorids, acrasids, Fonticula и labyrinthulids, now in Амебозоа, Rhizaria, Экскавата, Opisthokonta и Stramenopiles, respectively), water molds (oomycetes ) и hyphochytrids (both Stramenopiles ) were formerly classified in the kingdom Fungi, in groups like Mastigomycotina, Gymnomycota и Phycomycetes. The slime molds were studied also as protozoans, leading to an ambiregnal, duplicated taxonomy.
Unlike true fungi, the cell walls of oomycetes contain cellulose and lack хитин. Hyphochytrids have both chitin and cellulose. Slime molds lack a cell wall during the assimilative phase (except labyrinthulids, which have a wall of scales), and ingest nutrients by ingestion (phagocytosis, except labyrinthulids) rather than absorption (osmotrophy, as fungi, labyrinthulids, oomycetes and hyphochytrids). Neither water molds nor slime molds are closely related to the true fungi, and, therefore, taxonomists no longer group them in the kingdom Fungi. Nonetheless, studies of the oomycetes and myxomycetes are still often included in mycology textbooks and primary research literature.[154]
В Eccrinales и Amoebidiales находятся opisthokont protists, previously thought to be zygomycete fungi. Other groups now in Opisthokonta (например., Corallochytrium, Ichthyosporea ) were also at given time classified as fungi. Род Blastocystis, now in Stramenopiles, was originally classified as a yeast. Ellobiopsis, now in Alveolata, was considered a chytrid. В бактерии were also included in fungi in some classifications, as the group Schizomycetes.
В Rozellida clade, including the "ex-chytrid" Rozella, is a genetically disparate group known mostly from environmental DNA sequences that is a sister group to fungi. Members of the group that have been isolated lack the chitinous cell wall that is characteristic of fungi.
В nucleariids may be the next sister group to the eumycete clade, and as such could be included in an expanded fungal kingdom.[136]Много Actinomycetales (Актинобактерии ), a group with many filamentous bacteria, were also long believed to be fungi.[155][156]
Экология

Although often inconspicuous, fungi occur in every environment on земной шар and play very important roles in most ecosystems. Along with bacteria, fungi are the major decomposers in most terrestrial (and some aquatic) ecosystems, and therefore play a critical role in biogeochemical cycles[157] and in many food webs. As decomposers, they play an essential role in nutrient cycling, especially as saprotrophs и симбионты, degrading organic matter to inorganic molecules, which can then re-enter anabolic metabolic pathways in plants or other organisms.[158][159]
Симбиоз
Many fungi have important симбиотический relationships with organisms from most if not all kingdoms.[160][161][162] These interactions can be мутуалистический or antagonistic in nature, or in the case of commensal fungi are of no apparent benefit or detriment to the host.[163][164][165]
With plants
Микоризный symbiosis between растения and fungi is one of the most well-known plant–fungus associations and is of significant importance for plant growth and persistence in many ecosystems; over 90% of all plant species engage in mycorrhizal relationships with fungi and are dependent upon this relationship for survival.[166]
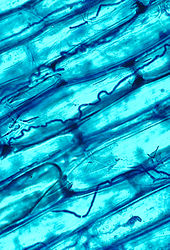
The mycorrhizal symbiosis is ancient, dating back to at least 400 million years.[146] It often increases the plant's uptake of inorganic compounds, such as nitrate и phosphate from soils having low concentrations of these key plant nutrients.[158][167] The fungal partners may also mediate plant-to-plant transfer of carbohydrates and other nutrients.[168] Such mycorrhizal communities are called "common mycorrhizal networks ".[169][170] A special case of mycorrhiza is myco-heterotrophy, whereby the plant parasitizes the fungus, obtaining all of its nutrients from its fungal symbiont.[171] Some fungal species inhabit the tissues inside roots, stems, and leaves, in which case they are called endophytes.[172] Similar to mycorrhiza, endophytic colonization by fungi may benefit both symbionts; for example, endophytes of grasses impart to their host increased resistance to herbivores and other environmental stresses and receive food and shelter from the plant in return.[173]
With algae and cyanobacteria

Лишайники are a symbiotic relationship between fungi and фотосинтетический водоросли или же cyanobacteria. The photosynthetic partner in the relationship is referred to in lichen terminology as a "photobiont". The fungal part of the relationship is composed mostly of various species of ascomycetes and a few basidiomycetes.[174] Lichens occur in every ecosystem on all continents, play a key role in soil formation and the initiation of biological succession,[175] and are prominent in some extreme environments, including polar, alpine, и semiarid desert regions.[176] They are able to grow on inhospitable surfaces, including bare soil, rocks, tree bark, wood, shells, barnacles and leaves.[177] As in mycorrhizas, the photobiont provides sugars and other carbohydrates via photosynthesis to the fungus, while the fungus provides minerals and water to the photobiont. The functions of both symbiotic organisms are so closely intertwined that they function almost as a single organism; in most cases the resulting organism differs greatly from the individual components. Lichenization is a common mode of nutrition for fungi; around 20% of fungi—between 17,500 and 20,000 described species—are lichenized.[178] Characteristics common to most lichens include obtaining organic carbon by photosynthesis, slow growth, small size, long life, long-lasting (seasonal) vegetative reproductive structures, mineral nutrition obtained largely from airborne sources, and greater tolerance of desiccation than most other photosynthetic organisms in the same habitat.[179]
With insects
Many insects also engage in mutualistic relationships with fungi. Several groups of ants cultivate fungi in the order Agaricales as their primary food source, while ambrosia beetles cultivate various species of fungi in the bark of trees that they infest.[180] Likewise, females of several wood wasp species (genus Sirex ) inject their eggs together with spores of the wood-rotting fungus Amylostereum areolatum into the sapwood из сосна trees; the growth of the fungus provides ideal nutritional conditions for the development of the wasp larvae.[181] At least one species of stingless bee has a relationship with a fungus in the genus Monascus, where the larvae consume and depend on fungus transferred from old to new nests.[182] Termites on the African savannah are also known to cultivate fungi,[160] and yeasts of the genera Candida и Lachancea inhabit the gut of a wide range of insects, including neuropterans, beetles, и cockroaches; it is not known whether these fungi benefit their hosts.[183] Fungi ingrowing dead wood are essential for xylophagous insects (e.g. woodboring beetles ).[184][185][186] They deliver nutrients needed by xylophages to nutritionally scarce dead wood.[187][185][186] Thanks to this nutritional enrichment the larvae of woodboring insect is able to grow and develop to adulthood.[184] The larvae of many families of fungicolous flies, particularly those within the superfamily Sciaroidea такой как Mycetophilidae и немного Keroplatidae feed on fungal fruiting bodies and sterile mycorrhizae.[188]
As pathogens and parasites

Many fungi are паразиты on plants, animals (including humans), and other fungi. Serious pathogens of many cultivated plants causing extensive damage and losses to agriculture and forestry include the rice blast грибок Magnaporthe oryzae,[189] tree pathogens such as Ophiostoma ulmi и Ophiostoma novo-ulmi causing Болезнь голландского вяза[190] и Cryphonectria parasitica responsible for chestnut blight,[191] and plant pathogens in the genera Fusarium, Ustilago, Alternaria, и Cochliobolus.[164] Немного carnivorous fungi, like Paecilomyces lilacinus, находятся хищники из нематоды, which they capture using an array of specialized structures such as constricting rings or adhesive nets.[192] Many fungi that are plant pathogens, such as Magnaporthe oryzae, can switch from being biotrophic (parasitic on living plants) to being necrotrophic (feeding on the dead tissues of plants they have killed).[193] This same principle is applied to fungi-feeding parasites, including Asterotremella albida, which feeds on the fruit bodies of other fungi both while they are living and after they are dead.[194]
Some fungi can cause serious diseases in humans, several of which may be fatal if untreated. К ним относятся aspergillosis, candidiasis, coccidioidomycosis, cryptococcosis, histoplasmosis, mycetomas, и paracoccidioidomycosis. Furthermore, persons with immuno-deficiencies are particularly susceptible to disease by genera such as Aspergillus, Candida, Cryptoccocus,[165][195][196] Histoplasma,[197] и Pneumocystis.[198] Other fungi can attack eyes, nails, hair, and especially skin, the so-called dermatophytic and keratinophilic fungi, and cause local infections such as ringworm и athlete's foot.[199] Fungal spores are also a cause of аллергия, and fungi from different taxonomic groups can evoke allergic reactions.[200]
As targets of mycoparasites
The organisms which parasitize fungi are known as mycoparasitic organisms. Certain species of the genus Pythium, which are oomycetes, have potential as biocontrol agents against certain fungi.[201] Fungi can also act as mycoparasites or antagonists of other fungi, such as Hypomyces chrysospermus, which grows on болет mushrooms.Fungi can also become the target of infection by mycoviruses.[202][203]
Микотоксины
![(6aR, 9R) -N - ((2R, 5S, 10aS, 10bS) -5-бензил-10b-гидрокси-2-метил-3,6-диоксооктагидро-2H-оксазоло [3,2-a] пирроло [2 , 1-c] пиразин-2-ил) -7-метил-4,6,6a, 7,8,9-гексагидроиндоло [4,3-fg] хинолин-9-карбоксамид](http://upload.wikimedia.org/wikipedia/commons/thumb/9/94/Ergotamine3.png/220px-Ergotamine3.png)
Many fungi produce biologically active compounds, several of which are toxic to animals or plants and are therefore called mycotoxins. Of particular relevance to humans are mycotoxins produced by molds causing food spoilage, and poisonous mushrooms (see above). Particularly infamous are the lethal аматоксины in some Мухомор mushrooms, and алкалоиды спорыньи, which have a long history of causing serious epidemics of эрготизм (St Anthony's Fire) in people consuming rye or related cereals contaminated with sclerotia of the ergot fungus, Claviceps purpurea.[204] Other notable mycotoxins include the aflatoxins, which are insidious liver toxins and highly carcinogenic metabolites produced by certain Aspergillus species often growing in or on grains and nuts consumed by humans, ochratoxins, patulin, и trichothecenes (например., T-2 mycotoxin ) и fumonisins, which have significant impact on human food supplies or animal домашний скот.[205]
Mycotoxins are secondary metabolites (or natural products ), and research has established the existence of biochemical pathways solely for the purpose of producing mycotoxins and other natural products in fungi.[32] Mycotoxins may provide фитнес benefits in terms of physiological adaptation, competition with other microbes and fungi, and protection from consumption (fungivory ).[206][207] Many fungal secondary metabolites (or derivatives) are used medically, as described under Human Use below.
