Митохондрия - Mitochondrion
| Клеточная биология | |
|---|---|
| В животная клетка | |
 Компоненты типичной животной клетки:
|
В митохондрия (/ˌмаɪтəˈkɒпdрɪəп/,[1] множественное число митохондрии) является двойныммембрана -граница органелла найдено в большинстве эукариотический организмы. Некоторые клетки в некоторых многоклеточный у организмов отсутствуют митохондрии (например, зрелые млекопитающие красные кровяные тельца ). Ряд одноклеточных организмов, таких как микроспоридии, парабазалиды, и дипломонады, уменьшили или преобразовали свои митохондрии в другие структуры.[2] На сегодняшний день только один эукариот, Monocercomonoides, как известно, полностью потеряли свои митохондрии,[3] и один многоклеточный организм, Henneguya salminicola, как известно, сохранили связанные с митохондриями органеллы в связи с полной потерей их митохондриального генома.[3][4][5]
Митохондрии производят большую часть запасов клеточной аденозинтрифосфат (АТФ), используемый как источник химическая энергия.[6] Таким образом, митохондрия называется электростанция ячейки.[7]
Митохондрии обычно составляют от 0,75 до 3мкм ² на площади[8] но значительно различаются по размеру и структуре. Если специально окрашенный, они не видны. Помимо обеспечения клеточной энергии митохондрии участвуют в других задачах, таких как сигнализация, клеточная дифференциация, и смерть клетки, а также сохранение контроля над клеточный цикл и рост клеток.[9] Митохондриальный биогенез в свою очередь, координируется во времени с этими клеточными процессами.[10][11] Митохондрии участвуют в нескольких заболеваниях и состояниях человека, таких как митохондриальные нарушения,[12] сердечная дисфункция,[13] сердечная недостаточность[14] и аутизм.[15]
Количество митохондрий в клетке может широко варьироваться в зависимости от организм, ткань, и тип ячейки. красные кровяные тельца не имеют митохондрий, тогда как клетки печени может быть больше 2000.[16][17] В органелла состоит из отсеков, которые выполняют специализированные функции. Эти отсеки или области включают внешнюю мембрану, межмембранное пространство, внутренняя мембрана, кристы и матрица.
Хотя большая часть клеток ДНК содержится в ядро клетки, митохондрия имеет свой геном («митогеном»), который по существу похож на бактериальный геномы.[18] Митохондриальные белки (белки, транскрибируемые с митохондриальная ДНК ) варьируются в зависимости от ткани и вида. У людей было идентифицировано 615 различных типов белков из сердечный митохондрии,[19] тогда как в крысы Сообщалось о 940 белках.[20] Митохондриальный протеом считается динамически регулируемым.[21]
Структура


{kind=link}
Митохондрия содержит внешнюю и внутреннюю мембраны, состоящие из фосфолипидные бислои и белки.[16] Две мембраны имеют разные свойства. Из-за этой двухмембранной организации митохондрия состоит из пяти отдельных частей:
- внешняя митохондриальная мембрана,
- межмембранное пространство (пространство между внешней и внутренней мембранами),
- внутренняя митохондриальная мембрана,
- то кристы пространство (образованное складками внутренней мембраны), и
- то матрица (пространство внутри внутренней мембраны).
Митохондрии, лишенные внешней мембраны, называются митопласты.
Внешняя мембрана
В внешняя митохондриальная мембрана, охватывающая всю органеллу, составляет от 60 до 75 ангстремы (Å) толщиной. Он имеет соотношение белков и фосфолипидов, аналогичное соотношению клеточная мембрана (примерно 1: 1 по весу). Он содержит большое количество интегральные мембранные белки называется порины. Основным транспортным белком является порообразующий зависимый от напряжения анионный канал (VDAC). В VDAC является основным транспортером нуклеотиды, ионы и метаболиты между цитозоль и межмембранное пространство.[23][24] Он сформирован как бета-баррель который охватывает внешнюю мембрану, как и в грамотрицательные бактерии мембрана.[25] Более крупные белки могут проникать в митохондрии, если сигнальная последовательность на их N-конец связывается с большой мультисубъединицей белок называется транслоказа в наружной мембране, который затем активно двигается их через мембрану.[26] Митохондриальный протеины импортируются через специализированные комплексы транслокаций.
Наружная мембрана также содержит ферменты участвует в таких разнообразных мероприятиях, как удлинение жирные кислоты, окисление из адреналин, а деградация из триптофан. Эти ферменты включают моноаминоксидаза, ротенон -нечувствительная НАДН-цитохром с-редуктаза, кинуренин гидроксилаза и жирная кислота Co-A лигаза. Нарушение внешней мембраны позволяет протеинам из межмембранного пространства просачиваться в цитозоль, что приводит к гибели клеток.[27] Наружная мембрана митохондрий может ассоциироваться с мембраной эндоплазматического ретикулума (ER) в структуре, называемой MAM (митохондриально-ассоциированная ER-мембрана). Это важно для передачи сигналов кальция в ER-митохондриях и участвует в переносе липидов между ER и митохондриями.[28] Снаружи внешней мембраны есть маленькие (диаметром 60 Å) частицы, называемые частичками Парсона.
Межмембранное пространство
В митохондриальное межмембранное пространство это пространство между внешней и внутренней мембранами. Оно также известно как перимитохондриальное пространство. Поскольку внешняя мембрана свободно проницаема для малых молекул, концентрация малых молекул, таких как ионы и сахара, в межмембранном пространстве такая же, как и в межмембранном пространстве. цитозоль.[16] Однако большие белки должны иметь специфическую сигнальную последовательность, чтобы транспортироваться через внешнюю мембрану, поэтому белковый состав этого пространства отличается от белкового состава цитозоль. Один белок который локализован в межмембранном пространстве таким образом, цитохром с.[27]
Внутренняя мембрана
Внутренняя митохондриальная мембрана содержит белки с тремя типами функций:[16]
- Те, кто выполняет электронная транспортная цепь редокс реакции
- АТФ-синтаза, который генерирует АТФ в матрице
- Специфический транспортные белки которые регулируют метаболит переход в и из митохондриальный матрикс
Он содержит более 151 различных полипептиды, и имеет очень высокое соотношение белка к фосфолипидам (более 3: 1 по весу, что составляет примерно 1 белок на 15 фосфолипидов). Внутренняя мембрана содержит около 1/5 всего белка митохондрии.[29] Кроме того, внутренняя мембрана богата необычным фосфолипидом, кардиолипин. Этот фосфолипид был первоначально открыт в корова сердца в 1942 г. и обычно характерен для плазматических мембран митохондрий и бактерий.[30] Кардиолипин содержит четыре жирные кислоты, а не две, и может помочь сделать внутреннюю мембрану непроницаемой.[16] В отличие от внешней мембраны, внутренняя мембрана не содержит поринов и очень непроницаема для всех молекул. Практически все ионы и молекулы требуют специальных мембранных транспортеров для входа в матрицу или выхода из нее. Белки попадают в матрицу через транслоказа внутренней мембраны (TIM) комплекс или через Oxa1.[26] Кроме того, существует мембранный потенциал на внутренней мембране, образованный действием ферменты из электронная транспортная цепь. Внутренняя мембрана слияние опосредуется белком внутренней мембраны OPA1.[31]
Кристы

Внутренняя митохондриальная мембрана разделена на многочисленные складки, называемые кристы, которые увеличивают площадь поверхности внутренней митохондриальной мембраны, увеличивая ее способность производить АТФ. Для типичных митохондрий печени площадь внутренней мембраны примерно в пять раз больше внешней мембраны. Это соотношение варьируется, и митохондрии из клеток, которые имеют большую потребность в АТФ, таких как мышечные клетки, содержат еще больше крист. Митохондрии в одной и той же клетке могут иметь существенно разную плотность криста, те, которые необходимы для производства большего количества энергии, имеют гораздо большую поверхность криста-мембраны.[32] Эти складки усеяны маленькими круглыми телами, известными как F1 частицы или оксисомы.[33]
Матрица
Матрица - это пространство, ограниченное внутренней мембраной. Он содержит около 2/3 всех белков митохондрии.[16] Матрикс играет важную роль в производстве АТФ с помощью АТФ-синтазы, содержащейся во внутренней мембране. Матрикс содержит высококонцентрированную смесь сотен ферментов, особенных митохондриальных рибосомы, тРНК, и несколько копий митохондриальная ДНК геном. Среди ферментов основные функции включают окисление пируват и жирные кислоты, а цикл лимонной кислоты.[16] Молекулы ДНК упаковываются в нуклеоиды с помощью белков, один из которых TFAM.[34]
Функция
Наиболее важная роль митохондрий заключается в производстве энергетической валюты клетки, АТФ (т.е. фосфорилирование ADP ) через дыхание и регулировать клеточную метаболизм.[17] Центральный набор реакций, участвующих в производстве АТФ, известен под общим названием цикл лимонной кислоты, или Кребс цикл. Однако митохондрия выполняет много других функций, помимо производства АТФ.
Преобразование энергии
Доминирующая роль митохондрий - производство АТФ, о чем свидетельствует большое количество белков во внутренней мембране для этой задачи. Это достигается путем окисления основных продуктов глюкоза: пируват, и НАДН, которые производятся в цитозоле.[17] Этот тип клеточное дыхание известный как аэробного дыхания, зависит от наличия кислород, который обеспечивает большую часть высвобождаемой энергии.[35] Когда кислород ограничен, гликолитические продукты будут метаболизироваться анаэробная ферментация, процесс, который не зависит от митохондрий.[17] Производство АТФ из глюкозы и кислорода дает примерно в 13 раз больший выход при аэробном дыхании по сравнению с ферментацией.[36] Митохондрии растений также могут производить ограниченное количество АТФ без кислорода, используя альтернативный субстрат. нитрит.[37] АТФ пересекает внутреннюю мембрану с помощью специфический белок, и через внешнюю мембрану через порины. ADP возвращается по тому же маршруту.
Пируват и цикл лимонной кислоты
Пируват молекулы, произведенные гликолиз находятся активно транспортируется через внутреннюю митохондриальную мембрану и в матрикс, где они могут быть окисленный и в сочетании с кофермент А образовать СО2, ацетил-КоА, и НАДН,[17] или они могут быть карбоксилированный (к пируваткарбоксилаза ) с образованием оксалоацетата. Эта последняя реакция «восполняет» количество оксалоацетата в цикле лимонной кислоты и, следовательно, является анаплеротическая реакция, увеличивая способность цикла метаболизировать ацетил-КоА, когда ткани необходимы энергии (например, в мышца ) внезапно усиливаются из-за активности.[38]
В цикле лимонной кислоты все промежуточные соединения (например, цитрат, изоцитрат, альфа-кетоглутарат, сукцинат, фумарат, малат и оксалоацетат) регенерируются во время каждого цикла цикла. Таким образом, добавление большего количества любого из этих промежуточных продуктов к митохондрии означает, что дополнительное количество остается в пределах цикла, увеличивая количество всех других промежуточных продуктов по мере того, как одно превращается в другое. Следовательно, добавление любого из них к циклу имеет анаплеротический эффект, а его удаление имеет катаплеротический эффект. Эти анаплеротические и катаплеротический реакции будут в течение цикла увеличивать или уменьшать количество оксалоацетата, доступного для объединения с ацетил-КоА с образованием лимонной кислоты. Это, в свою очередь, увеличивает или уменьшает скорость АТФ производство митохондрией и, следовательно, доступность АТФ для клетки.[38]
Ацетил-КоА, с другой стороны, образуется в результате окисления пирувата или бета-окисление из жирные кислоты, это единственное топливо, которое входит в цикл лимонной кислоты. При каждом повороте цикла одна молекула ацетил-КоА расходуется на каждую молекулу оксалоацетата, присутствующего в митохондриальном матриксе, и никогда не регенерируется. Это окисление ацетатной части ацетил-КоА, которое производит CO2 и вода, при этом высвобождаемая энергия улавливается в форме АТФ.[38]
В печени карбоксилирование из цитозольный пирувата в оксалоацетат внутри митохондрий является ранним шагом в глюконеогенный путь, который преобразует лактат и деаминированный аланин в глюкозу,[17][38] под воздействием высокого уровня глюкагон и / или адреналин в крови.[38] Здесь добавление оксалоацетата к митохондрии не имеет чистого анаплеротического эффекта, поскольку другой промежуточный продукт цикла лимонной кислоты (малат) немедленно удаляется из митохондрии и превращается в цитозольный оксалоацетат, который в конечном итоге превращается в глюкозу в процессе, который почти противоположно гликолиз.[38]
Ферменты цикла лимонной кислоты расположены в матриксе митохондрий, за исключением сукцинатдегидрогеназа, который связан с внутренней митохондриальной мембраной как часть Комплекса II.[39] Цикл лимонной кислоты окисляет ацетил-КоА до диоксида углерода и в процессе производит восстановленные кофакторы (три молекулы НАДН и одна молекула FADH2 ), которые являются источником электронов для электронная транспортная цепь, и молекула GTP (который легко превращается в АТФ).[17]
НАДН и ФАД2: цепь переноса электронов
В электроны от НАДН и ФАДН2 переходят в кислород (O2), богатая энергией молекула,[35] и водород (протоны) в несколько этапов через цепь переноса электронов. НАДН и ФАД2 молекулы производятся в матриксе через цикл лимонной кислоты, но также производятся в цитоплазме за счет гликолиз. Сокращающие эквиваленты из цитоплазмы может быть импортирован через малат-аспартатный челнок система антипортер белков или подавать в цепь переноса электронов с помощью глицеринфосфатный челнок.[17] Белковые комплексы во внутренней мембране (НАДН-дегидрогеназа (убихинон), цитохром с редуктаза, и цитохром с оксидаза ) выполнить передачу, и постепенное высвобождение энергии используется для накачки протоны (ЧАС+) в межмембранное пространство. Этот процесс эффективен, но небольшой процент электронов может преждевременно восстанавливать кислород, образуя активные формы кислорода Такие как супероксид.[17] Это может вызвать окислительный стресс в митохондриях и может способствовать снижению митохондриальной функции, связанной с процессом старения.[40]
