Прокариотический цитоскелет - Prokaryotic cytoskeleton
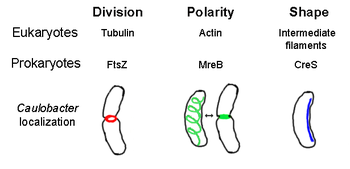
В прокариотический цитоскелет это собирательное название для всех структурных нити в прокариоты. Когда-то считалось, что прокариотические клетки не обладают цитоскелеты, но достижения в области технологий визуализации и определения структуры привели к открытию филаментов в этих клетках в начале 1990-х годов.[2] Не только аналоги для всех основных белков цитоскелета в эукариоты были обнаружены у прокариот, цитоскелетных белков, эукариотические белки неизвестны. гомологи также были обнаружены.[3][4][5][6] Элементы цитоскелета играют важную роль в деление клеток, защита, определение формы и определение полярности у различных прокариот.[7][8]
FtsZ
FtsZ, первый идентифицированный прокариотический цитоскелетный элемент, образует нитевидную кольцевую структуру, расположенную в середине клетки, называемую Z-кольцом, которое сжимается во время деление клеток, аналогично актин-миозиновое сократительное кольцо у эукариот.[2] Z-кольцо - это очень динамичная структура, которая состоит из многочисленных пучков протофиламентов, которые расширяются и сжимаются, хотя механизм сокращения Z-кольца и количество задействованных протофиламентов неясны.[1] FtsZ действует как белок-организатор и необходим для деления клеток. Это первый компонент перегородка в течение цитокинез, и он привлекает все другие известные белки деления клетки к участку деления.[9]
Несмотря на это функциональное сходство с актин, FtsZ гомологичен эукариалу тубулин. Хотя сравнение основные конструкции FtsZ и тубулина обнаруживают слабую взаимосвязь, их трехмерные структуры удивительно похожи. Кроме того, как и тубулин, мономерный FtsZ привязан к GTP и полимеризуется с другими мономерами FtsZ с гидролизом GTP по механизму, аналогичному димеризация тубулина.[10] Поскольку FtsZ необходим для деления клеток у бактерий, этот белок является мишенью для разработки новых антибиотики.[11]В настоящее время существует несколько моделей и механизмов, которые регулируют образование Z-кольца, но эти механизмы зависят от вида. Несколько видов стержней, в том числе кишечная палочка и Caulobacter crescentus, используйте один или несколько ингибиторов сборки FtsZ, которые образуют биполярный градиент в клетке, усиливая полимеризацию FtsZ в центре клетки.[12] Одна из этих градиентообразующих систем состоит из белков MinCDE (см. Ниже).
MreB
MreB бактериальный белок, который считается гомологичным эукариалам актин. MreB и актин обладают слабым первичная структура совпадают, но очень похожи с точки зрения трехмерной структуры и полимеризации волокон.