Pathogenic mechanisms
Ustilago maydis is a pathogenic plant fungus that causes smut disease in maize and teosinte. Plants have evolved efficient defense systems against pathogenic microbes such as U. maydis. A rapid defense reaction after pathogen attack is the oxidative burst where the plant produces reactive oxygen species at the site of the attempted invasion. U. maydis can respond to the oxidative burst with an oxidative stress response, regulated by the gene YAP1. The response protects U. maydis from the host defense, and is necessary for the pathogen's virulence.[208] Furthermore, U. maydis has a well-established recombinational Ремонт ДНК system which acts during mitosis and meiosis.[209] The system may assist the pathogen in surviving DNA damage arising from the host plant's oxidative defensive response to infection.[210]
Cryptococcus neoformans is an encapsulated yeast that can live in both plants and animals. С. neoformans usually infects the lungs, where it is phagocytosed by alveolar macrophages.[211] Немного С. neoformans can survive inside macrophages, which appears to be the basis for latency, disseminated disease, and resistance to antifungal agents. One mechanism by which С. neoformans survives the hostile macrophage environment is by up-regulating the expression of genes involved in the oxidative stress response.[211] Another mechanism involves meiosis. Большая часть чего-либо С. neoformans are mating "type a". Filaments of mating "type a" ordinarily have haploid nuclei, but they can become diploid (perhaps by endoduplication or by stimulated nuclear fusion) to form blastospores. The diploid nuclei of blastospores can undergo meiosis, including recombination, to form haploid basidiospores that can be dispersed.[212] This process is referred to as monokaryotic fruiting. This process requires a gene called DMC1, which is a conserved homologue of genes recA in bacteria and RAD51 in eukaryotes, that mediates homologous chromosome pairing during meiosis and repair of DNA double-strand breaks. Таким образом, С. neoformans can undergo a meiosis, monokaryotic fruiting, that promotes recombinational repair in the oxidative, DNA damaging environment of the host macrophage, and the repair capability may contribute to its virulence.[210][212]
Человеческое использование

The human use of fungi for food preparation or preservation and other purposes is extensive and has a long history. Mushroom farming и mushroom gathering are large industries in many countries. The study of the historical uses and sociological impact of fungi is known as ethnomycology. Because of the capacity of this group to produce an enormous range of natural products с antimicrobial or other biological activities, many species have long been used or are being developed for industrial production of antibiotics, vitamins, and anti-cancer и cholesterol-lowering drugs. More recently, methods have been developed for генная инженерия of fungi,[213] enabling metabolic engineering of fungal species. For example, genetic modification of yeast species[214]—which are easy to grow at fast rates in large fermentation vessels—has opened up ways of фармацевтический production that are potentially more efficient than production by the original source organisms.[215]
Therapeutic uses
Modern chemotherapeutics
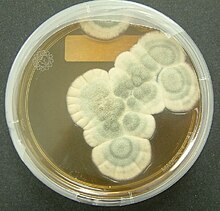
Many species produce metabolites that are major sources of pharmacologically active drugs. Particularly important are the antibiotics, including the penicillins, a structurally related group of β-lactam antibiotics that are synthesized from small peptides. Although naturally occurring penicillins such as penicillin G (produced by Penicillium chrysogenum ) have a relatively narrow spectrum of biological activity, a wide range of other penicillins can be produced by chemical modification of the natural penicillins. Modern penicillins are semisynthetic compounds, obtained initially from ферментация cultures, but then structurally altered for specific desirable properties.[216] Other antibiotics produced by fungi include: ciclosporin, commonly used as an immunosuppressant в течение transplant surgery; и fusidic acid, used to help control infection from methicillin-resistant Staphylococcus aureus бактерии.[217] Widespread use of antibiotics for the treatment of bacterial diseases, such as туберкулез, syphilis, leprosy, and others began in the early 20th century and continues to date. In nature, antibiotics of fungal or bacterial origin appear to play a dual role: at high concentrations they act as chemical defense against competition with other microorganisms in species-rich environments, such as the rhizosphere, and at low concentrations as quorum-sensing molecules for intra- or interspecies signaling.[218] Other drugs produced by fungi include griseofulvin isolated from Penicillium griseofulvum, used to treat fungal infections,[219] и statins (ГМГ-КоА редуктаза inhibitors), used to inhibit cholesterol synthesis. Examples of statins found in fungi include mevastatin из Penicillium citrinum и lovastatin из Aspergillus terreus и oyster mushroom.[220] Fungi produce compounds that inhibit вирусы[221][222] и cancer cells.[223][224] Specific metabolites, such as polysaccharide-K, эрготамин, и β-lactam antibiotics, are routinely used in clinical medicine. В шиитаке mushroom is a source of lentinan, a clinical drug approved for use in cancer treatments in several countries, including Япония.[225][226] В Европа и Япония, polysaccharide-K (brand name Krestin), a chemical derived from Trametes versicolor, is an approved adjuvant for cancer therapy.[227]
Traditional and folk medicine


Certain mushrooms enjoy usage as therapeutics in folk medicines, Такие как Traditional Chinese medicine. Notable medicinal mushrooms with a well-documented history of use include Agaricus subrufescens,[223][228] Ganoderma lucidum,[229] Псилоцибе и Ophiocordyceps sinensis.[230]
Cultured foods
пекарские дрожжи или же Saccharomyces cerevisiae, a unicellular fungus, is used to make bread and other wheat-based products, such as pizza dough and dumplings.[231] Yeast species of the genus Saccharomyces are also used to produce alcoholic beverages through fermentation.[232] Shoyu koji mold (Aspergillus oryzae ) is an essential ingredient in brewing Shoyu (соевый соус ) и sake, and the preparation of мисо,[233] пока Rhizopus species are used for making tempeh.[234] Several of these fungi are domesticated species that were разводится or selected according to their capacity to ferment food without producing harmful mycotoxins (see below), which are produced by very closely related Aspergilli.[235] Quorn, а meat substitute, is made from Fusarium venenatum.[236]
In food

Edible mushrooms include commercially raised and wild-harvested fungi. Agaricus bisporus, sold as button mushrooms when small or Portobello mushrooms when larger, is the most widely cultivated species in the West, used in salads, soups, and many other dishes. Many Asian fungi are commercially grown and have increased in popularity in the West. They are often available fresh in grocery stores and markets, including straw mushrooms (Volvariella volvacea ), oyster mushrooms (Pleurotus ostreatus ), shiitakes (Lentinula edodes ), и enokitake (Flammulina spp.).[237]

Many other mushroom species are harvested from the wild for personal consumption or commercial sale. Milk mushrooms, morels, chanterelles, truffles, black trumpets, и porcini mushrooms (Boletus edulis ) (also known as king boletes) demand a high price on the market. They are often used in gourmet dishes.[238]
Certain types of cheeses require inoculation of milk curds with fungal species that impart a unique flavor and texture to the cheese. Examples include the синий color in cheeses such as Stilton или же Roquefort, which are made by inoculation with Penicillium roqueforti.[239] Molds used in cheese production are non-toxic and are thus safe for human consumption; however, mycotoxins (e.g., aflatoxins, roquefortine C, patulin, or others) may accumulate because of growth of other fungi during cheese ripening or storage.[240]
Poisonous fungi

Many mushroom species are poisonous to humans and cause a range of reactions including slight digestive problems, allergic reactions, галлюцинации, severe organ failure, and death. Genera with mushrooms containing deadly toxins include Conocybe, Галерина, Lepiota, and, the most infamous, Мухомор.[241] The latter genus includes the destroying angel (А. вироса ) and the death cap (А. фаллоиды ), the most common cause of deadly mushroom poisoning.[242] The false morel (Gyromitra esculenta ) is occasionally considered a delicacy when cooked, yet can be highly toxic when eaten raw.[243] Tricholoma equestre was considered edible until it was implicated in serious poisonings causing rhabdomyolysis.[244] Fly agaric mushrooms (Мухомор мухомор) also cause occasional non-fatal poisonings, mostly as a result of ingestion for its hallucinogenic характеристики. Historically, fly agaric was used by different peoples in Europe and Asia and its present usage for religious or shamanic purposes is reported from some ethnic groups such as the Koryak people of northeastern Сибирь.[245]
As it is difficult to accurately identify a safe mushroom without proper training and knowledge, it is often advised to assume that a wild mushroom is poisonous and not to consume it.[246][247]
Pest control

In agriculture, fungi may be useful if they actively compete for nutrients and space with патогенный microorganisms such as bacteria or other fungi via the competitive exclusion principle,[248] or if they are паразиты of these pathogens. For example, certain species may be used to eliminate or suppress the growth of harmful plant pathogens, such as insects, клещи, weeds, нематоды, and other fungi that cause diseases of important crop plants.[249] This has generated strong interest in practical applications that use these fungi in the biological control of these agricultural pests. Entomopathogenic fungi can be used as biopesticides, as they actively kill insects.[250] Examples that have been used as biological insecticides находятся Beauveria bassiana, Metarhizium spp, Hirsutella spp, Paecilomyces (Isaria) spp, and Lecanicillium lecanii.[251][252] Endophytic fungi of grasses of the genus Неотифодиум, Такие как N. coenophialum, produce alkaloids that are toxic to a range of invertebrate and vertebrate травоядные животные. These alkaloids protect grass plants from травоядный, but several endophyte alkaloids can poison grazing animals, such as cattle and sheep.[253] Infecting cultivars of pasture или же корм grasses with Неотифодиум endophytes is one approach being used in grass breeding programs; the fungal strains are selected for producing only alkaloids that increase resistance to herbivores such as insects, while being non-toxic to livestock.[254][255]
Bioremediation
Certain fungi, in particular white-rot fungi, can degrade insecticides, herbicides, pentachlorophenol, creosote, coal tars, and heavy fuels and turn them into carbon dioxide, water, and basic elements.[256] Fungi have been shown to biomineralize уран oxides, suggesting they may have application in the bioremediation of radioactively polluted sites.[257][258][259]
Модельные организмы
Several pivotal discoveries in biology were made by researchers using fungi as model organisms, that is, fungi that grow and sexually reproduce rapidly in the laboratory. Например, one gene-one enzyme hypothesis was formulated by scientists using the bread mold Neurospora crassa to test their biochemical theories.[260] Other important model fungi are Aspergillus nidulans and the yeasts Saccharomyces cerevisiae и Schizosaccharomyces pombe, each of which with a long history of use to investigate issues in eukaryotic cell biology и genetics, Такие как cell cycle regulation, chromatin structure, and gene regulation. Other fungal models have more recently emerged that address specific biological questions relevant to medicine, plant pathology, and industrial uses; examples include Candida albicans, a dimorphic, opportunistic human pathogen,[261] Magnaporthe grisea, a plant pathogen,[262] и Pichia pastoris, a yeast widely used for eukaryotic protein production.[263]
Другие
Fungi are used extensively to produce industrial chemicals like лимонный, gluconic, lactic, и malic acids,[264] и industrial enzymes, Такие как lipases используется в biological detergents,[265] cellulases used in making cellulosic ethanol[266] и stonewashed jeans,[267] и amylases,[268] invertases, proteases и xylanases.[269]
Смотрите также
Рекомендации
- ^ Moore RT (1980). "Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts". Botanica Marina. 23: 361–373.
- ^ /ˈжʌпdʒаɪ/ (
 Слушать), /ˈжʌŋɡаɪ/ (Слушать), /ˈжʌŋɡя/ (Слушать) или же /ˈжʌпdʒя/ (Слушать). Первые два произношения предпочитаются больше в США, а другие - в Великобритании, однако любое произношение можно услышать в любой англоязычной стране.
Слушать), /ˈжʌŋɡаɪ/ (Слушать), /ˈжʌŋɡя/ (Слушать) или же /ˈжʌпdʒя/ (Слушать). Первые два произношения предпочитаются больше в США, а другие - в Великобритании, однако любое произношение можно услышать в любой англоязычной стране. - ^ «Грибок». Оксфордские словари. В архиве из оригинала 28 июля 2012 г.. Получено 26 февраля 2011.
- ^ а б Хоксворт Д.Л., Люкинг Р. (июль 2017 г.). «Пересмотренное разнообразие грибов: от 2,2 до 3,8 миллиона видов». Грибковое королевство. Микробиологический спектр. 5. С. 79–95. Дои:10.1128 / microbiolspec.FUNK-0052-2016. ISBN 978-1-55581-957-6. PMID 28752818.
- ^ «Хватит пренебрегать грибами». Природная микробиология. 2 (8): 17120.25 июля 2017 г. Дои:10.1038 / nmicrobiol.2017.120. PMID 28741610.
- ^ Симпсон Д.П. (1979). Латинский словарь Кассела (5-е изд.). Лондон, Великобритания: Cassell Ltd., стр. 883. ISBN 978-0-304-52257-6.
- ^ а б Эйнсворт, стр. 2.
- ^ Мицка В., изд. (1960). Etymologisches Wörterbuch der deutschen Sprache. Берлин: Вальтер де Грюйтер.
- ^ Алексопулос и другие., п. 1.
- ^ Персона, Христиан Хендрик (1796). Observationes Mycologicae (на латыни). Часть 1. Лейпциг (Германия): Питер Филипп Вольф. В архиве из оригинала 19 декабря 2013 г.. Получено 30 марта 2019.
- ^ Гревиль, Роберт Кэй (1824). Шотландская криптогамная флора: цветные рисунки и описания криптогамных растений, принадлежащих главным образом к отряду грибов. т. 2. Эдинбург, Шотландия: Маклахленд и Стюарт. п. 65. С п. 65: «Это маленькое растение, вероятно, не станет редкостью в Великобритании, когда микология будет более изучена».