По мере увеличения концентрации протонов в межмембранном пространстве сильное электрохимический градиент устанавливается через внутреннюю мембрану. Протоны могут вернуться в матрицу через АТФ-синтаза комплекса, а их потенциальная энергия используется для синтеза АТФ из АДФ и неорганического фосфата (Pя).[17] Этот процесс называется хемиосмос, и впервые был описан Питер Митчелл,[41][42] кто был награжден 1978 Нобелевская премия по химии за его работу. Позже часть Нобелевской премии по химии 1997 г. была присуждена Пол Д. Бойер и Джон Э. Уокер за их разъяснение механизма работы АТФ-синтазы.[43]
Производство тепла
При определенных условиях протоны могут повторно входить в матрикс митохондрий, не участвуя в синтезе АТФ. Этот процесс известен как утечка протонов или же митохондриальное разобщение и это связано с облегченное распространение протонов в матрицу. Результатом этого процесса является высвобождение потенциальной энергии протона. электрохимический градиент выделяется в виде тепла.[17] Процесс опосредуется протонным каналом, который называется термогенин, или же UCP1.[44] Термогенин в основном содержится в коричневая жировая ткань, или бурый жир, и отвечает за термогенез без дрожи. Коричневая жировая ткань обнаруживается у млекопитающих, и ее самый высокий уровень наблюдается в раннем возрасте и у животных, находящихся в спячке. У людей коричневая жировая ткань присутствует при рождении и уменьшается с возрастом.[44]
Хранение ионов кальция

Концентрации свободного кальция в клетке могут регулировать множество реакций и важны для преобразование сигнала в камере. Митохондрии могут временно хранить кальций, способствующий гомеостазу кальция в клетке.[45][46] Их способность быстро усваивать кальций для последующего высвобождения делает их хорошими «цитозольными буферами» для кальция.[47][48][49] Эндоплазматический ретикулум (ЭР) является наиболее важным местом хранения кальция,[50] и существует значительная взаимосвязь между митохондрией и ER в отношении кальция.[51] Кальций попадает в матрица посредством митохондриальный унипортер кальция на внутренняя митохондриальная мембрана.[52] Это в первую очередь обусловлено митохондриальной мембранный потенциал.[46] Высвобождение этого кальция обратно внутрь клетки может происходить через обменный белок натрий-кальций или через пути «кальций-индуцированного высвобождения кальция».[52] Это может вызвать скачки кальция или волны кальция с большими изменениями в мембранный потенциал. Они могут активировать серию вторая система обмена сообщениями белки, которые могут координировать процессы, такие как выпуск нейротрансмиттера в нервных клетках и высвобождение гормоны в эндокринных клетках.[53]
Ca2+ приток в митохондриальный матрикс недавно был задействован как механизм регуляции дыхательной биоэнергетика позволяя электрохимическому потенциалу на мембране временно "пульсировать" от ΔΨ-доминирующего к pH-доминирующему, облегчая снижение окислительный стресс.[54] В нейронах сопутствующее увеличение цитозольного и митохондриального кальция синхронизирует активность нейронов с митохондриальным энергетическим метаболизмом. Уровни кальция в митохондриальном матриксе могут достигать десятков микромолярных уровней, что необходимо для активации изоцитратдегидрогеназа, один из ключевых регуляторных ферментов Цикл Кребса.[55]
Регулирование клеточной пролиферации
Была исследована взаимосвязь между клеточной пролиферацией и митохондриями. Опухолевым клеткам требуется достаточное количество АТФ для синтеза биоактивных соединений, таких как липиды, белки, и нуклеотиды для быстрого распространения.[56] Большая часть АТФ в опухолевых клетках генерируется через окислительного фосфорилирования путь (OxPhos).[57] Взаимодействие с OxPhos вызывает остановку клеточного цикла, что позволяет предположить, что митохондрии играют роль в пролиферации клеток.[57] Производство митохондриального АТФ также жизненно важно для деление клеток и дифференциация инфекции [58] в дополнение к основным функциям клетки, включая регулирование объема клетки, растворенные вещества концентрация, и сотовая архитектура.[59][60][61] Уровни АТФ различаются на разных стадиях клеточного цикла, что позволяет предположить, что существует взаимосвязь между содержанием АТФ и способностью клетки вступать в новый клеточный цикл.[62] Роль АТФ в основных функциях клетки заставляет клеточный цикл чувствительны к изменениям в доступности митохондриального АТФ.[62] Различия в уровнях АТФ на разных стадиях клеточного цикла подтверждают гипотезу о том, что митохондрии играют важную роль в регуляции клеточного цикла.[62] Хотя конкретные механизмы между митохондриями и регуляцией клеточного цикла не совсем понятны, исследования показали, что контрольные точки клеточного цикла с низкой энергией контролируют энергетическую способность перед переходом к следующему раунду клеточного деления.[9]
Дополнительные функции
Митохондрии играют центральную роль во многих других метаболический задачи, такие как:
- Передача сигналов через митохондрии активные формы кислорода[63]
- Регулирование мембранный потенциал[17]
- Апоптоз -программированная гибель клеток[64]
- Передача сигналов кальция (включая апоптоз, вызванный кальцием)[65]
- Регулирование сотовой метаболизм[9]
- Определенный гем реакции синтеза[66] (смотрите также: порфирин )
- Стероидный препарат синтез.[47]
- Гормональная сигнализация [67] Митохондрии чувствительны и реагируют на гормоны, отчасти за счет действия митохондриальных рецепторов эстрогена (mtER). Эти рецепторы были обнаружены в различных тканях и типах клеток, включая мозг. [68] и сердце [69]
- Иммунная сигнализация [70]
- Митохондрии нейронов также способствуют контролю качества клеток, сообщая о статусе нейронов микроглии через специализированные соматические соединения.[71]
Некоторые митохондриальные функции выполняются только в определенных типах клеток. Например, митохондрии в печень клетки содержат ферменты, которые позволяют им выводить токсины аммиак, продукт жизнедеятельности белкового обмена. Мутация в генах, регулирующих любую из этих функций, может привести к митохондриальные заболевания.
Организация и распространение

Митохондрии (и связанные с ними структуры) встречаются во всех эукариоты (кроме двух - Оксимонад Monocercomonoides и Henneguya salminicola).[3][4][5][72] Хотя обычно изображаются как бобовые структуры, они образуют очень динамичную сеть в большинстве ячеек, где они постоянно подвергаются воздействию. деление и слияние. Популяция всех митохондрий данной клетки составляет хондриом.[73] Митохондрии различаются по количеству и расположению в зависимости от типа клеток. Одна митохондрия часто встречается у одноклеточных организмов, в то время как клетки печени человека содержат около 1000–2000 митохондрий на клетку, что составляет 1/5 объема клетки.[16] Митохондриальное содержание подобных клеток может существенно различаться по размеру и мембранному потенциалу.[74] с различиями, возникающими из источников, включая неравномерное разделение при делениях клеток, что приводит к внешние различия в АТФ уровни и последующие клеточные процессы.[75] Митохондрии можно найти между миофибриллы из мышца или обернутый вокруг сперма жгутик.[16] Часто они образуют сложную трехмерную разветвленную сеть внутри ячейки с цитоскелет. Связь с цитоскелет определяет форму митохондрий, что также может влиять на функцию:[76] различные структуры митохондриальной сети могут дать населению различные физические, химические и сигнальные преимущества или недостатки.[77] Митохондрии в клетках всегда распределяются по микротрубочкам, и распределение этих органелл также коррелирует с эндоплазматический ретикулум.[78] Последние данные свидетельствуют о том, что виментин, один из компонентов цитоскелета, также важен для ассоциации с цитоскелетом.[79]
Митохондриально-ассоциированная мембрана ER (MAM)
Митохондриально-ассоциированная мембрана ER (MAM) - еще один структурный элемент, который все больше признается за его критическую роль в клеточной физиологии и гомеостаз. Когда-то считавшиеся технической загвоздкой в методах клеточного фракционирования, предполагаемые контаминанты ER везикул, которые неизменно появлялись в митохондриальной фракции, были повторно идентифицированы как мембранные структуры, происходящие из MAM - интерфейса между митохондриями и ER.[80] Физическая связь между этими двумя органеллами ранее наблюдалась на электронных микрофотографиях, а позднее была исследована с помощью флуоресцентная микроскопия.[80] Такие исследования показывают, что в MAM, который может составлять до 20% внешней митохондриальной мембраны, ER и митохондрии разделены всего лишь на 10-25 нм и удерживаются вместе с помощью белковых связывающих комплексов.[80][28][81]
Очищенный МАМ от субклеточного фракционирования обогащен ферментами, участвующими в обмене фосфолипидов, в дополнение к каналам, связанным с Ca2+ сигнализация.[80][81] Эти намеки на выдающуюся роль MAM в регуляции клеточных липидных запасов и передачи сигналов были подтверждены со значительными последствиями для митохондриально-ассоциированных клеточных явлений, как обсуждается ниже. MAM не только предоставил понимание механистической основы, лежащей в основе таких физиологических процессов, как внутренний апоптоз и распространение кальциевых сигналов, но также способствует более точному представлению о митохондриях. Хотя его часто рассматривают как статические, изолированные `` электростанции '', используемые для клеточного метаболизма в результате древнего эндосимбиотического события, эволюция MAM подчеркивает степень интеграции митохондрий в общую физиологию клетки с тесным физическим и функциональным взаимодействием с эндомембранной системой.
Перенос фосфолипидов
МАМ обогащен ферментами, участвующими в биосинтезе липидов, такими как фосфатидилсеринсинтаза на поверхности ЭПР и фосфатидилсерин декарбоксилаза на поверхности митохондрий.[82][83] Поскольку митохондрии - это динамические органеллы, постоянно подвергающиеся деление и слияние событиях, они требуют постоянного и хорошо регулируемого поступления фосфолипидов для целостности мембраны.[84][85] Но митохондрии - это не только место назначения фосфолипидов, синтез которых они завершают; скорее, эта органелла также играет роль в перемещении между органеллами промежуточных продуктов и продуктов путей биосинтеза фосфолипидов, метаболизма церамидов и холестерина и анаболизма гликосфинголипидов.[83][85]
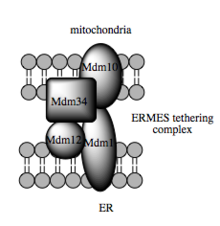
Такая пропускная способность зависит от MAM, который, как было показано, облегчает перенос липидных промежуточных соединений между органеллами.[82] В противоположность стандартному везикулярному механизму переноса липидов, данные указывают на то, что физическая близость ER и митохондриальных мембран в MAM делает возможным переключение липидов между противоположными бислоями.[85] Несмотря на этот необычный и, казалось бы, энергетически невыгодный механизм, такой транспорт не требует АТФ.[85] Вместо этого было показано, что в дрожжах он зависит от мультипротеин связывающая структура, называемая структурой встречи ER-митохондрий, или ERMES, хотя остается неясным, напрямую ли эта структура опосредует перенос липидов или требуется для удержания мембран в достаточно близком расстоянии, чтобы снизить энергетический барьер для липид листать.[85][86]
МАМ может также быть частью секреторного пути в дополнение к его роли во внутриклеточном перемещении липидов. В частности, MAM, по-видимому, является промежуточным пунктом назначения между грубым ER и Golgi на пути, который ведет к липопротеины очень низкой плотности, или VLDL, сборка и секреция.[83][87] Таким образом, МАМ служит важнейшим центром метаболизма и транспортировки липидов.
Сигнализация кальция
Критическая роль ER в передаче сигналов кальция была признана еще до того, как такая роль митохондрий получила широкое признание, отчасти из-за низкого сродства Ca2+ каналы, локализованные на внешней митохондриальной мембране, по-видимому, противоречат предполагаемой способности этой органеллы реагировать на изменения внутриклеточного Ca2+ поток.[80][50] Но присутствие МАМ разрешает это очевидное противоречие: тесная физическая ассоциация между двумя органеллами приводит к образованию Ca2+ микродомены в точках контакта, которые способствуют эффективному Ca2+ передача из ER в митохондрии.[80] Передача происходит в ответ на так называемый Ca2+ затяжки », вызванные спонтанной кластеризацией и активацией IP3R, каноническая мембрана ЭР Ca2+ канал.[80][28]
Судьба этих затяжек - в частности, остаются ли они ограниченными изолированными местами или интегрированы в Ca2+ волны для распространения по клетке - в значительной степени определяется динамикой МАМ. Хотя обратный захват Ca2+ ЭР (одновременно с его высвобождением) модулирует интенсивность затяжек, таким образом изолируя митохондрии до определенной степени от высокого содержания Ca2+ MAM часто служит брандмауэром, который, по сути, буферизует Ca2+ затяжки, действуя как сток, в который могут попадать свободные ионы, выпущенные в цитозоль.[80][88][89] Это Ca2+ туннелирование происходит через низкоаффинный Ca2+ рецептор VDAC1, который, как недавно было показано, физически привязанный к кластерам IP3R на мембране ER и обогащен на MAM.[80][28][90] Способность митохондрий служить Са2+ сток является результатом электрохимического градиента, генерируемого во время окислительного фосфорилирования, что делает туннелирование катиона экзэргоническим процессом.[90] Нормальный, умеренный приток кальция из цитозоля в митохондриальный матрикс вызывает временную деполяризацию, которая корректируется путем откачки протонов.
Но передача Са2+ не является однонаправленным; скорее, это улица с двусторонним движением.[50] Свойства Ca2+ насос SERCA и канал IP3R, присутствующий на мембране ER, облегчают регулирование обратной связи, координируемое функцией MAM. В частности, клиренс Ca2+ МАМ позволяет пространственно-временные паттерны Са2+ сигнализация, потому что Ca2+ изменяет активность IP3R двухфазным образом.[80] SERCA также зависит от митохондриальной обратной связи: поглощение Ca2+ МАМ стимулирует выработку АТФ, обеспечивая тем самым энергию, которая позволяет SERCA перезагружать ЭР Са2+ для продолжения Ca2+ отток в МАМ.[88][90] Таким образом, МАМ не является пассивным буфером для Ca2+ затяжки; скорее это помогает модулировать дальнейшее Ca2+ передача сигналов через петли обратной связи, которые влияют на динамику ER.