Почти все несферические бактерии полагаются на MreB для определения своей формы. MreB собирается в спиральную сеть нитчатых структур прямо под цитоплазматическая мембрана, покрывающего всю длину ячейки.[13] MreB определяет форму клетки, опосредуя положение и активность ферментов, которые синтезируют пептидогликан и действуя как жесткая нить под клеточной мембраной, которая оказывает внешнее давление, формируя и поддерживая клетку.[1] MreB конденсируется из своей нормальной спиральной сети и образует плотное кольцо на перегородка в Caulobacter crescentus непосредственно перед делением клетки, механизм, который, как полагают, помогает определить местонахождение перегородки, не соответствующей центру.[14] MreB также важен для определения полярности у полярных бактерий, поскольку он отвечает за правильное расположение как минимум четырех различных полярных белков в C. crescentus.[14]
Crescentin
Crescentin (закодировано CreS ген) является аналогом эукариотического промежуточные нити (ЕСЛИ). В отличие от других аналогичных взаимосвязей, обсуждаемых здесь, crescentin имеет довольно большую первичную гомологию с белками IF в дополнение к трехмерному сходству - последовательность CreS имеет 25% совпадение идентичности и 40% сходство с цитокератин 19 и совпадение на 24% и сходство на 40% с ядерный ламин А. Кроме того, филаменты сресцентина имеют диаметр примерно 10 нм и, таким образом, попадают в диапазон диаметров эукариальных IF (8-15 нм).[15] Кресцентин образует непрерывную нить от полюса к полюсу вдоль внутренней вогнутой стороны бактерии, имеющей форму полумесяца. Caulobacter crescentus. И MreB, и crescentin необходимы для C. crescentus существовать в своей характерной форме; Считается, что MreB придает клетке форму стержня, а полумесяц изгибает эту форму в виде полумесяца.[1]
ParM и SopA
ParM это элемент цитоскелета, который имеет структуру, аналогичную актин, хотя функционально ведет себя как тубулин. Кроме того, он двунаправленно полимеризуется и проявляет динамическая нестабильность, оба поведения характерны для полимеризации тубулина.[4][16] Он образует систему с ParR и парк что отвечает за Плазмида R1 разделение. ParM присоединяется к ParR, a ДНК-связывающий белок который специфически связывается с 10 прямыми повторами в парк область на плазмиде R1. Это связывание происходит на обоих концах нити ParM. Затем эта нить удлиняется, разделяя плазмиды.[17] Эта система аналогична эукариотической сегрегации хромосом, поскольку ParM действует как эукариотическая тубулин в митотическое веретено, ParR действует как кинетохора комплекс, и парк действует как центромера из хромосома.[18]
F плазмида сегрегация происходит в аналогичной системе, где SopA действует как филамент цитоскелета, а SopB связывается с sopC последовательность в плазмиде F, как кинетохора и центромера соответственно.[18] Недавно актин-подобный гомолог ParM был обнаружен в грамположительный бактерия Bacillus thuringiensis, который собирается в структуру, подобную микротрубочке, и участвует в плазмида сегрегация.[19]
Система MinCDE
В Система MinCDE система нитей, которая правильно позиционирует перегородка в середине камеры в кишечная палочка. Согласно Shih et al., MinC ингибирует образование перегородки, запрещая полимеризацию Z-кольца. MinC, MinD и MinE образуют спиральную структуру, которая наматывается вокруг клетки и связывается с мембраной с помощью MinD. Спираль MinCDE занимает полюс и оканчивается нитевидной структурой, называемой E-кольцом, сделанной из MinE, на самом среднем краю полярной зоны. В этой конфигурации E-кольцо будет сжиматься и двигаться к этому полюсу, разбирая спираль MinCDE по мере ее движения. Одновременно разобранные фрагменты снова соберутся на противоположном полярном конце, преобразовав катушку MinCDE на противоположном полюсе, в то время как текущая спираль MinCDE будет разрушена. Затем этот процесс повторяется, спираль MinCDE колеблется от полюса к полюсу. Это колебание происходит неоднократно в течение клеточного цикла, тем самым поддерживая MinC (и его эффект ингибирования перегородки) на более низкой усредненной по времени концентрации в середине клетки, чем на концах клетки.[20]