- ^ Смит, Джеймс Эдвард; Хукер, Уильям Джексон, изд. (1836 г.). Беркли, Майлз Джозеф (ред.). Английская Флора сэра Джеймса Эдварда Смита. т. 5, часть II: «Класс XXIV. Криптогамия». Лондон, Англия: Longman, Rees, Orme, Brown, Green & Longman. п. 7.CS1 maint: дополнительный текст: список авторов (связь) С п. 7: «Это возникло, как я понимаю, отчасти из-за практических трудностей сохранения образцов для гербария, отчасти из-за отсутствия какой-либо общей работы, адаптированной к огромным успехам, достигнутым в последние годы в изучении микологии».
- ^ «Глоссарий LIAS». В архиве из оригинала 11 декабря 2013 г.. Получено 14 августа 2013.
- ^ Брунс Т. (октябрь 2006 г.). «Эволюционная биология: новое царство». Природа. 443 (7113): 758–61. Bibcode:2006Натура.443..758Б. Дои:10.1038 / 443758a. PMID 17051197. S2CID 648881.
- ^ Baldauf SL, Palmer JD (декабрь 1993 г.). «Животные и грибы являются ближайшими родственниками друг друга: совпадающие доказательства из множества белков». Труды Национальной академии наук Соединенных Штатов Америки. 90 (24): 11558–62. Bibcode:1993ПНАС ... 9011558Б. Дои:10.1073 / пнас.90.24.11558. ЧВК 48023. PMID 8265589.
- ^ Дьякон, стр. 4.
- ^ а б Дьякон, стр. 128–129.
- ^ Алексопулос и другие., стр. 28–33.
- ^ Алексопулос и другие., стр. 31–32.
- ^ Сёдзи Дж.Й., Ариока М., Китамото К. (2006). «Возможное участие плейоморфных вакуолярных сетей в рециркуляции питательных веществ в мицелиальных грибах». Аутофагия. 2 (3): 226–7. Дои:10.4161 / авто.2695. PMID 16874107.
- ^ Дьякон, стр. 58.
- ^ Забриски TM, Джексон MD (февраль 2000 г.). «Биосинтез и метаболизм лизина в грибах». Отчеты о натуральных продуктах. 17 (1): 85–97. Дои:10.1039 / a801345d. PMID 10714900.
- ^ Сюй Х, Анди Б., Цянь Дж., Западный А. Х., Кук П. Ф. (2006). «Альфа-аминоадипатный путь биосинтеза лизина в грибах». Биохимия клетки и биофизика. 46 (1): 43–64. Дои:10.1385 / CBB: 46: 1: 43. PMID 16943623. S2CID 22370361.
- ^ Алексопулос и другие., стр. 27–28.
- ^ Алексопулос и другие., п. 685.
- ^ а б c Алексопулос и другие., п. 30.
- ^ Десжарден Д.Е., Перри Б.А., Лодж Диджей, Стевани CV, Нагасава Э. (2010). «Люминесцентный микен: новые и примечательные виды». Микология. 102 (2): 459–77. Дои:10.3852/09-197. PMID 20361513. S2CID 25377671. В архиве из оригинала 11 ноября 2018 г.. Получено 11 ноября 2018.
- ^ Алексопулос и другие., стр. 32–33.
- ^ Bowman SM, Free SJ (август 2006 г.). «Строение и синтез клеточной стенки грибов». BioEssays. 28 (8): 799–808. Дои:10.1002 / bies.20441. PMID 16927300. S2CID 22623524.
- ^ Алексопулос и другие., п. 33.
- ^ Михаил JD, Брюн JN (ноябрь 2005 г.). «Кормление систем ризоморф Armillaria». Микологические исследования. 109 (Pt 11): 1195–207. Дои:10.1017 / S0953756205003606. PMID 16279413.
- ^ а б c d Келлер Н.П., Тернер Дж., Беннетт Дж. В. (декабрь 2005 г.). «Вторичный метаболизм грибов - от биохимии к геномике». Обзоры природы. Микробиология. 3 (12): 937–47. Дои:10.1038 / nrmicro1286. PMID 16322742. S2CID 23537608.
- ^ Ву С., Шалк М., Кларк А., Майлз Р. Б., Коутс Р., Чаппелл Дж. (Ноябрь 2006 г.). «Перенаправление цитозольных или пластидных предшественников изопреноидов увеличивает продукцию терпена в растениях». Природа Биотехнологии. 24 (11): 1441–7. Дои:10.1038 / nbt1251. PMID 17057703. S2CID 23358348.
- ^ Тудзинский Б. (март 2005 г.). «Биосинтез гиббереллина в грибах: гены, ферменты, эволюция и влияние на биотехнологию». Прикладная микробиология и биотехнология. 66 (6): 597–611. Дои:10.1007 / s00253-004-1805-1. PMID 15578178. S2CID 11191347.
- ^ Vaupotic T, Veranic P, Jenoe P, Plemenitas A (июнь 2008 г.). «Митохондриальное посредничество дискриминации осмолитов окружающей среды во время осмоадаптации в чрезвычайно галотолерантных черных дрожжах Hortaea werneckii». Грибковая генетика и биология. 45 (6): 994–1007. Дои:10.1016 / j.fgb.2008.01.006. PMID 18343697.
- ^ а б Дадачева Э., Брайан Р.А., Хуанг Х, Моадель Т., Швейцер А.Д., Айзен П., Носанчук Дж.Д., Касадеваль А (2007). «Ионизирующее излучение изменяет электронные свойства меланина и усиливает рост меланизированных грибов». PLOS ONE. 2 (5): e457. Bibcode:2007PLoSO ... 2..457D. Дои:10.1371 / journal.pone.0000457. ЧВК 1866175. PMID 17520016.
- ^ Рагхукумар С., Рагхукумар С. (1998). «Баротолерантность грибов, выделенных из глубоководных отложений Индийского океана». Экология водных микробов. 15 (2): 153–163. Дои:10.3354 / ame015153.
- ^ Санчо Л.Г., де ла Торре Р., Хорнек Г., Аскасо С., де Лос Риос А., Пинтадо А., Вежчос Дж., Шустер М. (июнь 2007 г.). «Лишайники выживают в космосе: результаты эксперимента LICHENS 2005 г.». Астробиология. 7 (3): 443–54. Bibcode:2007AsBio ... 7..443S. Дои:10.1089 / аст.2006.0046. PMID 17630840. S2CID 4121180.
- ^ Brem FM, Lips KR (сентябрь 2008 г.). «Паттерны заражения Batrachochytrium dendrobatidis среди видов панамских амфибий, среды обитания и возвышенности во время эпизоотической и энзоотической стадий». Болезни водных организмов. 81 (3): 189–202. Дои:10.3354 / dao01960. PMID 18998584.
- ^ Ле Кальвез Т., Бурго Дж., Маэ С., Барбье Дж., Ванденкоорнхейз П. (октябрь 2009 г.). «Разнообразие грибов в глубоководных гидротермальных экосистемах». Прикладная и экологическая микробиология. 75 (20): 6415–21. Дои:10.1128 / AEM.00653-09. ЧВК 2765129. PMID 19633124.
- ^ а б Мюллер GM, Шмит JP (2006). «Биоразнообразие грибов: что мы знаем? Что мы можем предсказать?». Биоразнообразие и сохранение. 16: 1–5. Дои:10.1007 / s10531-006-9117-7. S2CID 23827807.
- ^ а б Кирк и другие., п. 489.
- ^ а б c d е ж грамм час Хиббетт Д.С., Биндер М., Бишофф Дж. Ф., Блэквелл М., Кэннон П. Ф., Эрикссон О. Э. и др. (Май 2007 г.). «Филогенетическая классификация грибов более высокого уровня» (PDF). Микологические исследования. 111 (Pt 5): 509–47. CiteSeerX 10.1.1.626.9582. Дои:10.1016 / j.mycres.2007.03.004. PMID 17572334. Архивировано из оригинал (PDF) 26 марта 2009 г.
- ^ По оценке 2001 года, известно около 10 000 грибковых заболеваний. Ударил C (2006). «Стратегии заражения растительными паразитарными грибами». В Cooke BM, Jones DG, Kaye B (ред.). Эпидемиология болезней растений. Берлин, Германия: Springer. п. 117. ISBN 978-1-4020-4580-6.
- ^ Пейнтнер У., Пёдер Р., Пюмпель Т. (1998). «Грибы ледяного человека». Микологические исследования. 102 (10): 1153–1162. Дои:10.1017 / S0953756298006546.
- ^ Эйнсворт, стр. 1.
- ^ Алексопулос и другие., стр. 1–2.
- ^ Эйнсворт, стр. 18.
- ^ Хоксворт DL (сентябрь 2006 г.). «Микологический ящик Пандоры: молекулярные последовательности против морфологии в понимании взаимоотношений грибов и биоразнообразия». Revista Iberoamericana de Micología. 23 (3): 127–33. Дои:10.1016 / S1130-1406 (06) 70031-6. PMID 17196017.
- ^ Харрис С.Д. (2008). «Ветвление гиф грибов: регуляция, механизмы и сравнение с другими системами ветвления». Микология. 100 (6): 823–32. Дои:10.3852/08-177. PMID 19202837. S2CID 2147525. В архиве из оригинала 12 апреля 2016 г.. Получено 5 июля 2011.
- ^ Дьякон, стр. 51.
- ^ Дьякон, стр. 57.
- ^ Чанг С.Т., Майлз П.Г. (2004). Грибы: выращивание, пищевая ценность, лечебное действие и воздействие на окружающую среду. Бока-Ратон, Флорида: CRC Press. ISBN 978-0-8493-1043-0.
- ^ Парниске М. (октябрь 2008 г.). «Арбускулярная микориза: мать корневых эндосимбиозов растений». Обзоры природы. Микробиология. 6 (10): 763–75. Дои:10.1038 / nrmicro1987. PMID 18794914. S2CID 5432120.
- ^ Стинкамп Е.Т., Райт Дж., Балдауф С.Л. (январь 2006 г.). «Протестированное происхождение животных и грибов». Молекулярная биология и эволюция. 23 (1): 93–106. Дои:10.1093 / molbev / msj011. PMID 16151185.
- ^ Стивенс Д.А., Ичиномия М., Коси Ю., Хориучи Х. (сентябрь 2006 г.). «Избавление Candida от ингибирования каспофунгина при концентрациях выше МПК (парадоксальный эффект), достигаемое за счет увеличения хитина клеточной стенки; доказательства ингибирования синтеза бета-1,6-глюкана каспофунгином». Противомикробные препараты и химиотерапия. 50 (9): 3160–1. Дои:10.1128 / AAC.00563-06. ЧВК 1563524. PMID 16940118.
- ^ Хэнсон, стр. 127–141.
- ^ Фергюсон Б.А., Драйсбах Т.А., Паркс К.Г., Филип Г.М., Шмитт К.Л. (2003). «Примерная популяционная структура патогенных Армиллярия вид в смешанном хвойном лесу в Голубых горах на северо-востоке Орегона ". Канадский журнал исследований леса. 33 (4): 612–623. Дои:10.1139 / x03-065. В архиве из оригинала на 3 июля 2019 г.. Получено 3 июля 2019.
- ^ Алексопулос и другие., стр. 204–205.
- ^ Мосс СТ (1986). Биология морских грибов. Кембридж, Великобритания: Издательство Кембриджского университета. п. 76. ISBN 978-0-521-30899-1.
- ^ Пеньяльва М.А., Арст Х.Н. (сентябрь 2002 г.). «Регулирование экспрессии генов с помощью pH окружающей среды в мицелиальных грибах и дрожжах». Обзоры микробиологии и молекулярной биологии. 66 (3): 426–46, содержание. Дои:10.1128 / MMBR.66.3.426-446.2002. ЧВК 120796. PMID 12208998.
- ^ а б Говард Р.Дж., Феррари М.А., Роуч Д.Х., Деньги Н.П. (декабрь 1991 г.). «Проникновение грибка в твердые субстраты с огромным тургорным давлением». Труды Национальной академии наук Соединенных Штатов Америки. 88 (24): 11281–4. Bibcode:1991PNAS ... 8811281H. Дои:10.1073 / пнас.88.24.11281. ЧВК 53118. PMID 1837147.