Регулирование ER высвобождения Ca2+ на МАМ особенно критично, потому что только определенное окно Са2+ поглощение поддерживает митохондрии и, следовательно, клетку в гомеостазе. Достаточное количество кальция в органеллах2+ Передача сигналов необходима для стимуляции метаболизма путем активации ферментов дегидрогеназы, критических для прохождения цикла лимонной кислоты.[91][92] Однако однажды Ca2+ передача сигналов в митохондриях преодолевает определенный порог, она стимулирует внутренний путь апоптоза частично за счет коллапса потенциала митохондриальной мембраны, необходимого для метаболизма.[80] Исследования, изучающие роль про- и антиапоптотических факторов, поддерживают эту модель; например, было показано, что антиапоптотический фактор Bcl-2 взаимодействует с IP3R для снижения содержания Ca2+ заполнение ER, что приводит к уменьшению оттока в MAM и предотвращению коллапса потенциала митохондриальной мембраны после постапоптотических стимулов.[80] Учитывая необходимость такого тонкого регулирования Ca2+ передачи сигналов, возможно, неудивительно, что дисрегуляция митохондриального Ca2+ был вовлечен в несколько нейродегенеративных заболеваний, в то время как каталог опухолевых супрессоров включает несколько, которые обогащены МАМ.[90]
Молекулярная основа привязки
Последние достижения в идентификации привязи между митохондриальной и ER мембранами предполагают, что функция каркаса вовлеченных молекулярных элементов вторична по отношению к другим, неструктурным функциям. У дрожжей ERMES, мультибелковый комплекс взаимодействующих ER- и митохондриально-резидентных мембранных белков, необходим для переноса липидов в MAM и иллюстрирует этот принцип. Один из его компонентов, например, также является составной частью белкового комплекса, необходимого для встраивания трансмембранных бета-стволовых белков в липидный бислой.[85] Однако гомолог комплекса ERMES еще не идентифицированы в клетках млекопитающих. Другие белки, участвующие в каркасе, также имеют функции, независимые от структурного связывания в МАМ; например, митофузины, резиденты ER и митохондрии, образуют гетерокомплексы, которые регулируют количество сайтов межорганических контактов, хотя митофузины были впервые идентифицированы из-за их роли в деление и слияние события между отдельными митохондриями.[80] Глюкоза Родственный белок 75 (grp75) является еще одним белком с двойной функцией. В дополнение к матричному пулу grp75, часть служит шапероном, который физически связывает митохондрии и ER Ca2+ каналы VDAC и IP3R для эффективного Ca2+ передача в МАМ.[80][28] Другой потенциальный трос Сигма-1Р, неопиоидный рецептор, чья стабилизация резидентного в ER IP3R может сохранять связь в МАМ во время реакции на метаболический стресс.[93][94]
Перспектива
МАМ является важнейшим центром передачи сигналов, метаболизма и трафика в клетке, который позволяет интегрировать ЭР и физиологию митохондрий. Связь между этими органеллами является не только структурной, но и функциональной, и имеет решающее значение для общей клеточной физиологии и гомеостаз. Таким образом, MAM предлагает взгляд на митохондрии, который расходится с традиционным взглядом на эту органеллу как на статическую изолированную единицу, приспособленную для ее метаболической способности клеткой.[95] Напротив, этот интерфейс митохондрий-ER подчеркивает интеграцию митохондрий, продуктов эндосимбиотического события, в различные клеточные процессы. Недавно было также показано, что митохондрии и MAM-ы в нейронах прикреплены к специализированным участкам межклеточной коммуникации (так называемые соматические соединения). Микроглиальный процессы контролируют и защищают функции нейронов на этих участках, и предполагается, что MAM-ы играют важную роль в этом типе контроля качества клеток.[71]
Происхождение и эволюция
Существует две гипотезы происхождения митохондрий: эндосимбиотический и автогенный. Эндосимбиотическая гипотеза предполагает, что митохондрии изначально были прокариотический клетки, способные реализовывать окислительные механизмы, недоступные для эукариотических клеток; они становятся эндосимбионты живущие внутри эукариота.[96] Согласно автогенной гипотезе, митохондрии возникли в результате отщепления части ДНК от ядра эукариотической клетки во время расхождения с прокариотами; эта часть ДНК была бы заключена в мембраны, которые не могли бы пересечься белки. Поскольку митохондрии имеют много общего с бактерии, эндосимбиотическая гипотеза получила большее распространение.[96][97]
А митохондрия содержит ДНК, который организован в виде нескольких копий одного, обычно круговой хромосома. Эта митохондриальная хромосома содержит гены редокс белки, например, дыхательной цепи. В Гипотеза CoRR предполагает, что это совместное расположение требуется для регуляции окислительно-восстановительного потенциала. Митохондриальный геном коды для некоторых РНК из рибосомы, а 22 тРНК необходимо для перевода мРНК в белок. Круглая структура также встречается у прокариот. Протомитохондрия, вероятно, была тесно связана с Риккетсия.[98][99] Однако точное родство предка митохондрий с альфапротеобактерии и вопрос о том, образовалась ли митохондрия одновременно или после ядра, остается спорным.[100] Например, было высказано предположение, что SAR11 клады бактерий имеет относительно недавнего общего предка с митохондриями,[101] пока филогеномный анализы показывают, что митохондрии произошли от протеобактерии родословная, которая тесно связана или является членом альфапротеобактерии.[102][103]
| Схема филогении рибосомных РНК Alphaproteobacteria | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кладограмма Rickettsidae была выведена Ферла. и другие. [104] из сравнения 16S + 23S последовательности рибосомной РНК. |
Рибосомы, кодируемые митохондриальной ДНК, похожи на рибосомы бактерий по размеру и структуре.[105] Они очень похожи на бактериальные 70S рибосома, а не 80-е годы цитоплазматический рибосомы, которые кодируются ядерный ДНК.
В эндосимбиотический связь митохондрий с их клетками-хозяевами была популяризирована Линн Маргулис.[106] В эндосимбиотическая гипотеза предполагает, что митохондрии произошли от бактерий, которые каким-то образом выжили эндоцитоз другой ячейкой, и стал частью цитоплазма. Способность этих бактерий проводить дыхание в хозяйских клетках, которые полагались на гликолиз и ферментация дало бы значительное эволюционное преимущество. Эти симбиотические отношения, вероятно, возникли от 1,7 до 2 миллиардов лет назад.[107][108]Некоторые группы одноклеточных эукариот имеют только рудиментарные митохондрии или производные структуры: микроспоридианы, метамонады, и архамебы.[109] Эти группы выглядят как самые примитивные эукариоты на филогенетические деревья построен с использованием рРНК информация, которая когда-то предполагала, что они появились до возникновения митохондрий. Однако теперь известно, что это артефакт притяжение длинных ветвей - они являются производными группами и сохраняют гены или органеллы, полученные из митохондрий (например, митосомы и гидрогеносомы ).[2] Гидрогеносомы, митосомы и родственные органеллы, обнаруженные в некоторых лорицифера (например, Спинолорикус )[110][111] и миксозоа (например, Henneguya zschokkei ) вместе классифицируются как MRO, органеллы, связанные с митохондриями.[112][113]
Monocercomonoides по-видимому, полностью утратили свои митохондрии, и, по крайней мере, некоторые из митохондриальных функций, похоже, теперь выполняются цитоплазматическими белками.[114]
Геном

Митохондрии содержат собственный геном. В человек митохондриальный геном это циркуляр ДНК молекула около 16килобазы.[115] Он кодирует 37 генов: 13 для подразделения респираторных комплексов I, III, IV и V, 22 для митохондриальных тРНК (для 20 стандартных аминокислот плюс дополнительный ген лейцина и серина) и 2 для рРНК.[115] Одна митохондрия может содержать от двух до десяти копий своей ДНК.[116]
Как и у прокариот, здесь очень высокая доля кодирующей ДНК и отсутствие повторов. Митохондриальные гены записано как мультигенные транскрипты, которые расщепляются и полиаденилированный давать зрелые мРНК. Большинство белков, необходимых для митохондриальной функции, кодируются генами в ядро клетки и соответствующие белки импортируются в митохондрии.[117] Точное количество генов, кодируемых ядром и митохондриальный геном отличается между видами. Большинство митохондриальных геномов имеют круглую форму.[118] В целом в митохондриальной ДНК отсутствует интроны, как в случае митохондриального генома человека;[117] однако интроны наблюдались в некоторых митохондриальных ДНК эукариот,[119] такой как у дрожжи[120] и протисты,[121] включая Диктиостелиум дискоидеум.[122] Между областями, кодирующими белок, присутствуют тРНК. Гены митохондриальной тРНК имеют последовательности, отличные от ядерных тРНК, но двойники митохондриальных тРНК были обнаружены в ядерных хромосомах с высоким сходством последовательностей.[123]
У животных митохондриальный геном обычно представляет собой одну кольцевую хромосому длиной примерно 16 т.п.н. и содержит 37 генов. Гены, хотя и являются высококонсервативными, могут различаться по местоположению. Любопытно, что у вшей человеческого тела такой закономерности нет (Pediculus humanus ). Вместо этого этот митохондриальный геном организован в 18 миникольцевых хромосом, каждая из которых имеет длину 3–4 т.п.н. и имеет от одного до трех генов.[124] Этот паттерн также встречается в других сосущие вши, но не в жевательные вши. Было показано, что между минихромосомами происходит рекомбинация.
Альтернативный генетический код
| Организм | Кодон | Стандарт | Митохондрии |
|---|---|---|---|
| Млекопитающие | AGA, AGG | Аргинин | Стоп-кодон |
| Беспозвоночные | AGA, AGG | Аргинин | Серин |
| Грибы | CUA | Лейцин | Треонин |
| Все вышеперечисленное | AUA | Изолейцин | Метионин |
| UGA | Стоп-кодон | Триптофан |
Хотя небольшие вариации стандартного генетического кода были предсказаны ранее,[125] ничего не было обнаружено до 1979 года, когда исследователи, изучавшие митохондриальные гены человека определили, что они использовали альтернативный код.[126] Однако митохондрии многих других эукариот, включая большинство растений, используют стандартный код.[127] С тех пор было обнаружено много небольших вариантов,[128] включая различные альтернативные митохондриальные коды.[129] Кроме того, все кодоны AUA, AUC и AUU являются допустимыми стартовыми кодонами.
Некоторые из этих различий следует рассматривать как псевдоизменения генетического кода из-за феномена Редактирование РНК, что часто встречается в митохондриях. Считалось, что у высших растений CGG кодирует триптофан и нет аргинин; однако было обнаружено, что кодон в процессированной РНК является кодоном UGG, что соответствует стандарту генетический код для триптофана.[130] Следует отметить, что генетический код митохондрий членистоногих претерпел параллельную эволюцию внутри филума, при этом некоторые организмы однозначно транслируют AGG в лизин.[131]
Репликация и наследование
Митохондрии делятся на двойное деление, похожие на бактерии.[132] Регуляция этого деления у эукариот разная. У многих одноклеточных эукариот их рост и деление связаны с клеточный цикл. Например, отдельная митохондрия может делиться синхронно с ядром. Этот процесс деления и сегрегации необходимо строго контролировать, чтобы каждая дочерняя клетка получила по крайней мере одну митохондрию. У других эукариот (например, у млекопитающих) митохондрии могут реплицировать свою ДНК и делиться в основном в ответ на энергетические потребности клетки, а не в фазе клеточного цикла. Когда потребности клетки в энергии высоки, митохондрии растут и делятся. Когда потребление энергии низкое, митохондрии разрушаются или становятся неактивными. В таких примерах митохондрии, по-видимому, случайным образом распределяются по дочерним клеткам во время деления цитоплазма. Митохондриальная динамика, баланс между митохондриальное слияние и деление, является важным фактором при патологиях, связанных с несколькими болезненными состояниями.[133]
Гипотеза бинарного деления митохондрий опиралась на визуализацию с помощью флуоресцентной микроскопии и обычных просвечивающая электронная микроскопия (ТЕМ). Разрешение флуоресцентной микроскопии (~ 200 нм) недостаточно для различения структурных деталей, таких как двойная митохондриальная мембрана в митохондриальном делении, или даже для различения отдельных митохондрий, когда несколько митохондрий расположены близко друг к другу. Обычный ТЕА также имеет некоторые технические ограничения.[который? ] при проверке деления митохондрий. Криоэлектронная томография недавно был использован для визуализации деления митохондрий в замороженных гидратированных интактных клетках. Выяснилось, что митохондрии делятся почкованием.[134]
Митохондриальные гены человека наследуются только от матери. У людей, когда яйцеклетка оплодотворяется спермой, митохондриями и, следовательно, митохондриальная ДНК, как правило, происходит только из яйцеклетки. Митохондрии сперматозоидов входят в яйцеклетку, но не передают эмбриону генетическую информацию.[135] Вместо этого отцовские митохондрии отмечены убиквитин чтобы выбрать их для последующего уничтожения внутри эмбрион.[136] Яйцеклетка содержит относительно мало митохондрий, но эти митохондрии делятся, чтобы заселить клетки взрослого организма. Этот режим наблюдается у большинства организмов, включая большинство животных. Однако митохондрии у некоторых видов иногда могут передаваться по отцовской линии. Это норма среди некоторых хвойный растения, хотя и не в сосны и тис.[137] За Митилиды отцовское наследование происходит только у самцов данного вида.[138][139][140] Было высказано предположение, что это происходит в очень низком уровне у людей.[141]
Однородительское наследование приводит к небольшой возможности для генетическая рекомбинация между разными линиями митохондрий, хотя одна митохондрия может содержать 2–10 копий своей ДНК.[116] То, что действительно происходит, поддерживает генетическую целостность, а не разнообразие. Однако есть исследования, показывающие доказательства рекомбинации в митохондриальной ДНК. Понятно, что ферменты, необходимые для рекомбинации, присутствуют в клетках млекопитающих.[142] Кроме того, данные свидетельствуют о том, что митохондрии животных могут подвергаться рекомбинации.[143] Данные о людях более противоречивые, хотя существуют косвенные доказательства рекомбинации.[144][145]
Можно ожидать, что объекты, подвергающиеся однопородному наследованию и практически не имеющие рекомбинации, будут подвергаться Трещотка Мюллера, накопление вредных мутаций до потери функциональности. Популяции животных митохондрий избегают этого накопления в процессе развития, известном как узкое место мтДНК. Эксплойты узких мест случайные процессы в клетке для увеличения межклеточной изменчивости в мутантный груз по мере развития организма: одна яйцеклетка с некоторой долей мутантной мтДНК, таким образом, дает эмбрион, в котором разные клетки имеют разные мутантные нагрузки. Затем отбор на клеточном уровне может действовать для удаления этих клеток с большим количеством мутантной мтДНК, что приводит к стабилизации или снижению мутантной нагрузки между поколениями. Обсуждается механизм, лежащий в основе узкого места,[146][147][148] с недавним математическим и экспериментальным метаисследованием, предоставившим доказательства комбинации случайного разделения мтДНК при делениях клетки и случайного оборота молекул мтДНК внутри клетки.[149]
Ремонт ДНК
Митохондрии могут восстанавливать окислительный Повреждение ДНК механизмами, аналогичными тем, которые происходят в ядро клетки. Белки, используемые в мтДНК ремонт кодируется ядерным гены, и перемещаются в митохондрии. В Ремонт ДНК пути в митохондриях млекопитающих включают базовая эксцизионная пластика, восстановление двухцепочечных разрывов, прямое обращение и ремонт несоответствия.[150][151] Также повреждения ДНК могут быть обойдены, а не восстановлены с помощью синтеза трансформации.