Динамическое поведение белков Min было реконструировано in vitro с использованием искусственного липидного бислоя в качестве имитации клеточной мембраны. MinE и MinD самоорганизуются в параллельные и спиральные белковые волны по механизму, подобному реакции-диффузии.[21]
Бактофилин
Бактофилин (ИнтерПро: IPR007607 ) представляет собой цитоскелетный элемент, который образует волокна в клетках стержневидный протеобактерии Myxococcus xanthus.[22] Белок бактофилин, BacM, необходим для поддержания правильной формы клетки и целостности клеточной стенки. М. xanthus клетки, лишенные BacM, имеют деформированную морфологию, характеризующуюся изогнутым телом клетки, и BACM мутанты обладают пониженной устойчивостью к антибиотикам, направленным на клеточную стенку бактерий. М. xanthus Белок BacM отщепляется от его полноразмерной формы, чтобы обеспечить полимеризацию. Бактофилины участвуют в регуляции формы клеток у других бактерий, включая искривление Протей мирабилис клетки[23] формирование стебля Caulobacter crescentus,[24] и винтовая форма Helicobacter pylori.[25]
Кренактин
Кренактин - гомолог актина, уникальный для царства архей. Crenarchaeota что было найдено в заказах Thermoproteales и Candidatus Korarchaeum.[26] Он имеет самое высокое сходство последовательностей с эукариотическими актинами из всех известных гомологов актина.[27] Кренактин хорошо охарактеризован в Pyryobaculum calidifontis (A3MWN5) и показали высокую специфичность к АТФ и ГТФ.[26] Все виды, содержащие кренактин, имеют форму стержня или иглы и имеют форму P. calidifontis Было показано, что кренактин формирует спиральные структуры, которые охватывают длину клетки, что указывает на роль кренактина в определении формы, аналогичную роли MreB у других прокариот.[26][28]
CfpA
Внутри филума Спирохеты, ряд видов разделяют структуру нитевидной цитоплазматической ленты, образованную отдельными филаментами, состоящими из белка CfpA (Cytoplasmic filament protein A, Q56336), соединенных между собой соединительными элементами и анкерами с внутренней мембраной.[29][30] Пока присутствует в родах Трепонема, Спирохета, Пиллотина, Лептонема, Голландия и Диплокаликс, однако у некоторых видов они отсутствуют, как на примере Treponema primitia.[31][32][33][34] При размере поперечного сечения 5 x 6 нм (горизонтально / вертикально) они попадают в диапазон диаметров эукариальных промежуточных филаментов (IF) (8-15 нм). Treponema denticola клетки, лишенные белка CfpA, образуют длинные сцепленные клетки с дефектом сегрегации хромосомной ДНК, фенотип, также влияющий на патогенность этого организма.[35][36] Отсутствие другой ультраструктуры клетки, пучка филаментов периплазматических жгутиков, не изменяет структуру цитоплазматической ленты.[37]
Смотрите также
Рекомендации
- ^ а б c d Gitai Z (март 2005 г.). «Новая биология бактериальных клеток: движущиеся части и субклеточная архитектура». Клетка. 120 (5): 577–86. Дои:10.1016 / j.cell.2005.02.026. PMID 15766522. S2CID 8894304.
- ^ а б Би EF, Lutkenhaus J (ноябрь 1991 г.). «Кольцевая структура FtsZ, связанная с делением у Escherichia coli». Природа. 354 (6349): 161–4. Дои:10.1038 / 354161a0. PMID 1944597. S2CID 4329947.
- ^ Gunning PW, Ghoshdastider U, Whitaker S, Popp D, Robinson RC (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых филаментов». Журнал клеточной науки. 128 (11): 2009–19. Дои:10.1242 / jcs.165563. PMID 25788699.
- ^ а б Попп Д., Нарита А., Ли Л.Дж., Гошдастидер Ю., Сюэ Б., Сринивасан Р., Баласубраманиан М.К., Танака Т., Робинсон Р.К. (июнь 2012 г.). «Новая актиноподобная структура филаментов из Clostridium tetani». Журнал биологической химии. 287 (25): 21121–9. Дои:10.1074 / jbc.M112.341016. ЧВК 3375535. PMID 22514279.
- ^ Попп Д., Нарита А., Гошдастидер У., Маеда К., Маеда Й, Ода Т., Фудзисава Т., Ониши Х., Ито К., Робинсон Р.С. (апрель 2010 г.). «Полимерные структуры и динамические свойства бактериального актина AlfA». Журнал молекулярной биологии. 397 (4): 1031–41. Дои:10.1016 / j.jmb.2010.02.010. PMID 20156449.
- ^ Уикстед Б., Чайка К. (август 2011 г.). «Эволюция цитоскелета». Журнал клеточной биологии. 194 (4): 513–25. Дои:10.1083 / jcb.201102065. ЧВК 3160578. PMID 21859859.