- ^ Деньги НП (1998). «Механика инвазивного роста грибов и значение тургора при заражении растений». Молекулярная генетика специфических для хозяев токсинов при болезнях растений: материалы 3-го Международного симпозиума Тоттори по специфическим для хозяина токсинам, Дайсен, Тоттори, Япония, 24–29 августа 1997 г.. Нидерланды: Kluwer Academic Publishers. С. 261–271. ISBN 978-0-7923-4981-5.
- ^ Ван З.Й., Дженкинсон Дж. М., Холкомб Л. Дж., Соанес Д. М., Вено-Фурри С., Бхамбра Г. К., Талбот, штат Нью-Джерси (апрель 2005 г.). «Молекулярная биология генерации тургора appressorium грибком Magnaporthe grisea». Сделки Биохимического Общества. 33 (Pt 2): 384–8. Дои:10.1042 / BST0330384. PMID 15787612. S2CID 7111935.
- ^ Перейра Дж. Л., Норонья Е. Ф., Миллер Р. Н., Франко О. Л. (июнь 2007 г.). «Новые идеи в использовании гидролитических ферментов, секретируемых грибами с биотехнологическим потенциалом». Письма по прикладной микробиологии. 44 (6): 573–81. Дои:10.1111 / j.1472-765X.2007.02151.x. PMID 17576216.
- ^ Шаллер М., Борелли С., Кортинг Х.С., Хубе Б. (ноябрь 2005 г.). «Гидролитические ферменты как факторы вирулентности Candida albicans». Микозы. 48 (6): 365–77. Дои:10.1111 / j.1439-0507.2005.01165.x. PMID 16262871. S2CID 1356254.
- ^ Фаррар Дж. Ф. (октябрь 1985 г.). «Углеводный обмен в патогенах биотрофных растений». Микробиологические науки. 2 (10): 314–7. PMID 3939987.
- ^ Фишер Р., Зекерт Н., Такешита Н. (май 2008 г.). «Поляризованный рост грибов - взаимодействие между цитоскелетом, позиционными маркерами и мембранными доменами». Молекулярная микробиология. 68 (4): 813–26. Дои:10.1111 / j.1365-2958.2008.06193.x. PMID 18399939. S2CID 205365895.
- ^ Кристенсен М.Дж., Беннетт Р.Дж., Ансари Н.А., Кога Н.Д., Джонсон Р.Д., Брайан Г.Т., Симпсон В.Р., Кулаард Дж. П., Никлесс Э.М., Voisey CR (февраль 2008 г.). «Эндофиты эпихлои растут за счет удлинения вставочного гифа в удлиненных листьях травы». Грибковая генетика и биология. 45 (2): 84–93. Дои:10.1016 / j.fgb.2007.07.013. PMID 17919950.
- ^ Деньги НП (октябрь 2002 г.). «Стволовые клетки грибов». BioEssays. 24 (10): 949–52. Дои:10.1002 / bies.10160. PMID 12325127.
- ^ Вилленсдорфер М (февраль 2009 г.). «Об эволюции дифференцированной многоклеточности». Эволюция; Международный журнал органической эволюции. 63 (2): 306–23. arXiv:0801.2610. Дои:10.1111 / j.1558-5646.2008.00541.x. PMID 19154376. S2CID 39155292.
- ^ Дэниэлс К.Дж., Сриканта Т., Локхарт С.Р., Пухоль С., Солл Д.Р. (май 2006 г.). "Непрозрачные клетки сигнализируют лейкоцитам о формировании биопленок в грибковые микроорганизмы албиканс". Журнал EMBO. 25 (10): 2240–52. Дои:10.1038 / sj.emboj.7601099. ЧВК 1462973. PMID 16628217.
- ^ Марзлуф Г.А. (сентябрь 1981 г.). «Регуляция азотистого обмена и экспрессии генов у грибов». Микробиологические обзоры. 45 (3): 437–61. Дои:10.1128 / MMBR.45.3.437-461.1981. ЧВК 281519. PMID 6117784.
- ^ Хайнс MJ (1994). "Регуляторные цепи гена amdS Aspergillus nidulans". Антони ван Левенгук. 65 (3): 179–82. Дои:10.1007 / BF00871944. PMID 7847883. S2CID 45815733.
- ^ Дадачева Е., Casadevall A (декабрь 2008 г.). «Ионизирующее излучение: как грибы справляются, адаптируются и эксплуатируются с помощью меланина». Текущее мнение в микробиологии. 11 (6): 525–31. Дои:10.1016 / j.mib.2008.09.013. ЧВК 2677413. PMID 18848901.
- ^ Алексопулос и другие., стр. 48–56.
- ^ Кирк и другие., п. 633.
- ^ Хейтман Дж (сентябрь 2006 г.). «Половое размножение и эволюция микробных патогенов». Текущая биология. 16 (17): R711–25. Дои:10.1016 / j.cub.2006.07.064. PMID 16950098. S2CID 2898102.
- ^ Алькамо И.Е., Поммервилл Дж. (2004). Основы микробиологии Алькамо. Бостон, Массачусетс: Джонс и Бартлетт. п.590. ISBN 978-0-7637-0067-6.
- ^ а б Редекер Д., Рааб П. (2006). «Филогения glomeromycota (арбускулярные микоризные грибы): последние разработки и новые генные маркеры». Микология. 98 (6): 885–95. Дои:10.3852 / mycologia.98.6.885. PMID 17486965. В архиве из оригинала 23 сентября 2015 г.. Получено 5 июля 2011.
- ^ Гуарро Дж., Счигель А.М. (июль 1999 г.). «Развитие грибковой систематики». Обзоры клинической микробиологии. 12 (3): 454–500. Дои:10.1128 / CMR.12.3.454. ЧВК 100249. PMID 10398676.
- ^ а б Тейлор Дж. В., Джейкобсон Д. Д., Крокен С., Касуга Т., Гейзер Д. М., Хиббетт Д. С., Фишер М.С. (октябрь 2000 г.). «Филогенетическое распознавание видов и видовые концепции грибов». Грибковая генетика и биология. 31 (1): 21–32. Дои:10.1006 / fgbi.2000.1228. PMID 11118132. S2CID 2551424.
- ^ Метценберг Р.Л., Гласс Н.Л. (февраль 1990 г.). «Тип спаривания и стратегии спаривания у Neurospora». BioEssays. 12 (2): 53–9. Дои:10.1002 / bies.950120202. PMID 2140508. S2CID 10818930.
- ^ Дженнингс и Лизек, стр. 107–114.
- ^ Дьякон, стр. 31.
- ^ Алексопулос и другие., стр. 492–493.
- ^ Дженнингс и Лизек, стр. 142.
- ^ Дьякон, стр. 21–24.
- ^ Линдер МБ, Сзилвай Г.Р., Накари-Сетяля Т., Пенттиля М.Э. (ноябрь 2005 г.). «Гидрофобины: белки-амфифилы мицелиальных грибов». Обзор микробиологии FEMS. 29 (5): 877–96. Дои:10.1016 / j.femsre.2005.01.004. PMID 16219510.
- ^ След F (ноябрь 2007 г.). «Грибковые пушки: взрывные разряды спор в Ascomycota». Письма о микробиологии FEMS. 276 (1): 12–8. Дои:10.1111 / j.1574-6968.2007.00900.x. PMID 17784861.
- ^ Прингл А., Патек С. Н., Фишер М., Штольце Дж., Деньги Н. П. (2005). «Захваченный запуск баллистоспоры». Микология. 97 (4): 866–71. Дои:10.3852 / mycologia.97.4.866. PMID 16457355. В архиве из оригинала 12 апреля 2016 г.. Получено 5 июля 2011.
- ^ Кирк и другие., п. 495.
- ^ «Стимулирующие гидноидные грибы, Хэмпширское партнерство по биоразнообразию» (PDF). В архиве (PDF) из оригинала 4 марта 2016 г.. Получено 13 ноября 2019.
- ^ Броди HJ (1975). Грибы птичьего гнезда. Торонто, Онтарио: Университет Торонто Пресс. п. 80. ISBN 978-0-8020-5307-7.
- ^ Алексопулос и другие., п. 545.
- ^ а б "Распространение спор в грибах". www.botany.hawaii.edu. В архиве из оригинала 17 ноября 2011 г.. Получено 28 декабря 2018.
- ^ «Разгон». herbarium.usu.edu. В архиве с оригинала 28 декабря 2018 г.. Получено 28 декабря 2018.
- ^ Hassett, Maribeth O .; Фишер, Марк В. Ф .; Деньги, Николай П. (28 октября 2015 г.). «Грибы как создатели дождя: как споры действуют как ядра для капель дождя». PLOS ONE. 10 (10): e0140407. Bibcode:2015PLoSO..1040407H. Дои:10.1371 / journal.pone.0140407. ISSN 1932-6203. ЧВК 4624964. PMID 26509436.
- ^ Ким, Сынхо; Пак, Хёнгон; Gruszewski, Hope A .; Шмале, Дэвид Дж .; Юнг, Сунгван (12 марта 2019 г.). «Вызванное вихрем распространение патогена растений под воздействием капель дождя». Труды Национальной академии наук. 116 (11): 4917–4922. Bibcode:2019PNAS..116.4917K. Дои:10.1073 / pnas.1820318116. ISSN 0027-8424. ЧВК 6421443. PMID 30804195.
- ^ Дайер П.С., О'Горман К.М. (январь 2012 г.). «Половое развитие и загадочная сексуальность у грибов: выводы из видов Aspergillus». FEMS Microbiol. Rev. 36 (1): 165–92. Дои:10.1111 / j.1574-6976.2011.00308.x. PMID 22091779.
- ^ Юн Ш., Берби М.Л., Йодер О.К., Тургеон Б.Г. (1999). «Эволюция самоплодотворяющего репродуктивного образа жизни грибов от самоплодородных предков». Proc Natl Acad Sci U S A. 96 (10): 5592–7. Bibcode:1999PNAS ... 96.5592Y. Дои:10.1073 / пнас.96.10.5592. ЧВК 21905. PMID 10318929.CS1 maint: несколько имен: список авторов (связь)
- ^ Ричард С., Almeida Jmgcf CO, Luraschi A, Nielsen O, Pagni M, Hauser PM (2018). «Функциональный анализ и анализ экспрессии генов MAT Pneumocystis предполагают обязательную сексуальность через первичный гомоталлизм в легких хозяина». мБио. 9 (1). Дои:10,1128 / мБио.02201-17. ЧВК 5821091. PMID 29463658.CS1 maint: несколько имен: список авторов (связь)
- ^ Хейтман Дж. (Декабрь 2015 г.). «Эволюция полового размножения: взгляд из Грибного царства поддерживает эволюционную эпоху с полом до пола». Грибной Биол Рев. 29 (3–4): 108–117. Дои:10.1016 / j.fbr.2015.08.002. ЧВК 4730888. PMID 26834823.
- ^ Дженнингс и Лизек, стр. 114–115.
- ^ Furlaneto MC, Pizzirani-Kleiner AA (январь 1992 г.). «Внутривидовая гибридизация Trichoderma pseudokoningii путем анастомоза и слияния протопластов». Письма о микробиологии FEMS. 69 (2): 191–5. Дои:10.1111 / j.1574-6968.1992.tb05150.x. PMID 1537549.
- ^ Schardl CL, Craven KD (ноябрь 2003 г.). «Межвидовая гибридизация у ассоциированных с растениями грибов и оомицетов: обзор». Молекулярная экология. 12 (11): 2861–73. Дои:10.1046 / j.1365-294X.2003.01965.x. PMID 14629368. S2CID 25879264.
- ^ Донохью MJ; Крафт J (2004). Сборка Древа Жизни. Оксфорд (Оксфордшир), Великобритания: Oxford University Press. п. 187. ISBN 978-0-19-517234-8.
- ^ Тейлор и Тейлор, стр. 19.
- ^ Тейлор и Тейлор, стр. 7–12.
- ^ Бенгтсон, Стефан; Расмуссен, Биргер; Иварссон, Магнус; Мюлинг, Джанет; Броман, Курт; Мароне, Федерика; Стампанони, Марко; Беккер, Андрей (24 апреля 2017 г.). «Грибоподобные окаменелости мицелия в везикулярном базальте возрастом 2,4 миллиарда лет». Природа Экология и эволюция. 1 (6): 0141. Дои:10.1038 / s41559-017-0141. PMID 28812648. S2CID 25586788. В архиве с оригинала 15 июля 2019 г.. Получено 15 июля 2019.