Из нескольких Ремонт ДНК процесс в митохондриях, базовая эксцизионная пластика путь был наиболее всесторонне изучен.[151] Эксцизионная репарация оснований осуществляется последовательностью стадий, катализируемых ферментативными методами, которые включают распознавание и удаление поврежденного основания ДНК, удаление образовавшегося абазического сайта, обработку концов, заполнение пробелов и лигирование. Обычное повреждение мтДНК, которое восстанавливается путем эксцизионной репарации оснований, - это 8-оксогуанин производится окислением гуанин.[152]
Двухрядные разрывы можно устранить с помощью гомологичный рекомбинационный ремонт в мтДНК млекопитающих[153] и мтДНК растений.[154] Двухцепочечные разрывы в мтДНК также можно исправить с помощью соединение концов, опосредованное микрогомологией.[155] Хотя есть доказательства процессов репарации прямой реверсии и репарации ошибочных спариваний в мтДНК, эти процессы недостаточно охарактеризованы.[151]
Отсутствие митохондриальной ДНК
Некоторые организмы полностью утратили митохондриальную ДНК. В этих случаях гены, кодируемые митохондриальной ДНК, были потеряны или перенесены в ядро.[115] Криптоспоридиум, имеют митохондрии, в которых отсутствует какая-либо ДНК, предположительно потому, что все их гены были потеряны или перенесены.[156] В Криптоспоридиуммитохондрии имеют измененный АТФ система генерации, которая делает паразита устойчивым ко многим классическим митохондриальным ингибиторы Такие как цианид, азид, и атоваквон.[156] Митохондрии, лишенные собственной ДНК, были обнаружены у морских паразитов. динофлагеллята из рода Амебофира. Этот микроорганизм, A. cerati, имеет функциональные митохондрии, в которых отсутствует геном.[157] У родственных видов в митохондриальном геноме все еще есть три гена, но в A. cerati только один митохондриальный ген - цитохром с оксидаза I ген (cox1) - обнаружен, и он перекочевал в геном ядра.[158]
Популяционно-генетические исследования
Практически полное отсутствие генетическая рекомбинация в митохондриальной ДНК делает его полезным источником информации для изучения популяционная генетика и эволюционная биология.[159] Потому что вся митохондриальная ДНК наследуется как единое целое, или гаплотип, отношения между митохондриальной ДНК разных людей можно представить как генное дерево. Паттерны в этих генных деревьях можно использовать для вывода эволюционной истории популяций. Классический пример этого - в эволюционная генетика человека, где молекулярные часы может использоваться для указания недавней даты для митохондриальная Ева.[160][161] Это часто интерпретируется как сильная поддержка недавней современной человеческой экспансии. из Африки.[162] Другой пример человека - секвенирование митохондриальной ДНК из Неандерталец кости. Относительно большое эволюционное расстояние между последовательностями митохондриальной ДНК неандертальцев и живых людей было интерпретировано как доказательство отсутствия межпородного скрещивания между неандертальцами и современными людьми.[163]
Однако митохондриальная ДНК отражает только историю самок в популяции. Частично это можно преодолеть за счет использования отцовских генетических последовательностей, таких как не рекомбинирование регион Y-хромосома.[162]
Недавние измерения молекулярные часы для митохондриальной ДНК[164] сообщили о 1 мутации каждые 7884 года, относящейся к последнему общему предку людей и обезьян, что согласуется с оценками частоты мутаций аутосомной ДНК (10−8 на базу на поколение).[165]
Дисфункция и болезнь
Митохондриальные заболевания
Повреждение и последующая дисфункция митохондрий является важным фактором ряда заболеваний человека из-за их влияния на метаболизм клеток. Митохондриальные расстройства часто проявляются в виде неврологических расстройств, в том числе аутизм.[15] Они также могут проявляться как миопатия, сахарный диабет, несколько эндокринопатия и множество других системных расстройств.[166] Заболевания, вызванные мутацией в мтДНК, включают: Синдром Кирнса – Сайра, Синдром MELAS и Наследственная оптическая нейропатия Лебера.[167] В подавляющем большинстве случаев эти заболевания передаются от женщины к ее детям, поскольку зигота получает свои митохондрии и, следовательно, свою мтДНК из яйцеклетки. Заболевания, такие как синдром Кернса-Сейра, Синдром Пирсона, и прогрессирующая наружная офтальмоплегия считается, что это связано с крупномасштабными перестройками мтДНК, тогда как другие заболевания, такие как синдром MELAS, наследственная оптическая нейропатия Лебера, Синдром MERRF, а другие связаны с точечные мутации в мтДНК.[166]
При других заболеваниях дефекты ядерных генов приводят к нарушению функции митохондриальных белков. Так обстоит дело в Атаксия Фридрейха, наследственная спастическая параплегия, и Болезнь Вильсона.[168] Эти заболевания передаются по наследству отношения доминирования, как и большинство других генетических заболеваний. Различные расстройства могут быть вызваны ядерными мутациями ферментов окислительного фосфорилирования, такими как Коэнзим Q10 дефицит и Синдром Барта.[166] Влияние окружающей среды может взаимодействовать с наследственной предрасположенностью и вызывать митохондриальные заболевания. Например, может быть связь между пестицид воздействие и более позднее начало болезнь Паркинсона.[169][170] Другие патологии с этиологией, связанной с дисфункцией митохондрий, включают: шизофрения, биполярное расстройство, слабоумие, Болезнь Альцгеймера,[171][172] Болезнь Паркинсона, эпилепсия, Инсульт, сердечно-сосудистые заболевания, Синдром хронической усталости, пигментный ретинит, и сахарный диабет.[173][174]
Окислительный стресс, опосредованный митохондриями, играет роль в кардиомиопатии в диабетики 2 типа. Повышенная доставка жирных кислот к сердцу увеличивает поглощение жирных кислот кардиомиоцитами, что приводит к усилению окисления жирных кислот в этих клетках. Этот процесс увеличивает количество восстанавливающих эквивалентов, доступных электронной транспортной цепи митохондрий, в конечном итоге увеличивая производство активных форм кислорода (АФК). ROS увеличивается разобщение белков (UCP) и усиливают утечку протонов через транслокатор адениновых нуклеотидов (ANT), комбинация которых разъединяет митохондрии. Затем разобщение увеличивает потребление кислорода митохондриями, усугубляя усиление окисления жирных кислот. Это создает порочный круг разъединения; более того, даже если потребление кислорода увеличивается, синтез АТФ не увеличивается пропорционально, потому что митохондрии не связаны. Меньшая доступность АТФ в конечном итоге приводит к дефициту энергии, проявляющемуся в снижении сердечной эффективности и сократительной дисфункции. Проблема усугубляется тем, что нарушение высвобождения кальция в саркоплазматическом ретикулуме и снижение обратного захвата митохондриями ограничивают пиковые уровни цитозольного важного сигнального иона во время сокращения мышц. Снижение внутримитохондриальной концентрации кальция увеличивает активацию дегидрогеназы и синтез АТФ.Таким образом, помимо снижения синтеза АТФ из-за окисления жирных кислот, синтез АТФ нарушается также из-за плохой передачи сигналов кальция, вызывая сердечные проблемы у диабетиков.[175]
Отношение к старению
Возможна утечка высокой энергии электроны в дыхательной цепи с образованием активные формы кислорода. Считалось, что это привело к значительному окислительный стресс в митохондриях с высокой частотой мутаций митохондриальной ДНК.[176] Предполагаемые связи между старением и окислительным стрессом не новы и были предложены в 1956 г.[177] который позже был переработан в митохондриальная свободнорадикальная теория старения.[178] Считалось, что возникает порочный круг, так как окислительный стресс приводит к мутациям митохондриальной ДНК, что может привести к ферментативным нарушениям и дальнейшему окислительному стрессу.
В процессе старения в митохондриях может произойти ряд изменений.[179] В тканях пожилых людей наблюдается снижение ферментативной активности белков дыхательной цепи.[180] Однако мутированная мтДНК может быть обнаружена только примерно в 0,2% очень старых клеток.[181] Предполагается, что большие делеции в митохондриальном геноме приводят к высоким уровням окислительный стресс и гибель нейронов в болезнь Паркинсона.[182] Также было показано, что дисфункция митохондрий возникает в боковой амиотрофический склероз.[183]
Поскольку митохондрии играют ключевую роль в функции яичников, обеспечивая АТФ, необходимый для развития от зародышевого пузырька до зрелого ооцита, снижение функции митохондрий может привести к воспалению, что приведет к преждевременной недостаточности яичников и ускоренному старению яичников. Вызванная дисфункция затем отражается как в количественном (например, количество копий мтДНК и делеции мтДНК), так и в качественном (например, мутации и разрывы цепей) и окислительных повреждениях (например, дисфункциональные митохондрии из-за АФК), которые имеют значение не только для старения яичников. , но нарушают перекрестные помехи между ооцитами и кумулюсами в яичнике, связаны с генетическими нарушениями (такими как Fragile X) и могут мешать отбору эмбрионов. [184]
История
Первые наблюдения внутриклеточных структур, которые, вероятно, представляли митохондрии, были опубликованы в 1840-х годах.[185] Ричард Альтманн в 1890 г. установил их как клеточные органеллы и назвал «биобластами».[185][186] В 1898 году Карл Бенда ввел термин «митохондрии» из Греческий μίτος, митос, "поток" и χονδρίον, хондрион, «гранула».[187][185][188] Леонор Михаэлис обнаружил, что Янус зеленый может использоваться как суправитальное пятно для митохондрий в 1900 г. В 1904 г. Фридрих Мевес, сделал первое зарегистрированное наблюдение митохондрий у растений в клетках белой кувшинки, Nymphaea alba[185][189] а в 1908 г. вместе с Клавдий Рего, предположил, что они содержат белки и липиды. Бенджамин Ф. Кингсбери в 1912 году впервые связал их с клеточным дыханием, но почти исключительно на основании морфологических наблюдений.[185] В 1913 году частицы экстрактов печени морских свинок были связаны с дыханием Отто Генрих Варбург, которую он назвал «грана». Варбург и Генрих Отто Виланд, которые также постулировали аналогичный механизм частиц, не согласились с химической природой дыхания. Так было до 1925 года, когда Дэвид Кейлин обнаруженный цитохромы, что дыхательная цепь был описан.[185]
В 1939 году эксперименты с измельченными мышечными клетками показали, что клеточное дыхание с использованием одного атом кислорода может образовать два аденозинтрифосфат (АТФ), а в 1941 году концепция фосфатных связей АТФ как формы энергии в клеточном метаболизме была развита Фриц Альберт Липманн. В последующие годы механизм клеточного дыхания получил дальнейшее развитие, хотя его связь с митохондриями не была известна.[185] Вступление к фракционирование тканей к Альбер Клод позволили изолировать митохондрии от других клеточных фракций и провести биохимический анализ только на них. В 1946 году он пришел к выводу, что цитохромоксидаза и другие ферменты, ответственные за дыхательную цепь, были выделены в митохондрии. Юджин Кеннеди и Альберт Ленингер обнаружил в 1948 году, что митохондрии являются местом окислительного фосфорилирования у эукариот. Со временем метод фракционирования получил дальнейшее развитие, улучшив качество изолированных митохондрий и других элементов клеточное дыхание было определено, что они происходят в митохондриях.[185]
Первый электрон высокого разрешения микрофотографии появился в 1952 году, заменив красители Януса Грин как предпочтительный способ визуализации митохондрий.[185] Это привело к более детальному анализу структуры митохондрий, включая подтверждение того, что они были окружены мембраной. Он также показал вторую мембрану внутри митохондрий, которая складывалась гребнями, разделяющими внутреннюю камеру, и что размер и форма митохондрий варьировались от клетки к клетке.
Популярный термин «электростанция клетки» был придуман Филип Зикевиц в 1957 г.[7]
В 1967 году было обнаружено, что митохондрии содержат рибосомы.[190] В 1968 году были разработаны методы картирования митохондриальных генов, а генетическая и физическая карта митохондриальной ДНК дрожжей завершена в 1976 году.[185]
В популярной культуре
Мадлен Л'Энгль 1973 год научная фантастика Роман Ветер в дверь заметно выделяет митохондрии главного героя Чарльз Уоллес Мерри, поскольку населен существами, известными как фарандола. В романе также есть другие персонажи, путешествующие внутри одной из митохондрий Марри.
1995 год фантастика ужасов Роман Ева-паразит к Хидеаки Сена изображает митохондрии как имеющие некоторые сознание и контроль над разумом способности, пытаясь использовать их, чтобы обогнать эукариот как доминирующую форму жизни. Этот текст был адаптирован в одноименный фильм, видео игра, и продолжение видеоигры все с похожей предпосылкой.
в Звездные войны франшиза микроорганизмы называемые «мидихлорианами», дают некоторым персонажам способность чувствовать и использовать сила. Джордж Лукас, режиссер фильма 1999 г. Звёздные войны: Эпизод I - Призрачная угроза, в который были введены мидихлорианы, описал их как «расплывчатое изображение митохондрий».[191] Род бактерий Мидихлория позже был назван в честь мидихлорианов.
В результате того, что митохондрии в современном американском научное образование фраза «митохондрии - электростанция клетки» стала интернет-мем.
Смотрите также
Рекомендации
- ^ «Митохондрия | Определение митохондрии с помощью Lexico». Словари Lexico | английский.
- ^ а б Хенце К., Мартин В. (ноябрь 2003 г.). «Эволюционная биология: сущность митохондрий». Природа. 426 (6963): 127–128. Bibcode:2003Натура 426..127H. Дои:10.1038 / 426127a. PMID 14614484. S2CID 862398.
- ^ а б c Karnkowska A, Vacek V, Zubáčová Z, Treitli SC, Petrželková R, Eme L, Novák L, árský V, Barlow LD, Herman EK, Soukal P, Hroudová M, Doležal P, Stairs CW, Roger AJ, Eliáš M, Dacks , Влчек Ч, Хампл В. (май 2016 г.). «Эукариот без митохондриальной органеллы». Текущая биология. 26 (10): 1274–1284. Дои:10.1016 / j.cub.2016.03.053. PMID 27185558.
- ^ а б «Животное, которому не нужен кислород, чтобы выжить, обнаружил New Scientist». www.newscientist.com. Получено 2020-02-25.
- ^ а б Яхалом, Даяна; Аткинсон, Стивен Д .; Нойхоф, Моран; Чанг, Э. Салли; Филипп, Эрве; Картрайт, Полин; Bartholomew, Jerri L .; Хюшон, Дороти (19 февраля 2020 г.). «Книдарийный паразит лосося (Myxozoa: Henneguya) не имеет митохондриального генома». Труды Национальной академии наук. 117 (10): 5358–5363. Дои:10.1073 / pnas.1909907117. ISSN 0027-8424. ЧВК 7071853. PMID 32094163.
- ^ Кэмпбелл Н.А., Уильямсон Б., Хейден Р.Дж. (2006). Биология: изучение жизни. Бостон, Массачусетс: Пирсон Прентис Холл. ISBN 978-0-13-250882-7.
- ^ а б Зикевиц П. (1957). «Электростанция клетки». Scientific American. 197 (1): 131–140. Bibcode:1957SciAm.197a.131S. Дои:10.1038 / scientificamerican0757-131.
- ^ Wiemerslage L, Lee D (март 2016 г.). «Количественная оценка морфологии митохондрий в нейритах дофаминергических нейронов с использованием нескольких параметров». Журнал методов неврологии. 262: 56–65. Дои:10.1016 / j.jneumeth.2016.01.008. ЧВК 4775301. PMID 26777473.
- ^ а б c McBride HM, Neuspiel M, Wasiak S (июль 2006 г.). «Митохондрии: больше, чем просто электростанция». Текущая биология. 16 (14): R551–60. Дои:10.1016 / j.cub.2006.06.054. PMID 16860735. S2CID 16252290.
- ^ Валеро Т (2014). «Митохондриальный биогенез: фармакологические подходы». Текущий фармацевтический дизайн. 20 (35): 5507–9. Дои:10.2174/138161282035140911142118. HDL:10454/13341. PMID 24606795.
Таким образом, митохондриальный биогенез определяется как процесс, посредством которого клетки увеличивают свою индивидуальную митохондриальную массу [3]. ... Биогенез митохондрий происходит за счет роста и деления ранее существовавших органелл и временно координируется с событиями клеточного цикла [1].
- ^ Санчис-Гомар Ф., Гарсиа-Хименес Дж. Л., Гомес-Кабрера М.С., Паллардо Ф.В. (2014). «Митохондриальный биогенез в здоровье и болезни. Молекулярные и терапевтические подходы». Текущий фармацевтический дизайн. 20 (35): 5619–33. Дои:10.2174/1381612820666140306095106. PMID 24606801.
Митохондриальный биогенез (МБ) - важный механизм, с помощью которого клетки контролируют количество митохондрий.