- ^ Ши Ю.Л., Ротфилд Л. (сентябрь 2006 г.). «Бактериальный цитоскелет». Обзоры микробиологии и молекулярной биологии. 70 (3): 729–54. Дои:10.1128 / MMBR.00017-06. ЧВК 1594594. PMID 16959967.
- ^ Мичи КА, Лёве Дж (2006). «Динамические нити цитоскелета бактерий» (PDF). Ежегодный обзор биохимии. 75: 467–92. Дои:10.1146 / annurev.biochem.75.103004.142452. PMID 16756499. Архивировано из оригинал (PDF) 17 ноября 2006 г.
- ^ Грауманн П.Л. (декабрь 2004 г.). «Элементы цитоскелета у бактерий». Текущее мнение в микробиологии. 7 (6): 565–71. Дои:10.1016 / j.mib.2004.10.010. PMID 15556027.
- ^ Десаи А., Митчисон Т.Дж. (июль 1998 г.). «Структуры тубулина и ФцЗ: функциональное и терапевтическое значение». BioEssays. 20 (7): 523–7. Дои:10.1002 / (SICI) 1521-1878 (199807) 20: 7 <523 :: AID-BIES1> 3.0.CO; 2-L. PMID 9722999.
- ^ Haydon DJ, Stokes NR, Ure R, Galbraith G, Bennett JM, Brown DR, Baker PJ, Barynin VV, Rice DW, Sedelnikova SE, Heal JR, Sheridan JM, Aiwale ST, Chauhan PK, Srivastava A, Taneja A, Collins I. , Эррингтон Дж., Чаплевски Л.Г. (сентябрь 2008 г.). «Ингибитор FtsZ с сильной и селективной антистафилококковой активностью». Наука. 321 (5896): 1673–5. Дои:10.1126 / science.1159961. PMID 18801997. S2CID 7878853.
- ^ Хойссер Д.П., Марголин В. (апрель 2016 г.). «Сплитсвилл: структурное и функциональное понимание динамического Z-кольца бактерий». Обзоры природы. Микробиология. 14 (5): 305–19. Дои:10.1038 / nrmicro.2016.26. ЧВК 5290750. PMID 27040757.
- ^ Курнер Дж., Медалия О., Линарудис А.А., Баумейстер В. (ноябрь 2004 г.). «Новое понимание структурной организации эукариотических и прокариотических цитоскелетов с использованием криоэлектронной томографии». Экспериментальные исследования клеток. 301 (1): 38–42. Дои:10.1016 / j.yexcr.2004.08.005. PMID 15501443.
- ^ а б Gitai Z, Dye N, Shapiro L (июнь 2004 г.). «Актин-подобный ген может определять полярность клеток у бактерий». Труды Национальной академии наук Соединенных Штатов Америки. 101 (23): 8643–8. Дои:10.1073 / pnas.0402638101. ЧВК 423248. PMID 15159537.
- ^ Осмис Н., Кун Дж. Р., Якобс-Вагнер С. (декабрь 2003 г.). «Бактериальный цитоскелет: промежуточная филаментоподобная функция в форме клетки». Клетка. 115 (6): 705–13. Дои:10.1016 / S0092-8674 (03) 00935-8. PMID 14675535. S2CID 14459851.
- ^ Гарнер ЕС, Кэмпбелл С.С., Маллинз Р.Д. (ноябрь 2004 г.). «Динамическая нестабильность в ДНК-сегрегации гомолога прокариотического актина». Наука. 306 (5698): 1021–5. Дои:10.1126 / science.1101313. PMID 15528442. S2CID 14032209.
- ^ Мёллер-Йенсен Дж., Йенсен Р. Б., Лёве Дж., Гердес К. (июнь 2002 г.). «Прокариотическая сегрегация ДНК актиноподобным филаментом». Журнал EMBO. 21 (12): 3119–27. Дои:10.1093 / emboj / cdf320. ЧВК 126073. PMID 12065424.