- ^ Lücking R, Huhndorf S, Pfister DH, Plata ER, Lumbsch HT (2009). «Грибки эволюционировали прямо на ходу». Микология. 101 (6): 810–22. Дои:10.3852/09-016. PMID 19927746. S2CID 6689439.
- ^ а б c Джеймс Т.Ю., Кауфф Ф., Шох К.Л., Матени П.Б., Хофстеттер В., Кокс С.Дж. и др. (Октябрь 2006 г.). «Реконструкция ранней эволюции грибов с использованием филогении с шестью генами». Природа. 443 (7113): 818–22. Bibcode:2006Натура.443..818J. Дои:10.1038 / природа05110. PMID 17051209. S2CID 4302864.
- ^ Тейлор и Тейлор, стр. 84–94 и 106–107.
- ^ Schoch CL, Sung GH, López-Giráldez F, Townsend JP, Miadlikowska J, Hofstetter V, et al. (Апрель 2009 г.). «Древо жизни Ascomycota: филогенетическая филогения проясняет происхождение и эволюцию основных репродуктивных и экологических черт». Систематическая биология. 58 (2): 224–39. Дои:10.1093 / sysbio / syp020. PMID 20525580.
- ^ Циммер, Карл (22 мая 2019 г.). «Как жизнь появилась на суше? Гриб, которому миллиард лет, может содержать ключи - кладезь микроскопических окаменелостей из Арктики указывает на то, что грибы достигли суши задолго до растений». Нью-Йорк Таймс. В архиве с оригинала 23 мая 2019 г.. Получено 23 мая 2019.
- ^ Loron, Corentin C .; Франсуа, Камилла; Rainbird, Роберт Х .; Тернер, Элизабет С .; Боренштайн, Стефан; Хаво, Эммануэль Дж. (22 мая 2019 г.). «Ранние грибы протерозойской эры в арктической Канаде». Природа. ООО "Спрингер Сайенс энд Бизнес Медиа". 570 (7760): 232–235. Bibcode:2019Натуральная 570..232Л. Дои:10.1038 / s41586-019-1217-0. ISSN 0028-0836. PMID 31118507. S2CID 162180486.
- ^ Тиммер, Джон (22 мая 2019 г.). «Окаменелости возрастом в миллиард лет могут быть ранним грибком». Ars Technica. В архиве с оригинала 23 мая 2019 г.. Получено 23 мая 2019.
- ^ а б Брандретт MC (2002). «Коэволюция корней и микориз наземных растений». Новый Фитолог. 154 (2): 275–304. Дои:10.1046 / j.1469-8137.2002.00397.x.
- ^ Редекер Д., Коднер Р., Грэм Л. Е. (сентябрь 2000 г.). «Гломалийские грибы ордовика». Наука. 289 (5486): 1920–1. Bibcode:2000Sci ... 289.1920R. Дои:10.1126 / science.289.5486.1920. PMID 10988069. S2CID 43553633.
- ^ Тейлор Т.Н., Тейлор Э.Л. (1996). «Распространение и взаимодействие некоторых палеозойских грибов». Обзор палеоботаники и палинологии. 95 (1–4): 83–94. Дои:10.1016 / S0034-6667 (96) 00029-2.
- ^ Доцлер Н., Уокер К., Крингс М., Хасс Х, Керп Х, Тейлор Т. Н., Агерер Р. (2009). «Акаулоспороидные споры гломеромикотана с защитой от прорастания из Rhynie chert возрастом 400 миллионов лет» (PDF). Микологический прогресс. 8 (1): 9–18. Дои:10.1007 / s11557-008-0573-1. HDL:1808/13680. S2CID 1746303.
- ^ Тейлор JW, Берби ML (2006). «Расхождения в датах на грибном древе жизни: обзор и новый анализ». Микология. 98 (6): 838–49. Дои:10.3852 / mycologia.98.6.838. PMID 17486961. В архиве из оригинала 12 апреля 2016 г.. Получено 5 июля 2011.
- ^ Блэквелл М., Вилгалис Р., Джеймс Т.Ю., Тейлор Дж. В. (2009). «Грибы. Eumycota: грибы, мешочные грибки, дрожжи, плесень, ржавчина, головня и т. Д.». Веб-проект «Древо жизни». В архиве из оригинала 13 апреля 2009 г.. Получено 25 апреля 2009.
- ^ Юань X, Сяо С., Тейлор Т.Н. (май 2005 г.). «Симбиоз типа лишайников 600 миллионов лет назад». Наука. 308 (5724): 1017–20. Bibcode:2005Наука ... 308.1017Y. Дои:10.1126 / science.1111347. PMID 15890881. S2CID 27083645.
- ^ Каратыгин И.В., Снигиревская Н.С., Викулин С.В. (2009). «Древнейший наземный лишайник Winfrenatia reticulata: Новая находка и новая интерпретация ». Палеонтологический журнал. 43 (1): 107–114. Дои:10.1134 / S0031030109010110. S2CID 85262818.
- ^ Тейлор Т. Н., Хасс Х, Керп Х, Крингс М., Хэнлин Р. Т. (2005). «Перитециальные аскомицеты из Rhynie chert возрастом 400 миллионов лет: пример предкового полиморфизма». Микология. 97 (1): 269–85. Дои:10.3852 / mycologia.97.1.269. HDL:1808/16786. PMID 16389979. В архиве из оригинала 12 апреля 2016 г.. Получено 5 июля 2011.
- ^ Деннис Р.Л. (1970). «Мицелий базидиомицета Среднего Пенсильвании с зажимными соединениями». Микология. 62 (3): 578–584. Дои:10.2307/3757529. JSTOR 3757529. В архиве из оригинала 29 сентября 2018 г.. Получено 5 июля 2011.
- ^ Хиббетт Д.С., Гримальди Д., Донохью М.Дж. (1995). «Меловые грибы в янтаре». Природа. 377 (6549): 487. Bibcode:1995Натура.377..487H. Дои:10.1038 / 377487a0. S2CID 4346359.
- ^ Хиббетт Д.С., Гримальди Д., Донохью М.Дж. (1997). «Ископаемые грибы из миоценового и мелового янтаря и эволюция гомобазидиомицетов». Американский журнал ботаники. 84 (7): 981–991. Дои:10.2307/2446289. JSTOR 2446289. PMID 21708653. S2CID 22011469.
- ^ Эшет Y, Рампино MR, Visscher H (1995). «Грибковые явления и палинологическая запись экологического кризиса и восстановления на границе перми и триаса». Геология. 23 (1): 967–970. Bibcode:1995Гео .... 23..967E. Дои:10.1130 / 0091-7613 (1995) 023 <0967: FEAPRO> 2.3.CO; 2. S2CID 58937537.
- ^ Фостер CB, Стивенсон MH, Маршалл C, Логан GA, Greenwood PF (2002). «Пересмотр Reduviasporonites Wilson 1962: описание, иллюстрации, сравнение и биологическое родство». Палинология. 26 (1): 35–58. Дои:10.2113/0260035.
- ^ Лопес-Гомес Дж, Тейлор Э.Л. (2005). «Пермско-триасовый переход в Испании: мультидисциплинарный подход». Палеогеография, палеоклиматология, палеоэкология. 229 (1–2): 1–2. Дои:10.1016 / j.palaeo.2005.06.028.
- ^ Looy CV, Twitchett RJ, Dilcher DL, Van Konijnenburg-Van Cittert JH, Visscher H (июль 2001 г.). «Жизнь в конце пермской мертвой зоны». Труды Национальной академии наук Соединенных Штатов Америки. 98 (14): 7879–83. Bibcode:2001PNAS ... 98.7879L. Дои:10.1073 / pnas.131218098. ЧВК 35436. PMID 11427710.
См. Изображение 2
- ^ Полицейский округа, Бота Дж., Бьюик Р., Де Кок, МО, Эрвин Д.Х., Гаррисон Г.Х., Киршвинк Дж. Л., Смит Р. (февраль 2005 г.). «Резкое и постепенное вымирание среди позднепермских наземных позвоночных в бассейне Кару, Южная Африка». Наука. 307 (5710): 709–14. Bibcode:2005Sci ... 307..709W. CiteSeerX 10.1.1.503.2065. Дои:10.1126 / science.1107068. PMID 15661973. S2CID 46198018.
- ^ Касадеваль, Артуро; Хейтман, Джозеф (16 августа 2012 г.). "Грибы и рост млекопитающих". Патогены PLOS. 8 (8): e1002808. Дои:10.1371 / journal.ppat.1002808. ЧВК 3420938. PMID 22916007.
Это экологическое бедствие сопровождалось массовым обезлесением, за которым последовало цветение грибов, когда земля превратилась в массивный компост.
- ^ а б Шалчиан-Тебризи К., Минге М.А., Эспелунд М., Орр Р., Руден Т., Якобсен К.С., Кавальер-Смит Т. (2008). «Мультигенная филогения хоанозоа и происхождение животных». PLOS ONE. 3 (5): e2098. Bibcode:2008PLoSO ... 3.2098S. Дои:10.1371 / journal.pone.0002098. ЧВК 2346548. PMID 18461162.
- ^ Видеть Palaeos Fungi: Грибы В архиве 20 июня 2012 г. Wayback Machine для введения в таксономию грибов, включая недавние споры. архив
- ^ Celio GJ, Padamsee M, Dentinger BT, Bauer R, McLaughlin DJ (2006). «Сборка грибного древа жизни: построение структурной и биохимической базы данных». Микология. 98 (6): 850–9. Дои:10.3852 / mycologia.98.6.850. PMID 17486962. S2CID 23123595.
- ^ Силар П. (2016). Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes. HAL. п. 462. ISBN 978-2-9555841-0-1. В архиве из оригинала 25 сентября 2017 г.. Получено 7 апреля 2016.
- ^ Эссер К. (2014). Mycota VII A: Систематика и эволюция (2-е изд.). Springer. п. 461. ISBN 978-3-642-55317-2.
- ^ Тедерсоо, Лехо; Санчес-Рамирес, Сантьяго; Кольялг, Урмас; Бахрам, Мохаммад; Доринг, Маркус; Шигель, Дмитрий; Мэй, Том; Риберг, Мартин; Абаренков, Кесси (22 февраля 2018 г.). «Классификация грибов высокого уровня и инструмент для эволюционного экологического анализа». Грибковое разнообразие. 90 (1): 135–159. Дои:10.1007 / s13225-018-0401-0.
- ^ Gill EE, Fast NM (июнь 2006 г.). «Оценка взаимосвязи микроспоридий и грибов: комбинированный филогенетический анализ восьми генов». Ген. 375: 103–9. Дои:10.1016 / j.gene.2006.02.023. PMID 16626896.
- ^ Лю Ю.Дж., Ходсон М.С., Холл Б.Д. (2006). «Потеря жгутика произошла только один раз в линии грибов: филогенетическая структура царства грибов, выведенная из генов субъединиц РНК-полимеразы II». BMC Эволюционная биология. 6: 74. Дои:10.1186/1471-2148-6-74. ЧВК 1599754. PMID 17010206.
- ^ Джеймс Т.Ю., Летчер П.М., Лонгкор Дж. Э., Мозли-Стэндридж С. Е., Портер Д., Пауэлл М. Дж., Гриффит Г. В., Вилгалис Р. (2006). «Молекулярная филогения жгутиковых грибов (Chytridiomycota) и описание нового типа (Blastocladiomycota)». Микология. 98 (6): 860–71. Дои:10.3852 / mycologia.98.6.860. PMID 17486963. В архиве из оригинала 23 сентября 2015 г.. Получено 5 июля 2011.
- ^ Локхарт Р.Дж., Ван Дайк М.И., Бидл И.Р., Хамфрис П., Маккарти А.Дж. (август 2006 г.). «Молекулярно-биологическое обнаружение анаэробных кишечных грибов (Neocallimastigales) со свалок». Прикладная и экологическая микробиология. 72 (8): 5659–61. Дои:10.1128 / AEM.01057-06. ЧВК 1538735. PMID 16885325.