- ^ Гарднер А., Болес Р.Г. (2005). «Будет ли« митохондриальная психиатрия »в будущем? Обзор». Curr. Психиатрия Rev. 1 (3): 255–271. Дои:10.2174/157340005774575064.
- ^ Леснефски Э.Дж., Могаддас С., Тандлер Б., Кернер Дж., Хоппель С.Л. (июнь 2001 г.). «Митохондриальная дисфункция при сердечных заболеваниях: ишемия - реперфузия, старение и сердечная недостаточность». Журнал молекулярной и клеточной кардиологии. 33 (6): 1065–89. Дои:10.1006 / jmcc.2001.1378. PMID 11444914.
- ^ Дорн Г.В., Вега РБ, Келли Д.П. (октябрь 2015 г.). «Митохондриальный биогенез и динамика в развивающемся и больном сердце». Гены и развитие. 29 (19): 1981–91. Дои:10.1101 / gad.269894.115. ЧВК 4604339. PMID 26443844.
- ^ а б Гриффитс К.К., Леви Р.Дж. (2017). «Доказательства митохондриальной дисфункции при аутизме: биохимические связи, генетические ассоциации и механизмы, не связанные с энергией». Окислительная медицина и клеточное долголетие. 2017: 4314025. Дои:10.1155/2017/4314025. ЧВК 5467355. PMID 28630658.
- ^ а б c d е ж грамм час я j Альбертс Б., Джонсон А., Льюис Дж, Рафф М., Робертс К., Уолтер П. (2005). Молекулярная биология клетки. Нью-Йорк: Garland Publishing Inc. ISBN 978-0-8153-4105-5.
- ^ а б c d е ж грамм час я j k л Воет Д., Воет Дж. Г., Пратт К. В. (2006). Основы биохимии (2-е изд.). John Wiley and Sons, Inc., стр.547, 556. ISBN 978-0-471-21495-3.
- ^ Андерссон С.Г., Карлберг О., Канбек Б., Курланд С.Г. (январь 2003 г.). «О происхождении митохондрий: перспективы геномики». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1429): 165–77, обсуждение 177–9. Дои:10.1098 / rstb.2002.1193. ЧВК 1693097. PMID 12594925.
- ^ Тейлор С.В., Фахи Э., Чжан Б., Гленн Г.М., Варнок Д.Е., Уайли С., Мерфи А.Н., Гоше С.П., Капальди Р.А., Гибсон Б.В., Гош С.С. (март 2003 г.). «Характеристика митохондриального протеома сердца человека». Природа Биотехнологии. 21 (3): 281–6. Дои:10.1038 / nbt793. PMID 12592411. S2CID 27329521.
- ^ Чжан Дж., Ли Х, Мюллер М., Ван И, Зонг С., Дэн Н., Вондриска Т.М., Лием Д.А., Ян Джи, Корхе П., Хонда Х, Вайс Дж. Н., Апвейлер Р., Пинг П. (апрель 2008 г.). «Систематическая характеристика митохондриального протеома мышей с использованием функционально подтвержденных сердечных митохондрий». Протеомика. 8 (8): 1564–75. Дои:10.1002 / pmic.200700851. ЧВК 2799225. PMID 18348319.
- ^ Чжан Дж., Лием Д.А., Мюллер М., Ван И, Зонг С., Дэн Н., Вондриска Т.М., Корхе П., Дрюс О., Маклеллан В. Р., Хонда Х, Вайс Дж. Н., Апвейлер Р., Пинг П. (июнь 2008 г.). «Измененная протеомная биология митохондрий сердца в стрессовых условиях». Журнал протеомных исследований. 7 (6): 2204–14. Дои:10.1021 / pr070371f. ЧВК 3805274. PMID 18484766.
- ^ «Митохондрия - гораздо больше, чем преобразователь энергии». Британское общество клеточной биологии. Получено 19 августа 2013.
- ^ Blachly-Dyson E, Forte M (сентябрь 2001 г.). «Каналы VDAC». IUBMB Life. 52 (3–5): 113–8. Дои:10.1080/15216540152845902. PMID 11798022. S2CID 38314888.
- ^ Hoogenboom BW, Suda K, Engel A, Fotiadis D (июль 2007 г.). «Супрамолекулярные сборки потенциал-зависимых анионных каналов в нативной мембране». Журнал молекулярной биологии. 370 (2): 246–55. Дои:10.1016 / j.jmb.2007.04.073. PMID 17524423.
- ^ Зет К. (июнь 2010 г.). «Структура и эволюция белков внешней мембраны митохондрий бета-бочкообразной топологии». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1797 (6–7): 1292–9. Дои:10.1016 / j.bbabio.2010.04.019. PMID 20450883.
- ^ а б Herrmann JM, Neupert W (апрель 2000 г.). «Транспорт белка в митохондрии». Текущее мнение в микробиологии. 3 (2): 210–4. Дои:10.1016 / S1369-5274 (00) 00077-1. PMID 10744987.
- ^ а б Чипук JE, Bouchier-Hayes L, Green DR (август 2006 г.). «Проницаемость внешней мембраны митохондрий во время апоптоза: сценарий невинного свидетеля». Гибель клеток и дифференциация. 13 (8): 1396–1402. Дои:10.1038 / sj.cdd.4401963. PMID 16710362.
- ^ а б c d е Хаяси Т., Риццуто Р., Хайноцки Г., Су Т.П. (февраль 2009 г.). «МАМ: больше, чем просто домработница». Тенденции в клеточной биологии. 19 (2): 81–88. Дои:10.1016 / j.tcb.2008.12.002. ЧВК 2750097. PMID 19144519.
- ^ Schenkel LC, Bakovic M (январь 2014 г.). «Формирование и регуляция митохондриальных мембран». Международный журнал клеточной биологии. 2014: 709828. Дои:10.1155/2014/709828. ЧВК 3918842. PMID 24578708.
- ^ Макмиллин Дж. Б., Доухан В. (декабрь 2002 г.). «Кардиолипин и апоптоз». Biochimica et Biophysica Acta (BBA) - молекулярная и клеточная биология липидов. 1585 (2–3): 97–107. Дои:10.1016 / S1388-1981 (02) 00329-3. PMID 12531542.
- ^ Юле Р.Дж., ван дер Блик AM (2012). «Митохондриальное деление, слияние и стресс». Наука. 337 (6098): 1062–1065. Bibcode:2012Научный ... 337.1062Y. Дои:10.1126 / наука.1219855. ЧВК 4762028. PMID 22936770.
- ^ Cserép C, Pósfai B, Schwarcz AD, Dénes Á (2018). «Ультраструктура митохондрий связана с синаптическими характеристиками на участках высвобождения аксонов». eNeuro. 5 (1): ENEURO.0390–17.2018. Дои:10.1523 / ENEURO.0390-17.2018. ЧВК 5788698. PMID 29383328.
- ^ Маннелла CA (2006). «Структура и динамика крист внутренней мембраны митохондрий». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1763 (5–6): 542–548. Дои:10.1016 / j.bbamcr.2006.04.006. PMID 16730811.
- ^ Богенхаген Д.Ф. (сентябрь 2012 г.). «Структура нуклеоида митохондриальной ДНК». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов. 1819 (9–10): 914–20. Дои:10.1016 / j.bbagrm.2011.11.005. PMID 22142616.
- ^ а б Шмидт-Рор К (2020). «Кислород - высокоэнергетическая молекула, питающая сложную многоклеточную жизнь: фундаментальные поправки к традиционной биоэнергетике». СКУД Омега. 5 (5): 2221–2233. Дои:10.1021 / acsomega.9b03352. ЧВК 7016920. PMID 32064383.
- ^ Богатый пиар (декабрь 2003 г.). «Молекулярный аппарат дыхательной цепи Кейлина». Сделки Биохимического Общества. 31 (Pt 6): 1095–1105. Дои:10.1042 / BST0311095. PMID 14641005.
- ^ Стойменова М., Игамбердиев А.Ю., Гупта К.Дж., Хилл Р.Д. (июль 2007 г.). «Управляемый нитритом анаэробный синтез АТФ в митохондриях корня ячменя и риса». Planta. 226 (2): 465–474. Дои:10.1007 / s00425-007-0496-0. PMID 17333252. S2CID 8963850.
- ^ а б c d е ж Страйер Л. (1995). «Цикл лимонной кислоты». В: Биохимия. (Четвертое изд.). Нью-Йорк: W.H. Фримен и компания. С. 509–527, 569–579, 614–616, 638–641, 732–735, 739–748, 770–773. ISBN 0-7167-2009-4.
- ^ Король А, Селак М.А., Готлиб Э. (август 2006 г.). «Сукцинатдегидрогеназа и фумаратгидратаза: связь митохондриальной дисфункции и рака». Онкоген. 25 (34): 4675–4682. Дои:10.1038 / sj.onc.1209594. PMID 16892081.
- ^ Хуанг Х., Manton KG (май 2004 г.). «Роль окислительного повреждения митохондрий при старении: обзор» (PDF). Границы биологических наук. 9 (1–3): 1100–1117. Дои:10.2741/1298. PMID 14977532. S2CID 2278219.
- ^ Митчелл П., Мойл Дж. (Январь 1967 г.). «Хемиосмотическая гипотеза окислительного фосфорилирования». Природа. 213 (5072): 137–139. Bibcode:1967Натура.213..137М. Дои:10.1038 / 213137a0. PMID 4291593. S2CID 4149605.
- ^ Митчелл П. (июнь 1967). «Протонный ток в митохондриальных системах». Природа. 214 (5095): 1327–1328. Bibcode:1967Натура.214.1327М. Дои:10.1038 / 2141327a0. PMID 6056845. S2CID 4160146.
- ^ Нобелевский фонд. «Химия 1997». Получено 2007-12-16.
- ^ а б Mozo J, Emre Y, Bouillaud F, Ricquier D, Criscuolo F (ноябрь 2005 г.). «Терморегуляция: какова роль UCP у млекопитающих и птиц?». Отчеты по бионауке. 25 (3–4): 227–249. Дои:10.1007 / s10540-005-2887-4. PMID 16283555. S2CID 164450.
- ^ Сантулли Г., Се В., Рейкен С. Р., Маркс А. Р. (сентябрь 2015 г.). «Перегрузка митохондрий кальцием является ключевым фактором сердечной недостаточности». Труды Национальной академии наук Соединенных Штатов Америки. 112 (36): 11389–11394. Bibcode:2015ПНАС..11211389С. Дои:10.1073 / pnas.1513047112. ЧВК 4568687. PMID 26217001.
- ^ а б Сигел Дж. Дж., Агранофф Б. В., Фишер СК, Альберс Р. В., Улер М. Д., ред. (1999). Основы нейрохимии (6 изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-397-51820-3. Иллюстрации Лори М. Гавулич
- ^ а б Россье М.Ф. (август 2006 г.). «Т-каналы и биосинтез стероидов: в поисках связи с митохондриями». Клеточный кальций. 40 (2): 155–164. Дои:10.1016 / j.ceca.2006.04.020. PMID 16759697.
- ^ Брайтон, Коннектикут, Хант Р.М. (май 1974 г.). «Митохондриальный кальций и его роль в кальцификации. Гистохимическая локализация кальция на электронных микрофотографиях эпифизарной ростовой пластинки с K-пироантимонатом». Клиническая ортопедия и смежные исследования. 100 (5): 406–416. Дои:10.1097/00003086-197405000-00057. PMID 4134194.
- ^ Брайтон, Коннектикут, Хант, Р.М. (июль 1978 г.). «Роль митохондрий в кальцификации ростовых пластинок, как показано на рахитической модели». Журнал костной и суставной хирургии. Американский объем. 60 (5): 630–639. Дои:10.2106/00004623-197860050-00007. PMID 681381.
- ^ а б c Сантулли Г, Маркс АР (2015). «Основные роли внутриклеточных каналов высвобождения кальция в мышцах, мозге, метаболизме и старении». Современная молекулярная фармакология. 8 (2): 206–222. Дои:10.2174/1874467208666150507105105. PMID 25966694.
- ^ Пиццо П., Поццан Т. (октябрь 2007 г.). «Хореография митохондрий и эндоплазматического ретикулума: структура и сигнальная динамика». Тенденции в клеточной биологии. 17 (10): 511–517. Дои:10.1016 / j.tcb.2007.07.011. PMID 17851078.
- ^ а б Миллер Р.Дж. (1 марта 1998 г.). «Митохондрии - кракен просыпается!». Тенденции Neurosci. 21 (3): 95–97. Дои:10.1016 / S0166-2236 (97) 01206-X. PMID 9530913. S2CID 5193821.
- ^ Santulli G, Pagano G, Sardu C, Xie W, Reiken S, D'Ascia SL, Cannone M, Marziliano N, Trimarco B, Guise TA, Lacampagne A, Marks AR (май 2015 г.). «Канал высвобождения кальция RyR2 регулирует высвобождение инсулина и гомеостаз глюкозы». Журнал клинических исследований. 125 (5): 1968–1978. Дои:10.1172 / JCI79273. ЧВК 4463204. PMID 25844899.
- ^ Шварцлендер М., Логан, округ Колумбия, Джонстон И.Г., Джонс Н.С., Мейер А.Дж., Фрикер М.Д., Sweetlove LJ (март 2012 г.). «Пульсация мембранного потенциала в отдельных митохондриях: стресс-индуцированный механизм регулирования биоэнергетики дыхания у Arabidopsis». Растительная клетка. 24 (3): 1188–1201. Дои:10.1105 / tpc.112.096438. ЧВК 3336130. PMID 22395486.
- ^ Иванников М.В., Маклеод Г.Т. (июнь 2013 г.). «Уровни свободного Ca²⁺ в митохондриях и их влияние на энергетический метаболизм в двигательных нервных окончаниях дрозофилы». Биофизический журнал. 104 (11): 2353–2361. Bibcode:2013BpJ ... 104.2353I. Дои:10.1016 / j.bpj.2013.03.064. ЧВК 3672877. PMID 23746507.
- ^ Вайнберг Ф., Чандель Н.С. (октябрь 2009 г.). «Митохондриальный метаболизм и рак». Летопись Нью-Йоркской академии наук. 1177 (1): 66–73. Bibcode:2009НЯСА1177 ... 66Вт. Дои:10.1111 / j.1749-6632.2009.05039.x. PMID 19845608. S2CID 29827252.
- ^ а б Морено-Санчес Р., Родригес-Энрикес С., Марин-Эрнандес А., Сааведра Е. (март 2007 г.). «Энергетический обмен в опухолевых клетках». Журнал FEBS. 274 (6): 1393–1418. Дои:10.1111 / j.1742-4658.2007.05686.x. PMID 17302740. S2CID 7748115.
- ^ Мистри Дж. Дж., Марлейн С. Р., Мур Дж., Хельмих К., Войтович Э., Смит Дж. Г., Маколей I, Сан Y, Морфакис А., Паттерсон А., Хортон Р. Х., Дивекар Д., Моррис С. Дж., Хэстьер А., Ди Пальма Ф, Бераза Н., Боулз К.М., Рашворт С.А. (ноябрь 2019 г.). «Активация PI3K, опосредованная ROS, запускает перенос митохондрий от стромальных клеток к гемопоэтическим стволовым клеткам в ответ на инфекцию». PNAS. 116 (49): 24610–24619. Дои:10.1073 / pnas.1913278116. ЧВК 6900710. PMID 31727843.