- ^ а б Gitai Z (февраль 2006 г.). «Плазмидная сегрегация: появляется новый класс цитоскелетных белков». Текущая биология. 16 (4): R133-6. Дои:10.1016 / j.cub.2006.02.007. PMID 16488865.
- ^ Jiang S, Narita A, Popp D, Ghoshdastider U, Lee LJ, Srinivasan R, Balasubramanian MK, Oda T, Koh F, Larsson M, Robinson RC (март 2016 г.). «Новые актиновые филаменты из Bacillus thuringiensis образуют нанотрубочки для сегрегации плазмидной ДНК». Труды Национальной академии наук Соединенных Штатов Америки. 113 (9): E1200-5. Дои:10.1073 / pnas.1600129113. ЧВК 4780641. PMID 26873105.
- ^ Ши Ю.Л., Ле Т., Ротфилд Л. (июнь 2003 г.). «Выбор места деления в Escherichia coli включает динамическое перераспределение белков Min в спиральных структурах, которые простираются между двумя полюсами клетки». Труды Национальной академии наук Соединенных Штатов Америки. 100 (13): 7865–70. Дои:10.1073 / pnas.1232225100. ЧВК 164679. PMID 12766229.
- ^ Loose M, Fischer-Friedrich E, Ries J, Kruse K, Schwille P (май 2008 г.). «Пространственные регуляторы для самоорганизации бактериальных клеток в поверхностные волны in vitro». Наука. 320 (5877): 789–92. Дои:10.1126 / science.1154413. PMID 18467587. S2CID 27134918.
- ^ Koch MK, McHugh CA, Hoiczyk E (май 2011 г.). «BacM, бактофилин Myxococcus xanthus, процессируемый на N-конце, имеет решающее значение для правильной формы клеток». Молекулярная микробиология. 80 (4): 1031–51. Дои:10.1111 / j.1365-2958.2011.07629.x. ЧВК 3091990. PMID 21414039.
- ^ Hay NA, Tipper DJ, Gygi D, Hughes C (апрель 1999 г.). «Новый мембранный белок, влияющий на форму клеток и многоклеточное скопление Proteus mirabilis». Журнал бактериологии. 181 (7): 2008–16. Дои:10.1128 / JB.181.7.2008-2016.1999. ЧВК 93611. PMID 10094676.
- ^ Kühn J, Briegel A, Mörschel E, Kahnt J, Leser K, Wick S, Jensen GJ, Thanbichler M (январь 2010 г.). «Бактофилины, повсеместный класс цитоскелетных белков, опосредующих полярную локализацию синтазы клеточной стенки у Caulobacter crescentus». Журнал EMBO. 29 (2): 327–39. Дои:10.1038 / emboj.2009.358. ЧВК 2824468. PMID 19959992.
- ^ Sycuro LK, Pincus Z, Gutierrez KD, Biboy J, Stern CA, Vollmer W., Salama NR (май 2010 г.). «Релаксация сшивания пептидогликанов способствует формированию спиральной формы Helicobacter pylori и колонизации желудка». Клетка. 141 (5): 822–33. Дои:10.1016 / j.cell.2010.03.046. ЧВК 2920535. PMID 20510929.
- ^ а б c Эттема Т.Дж., Линдос А.С., Бернандер Р. (май 2011 г.). «Актиновый цитоскелет архей». Молекулярная микробиология. 80 (4): 1052–61. Дои:10.1111 / j.1365-2958.2011.07635.x. PMID 21414041.
- ^ Ютин Н., Вольф М.Ю., Вольф Ю.И., Кунин Е.В. (февраль 2009 г.). «Истоки фагоцитоза и эукариогенеза». Биология Директ. 4: 9. Дои:10.1186/1745-6150-4-9. ЧВК 2651865. PMID 19245710.