- ^ а б Реми В., Тейлор Т.Н., Хасс Х, Керп Х (декабрь 1994 г.). «Везикулярная арбускулярная микориза возрастом четыреста миллионов лет». Труды Национальной академии наук Соединенных Штатов Америки. 91 (25): 11841–3. Bibcode:1994PNAS ... 9111841R. Дои:10.1073 / пнас.91.25.11841. ЧВК 45331. PMID 11607500.
- ^ Шюсслер А., Шварцотт Д., Уокер С. (2001). "Новый грибной тип, Гломеромикота: филогения и эволюция ». Микологические исследования. 105 (12): 1413–1421. Дои:10.1017 / S0953756201005196. S2CID 82128210.
- ^ Алексопулос и другие., п. 145.
- ^ Для примера см. Сэмюэлс Дж. Дж. (Февраль 2006 г.). «Триходермия: систематика, половое состояние, экология». Фитопатология. 96 (2): 195–206. Дои:10.1094 / PHYTO-96-0195. PMID 18943925.
- ^ Рэдфорд А., Округ JH (июнь 1997 г.). «Геном и гены Neurospora crassa». Грибковая генетика и биология. 21 (3): 258–66. Дои:10.1006 / fgbi.1997.0979. PMID 9290240.
- ^ Вальверде М.Э., Паредес-Лопес О., Патаки Дж. К., Гевара-Лара Ф. (январь 1995 г.). «Уитлакоче (Ustilago maydis) как источник пищи - биология, состав и производство». Критические обзоры в области пищевой науки и питания. 35 (3): 191–229. Дои:10.1080/10408399509527699. PMID 7632354.
- ^ Зисова Л.Г. (2009). «Виды Malassezia и себорейный дерматит». Folia Medica. 51 (1): 23–33. PMID 19437895.
- ^ Perfect JR (июнь 2006 г.). "Криптококк neoformans: дрожжи, которые любят остро ". FEMS дрожжевые исследования. 6 (4): 463–8. Дои:10.1111 / j.1567-1364.2006.00051.x. PMID 16696642.
- ^ Блэквелл М., Спатафора Дж. В. (2004). «Грибки и их союзники». В Bills GF, Mueller GM, Foster MS (ред.). Биоразнообразие грибов: методы инвентаризации и мониторинга. Амстердам: Elsevier Academic Press. С. 18–20. ISBN 978-0-12-509551-8.
- ^ Аморосо, Мария Джулия; Бенимели, Клаудиа Сусана; Куоццо, Серджио Антонио (2013). Актинобактерии: применение в биоремедиации и производстве промышленных ферментов. CRC Press, Taylor & Francis Group. п. 33. ISBN 9781466578739.
- ^ Введение в почвенную биологию, Актинобактерии
- ^ Гэдд GM (январь 2007 г.). «Геомикология: биогеохимические преобразования горных пород, минералов, металлов и радионуклидов грибами, биопоглощение и биоремедиация». Микологические исследования. 111 (Чт 1): 3–49. Дои:10.1016 / j.mycres.2006.12.001. PMID 17307120.
- ^ а б Lindahl BD, Ihrmark K, Boberg J, Trumbore SE, Högberg P, Stenlid J, Finlay RD (2007). «Пространственное разделение процессов разложения подстилки и микоризного поглощения азота в бореальных лесах». Новый Фитолог. 173 (3): 611–20. Дои:10.1111 / j.1469-8137.2006.01936.x. PMID 17244056.
- ^ Бареа Дж. М., Посо М. Дж., Азкон Р., Азкон-Агилар К. (июль 2005 г.). «Сотрудничество микробов в ризосфере». Журнал экспериментальной ботаники. 56 (417): 1761–78. Дои:10.1093 / jxb / eri197. PMID 15911555.
- ^ а б Aanen DK (июнь 2006 г.). «Что пожнешь, то посеешь: сочетание уборки урожая и инокуляции стабилизирует взаимопонимание между термитами и грибами». Письма о биологии. 2 (2): 209–12. Дои:10.1098 / rsbl.2005.0424. ЧВК 1618886. PMID 17148364.
- ^ Никох Н., Фукацу Т. (апрель 2000 г.). «Межцарский хозяин, прыгающий под землю: филогенетический анализ энтомопаразитарных грибов рода кордицепс». Молекулярная биология и эволюция. 17 (4): 629–38. Дои:10.1093 / oxfordjournals.molbev.a026341. PMID 10742053.
- ^ Перотто С., Бонфанте П. (декабрь 1997 г.). «Бактериальные ассоциации с микоризными грибами: близкие и далекие друзья в ризосфере». Тенденции в микробиологии. 5 (12): 496–501. Дои:10.1016 / S0966-842X (97) 01154-2. PMID 9447662.
- ^ Арнольд А.Е., Мехия Л.С., Килло Д., Рохас Е.И., Мейнард З., Роббинс Н., Херре Э.А. (декабрь 2003 г.). «Эндофиты грибов ограничивают повреждение тропического дерева патогенами». Труды Национальной академии наук Соединенных Штатов Америки. 100 (26): 15649–54. Bibcode:2003ПНАС..10015649А. Дои:10.1073 / pnas.2533483100. ЧВК 307622. PMID 14671327.
- ^ а б Paszkowski U (август 2006 г.). «Мутуализм и паразитизм: инь и янь растительных симбиозов». Текущее мнение в области биологии растений. 9 (4): 364–70. Дои:10.1016 / j.pbi.2006.05.008. PMID 16713732.
- ^ а б Hube B (август 2004 г.). "От комменсала к патогену: стадийная и тканеспецифическая экспрессия гена грибковые микроорганизмы албиканс". Текущее мнение в микробиологии. 7 (4): 336–41. Дои:10.1016 / j.mib.2004.06.003. PMID 15288621.
- ^ Bonfante P (апрель 2003 г.). «Растения, микоризные грибы и эндобактерии: диалог между клетками и геномами». Биологический бюллетень. 204 (2): 215–20. Дои:10.2307/1543562. JSTOR 1543562. PMID 12700157. S2CID 12377410.
- ^ ван дер Хейден М.Г., Стрейтвольф-Энгель Р., Ридл Р., Зигрист С., Нойдекер А., Инейхен К., Боллер Т., Вимкен А., Сандерс И. Р. (2006). «Вклад микоризы в продуктивность растений, питание растений и структуру почвы на экспериментальных пастбищах». Новый Фитолог. 172 (4): 739–52. Дои:10.1111 / j.1469-8137.2006.01862.x. PMID 17096799. S2CID 17048094.
- ^ Хейден, Марсель Г. А. ван дер (15 апреля 2016 г.). «Подземные сети». Наука. 352 (6283): 290–291. Bibcode:2016Научный ... 352..290H. Дои:10.1126 / science.aaf4694. ISSN 0036-8075. PMID 27081054. S2CID 133399719.
- ^ Selosse MA, Ричард Ф., He X, Simard SW (ноябрь 2006 г.). «Микоризные сети: опасные связи?». Тенденции в экологии и эволюции. 21 (11): 621–8. Дои:10.1016 / j.tree.2006.07.003. PMID 16843567.
- ^ Йонг, Эд (14 апреля 2016 г.). «У деревьев есть свой Интернет». Атлантический океан. В архиве с оригинала 28 марта 2019 г.. Получено 9 марта 2019.
- ^ Merckx V, Bidartondo MI, Hynson NA (декабрь 2009 г.). «Микогетеротрофия: когда грибы являются хозяевами растений». Анналы ботаники. 104 (7): 1255–61. Дои:10.1093 / aob / mcp235. ЧВК 2778383. PMID 19767309.
- ^ Шульц Б., Бойл С. (июнь 2005 г.). «Эндофитный континуум». Микологические исследования. 109 (Pt 6): 661–86. Дои:10.1017 / S095375620500273X. PMID 16080390. S2CID 23182632.
- ^ Clay K, Schardl C (октябрь 2002 г.). «Эволюционные истоки и экологические последствия симбиоза эндофитов с травами». Американский натуралист. 160 Дополнение 4 (Дополнение 4): S99 – S127. Дои:10.1086/342161. PMID 18707456. S2CID 23909652.
- ^ Бродо И.М., Шарнофф С.Д. (2001). Лишайники Северной Америки. Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 978-0-300-08249-4.
- ^ Raven PH, Evert RF, Eichhorn, SE (2005). «14 - Грибки». Биология растений (7-е изд.). В. Х. Фриман. п.290. ISBN 978-0-7167-1007-3.
- ^ Дьякон, стр. 267.
- ^ Первис В. (2000). Лишайники. Вашингтон, округ Колумбия: издательство Smithsonian Institution Press совместно с Музеем естественной истории в Лондоне. стр.49–75. ISBN 978-1-56098-879-3.
- ^ Кирк и другие., п. 378.
- ^ Дьякон, стр. 267–276.
- ^ Дуглас А.Е. (ноябрь 1989 г.). «Симбиоз мицетоцитов у насекомых». Биологические обзоры Кембриджского философского общества. 64 (4): 409–34. Дои:10.1111 / j.1469-185X.1989.tb00682.x. PMID 2696562. S2CID 28345783.
- ^ Дьякон, стр. 277.
- ^ «Энтомологи: бразильская безжальная пчела должна выращивать особый вид грибов, чтобы выжить». Sci-News.com. 23 октября 2015 г. В архиве из оригинала 25 октября 2015 г.. Получено 25 октября 2015.
- ^ Нгуен Н.Х., Су СО, Блэквелл М. (2007). «Пять новых видов Candida в дрожжевых кладах, связанных с насекомыми, выделенных из Neuroptera и других насекомых». Микология. 99 (6): 842–58. Дои:10.3852 / mycologia.99.6.842. PMID 18333508. В архиве из оригинала 7 мая 2017 г.. Получено 5 июля 2011.
- ^ а б Филипяк, Михал; Вайнер, январь (март 2017 г.). «Динамика питания в процессе развития жуков-ксилофагов, связанная с изменением стехиометрии 11 элементов». Физиологическая энтомология. 42 (1): 73–84. Дои:10.1111 / phen.12168.
- ^ а б Улышен, Майкл Д. (2018). Сапроксильные насекомые: разнообразие, экология и сохранение. Спрингер, Чам. С. 429–469. Дои:10.1007/978-3-319-75937-1_13. ISBN 978-3-319-75937-1.
- ^ а б Улышен, Майкл Д. (2018). Разнообразие, экология и сохранение сапроксильных насекомых. Спрингер, Чам. С. 377–427. Дои:10.1007/978-3-319-75937-1_12. ISBN 978-3-319-75936-4.
- ^ Филипяк, Михал; Собчик, Лукаш; Вайнер, январь (2016). «Грибковая трансформация пней в подходящий ресурс для жуков-ксилофагов через изменения в соотношении элементов». Насекомые. 7 (2): 13. Дои:10.3390 / насекомые7020013. ЧВК 4931425.
- ^ Чендлер П.Дж. (2010). Справочник диптериста (2-е изд.). Великобритания: Общество энтомологов-любителей. С. 1–525.
- ^ Талбот, штат Нью-Джерси (2003). «По следу убийцы злаков: изучение биологии Magnaporthe grisea». Ежегодный обзор микробиологии. 57: 177–202. Дои:10.1146 / annurev.micro.57.030502.090957. PMID 14527276. S2CID 10582117.
- ^ Паолетти М., Бак К.В., Бразье К.М. (январь 2006 г.). "Селективное приобретение генов нового типа спаривания и вегетативной несовместимости посредством межвидового переноса генов у глобально вторгающихся эукариот. Офиостома новоульми". Молекулярная экология. 15 (1): 249–62. Дои:10.1111 / j.1365-294X.2005.02728.x. PMID 16367844. S2CID 22957337.
- ^ Gryzenhout M, Wingfield BD, Wingfield MJ (май 2006 г.). «Новые таксономические концепции важного лесного патогена Cryphonectria parasitica и родственных грибов». Письма о микробиологии FEMS. 258 (2): 161–72. Дои:10.1111 / j.1574-6968.2006.00170.x. PMID 16640568.