- ^ Педерсен П.Л. (декабрь 1994 г.). «АТФ-синтаза. Машина, производящая АТФ». Текущая биология. 4 (12): 1138–1141. Дои:10.1016 / S0960-9822 (00) 00257-8. PMID 7704582. S2CID 10279742.
- ^ Паттаппа Дж., Хейвуд ХК, де Брейн Дж. Д., Ли Д. А. (октябрь 2011 г.). «Метаболизм мезенхимальных стволовых клеток человека во время пролиферации и дифференцировки». Журнал клеточной физиологии. 226 (10): 2562–2570. Дои:10.1002 / jcp.22605. PMID 21792913. S2CID 22259833.
- ^ Agarwal B (июнь 2011 г.). «Роль анионов в синтезе АТФ и его молекулярно-механистическая интерпретация». Журнал биоэнергетики и биомембран. 43 (3): 299–310. Дои:10.1007 / s10863-011-9358-3. PMID 21647635. S2CID 29715383.
- ^ а б c Sweet S, Singh G (июль 1999 г.). «Изменения митохондриальной массы, мембранного потенциала и клеточного содержания аденозинтрифосфата во время клеточного цикла лейкозных клеток человека (HL-60)». Журнал клеточной физиологии. 180 (1): 91–96. Дои:10.1002 / (SICI) 1097-4652 (199907) 180: 1 <91 :: AID-JCP10> 3.0.CO; 2-6. PMID 10362021.
- ^ Ли Х, Фанг П, Май Дж, Чой Э., Ван Х, Ян Х Ф (февраль 2013 г.). «Нацеленность на активные формы кислорода в митохондриях в качестве новой терапии воспалительных заболеваний и рака». Журнал гематологии и онкологии. 6 (19): 19. Дои:10.1186/1756-8722-6-19. ЧВК 3599349. PMID 23442817.
- ^ Зеленый DR (сентябрь 1998 г.). «Пути апоптоза: дороги к гибели». Клетка. 94 (6): 695–698. Дои:10.1016 / S0092-8674 (00) 81728-6. PMID 9753316. S2CID 16654082.
- ^ Хайноцкий Г., Чордас Г., Дас С., Гарсия-Перес К., Саотоме М., Синха Рой С., Йи М. (2006). «Передача сигналов кальция в митохондриях и гибель клеток: подходы к оценке роли захвата митохондриального Ca2 + в апоптозе». Клеточный кальций. 40 (5–6): 553–560. Дои:10.1016 / j.ceca.2006.08.016. ЧВК 2692319. PMID 17074387.
- ^ О-хама Т. (август 1997 г.). «Эволюционное рассмотрение 5-аминолевулинатсинтазы в природе». Истоки жизни и эволюция биосферы. 27 (4): 405–412. Дои:10.1023 / А: 1006583601341. PMID 9249985. S2CID 13602877.
- ^ Klinge CM (декабрь 2008 г.). «Эстрогенный контроль функции митохондрий и биогенеза». Журнал клеточной биохимии. 105 (6): 1342–1351. Дои:10.1002 / jcb.21936. ЧВК 2593138. PMID 18846505.
- ^ Альварес-Дельгадо C, Мендоса-Родригес, Калифорния, Picazo O, Cerbón M (август 2010 г.). «Различная экспрессия альфа- и бета-митохондриальных рецепторов эстрогена в стареющем мозге крысы: взаимодействие с респираторным комплексом V». Экспериментальная геронтология. 45 (7–8): 580–585. Дои:10.1016 / j.exger.2010.01.015. PMID 20096765. S2CID 30841790.
- ^ Павон Н., Мартинес-Абундис Е., Эрнандес Л., Галлардо-Перес Дж. К., Альварес-Дельгадо С., Цербон М., Перес-Торрес I, Аранда А., Чавес Е. (октябрь 2012 г.). «Половые гормоны: влияние на сердечную и митохондриальную активность после ишемии-реперфузии у взрослых крыс. Гендерные различия». Журнал стероидной биохимии и молекулярной биологии. 132 (1–2): 135–146. Дои:10.1016 / j.jsbmb.2012.05.003. PMID 22609314. S2CID 24794040.
- ^ Naffah et. аль, https://doi.org/10.1016/j.redox.2019.101255
- ^ а б Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z, et al. (Январь 2020 г.). «Микроглия контролирует и защищает функцию нейронов с помощью специализированных соматических пуринергических соединений». Наука. 367 (6477): 528–537. Bibcode:2020Sci ... 367..528C. Дои:10.1126 / science.aax6752. PMID 31831638. S2CID 209343260.
- ^ Эукариот Лямблии лямблии, например, не содержит митохондрий, но имеет митохондриально-подобный ген, что позволяет предположить, что когда-то он включал либо митохондрии, либо их эндосимбиотический предшественник Роджер А.Дж., Свэрд С.Г., Товар Дж., Кларк К.Г., Смит М.В., Гиллин Ф.Д., Согин М.Л. (январь 1998 г.). «Митохондриально-подобный ген шаперонина 60 в Giardia lamblia: свидетельство того, что на дипломонаде когда-то был эндосимбионт, родственный предку митохондрий». Труды Национальной академии наук Соединенных Штатов Америки. 95 (1): 229–234. Bibcode:1998ПНАС ... 95..229Р. Дои:10.1073 / пнас.95.1.229. ЧВК 18184. PMID 9419358.
- ^ Логан, округ Колумбия (июнь 2010 г.). «Слияние, деление и расположение митохондрий в растениях». Сделки Биохимического Общества. 38 (3): 789–95. Дои:10.1042 / bst0380789. PMID 20491666.
- ^ das Neves RP, Jones NS, Andreu L, Gupta R, Enver T., Iborra FJ (декабрь 2010 г.). Вайсман Дж.С. (ред.). «Связь изменчивости глобальной скорости транскрипции с митохондриальной изменчивостью». PLOS Биология. 8 (12): e1000560. Дои:10.1371 / journal.pbio.1000560. ЧВК 3001896. PMID 21179497.
- ^ Джонстон И.Г., Гаал Б., Невес Р.П., Энвер Т., Иборра Ф.Дж., Джонс Н.С. (2012). Haugh JM (ред.). «Митохондриальная изменчивость как источник внешнего клеточного шума». PLOS вычислительная биология. 8 (3): e1002416. arXiv:1107.4499. Bibcode:2012PLSCB ... 8E2416J. Дои:10.1371 / journal.pcbi.1002416. ЧВК 3297557. PMID 22412363.
- ^ Раппапорт Л., Оливьеро П., Сэмюэл Дж. Л. (1998). «Цитоскелет, морфология и функция митохондрий». Мол. Клетка. Биохим. 184 (1–2): 101–105. Дои:10.1023 / А: 1006843113166. PMID 9746315. S2CID 28165195.
- ^ Хойцинг Х., Джонстон И.Г., Джонс Н.С. (июнь 2015 г.). «Какова функция митохондриальных сетей? Теоретическая оценка гипотез и предложение для будущих исследований». BioEssays. 37 (6): 687–700. Дои:10.1002 / bies.201400188. ЧВК 4672710. PMID 25847815.
- ^ Солтыс Б.Дж., Гупта Р.С. (1992). «Взаимоотношения эндоплазматического ретикулума, митохондрий, промежуточных волокон и микротрубочек - исследование четырехкратного флуоресцентного мечения». Биохимия и клеточная биология. 70 (10–11): 1174–1186. Дои:10.1139 / o92-163. PMID 1363623.
- ^ Тан Х.Л., Лунг Х.Л., Ву К.С., Ле АХ, Тан Х.М., Фунг М.С. (февраль 2008 г.). «Виментин поддерживает морфологию и организацию митохондрий». Биохимический журнал. 410 (1): 141–146. Дои:10.1042 / BJ20071072. PMID 17983357.
- ^ а б c d е ж грамм час я j k л м п Риццуто Р., Марчи С., Бонора М., Агуари П., Бонони А., Де Стефани Д., Джорджи С., Лео С., Римесси А., Сивьеро Р., Зеккини Е., Пинтон П. (ноябрь 2009 г.). «Перенос Са (2+) из ЭПР в митохондрии: когда, как и почему». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1787 (11): 1342–1351. Дои:10.1016 / j.bbabio.2009.03.015. ЧВК 2730423. PMID 19341702.
- ^ а б де Брито О.М., Скоррано Л. (август 2010 г.). «Интимная связь: пространственная организация взаимоотношений эндоплазматического ретикулума и митохондрий». Журнал EMBO. 29 (16): 2715–2723. Дои:10.1038 / emboj.2010.177. ЧВК 2924651. PMID 20717141.
- ^ а б Вэнс Дж. Э., Шиао Ю. Дж. (1996).«Внутриклеточный трафик фосфолипидов: импорт фосфатидилсерина в митохондрии». Противораковые исследования. 16 (3B): 1333–1339. PMID 8694499.
- ^ а б c Lebiedzinska M, Szabadkai G, Jones AW, Duszynski J, Wieckowski MR (октябрь 2009 г.). «Взаимодействие между эндоплазматическим ретикулумом, митохондриями, плазматической мембраной и другими субклеточными органеллами». Международный журнал биохимии и клеточной биологии. 41 (10): 1805–1816. Дои:10.1016 / j.biocel.2009.02.017. PMID 19703651.
- ^ Twig G, Elorza A, Molina AJ, Mohamed H, Wikstrom JD, Walzer G, Stiles L, Haigh SE, Katz S, Las G, Alroy J, Wu M, Py BF, Yuan J, Deeney JT, Corkey BE, Shirihai OS (Январь 2008 г.). «Деление и селективное слияние регулируют сегрегацию и удаление митохондрий путем аутофагии». Журнал EMBO. 27 (2): 433–446. Дои:10.1038 / sj.emboj.7601963. ЧВК 2234339. PMID 18200046.
- ^ а б c d е ж Осман К., Фолькер Д. Р., Лангер Т. (январь 2011 г.). «Создание головы или хвоста фосфолипидов в митохондриях». Журнал клеточной биологии. 192 (1): 7–16. Дои:10.1083 / jcb.201006159. ЧВК 3019561. PMID 21220505.
- ^ Корнманн Б., Карри Е., Коллинз С. Р., Шульдинер М., Нуннари Дж., Вайсман Дж. С., Уолтер П. (июль 2009 г.). «Комплекс связывания ER-митохондрий, обнаруженный с помощью синтетического биологического экрана». Наука. 325 (5939): 477–481. Bibcode:2009Наука ... 325..477K. Дои:10.1126 / science.1175088. ЧВК 2933203. PMID 19556461.
- ^ Rusiñol AE, Cui Z, Chen MH, Vance JE (ноябрь 1994). «Уникальная мембранная фракция, ассоциированная с митохондриями из печени крысы, обладает высокой способностью к синтезу липидов и содержит секреторные белки до Гольджи, включая возникающие липопротеины». Журнал биологической химии. 269 (44): 27494–27502. PMID 7961664.
- ^ а б Копач О., Кругликов И., Пивнева Т., Войтенко Н., Федирко Н. (май 2008 г.). «Функциональная связь между рианодиновыми рецепторами, митохондриями и Са (2+) АТФазами в подчелюстных ацинарных клетках крыс». Клеточный кальций. 43 (5): 469–481. Дои:10.1016 / j.ceca.2007.08.001. PMID 17889347.
- ^ Csordás G, Hajnóczky G (апрель 2001 г.). «Сортировка кальциевых сигналов на стыках эндоплазматического ретикулума и митохондрий». Клеточный кальций. 29 (4): 249–262. Дои:10.1054 / ceca.2000.0191. PMID 11243933.
- ^ а б c d Decuypere JP, Monaco G, Bultynck G, Missiaen L, De Smedt H, Parys JB (май 2011 г.). «Связь рецептора IP (3) с митохондриями в апоптозе и аутофагии». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1813 (5): 1003–1013. Дои:10.1016 / j.bbamcr.2010.11.023. PMID 21146562.
- ^ Diercks BP, Fliegert R, Guse AH (июнь 2017 г.). «Mag-Fluo4 в Т-клетках: визуализация свободного Ca внутри органелл»2+ концентрации ». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1864 (6): 977–986. Дои:10.1016 / j.bbamcr.2016.11.026. PMID 27913206.
- ^ Хайноцкий Г., Чордаш Г., Йи М. (2011). «Старые игроки в новой роли: ассоциированные с митохондриями мембраны, VDAC и рецепторы рианодина как участники распространения кальциевого сигнала от эндоплазматического ретикулума к митохондриям». Клеточный кальций. 32 (5–6): 363–377. Дои:10.1016 / S0143416002001872. PMID 12543096.
- ^ Marriott KS, Prasad M, Thapliyal V, Bose HS (декабрь 2012 г.). «Рецептор σ-1 на мембране митохондриально-ассоциированного эндоплазматического ретикулума отвечает за регуляцию метаболизма митохондрий». Журнал фармакологии и экспериментальной терапии. 343 (3): 578–586. Дои:10.1124 / jpet.112.198168. ЧВК 3500540. PMID 22923735.
- ^ Хаяси Т., Су ТП (ноябрь 2007 г.). «Шапероны рецептора сигма-1 на интерфейсе ER-митохондрии регулируют передачу сигналов Ca (2+) и выживаемость клеток». Клетка. 131 (3): 596–610. Дои:10.1016 / j.cell.2007.08.036. PMID 17981125. S2CID 18885068.
- ^ Csordás et al., Trends Cell Biol. 2018 июл; 28 (7): 523-540. DOI: 10.1016 / j.tcb.2018.02.009. Epub 2018 24 марта.
- ^ а б Маргулис Л., Саган Д. (1986). Истоки секса. Три миллиарда лет генетической рекомбинации. Нью-Хейвен: издательство Йельского университета. стр.69–71, 87. ISBN 978-0-300-03340-3.
- ^ Мартин В.Ф., Мюллер М (2007). Происхождение митохондрий и гидрогеносом. Гейдельберг: Springer Verlag.
- ^ Емельянов В.В. (апрель 2003 г.). «Митохондриальная связь с происхождением эукариотической клетки». Европейский журнал биохимии. 270 (8): 1599–1618. Дои:10.1046 / j.1432-1033.2003.03499.x. PMID 12694174.
- ^ Мюллер М., Мартин В. (май 1999 г.). «Геном Rickettsia prowazekii и некоторые мысли о происхождении митохондрий и гидрогеносом» (PDF). BioEssays. 21 (5): 377–381. Дои:10.1002 / (sici) 1521-1878 (199905) 21: 5 <377 :: aid-bies4> 3.0.co; 2-нед.. PMID 10376009.
- ^ Серый MW, Burger G, Lang BF (март 1999 г.). «Митохондриальная эволюция». Наука. 283 (5407): 1476–1481. Bibcode:1999Научный ... 283.1476G. Дои:10.1126 / science.283.5407.1476. ЧВК 3428767. PMID 10066161.