- ^ Ghoshdastider U, Jiang S, Popp D, Robinson RC (июль 2015 г.). «В поисках первозданной актиновой нити». Труды Национальной академии наук Соединенных Штатов Америки. 112 (30): 9150–1. Дои:10.1073 / pnas.1511568112. ЧВК 4522752. PMID 26178194.
- ^ Изард Дж., МакИвен Б.Ф., Барнард Р.М., Португальский Т., Самсонов В.А., Лимбергер Р.Дж. (февраль 2004 г.). «Томографическая реконструкция трепонемных цитоплазматических филаментов выявляет новые мостиковые и закрепляющие компоненты». Молекулярная микробиология. 51 (3): 609–18. Дои:10.1046 / j.1365-2958.2003.03864.x. PMID 14731266.
- ^ Ю И, Элмор С., Колтон Л.Л., Маккензи С., Ступс Дж. К., Вайншток Г.М., Норрис С.Дж. (июнь 1996 г.). «Характеристика гена белка цитоплазматических филаментов (cfpA) Treponema pallidum subsp. Pallidum». Журнал бактериологии. 178 (11): 3177–87. Дои:10.1128 / jb.178.11.3177-3187.1996. ЧВК 178068. PMID 8655496.
- ^ Изард Дж (2006). «Цитоскелетная цитоплазматическая филаментная лента Treponema: член семейства промежуточных филаментных белков». Журнал молекулярной микробиологии и биотехнологии. 11 (3–5): 159–66. Дои:10.1159/000094052. PMID 16983193. S2CID 40913042.
- ^ Murphy GE, Matson EG, Leadbetter JR, Berg HC, Jensen GJ (март 2008 г.). «Новые ультраструктуры Treponema primitia и их значение для подвижности». Молекулярная микробиология. 67 (6): 1184–95. Дои:10.1111 / j.1365-2958.2008.06120.x. ЧВК 3082362. PMID 18248579.
- ^ Изард Дж., Ренкен С., Се CE, Дерозье, округ Колумбия, Данхэм-Эмс, С., Ла Ваке, Гебхардт Л.Л., Лимбергер Р.Дж., Кокс Д.Л., Марко М., Радольф, Д.Д. (декабрь 2009 г.). «Криоэлектронная томография проясняет молекулярную архитектуру Treponema pallidum, спирохеты сифилиса». Журнал бактериологии. 191 (24): 7566–80. Дои:10.1128 / JB.01031-09. ЧВК 2786590. PMID 19820083.
- ^ Изард Дж., Се С.Е., Лимбергер Р.Дж., Маннелла К.А., Марко М. (июль 2008 г.). «Нативная клеточная архитектура Treponema denticola, выявленная с помощью криоэлектронной томографии». Журнал структурной биологии. 163 (1): 10–7. Дои:10.1016 / j.jsb.2008.03.009. ЧВК 2519799. PMID 18468917.
- ^ Изард Дж., Самсонов В.А., Лимбергер Р.Дж. (февраль 2001 г.). «Мутант Treponema denticola с дефицитом цитоплазматических филаментов имеет плейотропные дефекты». Журнал бактериологии. 183 (3): 1078–84. CiteSeerX 10.1.1.488.5178. Дои:10.1128 / JB.183.3.1078-1084.2001. ЧВК 94976. PMID 11208807.
- ^ Изард Дж, Сасаки Х, Кент Р. (2012). «Патогенность Treponema denticola дикого типа и мутантного штамма, проверенная активным режимом пародонтальной инфекции с использованием микроинъекции». Международный журнал стоматологии. 2012: 549169. Дои:10.1155/2012/549169. ЧВК 3398590. PMID 22829826.
- ^ Изард Дж., Самсонов В.А., Киношита МБ, Лимбергер Р.Дж. (ноябрь 1999 г.). «Генетические и структурные анализы цитоплазматических нитей Treponema phagedenis дикого типа и мутанта с дефицитом жгутиковых нитей». Журнал бактериологии. 181 (21): 6739–46. Дои:10.1128 / JB.181.21.6739-6746.1999. ЧВК 94139. PMID 10542176.