- ^ Ян И, Ян Э, Ан З, Лю Х (май 2007 г.). «Эволюция улавливающих нематод клеток хищных грибов Orbiliaceae на основе данных ДНК, кодирующих рРНК, и мультипротеиновых последовательностей». Труды Национальной академии наук Соединенных Штатов Америки. 104 (20): 8379–84. Bibcode:2007ПНАС..104.8379Y. Дои:10.1073 / pnas.0702770104. ЧВК 1895958. PMID 17494736.
- ^ Koeck, M .; Hardham, A.R .; Доддс; П.Н. (2011). «Роль эффекторов биотрофных и гемибиотрофных грибов в инфекции». Клеточная микробиология. 13 (12): 1849–1857. Дои:10.1111 / j.1462-5822.2011.01665.x. ЧВК 3218205. PMID 21848815.
- ^ «Asterotremella gen. Nov. Albida, анаморфные тремеллоидные дрожжи, выделенные из агариков Asterophora lycoperdoides и Asterophora parasitica». ResearchGate. Получено 19 апреля 2019.
- ^ Нильсен К., Хейтман Дж. (2007). «Пол и вирулентность патогенных грибов человека». Грибковая геномика. Успехи в генетике. 57. С. 143–73. Дои:10.1016 / S0065-2660 (06) 57004-X. ISBN 978-0-12-017657-1. PMID 17352904.
- ^ Брахейдж А.А. (декабрь 2005 г.). «Системные грибковые инфекции, вызываемые видами Aspergillus: эпидемиология, инфекционный процесс и детерминанты вирулентности». Текущие цели в отношении лекарств. 6 (8): 875–86. Дои:10.2174/138945005774912717. PMID 16375671.
- ^ Кауфман CA (январь 2007 г.). «Гистоплазмоз: клинические и лабораторные обновления». Обзоры клинической микробиологии. 20 (1): 115–32. Дои:10.1128 / CMR.00027-06. ЧВК 1797635. PMID 17223625.
- ^ Cushion MT, Smulian AG, Slaven BE, Sesterhenn T, Arnold J, Staben C, Porollo A, Adamczak R, Meller J (2007). «Транскриптом Pneumocystis carinii при молниеносной инфекции: углеводный обмен и концепция совместимого паразита». PLOS ONE. 2 (5): e423. Bibcode:2007PLoSO ... 2..423C. Дои:10.1371 / journal.pone.0000423. ЧВК 1855432. PMID 17487271.
- ^ Повар GC, Zumla AI (2008). Тропические болезни Мэнсона: консультации экспертов. Эдинбург, Шотландия: Saunders Ltd. стр. 347. ISBN 978-1-4160-4470-3.
- ^ Симон-Ноббе Б., Денк Ю., Пёлль В., Рид Р., Брайтенбах М. (2008). «Спектр грибковой аллергии». Международный архив аллергии и иммунологии. 145 (1): 58–86. Дои:10.1159/000107578. PMID 17709917.
- ^ Le Floch G, Rey P, Benizri E, Benhamou N, Tirilly Y (2003). «Воздействие ауксиновых соединений, продуцируемых антагонистическим грибком. Pythium oligandrum или второстепенный патоген Пифий группа F по росту растений ». Растительная почва. 257 (2): 459–470. Дои:10.1023 / а: 1027330024834. S2CID 22898373.
- ^ Пирсон М. Н., Бивер Р. Э., Бойн Б., Артур К. (январь 2009 г.). «Миковирусы мицелиальных грибов и их значение для патологии растений». Молекулярная патология растений. 10 (1): 115–28. Дои:10.1111 / j.1364-3703.2008.00503.x. ЧВК 6640375. PMID 19161358.
- ^ Бозарт РФ (октябрь 1972 г.). «Миковирусы: новое измерение в микробиологии». Перспективы гигиены окружающей среды. 2 (1): 23–39. Дои:10.1289 / ehp.720223. ЧВК 1474899. PMID 4628853.
- ^ Schardl CL, Panaccione DG, Tudzynski P (2006). Алкалоиды спорыньи - биология и молекулярная биология. Алкалоиды: химия и биология. 63. С. 45–86. Дои:10.1016 / S1099-4831 (06) 63002-2. ISBN 978-0-12-469563-4. PMID 17133714.
- ^ ван Эгмонд HP, Schothorst RC, Jonker MA (сентябрь 2007 г.). «Правила, касающиеся микотоксинов в пищевых продуктах: перспективы в глобальном и европейском контексте». Аналитическая и биоаналитическая химия. 389 (1): 147–57. Дои:10.1007 / s00216-007-1317-9. PMID 17508207.
- ^ Демейн А.Л., Фанг А. (2000). «Естественные функции вторичных метаболитов». История современной биотехнологии I. Достижения в области биохимической инженерии / биотехнологии. 69. С. 1–39. Дои:10.1007/3-540-44964-7_1. ISBN 978-3-540-67793-2. PMID 11036689.
- ^ Рольфс М., Альберт М., Келлер Н.П., Кемпкен Ф. (октябрь 2007 г.). «Вторичные химические вещества защищают плесень от грибка». Письма о биологии. 3 (5): 523–5. Дои:10.1098 / rsbl.2007.0338. ЧВК 2391202. PMID 17686752.
- ^ Молина Л., Кахманн Р. (июль 2007 г.). "Ген Ustilago maydis, участвующий в H2О2 детоксикация необходима для вирулентности ". Растительная клетка. 19 (7): 2293–309. Дои:10.1105 / tpc.107.052332. ЧВК 1955693. PMID 17616735.
- ^ Kojic M, Zhou Q, Lisby M, Holloman WK (январь 2006 г.). «Взаимодействие Rec2 с Brh2 и Rad51 уравновешивает рекомбинационную репарацию в Ustilago maydis». Молекулярная и клеточная биология. 26 (2): 678–88. Дои:10.1128 / MCB.26.2.678-688.2006. ЧВК 1346908. PMID 16382157.
- ^ а б Мичод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптивное значение секса у микробных возбудителей» (PDF). Инфекция, генетика и эволюция. 8 (3): 267–85. Дои:10.1016 / j.meegid.2008.01.002. PMID 18295550. В архиве (PDF) из оригинала 16 мая 2017 г.. Получено 22 июля 2013.
- ^ а б Fan W, Kraus PR, Boily MJ, Heitman J (август 2005 г.). "Криптококк neoformans экспрессия гена при инфицировании макрофагами мышей ". Эукариотическая клетка. 4 (8): 1420–33. Дои:10.1128 / EC.4.8.1420-1433.2005. ЧВК 1214536. PMID 16087747.
- ^ а б Лин X, Халл CM, Хейтман Дж. (Апрель 2005 г.). "Половое размножение между партнерами одного типа спаривания в Криптококк neoformans". Природа. 434 (7036): 1017–21. Bibcode:2005 Натур.434.1017L. Дои:10.1038 / природа03448. PMID 15846346. S2CID 52857557.
- ^ Финчем-младший (март 1989 г.). «Трансформация в грибах». Микробиологические обзоры. 53 (1): 148–70. Дои:10.1128 / MMBR.53.1.148-170.1989. ЧВК 372721. PMID 2651864.
- ^ Хокинс К.М., Смолке CD (сентябрь 2008 г.). "Производство бензилизохинолиновых алкалоидов в Saccharomyces cerevisiae". Природа Химическая Биология. 4 (9): 564–73. Дои:10.1038 / nchembio.105. ЧВК 2830865. PMID 18690217.
- ^ Хуан Б., Го Дж., Йи Б., Ю X, Сун Л., Чен В. (июль 2008 г.). «Гетерологичное производство вторичных метаболитов в качестве фармацевтических препаратов в Saccharomyces cerevisiae". Письма о биотехнологии. 30 (7): 1121–37. Дои:10.1007 / s10529-008-9663-z. PMID 18512022. S2CID 2222358.
- ^ Brakhage AA, Spröte P, Al-Abdallah Q, Gehrke A, Plattner H, Tüncher A (2004). «Регуляция биосинтеза пенициллина в нитчатых грибах». Молекулярная биотехнология грибковых бета-лактамных антибиотиков и родственных пептидных синтетаз. Достижения в области биохимической инженерии / биотехнологии. 88. С. 45–90. Дои:10.1007 / b99257. ISBN 978-3-540-22032-9. PMID 15719552.
- ^ Пан А., Лоренцотти С., Зонкада А. (январь 2008 г.). "Registered and investigational drugs for the treatment of methicillin-resistant Staphylococcus aureus infection". Recent Patents on Anti-Infective Drug Discovery. 3 (1): 10–33. Дои:10.2174/157489108783413173. PMID 18221183.
- ^ Fajardo A, Martínez JL (April 2008). "Antibiotics as signals that trigger specific bacterial responses". Current Opinion in Microbiology. 11 (2): 161–7. Дои:10.1016/j.mib.2008.02.006. PMID 18373943.
- ^ Loo DS (2006). "Systemic antifungal agents: an update of established and new therapies". Advances in Dermatology. 22: 101–24. Дои:10.1016/j.yadr.2006.07.001. PMID 17249298.
- ^ Manzoni M, Rollini M (April 2002). "Biosynthesis and biotechnological production of statins by filamentous fungi and application of these cholesterol-lowering drugs". Applied Microbiology and Biotechnology. 58 (5): 555–64. Дои:10.1007/s00253-002-0932-9. PMID 11956737. S2CID 5761188.
- ^ el-Mekkawy S, Meselhy MR, Nakamura N, Tezuka Y, Hattori M, Kakiuchi N, Shimotohno K, Kawahata T, Otake T (November 1998). "Anti-HIV-1 and anti-HIV-1-protease substances from Ganoderma lucidum". Phytochemistry. 49 (6): 1651–7. Дои:10.1016/S0031-9422(98)00254-4. PMID 9862140.
- ^ El Dine RS, El Halawany AM, Ma CM, Hattori M (June 2008). "Anti-HIV-1 protease activity of lanostane triterpenes from the vietnamese mushroom Ganoderma colossum". Journal of Natural Products. 71 (6): 1022–6. Дои:10.1021/np8001139. PMID 18547117.
- ^ а б Hetland G, Johnson E, Lyberg T, Bernardshaw S, Tryggestad AM, Grinde B (October 2008). "Effects of the medicinal mushroom Agaricus blazei Murill on immunity, infection and cancer". Scandinavian Journal of Immunology. 68 (4): 363–70. Дои:10.1111/j.1365-3083.2008.02156.x. PMID 18782264. S2CID 3866471.
- ^ Yuen JW, Gohel MD (2005). "Anticancer effects of Ganoderma lucidum: a review of scientific evidence". Nutrition and Cancer. 53 (1): 11–7. Дои:10.1207/s15327914nc5301_2. HDL:10397/26313. PMID 16351502. S2CID 24314616.
- ^ Sullivan R, Smith JE, Rowan NJ (2006). "Medicinal mushrooms and cancer therapy: translating a traditional practice into Western medicine". Perspectives in Biology and Medicine. 49 (2): 159–70. Дои:10.1353/pbm.2006.0034. PMID 16702701. S2CID 29723996.
- ^ Halpern GM, Miller A (2002). Medicinal Mushrooms: Ancient Remedies for Modern Ailments. New York, New York: M. Evans and Co. p. 116. ISBN 978-0-87131-981-4.
- ^ Fisher M, Yang LX (2002). "Anticancer effects and mechanisms of polysaccharide-K (PSK): implications of cancer immunotherapy". Anticancer Research. 22 (3): 1737–54. PMID 12168863.
- ^ Firenzuoli F, Gori L, Lombardo G (March 2008). "The Medicinal Mushroom Agaricus blazei Murrill: Review of Literature and Pharmaco-Toxicological Problems". Evidence-Based Complementary and Alternative Medicine. 5 (1): 3–15. Дои:10.1093/ecam/nem007. ЧВК 2249742. PMID 18317543.
- ^ Paterson RR (September 2006). "Ganoderma - a therapeutic fungal biofactory". Phytochemistry. 67 (18): 1985–2001. Дои:10.1016/j.phytochem.2006.07.004. HDL:1822/5522. PMID 16905165.
- ^ Paterson RR (May 2008). "Cordyceps: a traditional Chinese medicine and another fungal therapeutic biofactory?". Phytochemistry. 69 (7): 1469–95. Дои:10.1016/j.phytochem.2008.01.027. HDL:1822/7896. ЧВК 7111646. PMID 18343466.