- ^ Трэш Дж.С., Бойд А., Хаггетт М.Дж., Гроте Дж., Карини П., Йодер Р.Дж., Роббертсе Б., Спатафора Дж.В., Раппе М.С., Джованнони С.Дж. (14.06.2011). «Филогеномное свидетельство общего предка митохондрий и клады SAR11». Научные отчеты. 1 (1): 13. Bibcode:2011НатСР ... 1Е..13Т. Дои:10.1038 / srep00013. ЧВК 3216501. PMID 22355532.
- ^ Мартейн Дж., Фоссеберг Дж., Гай Л., Оффре П., Эттема Т. Дж. (Апрель 2018 г.). «Глубокое митохондриальное происхождение за пределами отобранных альфа-протеобактерий». Природа. 557 (7703): 101–105. Bibcode:2018Натура.557..101M. Дои:10.1038 / s41586-018-0059-5. PMID 29695865. S2CID 13740626.
- ^ Fan, Lu; Ву, Динфэн; Горемыкин, Вадим; Сяо, Цзин; Сюй, Янбинь; Гарг, Шрирам; Чжан, Чуаньлунь; Мартин, Уильям Ф .; Чжу, Жуйсинь (26.07.2019). «Ветвь митохондрий внутри Alphaproteobacteria». bioRxiv: 715870. Дои:10.1101/715870.
- ^ Ферла МП, Трэш Дж. К., Джованнони С. Дж., Патрик В. М. (2013). «Новые основанные на генах рРНК филогении Alphaproteobacteria дают представление об основных группах, митохондриальном происхождении и филогенетической нестабильности». PLOS One. 8 (12): e83383. Дои:10.1371 / journal.pone.0083383. ЧВК 3859672. PMID 24349502.
- ^ О'Брайен Т.В. (сентябрь 2003 г.). «Свойства митохондриальных рибосом человека». IUBMB Life. 55 (9): 505–513. Дои:10.1080/15216540310001626610. PMID 14658756. S2CID 43415449.
- ^ Саган Л. (март 1967). «О происхождении митозирующих клеток». Журнал теоретической биологии. 14 (3): 255–274. Дои:10.1016/0022-5193(67)90079-3. PMID 11541392.
- ^ Емельянов В.В. (февраль 2001 г.). «Rickettsiaceae, риккетсии-подобные эндосимбионты и происхождение митохондрий». Отчеты по бионауке. 21 (1): 1–17. Дои:10.1023 / А: 1010409415723. PMID 11508688. S2CID 10144020.
- ^ Фэн Д.Ф., Чо Г., Дулиттл Р.Ф. (ноябрь 1997 г.). «Определение времени расхождения с помощью белковых часов: обновление и переоценка». Труды Национальной академии наук Соединенных Штатов Америки. 94 (24): 13028–13033. Bibcode:1997PNAS ... 9413028F. Дои:10.1073 / пнас.94.24.13028. ЧВК 24257. PMID 9371794.
- ^ Кавальер-Смит Т. (1991). «Archamoebae: предки эукариот?». Биосистемы. 25 (1–2): 25–38. Дои:10.1016 / 0303-2647 (91) 90010-И. PMID 1854912.
- ^ Дановаро Р. (2010). «Первые метазоа, постоянно живущие в бескислородных условиях». BMC Биология. 8: 30. Дои:10.1186/1741-7007-8-30. ЧВК 2907586. PMID 20370908.
- ^ Энди Когхалн: Зоологгер: грязевое существо, живущее без кислорода., NewScientist, 7 апреля 2010 г.
- ^ Яхаломи Д., Аткинсон С.Д., Нойхоф М., Чанг Е.С., Филипп Х., Картрайт П., Бартоломью Д.Л., Хучон Д. (2020). "Книдарийный паразит лосося (Myxozoa: Хеннегуя) отсутствует митохондриальный геном ". Proc Natl Acad Sci U S A. 117 (10): 5358–5363. Дои:10.1073 / pnas.1909907117. ЧВК 7071853. PMID 32094163.
- ^ Шифлетт AM; Джонсон, П.Дж. (2010). «Органеллы, связанные с митохондриями, у эукариотических протистов». Ежегодный обзор микробиологии. 64: 409–29. Дои:10.1146 / annurev.micro.62.081307.162826. ЧВК 3208401. PMID 20528687.
- ^ Karnkowska A, Vacek V, Zubáčová Z, Treitli SC, Petrželková R, Eme L, Novák L, árský V, Barlow LD, Herman EK, Soukal P, Hroudová M, Doležal P, Stairs CW, Roger AJ, Eliáš M, Dacks , Влчек Ч, Хампл В. (май 2016 г.). «Эукариот без митохондриальной органеллы». Текущая биология. 26 (10): 1274–1284. Дои:10.1016 / j.cub.2016.03.053. PMID 27185558.
- ^ а б c Чан, округ Колумбия (июнь 2006 г.). «Митохондрии: динамические органеллы при заболеваниях, старении и развитии». Клетка. 125 (7): 1241–1252. Дои:10.1016 / j.cell.2006.06.010. PMID 16814712. S2CID 8551160.
- ^ а б Wiesner RJ, Rüegg JC, Morano I (март 1992 г.). «Подсчет целевых молекул с помощью экспоненциальной полимеразной цепной реакции: число копий митохондриальной ДНК в тканях крысы». Сообщения о биохимических и биофизических исследованиях. 183 (2): 553–559. Дои:10.1016 / 0006-291X (92) 90517-O. PMID 1550563.
- ^ а б Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брюин М.Х., Колсон А.Р., Друин Дж., Эперон И.К., Нирлих Д.П., Роу Б.А., Сэнгер Ф., Шрайер П.Х., Смит А.Дж., Стаден Р., Янг И.Г. (апрель 1981 г.) «Последовательность и организация митохондриального генома человека». Природа. 290 (5806): 457–465. Bibcode:1981Натура.290..457A. Дои:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ Фукухара Х., Сор Ф., Дрисси Р., Динуэль Н., Миякава И., Руссе С., Виола А.М. (апрель 1993 г.). «Линейные митохондриальные ДНК дрожжей: частота встречаемости и общие черты». Молекулярная и клеточная биология. 13 (4): 2309–2314. Дои:10.1128 / mcb.13.4.2309. ЧВК 359551. PMID 8455612.
- ^ Бернарди Г. (декабрь 1978 г.). «Промежуточные последовательности в митохондриальном геноме». Природа. 276 (5688): 558–559. Bibcode:1978Натура.276..558Б. Дои:10.1038 / 276558a0. PMID 214710. S2CID 4314378.
- ^ Хеббар С.К., Белчер С.М., Перлман П.С. (апрель 1992 г.). «Кодирующий матуразу интрон группы IIA митохондрий дрожжей самосплайсируется in vitro». Исследования нуклеиновых кислот. 20 (7): 1747–1754. Дои:10.1093 / nar / 20.7.1747. ЧВК 312266. PMID 1579468.
- ^ Gray MW, Lang BF, Cedergren R, Golding GB, Lemieux C, Sankoff D, Turmel M, Brossard N, Delage E, Littlejohn TG, Plante I, Rioux P, Saint-Louis D, Zhu Y, Burger G (февраль 1998 г.) . «Структура генома и содержание генов в митохондриальных ДНК протистов». Исследования нуклеиновых кислот. 26 (4): 865–878. Дои:10.1093 / nar / 26.4.865. ЧВК 147373. PMID 9461442.
- ^ Грей М.В., Ланг Б.Ф., Бургер Г. (2004). «Митохондрии простейших». Ежегодный обзор генетики. 38: 477–524. Дои:10.1146 / annurev.genet.37.110801.142526. PMID 15568984.
- ^ Telonis AG, Loher P, Kirino Y, Rigoutsos I (2014). «Ядерные и митохондриальные тРНК-двойники в геноме человека». Границы генетики. 5: 344. Дои:10.3389 / fgene.2014.00344. ЧВК 4189335. PMID 25339973.
- ^ Шао Р., Киркнесс Э. Ф., Баркер СК (май 2009 г.). «Единая митохондриальная хромосома, типичная для животных, превратилась в 18 минихромосом у человеческой вши Pediculus humanus». Геномные исследования. 19 (5): 904–912. Дои:10.1101 / гр.083188.108. ЧВК 2675979. PMID 19336451.
- ^ Крик Ф. Х., Оргель Л. Е. (1973). «Направленная панспермия» (PDF). Икар. 19 (3): 341–346. Bibcode:1973Icar ... 19..341C. Дои:10.1016/0019-1035(73)90110-3.
п. 344: Немного удивительно, что организмы с несколько разными кодами не сосуществуют.
Дальнейшее обсуждение. - ^ Баррелл Б.Г., Банкир А.Т., Друин Дж. (Ноябрь 1979 г.). «Другой генетический код в митохондриях человека». Природа. 282 (5735): 189–194. Bibcode:1979Натура.282..189Б. Дои:10.1038 / 282189a0. PMID 226894. S2CID 4335828.
- ^ Митохондриальный генетический код в дереве таксономии. NCBI
- ^ Эльзановский, Анджей и Остелл, Джим. Генетические коды. NCBI
- ^ Jukes TH, Osawa S (декабрь 1990 г.). «Генетический код митохондрий и хлоропластов». Experientia. 46 (11–12): 1117–1126. Дои:10.1007 / BF01936921. PMID 2253709. S2CID 19264964.
- ^ Хизель Р., Виссинджер Б., Шустер В., Бреннике А. (декабрь 1989 г.). «Редактирование РНК в митохондриях растений». Наука. 246 (4937): 1632–1634. Bibcode:1989Sci ... 246.1632H. Дои:10.1126 / science.2480644. PMID 2480644.
- ^ Abascal F, Posada D, Knight RD, Zardoya R (май 2006 г.). «Параллельная эволюция генетического кода в митохондриальных геномах членистоногих». PLOS Биология. 4 (5): e127. Дои:10.1371 / journal.pbio.0040127. ЧВК 1440934. PMID 16620150.
- ^ Пфайффер РФ (2012). Болезнь Паркинсона. CRC Press. п. 583. ISBN 9781439807149.
- ^ Со А. Ю., Джозеф А. М., Датта Д., Хван Дж. К., Арис Дж. П., Лиувенбург К. (август 2010 г.). «Новое понимание роли митохондрий в старении: митохондриальная динамика и многое другое». Журнал клеточной науки. 123 (Pt 15): 2533–2542. Дои:10.1242 / jcs.070490. ЧВК 2912461. PMID 20940129.
- ^ Ху ГБ (август 2014 г.). «Криоэлектронная томография всей клетки предполагает, что митохондрии делятся почкованием». Микроскопия и микроанализ. 20 (4): 1180–1187. Bibcode:2014MiMic..20.1180H. Дои:10.1017 / S1431927614001317. PMID 24870811.
- ^ Кимбалл, Дж. (2006) «Половое размножение у людей: совокупление и оплодотворение», Страницы биологии Кимбалла (на основе Биология, 6 изд., 1996 г.)
- ^ Сутовский П., Морено Р.Д., Рамальо-Сантос Дж., Доминко Т., Симерли С., Шаттен Дж. (Ноябрь 1999 г.). «Убиквитиновая метка для митохондрий сперматозоидов». Природа. 402 (6760): 371–372. Bibcode:1999Натура.402..371С. Дои:10.1038/46466. PMID 10586873. S2CID 205054671. Обсуждается в Новости науки.
- ^ Могенсен Х.Л. (1996). "Как и почему наследование цитоплазмы в семенных растениях". Американский журнал ботаники. 83 (3): 383–404. Дои:10.2307/2446172. JSTOR 2446172.
- ^ Зурос Э. (декабрь 2000 г.). «Исключительная система митохондриальной ДНК семейства мидий Mytilidae». Гены и генетические системы. 75 (6): 313–318. Дои:10.1266 / ggs.75.313. PMID 11280005.
- ^ Сазерленд Б., Стюарт Д., Кенчингтон Э. Р., Зурос Э. (январь 1998 г.). «Судьба отцовской митохондриальной ДНК в развивающихся самках мидий, Mytilus edulis: значение для механизма дважды монородительского наследования митохондриальной ДНК». Генетика. 148 (1): 341–347. ЧВК 1459795. PMID 9475744.
- ^ Митохондриальные линии ДНК самцов и самок голубых мидий (Mytilus edulis) Группа видов В архиве 2013-05-18 в Wayback Machine Дональд Т. Стюарт, Карлос Сааведра, Ребекка Р. Стэнвуд, Эми 0. Болл и Элефтериос Зурос
- ^ Джонс Д.Р. (октябрь 2003 г.). «Отцовская передача митохондриальной ДНК (к счастью) редка». Анналы неврологии. 54 (4): 422–424. Дои:10.1002 / ana.10771. PMID 14520651. S2CID 90422.
- ^ Тьягараджан Б., Падуя, Р. А., Кэмпбелл С. (ноябрь 1996 г.). «Митохондрии млекопитающих обладают активностью гомологичной рекомбинации ДНК». Журнал биологической химии. 271 (44): 27536–27543. Дои:10.1074 / jbc.271.44.27536. PMID 8910339.
- ^ Лант Д.Х., Хайман Британская Колумбия (май 1997 г.). «Рекомбинация митохондриальной ДНК животных». Природа. 387 (6630): 247. Bibcode:1997Натура.387..247л. Дои:10.1038 / 387247a0. PMID 9153388. S2CID 4318161.
- ^ Эйр-Уокер А., Смит Н.Х., Смит Дж. М. (март 1999 г.). "Насколько клональны митохондрии человека?". Ход работы. Биологические науки. 266 (1418): 477–483. Дои:10.1098 / rspb.1999.0662. ЧВК 1689787. PMID 10189711.
- ^ Авадалла П., Эйр-Уокер А., Смит Дж. М. (декабрь 1999 г.). «Нарушение равновесия по сцеплению и рекомбинация в митохондриальной ДНК гоминид». Наука. 286 (5449): 2524–2525. Дои:10.1126 / science.286.5449.2524. PMID 10617471. S2CID 27685285.
- ^ Кри Л.М., Сэмюэлс, округ Колумбия, де Соуза Лопес, СК, Раджасимха, Гонконг, Воннапинидж П., Манн Дж. Р., Даль Х. Х., Чинери П. Ф. (февраль 2008 г.) «Уменьшение количества молекул митохондриальной ДНК во время эмбриогенеза объясняет быструю сегрегацию генотипов». Природа Генетика. 40 (2): 249–254. Дои:10,1038 / нг.2007,63. PMID 18223651. S2CID 205344980.
- ^ Цао Л., Ситара Х, Хории Т., Нагао Й, Имаи Х, Абе К., Хара Т., Хаяси Дж., Ёнекава Х. (март 2007 г.). «Узкое место в митохондриях происходит без снижения содержания мтДНК в половых клетках самок мышей». Природа Генетика. 39 (3): 386–390. Дои:10,1038 / нг1970. PMID 17293866. S2CID 10686347.
- ^ Вай Т., Теоли Д., Шубридж Е.А. (декабрь 2008 г.). «Генетическое« узкое место »митохондриальной ДНК является результатом репликации субпопуляции геномов». Природа Генетика. 40 (12): 1484–1488. Дои:10,1038 / нг.258. PMID 19029901. S2CID 225349.
- ^ Джонстон И.Г., Бургсталлер Дж. П., Хавличек В., Кольбе Т., Рюликке Т., Брем Г., Поултон Дж., Джонс Н. С. (июнь 2015 г.). «Стохастическое моделирование, байесовский вывод и новые измерения in vivo проливают свет на обсуждаемый механизм узкого места мтДНК». eLife. 4: e07464. arXiv:1512.02988. Дои:10.7554 / eLife.07464. ЧВК 4486817. PMID 26035426.
- ^ Гредилла Р., Гарм С., Стевнснер Т. (2012). «Ядерная и митохондриальная репарация ДНК в избранных модельных системах старения эукариот». Oxid Med Cell Longev. 2012: 282438. Дои:10.1155/2012/282438. ЧВК 3462412. PMID 23050036.
- ^ а б c Саки М., Пракаш А. (июнь 2017 г.). «Перекрестные помехи, связанные с повреждением ДНК, между ядром и митохондриями». Свободный Радич. Биол. Med. 107: 216–227. Дои:10.1016 / j.freeradbiomed.2016.11.050. ЧВК 5449269. PMID 27915046.
- ^ Леон Дж., Сакуми К., Кастильо Е., Шенг З., Ока С., Накабеппу Ю. (февраль 2016 г.). «Накопление 8-оксогуанина в митохондриальной ДНК вызывает митохондриальную дисфункцию и нарушает нейритогенез в культивируемых корковых нейронах взрослых мышей в окислительных условиях». Научный представитель. 6: 22086. Bibcode:2016НатСР ... 622086Л. Дои:10.1038 / srep22086. ЧВК 4766534. PMID 26912170.
- ^ Дахал С., Дубей С., Рагхаван СК (май 2018 г.). «Опосредованная гомологичной рекомбинацией репарация двухцепочечных разрывов ДНК работает в митохондриях млекопитающих». Клетка. Мол. Life Sci. 75 (9): 1641–1655. Дои:10.1007 / s00018-017-2702-у. PMID 29116362. S2CID 4779650.
- ^ Одахара М., Иноуэ Т., Фудзита Т., Хасебе М., Секин Ю. (февраль 2007 г.). «Участие RecA, нацеленного на митохондрии, в репарацию митохондриальной ДНК мха Physcomitrella patens». Genes Genet. Syst. 82 (1): 43–51. Дои:10.1266 / ggs.82.43. PMID 17396019.
- ^ Тади СК, Себастьян Р., Дахал С., Бабу Р.К., Чоудхари Б., Рагхаван СК (январь 2016 г.). «Опосредованное микрогомологией соединение концов является основным медиатором репарации двухцепочечных разрывов при повреждениях митохондриальной ДНК». Мол. Биол. Клетка. 27 (2): 223–35. Дои:10.1091 / mbc.E15-05-0260. ЧВК 4713127. PMID 26609070.
- ^ а б Энрикес, Флорида, Ричардс Т.А., Робертс Ф., Маклеод Р., Робертс К.В. (февраль 2005 г.). «Необычный митохондриальный компартмент Cryptosporidium parvum». Тенденции в паразитологии. 21 (2): 68–74. Дои:10.1016 / j.pt.2004.11.010. PMID 15664529.
- ^ Джон У, Лу И, Вольраб С., Грот М, Яноушковец Дж., Кохли Г.С., Марк ФК, Бикмейер У., Фархат С., Фельдер М., Фриккенхаус С., Гийу Л., Килинг П.Дж., Мустафа А., Фарсель Б.М., Валентин К., Глёкнер Г. (Апрель 2019 г.). «Аэробный эукариотический паразит с функциональными митохондриями, у которого, вероятно, отсутствует митохондриальный геном». Достижения науки. 5 (4): eaav1110. Bibcode:2019SciA .... 5.1110J. Дои:10.1126 / sciadv.aav1110. ЧВК 6482013. PMID 31032404.
- ^ «Настоящие электростанции - даже без ДНК: паразитические водоросли из линии динофлагеллят организовали свой генетический материал беспрецедентным образом». ScienceDaily.
- ^ Кастро Дж. А., Пикорнелл А., Рамон М. (декабрь 1998 г.). «Митохондриальная ДНК: инструмент для изучения популяционной генетики». Международная микробиология. 1 (4): 327–332. PMID 10943382.
- ^ Канн Р.Л., Стоункинг М., Уилсон А.С. (январь 1987 г.). «Митохондриальная ДНК и эволюция человека». Природа. 325 (6099): 31–36. Bibcode:1987 Натур. 325 ... 31C. Дои:10.1038 / 325031a0. PMID 3025745. S2CID 4285418.
- ^ Торрони А., Ахилли А., Маколей В., Ричардс М., Бандельт Х. Дж. (Июнь 2006 г.). «Сбор плодов дерева мтДНК человека». Тенденции в генетике. 22 (6): 339–345. Дои:10.1016 / j.tig.2006.04.001. PMID 16678300.
- ^ а б Гарриган Д., Хаммер М.Ф. (сентябрь 2006 г.). «Реконструкция происхождения человека в эпоху генома». Природа Обзоры Генетика. 7 (9): 669–680. Дои:10.1038 / nrg1941. PMID 16921345. S2CID 176541.
- ^ Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S (июль 1997 г.). «Последовательности ДНК неандертальцев и происхождение современного человека». Клетка. 90 (1): 19–30. Дои:10.1016 / S0092-8674 (00) 80310-4. HDL:11858 / 00-001M-0000-0025-0960-8. PMID 9230299. S2CID 13581775.
- ^ Соарес П., Эрмини Л., Томсон Н., Мормина М., Рито Т., Рёль А., Салас А., Оппенгеймер С., Маколей В., Ричардс МБ (июнь 2009 г.). «Поправка на очищающую селекцию: улучшенные митохондриальные молекулярные часы человека». Американский журнал генетики человека. 84 (6): 740–759. Дои:10.1016 / j.ajhg.2009.05.001. ЧВК 2694979. PMID 19500773.
- ^ Нахман М.В., Crowell SL (сентябрь 2000 г.). «Оценка частоты мутаций на нуклеотид у человека». Генетика. 156 (1): 297–304. ЧВК 1461236. PMID 10978293.
- ^ а б c Зевиани М., Ди Донато С. (октябрь 2004 г.). «Митохондриальные нарушения». Мозг. 127 (Пт 10): 2153–2172. Дои:10.1093 / мозг / awh259. PMID 15358637.
- ^ Тейлор RW, Тернбулл DM (май 2005 г.). «Мутации митохондриальной ДНК при заболеваниях человека». Природа Обзоры Генетика. 6 (5): 389–402. Дои:10.1038 / nrg1606. ЧВК 1762815. PMID 15861210.
- ^ Чиннеры П.Ф., Шон Э.А. (сентябрь 2003 г.). «Митохондрии». Журнал неврологии, нейрохирургии и психиатрии. 74 (9): 1188–1199. Дои:10.1136 / jnnp.74.9.1188. ЧВК 1738655. PMID 12933917.
- ^ Шерер ТБ, Бетарбет Р., Гринамир Дж. Т. (июнь 2002 г.). «Окружающая среда, митохондрии и болезнь Паркинсона». Нейробиолог. 8 (3): 192–197. Дои:10.1177/1073858402008003004. PMID 12061498. S2CID 24318009.
- ^ Гомес К., Бандез М.Дж., Наварро А. (январь 2007 г.). «Пестициды и нарушение функции митохондрий в связи с паркинсоническим синдромом». Границы биологических наук. 12: 1079–1093. Дои:10.2741/2128. PMID 17127363.
- ^ Лим Я.А., Райн В., Байсанг Г., Мейер Ф., Поляк А., Рафтери М.Дж., Гильхаус М., Иттнер Л.М., Экерт А., Гетц Дж. (Апрель 2010 г.). «Abeta и человеческий амилин имеют общий путь токсичности через митохондриальную дисфункцию». Протеомика. 10 (8): 1621–1633. Дои:10.1002 / pmic.200900651. PMID 20186753. S2CID 33077667.
- ^ Кинг, Джон V .; Liang, Wenguang G .; Scherpelz, Kathryn P .; Шиллинг, Александр Б .; Мередит, Стивен С .; Тан, Вэй-Джен (2014-07-08). «Молекулярная основа распознавания и деградации субстрата протеазой предпоследовательности человека». Структура (Лондон, Англия: 1993). 22 (7): 996–1007. Дои:10.1016 / j.str.2014.05.003. ISSN 1878-4186. ЧВК 4128088. PMID 24931469.
- ^ Шапира А.Х. (июль 2006 г.). «Митохондриальная болезнь». Ланцет. 368 (9529): 70–82. Дои:10.1016 / S0140-6736 (06) 68970-8. PMID 16815381. S2CID 27039617.
- ^ Pieczenik SR, Neustadt J (август 2007 г.). «Митохондриальная дисфункция и молекулярные пути заболевания». Экспериментальная и молекулярная патология. 83 (1): 84–92. Дои:10.1016 / j.yexmp.2006.09.008. PMID 17239370.
- ^ Баггер Х., Абель Э.Д. (ноябрь 2010 г.). «Митохондрии в сердце диабетика». Сердечно-сосудистые исследования. 88 (2): 229–240. Дои:10.1093 / cvr / cvq239. ЧВК 2952534. PMID 20639213.
- ^ Рихтер С., Парк Дж. У., Эймс Б. Н. (сентябрь 1988 г.). «Нормальное окислительное повреждение митохондриальной и ядерной ДНК обширно». Труды Национальной академии наук Соединенных Штатов Америки. 85 (17): 6465–6467. Bibcode:1988PNAS ... 85.6465R. Дои:10.1073 / пнас.85.17.6465. ЧВК 281993. PMID 3413108.
- ^ Харман Д. (июль 1956 г.). «Старение: теория, основанная на свободнорадикальной и радиационной химии». Журнал геронтологии. 11 (3): 298–300. CiteSeerX 10.1.1.663.3809. Дои:10.1093 / geronj / 11.3.298. PMID 13332224.
- ^ Харман Д. (апрель 1972 г.). «Биологические часы: митохондрии?». Журнал Американского гериатрического общества. 20 (4): 145–147. Дои:10.1111 / j.1532-5415.1972.tb00787.x. PMID 5016631. S2CID 396830.
- ^ «Митохондрии и старение». circuitblue.co.
- ^ Boffoli D, Scacco SC, Vergari R, Solarino G, Santacroce G, Papa S (апрель 1994). «Снижение с возрастом активности дыхательной цепи в скелетных мышцах человека». Biochimica et Biophysica Acta (BBA) - Молекулярная основа болезни. 1226 (1): 73–82. Дои:10.1016/0925-4439(94)90061-2. PMID 8155742.
- ^ де Грей AD (2004). «Митохондриальные мутации при старении млекопитающих: слишком поспешный поворот?». Исследования омоложения. 7 (3): 171–174. Дои:10.1089 / rej.2004.7.171. PMID 15588517.
- ^ Бендер А., Кришнан К.Дж., Моррис С.М., Тейлор Г.А., Рив А.К., Перри Р.Х., Ярос Э., Хершсон Дж.С., Беттс Дж., Клопсток Т., Тейлор Р.В., Тернбулл Д.М. (май 2006 г.). «Высокий уровень делеций митохондриальной ДНК в нейронах черного вещества при старении и болезни Паркинсона». Природа Генетика. 38 (5): 515–517. Дои:10,1038 / ng1769. PMID 16604074. S2CID 13956928.
- ^ Mehta, Arpan R .; Уолтерс, Рэйчел; Waldron, Fergal M .; Пал, Суванкар; Selvaraj, Bhuvaneish T .; Macleod, Malcolm R .; Hardingham, Giles E .; Чандран, Сиддхартхан; Грегори, Дженна М. (01.01.2019). «Устранение митохондриальной дисфункции при боковом амиотрофическом склерозе: систематический обзор и метаанализ». Мозговые коммуникации. 1 (1). Дои:10.1093 / braincomms / fcz009. PMID 32133457. S2CID 201168692.
- ^ Chiang, J .; Шукла П. (ноябрь 2020 г.). «Митохондрии в старении яичников и репродуктивном долголетии». Обзоры исследований старения. 63. Дои:10.1016 / j.arr.2020.101168.
- ^ а б c d е ж грамм час я j Эрнстер Л., Шац Г. (декабрь 1981 г.). «Митохондрии: исторический обзор». Журнал клеточной биологии. 91 (3, часть 2): 227–255 с. Дои:10.1083 / jcb.91.3.227s. ЧВК 2112799. PMID 7033239.
- ^ Альтманн, Р. 1890. Die Elementarorganismen und ihre Beziehungen zu den Zellen. Файт, Лейпциг, [1].
- ^ «митохондрии». Интернет-словарь этимологии.
- ^ Бенда С. (1898). "Ueber die Spermatogense der Vertebraten und höherer Evertebraten. II. Theil: Die Histiogense der Spermien". Arch. Анальный. Physiol.: 393–398.
- ^ Цитата Эрнстера Мевес, Фридрих (май 1908 г.). "Die Chondriosomen als Träger erblicher Anlagen. Cytologische Studien am Hühnerembryo". Archiv für Mikroskopische Anatomie. 72 (1): 816–867. Дои:10.1007 / BF02982402. S2CID 85047330. неправильно, правильная цитата Мевес, Фридрих (1904). "Über das Vorkommen von Mitochondrien bezw. Chondromiten in Pflanzenzellen". Бер. Dtsch. Бот. Ges. 22: 284–286., цитируется в статье Мевеса 1908 г. и в Шмидт, Эрнст Вилли (1913). "Pflanzliche Mitochondrien". Progressus Rei Botanicae. 4: 164–183. Получено 21 сентября 2012., с подтверждением Nymphaea alba
- ^ Мартин В.Ф., Гарг С., Зиморский В. (сентябрь 2015 г.). "Эндосимбиотические теории происхождения эукариот". Философские труды Лондонского королевского общества. Серия B, Биологические науки. 370 (1678): 20140330. Дои:10.1098 / rstb.2014.0330. ЧВК 4571569. PMID 26323761.
- ^ Нарцисс Э (10 августа 2010 г.). «20 000 на ячейку: почему мидихлорианы - отстой». Время. Получено 19 июня, 2016.
Общий
 Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».
Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».
внешняя ссылка
- Лейн, Ник (2016). Жизненно важный вопрос: энергия, эволюция и происхождение сложной жизни. WW Norton & Company. ISBN 978-0393352979.
- Mitodb.com - База данных митохондриальных заболеваний.
- Атлас митохондрий в Университет Майнца
- Портал исследования митохондрий на mitochondrial.net
- Митохондрии: архитектура определяет функцию на cytochemistry.net
- Связи митохондрий в Университет Алабамы
- MIP Общество митохондриальной физиологии
- 3D-структуры белков внутренней митохондриальной мембраны в университет Мичигана
- Трехмерные структуры белков, связанных с внешней митохондриальной мембраной в университет Мичигана
- Партнерство митохондриальных белков в Университет Висконсина
- MitoMiner - база данных митохондриальной протеомики в MRC Отделение митохондриальной биологии
- Митохондрия - база данных, ориентированная на клетки
- Митохондрия, реконструированная с помощью электронной томографии в Государственный университет Сан-Диего
- Видеоклип митохондрии печени крысы с криоэлектронной томографии