- ^ Kulp K (2000). Handbook of Cereal Science and Technology. CRC Press. ISBN 978-0-8247-8294-8.
- ^ Piskur J, Rozpedowska E, Polakova S, Merico A, Compagno C (April 2006). "How did Saccharomyces evolve to become a good brewer?". Trends in Genetics. 22 (4): 183–6. Дои:10.1016/j.tig.2006.02.002. PMID 16499989.
- ^ Abe K, Gomi K, Hasegawa F, Machida M (September 2006). "Impact of Aspergillus oryzae genomics on industrial production of metabolites". Mycopathologia. 162 (3): 143–53. Дои:10.1007/s11046-006-0049-2. PMID 16944282. S2CID 36874528.
- ^ Hachmeister KA, Fung DY (1993). "Tempeh: a mold-modified indigenous fermented food made from soybeans and/or cereal grains". Critical Reviews in Microbiology. 19 (3): 137–88. Дои:10.3109/10408419309113527. PMID 8267862.
- ^ Jørgensen TR (December 2007). "Identification and toxigenic potential of the industrially important fungi, Aspergillus oryzae и Aspergillus sojae". Journal of Food Protection. 70 (12): 2916–34. Дои:10.4315/0362-028X-70.12.2916. PMID 18095455.
- ^ O'Donnell K, Cigelnik E, Casper HH (February 1998). "Molecular phylogenetic, morphological, and mycotoxin data support reidentification of the Quorn mycoprotein fungus as Fusarium venenatum". Fungal Genetics and Biology. 23 (1): 57–67. Дои:10.1006/fgbi.1997.1018. PMID 9501477.
- ^ Stamets P (2000). Growing Gourmet and Medicinal Mushrooms=[Shokuyō oyobi yakuyō kinoko no saibai]. Berkeley, California: Ten Speed Press. pp. 233–248. ISBN 978-1-58008-175-7.
- ^ Hall, pp. 13–26.
- ^ Kinsella JE, Hwang DH (November 1976). "Enzymes of Penicillium roqueforti involved in the biosynthesis of cheese flavor". Критические обзоры в области пищевой науки и питания. 8 (2): 191–228. Дои:10.1080/10408397609527222. PMID 21770.
- ^ Erdogan A, Gurses M, Sert S (August 2003). "Isolation of moulds capable of producing mycotoxins from blue mouldy Tulum cheeses produced in Turkey". International Journal of Food Microbiology. 85 (1–2): 83–5. Дои:10.1016/S0168-1605(02)00485-3. PMID 12810273.
- ^ Orr DB, Orr RT (1979). Mushrooms of Western North America. Berkeley, California: University of California Press. п. 17. ISBN 978-0-520-03656-7.
- ^ Vetter J (January 1998). "Toxins of Amanita phalloides". Токсикон. 36 (1): 13–24. Дои:10.1016/S0041-0101(97)00074-3. PMID 9604278.
- ^ Leathem AM, Dorran TJ (March 2007). "Poisoning due to raw Gyromitra esculenta (false morels) west of the Rockies". Canadian Journal of Emergency Medicine. 9 (2): 127–30. Дои:10.1017/s1481803500014937. PMID 17391587.
- ^ Karlson-Stiber C, Persson H (September 2003). "Cytotoxic fungi--an overview". Токсикон. 42 (4): 339–49. Дои:10.1016/S0041-0101(03)00238-1. PMID 14505933.
- ^ Michelot D, Melendez-Howell LM (February 2003). "Amanita muscaria: chemistry, biology, toxicology, and ethnomycology". Mycological Research. 107 (Pt 2): 131–46. Дои:10.1017/S0953756203007305. PMID 12747324. S2CID 41451034.
- ^ Hall, p. 7.
- ^ Ammirati JF, McKenny M, Stuntz DE (1987). The New Savory Wild Mushroom. Seattle, Washington: University of Washington Press. pp. xii–xiii. ISBN 978-0-295-96480-5.
- ^ López-Gómez J, Molina-Meyer M (February 2006). "The competitive exclusion principle versus biodiversity through competitive segregation and further adaptation to spatial heterogeneities". Theoretical Population Biology. 69 (1): 94–109. Дои:10.1016/j.tpb.2005.08.004. PMID 16223517.
- ^ Becker H (1998). "Setting the Stage To Screen Biocontrol Fungi". United States Department of Agriculture, Agricultural Research Service. В архиве from the original on 16 January 2009. Получено 23 февраля 2009.
- ^ Keiller TS. "Whey-based fungal microfactory technology for enhanced biological pest management using fungi" (PDF). UVM Innovations. Архивировано из оригинал (PDF) on 29 October 2011. Получено 29 октября 2011.
- ^ Deshpande MV (1999). "Mycopesticide production by fermentation: potential and challenges". Critical Reviews in Microbiology. 25 (3): 229–43. Дои:10.1080/10408419991299220. PMID 10524330.
- ^ Thomas MB, Read AF (May 2007). "Can fungal biopesticides control malaria?". Обзоры природы. Микробиология. 5 (5): 377–83. Дои:10.1038/nrmicro1638. HDL:1842/2089. PMID 17426726. S2CID 14460348.
- ^ Bush LP, Wilkinson HH, Schardl CL (May 1997). "Bioprotective Alkaloids of Grass-Fungal Endophyte Symbioses". Физиология растений. 114 (1): 1–7. Дои:10.1104/pp.114.1.1. ЧВК 158272. PMID 12223685.
- ^ Bouton JH, Latch GC, Hill NS, Hoveland CS, McCann MA, Watson RH, Parish JA, Hawkins LL, Thompson FN (2002). "Reinfection of Tall Fescue Cultivars with Non-Ergot Alkaloid–Producing Endophytes". Agronomy Journal. 94 (3): 567–574. Дои:10.2134/agronj2002.5670.
- ^ Parish JA, McCann MA, Watson RH, Hoveland CS, Hawkins LL, Hill NS, Bouton JH (May 2003). "Use of nonergot alkaloid-producing endophytes for alleviating tall fescue toxicosis in sheep". Journal of Animal Science. 81 (5): 1316–22. Дои:10.2527/2003.8151316x. PMID 12772860.
- ^ Christian V, Shrivastava R, Shukla D, Modi HA, Vyas BR (April 2005). "Degradation of xenobiotic compounds by lignin-degrading white-rot fungi: enzymology and mechanisms involved". Indian Journal of Experimental Biology. 43 (4): 301–12. PMID 15875713.
- ^ "Fungi to fight 'toxic war zones'". Новости BBC. 5 May 2008. В архиве from the original on 15 September 2017. Получено 12 мая 2008.
- ^ Fomina M, Charnock JM, Hillier S, Alvarez R, Gadd GM (July 2007). "Fungal transformations of uranium oxides". Environmental Microbiology. 9 (7): 1696–710. Дои:10.1111/j.1462-2920.2007.01288.x. PMID 17564604.
- ^ Fomina M, Charnock JM, Hillier S, Alvarez R, Livens F, Gadd GM (May 2008). "Role of fungi in the biogeochemical fate of depleted uranium". Текущая биология. 18 (9): R375–7. Дои:10.1016/j.cub.2008.03.011. PMID 18460315. S2CID 52805144.
- ^ Beadle GW, Tatum EL (November 1941). "Genetic Control of Biochemical Reactions in Neurospora". Труды Национальной академии наук Соединенных Штатов Америки. 27 (11): 499–506. Bibcode:1941PNAS...27..499B. Дои:10.1073/pnas.27.11.499. ЧВК 1078370. PMID 16588492.
- ^ Datta A, Ganesan K, Natarajan K (1989). "Current Trends in Candida albicans Research". Current trends in Candida albicans исследование. Advances in Microbial Physiology. 30. pp. 53–88. Дои:10.1016/S0065-2911(08)60110-1. ISBN 978-0-12-027730-8. PMID 2700541.
- ^ Dean RA, Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, Orbach MJ, et al. (April 2005). "The genome sequence of the rice blast fungus Magnaporthe grisea". Природа. 434 (7036): 980–6. Bibcode:2005Natur.434..980D. Дои:10.1038/nature03449. PMID 15846337.
- ^ Daly R, Hearn MT (2005). "Expression of heterologous proteins in Pichia pastoris: a useful experimental tool in protein engineering and production". Journal of Molecular Recognition. 18 (2): 119–38. Дои:10.1002/jmr.687. PMID 15565717. S2CID 7476149.
- ^ Schlegel HG (1993). General Microbiology. Кембридж, Великобритания: Издательство Кембриджского университета. п. 360. ISBN 978-0-521-43980-0.
- ^ Joseph B, Ramteke PW, Thomas G (2008). "Cold active microbial lipases: some hot issues and recent developments". Biotechnology Advances. 26 (5): 457–70. Дои:10.1016/j.biotechadv.2008.05.003. PMID 18571355.
- ^ Kumar R, Singh S, Singh OV (May 2008). "Bioconversion of lignocellulosic biomass: biochemical and molecular perspectives". Journal of Industrial Microbiology & Biotechnology. 35 (5): 377–91. Дои:10.1007/s10295-008-0327-8. PMID 18338189. S2CID 4830678.
- ^ "Trichoderma spp., including T. harzianum, T. viride, T. koningii, T. hamatum and other spp. Deuteromycetes, Moniliales (asexual classification system)". Biological Control: A Guide to Natural Enemies in North America. Архивировано из оригинал on 14 April 2011. Получено 10 июля 2007.
- ^ Olempska-Beer ZS, Merker RI, Ditto MD, DiNovi MJ (July 2006). "Food-processing enzymes from recombinant microorganisms--a review". Regulatory Toxicology and Pharmacology. 45 (2): 144–158. Дои:10.1016/j.yrtph.2006.05.001. PMID 16769167. В архиве from the original on 3 July 2019. Получено 3 июля 2019.
- ^ Polizeli ML, Rizzatti AC, Monti R, Terenzi HF, Jorge JA, Amorim DS (June 2005). "Xylanases from fungi: properties and industrial applications". Applied Microbiology and Biotechnology. 67 (5): 577–91. Дои:10.1007/s00253-005-1904-7. PMID 15944805. S2CID 22956.
Цитированная литература
- Ainsworth GC (1976). Introduction to the History of Mycology. Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0-521-11295-6.
- Alexopoulos CJ, Mims CW, Blackwell M (1996). Introductory Mycology. Джон Уайли и сыновья. ISBN 978-0-471-52229-4.
- Chandler PJ (2010). A Dipterist's Handbook (2-е изд.). The Amateur Entomologists' Society. pp. 1–525.
- Deacon J (2005). Fungal Biology. Cambridge, Massachusetts: Blackwell Publishers. ISBN 978-1-4051-3066-0.
- Hall IR (2003). Edible and Poisonous Mushrooms of the World. Portland, Oregon: Timber Press. ISBN 978-0-88192-586-9.
- Hanson JR (2008). The Chemistry of Fungi. Royal Society Of Chemistry. ISBN 978-0-85404-136-7.
- Jennings DH, Lysek G (1996). Fungal Biology: Understanding the Fungal Lifestyle. Guildford, UK: Bios Scientific Publishers Ltd. ISBN 978-1-85996-150-6.
- Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008). Словарь грибов (10-е изд.). Уоллингфорд, Великобритания: CAB International. ISBN 978-0-85199-826-8.
- Taylor EL, Taylor TN (1993). The Biology and Evolution of Fossil Plants. Englewood Cliffs, New Jersey: Prentice Hall. ISBN 978-0-13-651589-0.
внешняя ссылка
| Library resources about Грибок |
- Entangle Life: How Fungi Make Our Worlds, Change Our Minds, & Shape Our Futures Book by Merlin Sheldrake (2020), Random House. ISBN 978-0525510314
- Tree of Life web project: Fungi
- Mushroom Observer (mushroomobserver.org ), a collaborative fungus recording and identification project
- FUNGI в BoDD – Botanical Dermatology Database
Elements of nature | |||||
|---|---|---|---|---|---|
| Вселенная | |||||
| земной шар | |||||
| Погода | |||||
| Natural environment | |||||
| Жизнь | |||||
| |||||