Пластид - Plastid
| Пластид | |
|---|---|
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Цианобактерии |
| Clade: | Пластиды |

В пластида (Греческий: πλαστός; plastós: сформированный, формованный - множественное число пластиды) это мембраносвязанный органелла[1] найдено в клетки из растения, водоросли, и некоторые другие эукариотический организмы. Они считаются эндосимбиотический Цианобактерии, связанный с Gloeomargarita.[2] Событие постоянного эндосимбиоза, вероятно, произошло с цианобионт.[3] Пластиды были открыты и названы Эрнст Геккель, но А. Ф. В. Шимпер был первым, кто дал четкое определение. Пластиды - это место производства и хранения важных химических соединений, используемых клетками автотрофный эукариоты. Они часто содержат пигменты используется в фотосинтез, а типы пигментов в пластиде определяют цвет клетки. Они имеют общее эволюционное происхождение и обладают двухцепочечная ДНК молекула круглая, как у круговая хромосома из прокариотические клетки.
В растениях

Пластиды, содержащие хлорофилл может выполнять фотосинтез и называются хлоропласты. Пластиды также могут хранить такие продукты, как крахмал и может синтезировать жирные кислоты и терпены, который можно использовать для производства энергии и в качестве сырья для синтеза других молекул. Например, компоненты кутикула растений и это эпикутикулярный воск синтезируются эпидермальные клетки от пальмитиновая кислота, который синтезируется в хлоропластах ткань мезофилла.[4] Все пластиды происходят из пропластидов, которые присутствуют в меристематический регионы завода. Пропластиды и молодые хлоропласты обычно делятся на двойное деление, но более зрелые хлоропласты также обладают этой способностью.
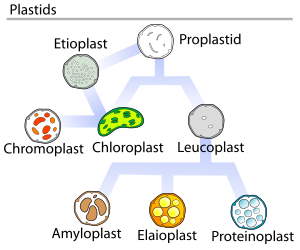
В растения, пропластиды (недифференцированные пластиды) могут различать на несколько форм, в зависимости от того, какую функцию они выполняют в клетке. Они могут развиться в любой из следующих вариантов:[5]
- Хлоропласты: зеленые пластиды для фотосинтез. Этиопласты являются предшественниками хлоропластов.
- Родопласты: красные хлоропласты, обнаруженные в красные водоросли
- Муропласты (также известные как цианопласты или цианеллы): хлоропласты глаукофиты
- Вторичные и третичные хлоропласты: от эндосимбиоза зеленые водоросли и красные водоросли
- Хромопласты: цветные пластиды для синтеза и хранения пигментов
- Геронтопласты: контролировать демонтаж фотосинтетического аппарата во время старение растений
- Лейкопласты: бесцветные пластиды для монотерпен синтез; лейкопласты иногда дифференцируются в более специализированные пластиды:
- Амилопласты: для крахмал хранение и обнаружение сила тяжести (для геотропизм )
- Элайопласты: для хранения жир
- Протеинопласты: для хранения и изменения белок
- Танносомы: для синтеза и производства дубильные вещества и полифенолы
В зависимости от своей морфологии и функции пластиды обладают способностью дифференцировать или повторно дифференцироваться между этими и другими формами.
Каждая пластида создает несколько копий круговой 10–250 килобаза пластом [6][7]. Число копий генома на пластиду варьирует от более 1000 в кратчайшие сроки. делящиеся клетки, которые, как правило, содержат мало пластид, до 100 или меньше в зрелых клетках, где пластидные деления дали начало большому количеству пластид. В пластоме содержится около 100 гены кодирование рибосом и перенос рибонуклеиновые кислоты (рРНК и тРНК ), а также белки, участвующие в фотосинтезе, и пластидный ген транскрипция и перевод. Однако эти белки представляют собой лишь небольшую часть общей белковой структуры, необходимой для создания и поддержания структуры и функции определенного типа пластиды. Завод ядерный гены кодируют подавляющее большинство пластидных белков, а экспрессия пластидных генов и ядерных генов тесно ко-регулируется для координации правильного развития пластид по отношению к дифференциация клеток.
Пластидная ДНК существует в виде больших комплексов белок-ДНК, связанных с внутренней оболочкой. мембрана и называется «пластидными нуклеоидами». Каждая частица нуклеоида может содержать более 10 копий пластидной ДНК. Пропластида содержит единственный нуклеоид, расположенный в центре пластиды. Развивающаяся пластида имеет множество нуклеоидов, локализованных на периферии пластиды, связанных с внутренней оболочкой мембраны. Во время развития пропластидов в хлоропласты и когда пластиды превращаются из одного типа в другой, нуклеоиды изменяются по морфологии, размеру и расположению в органелле. Считается, что ремоделирование нуклеоидов происходит за счет модификации состава и количества нуклеоидных белков.
Многие пластиды, особенно те, которые отвечают за фотосинтез, имеют многочисленные внутренние мембранные слои.
В клетки растений, длинные тонкие выступы, называемые стромулы иногда образуются и простираются от основного пластидного тела в цитозоль и соединить между собой несколько пластид. Белки и, предположительно, более мелкие молекулы могут перемещаться внутри стромул. Большинство культивируемых клеток, которые являются относительно большими по сравнению с другими растительными клетками, имеют очень длинные и многочисленные стромулы, которые доходят до периферии клетки.
В 2014 г. были обнаружены свидетельства возможной потери генома пластид у Раффлезия лагаски, нефотосинтетический паразитический цветущее растение, а в Политомелла, род нефотосинтетических зеленые водоросли. Обширные поиски пластидных генов в обоих Раффлезия и Политомелла не дали результатов, однако вывод о том, что их пластомы полностью отсутствуют, все еще остается спорным.[8] Некоторые ученые утверждают, что потеря пластидного генома маловероятна, поскольку даже нефотосинтетические пластиды содержат гены, необходимые для завершения различных биосинтетические пути, например, биосинтез гема.[8][9]
В водорослях
В водоросли термин лейкопласт используется для всех непигментированных пластид. Их функция отличается от лейкопластов растений. Этиопласты, амилопласты и хромопласты специфичны для растений и не встречаются в водорослях.[нужна цитата ] Пластиды в водорослях и роголистник также могут отличаться от пластид растений тем, что содержат пиреноиды. Апикопласты нефотосинтетические пластиды Apicomplexa происходит из вторичного эндосимбиоза.
Глаукофит водоросли содержат муропласты, похожие на хлоропласты, за исключением того, что они имеют пептидогликан клеточная стенка, похожая на прокариоты. Красные водоросли содержат родопласты - красные хлоропласты, которые позволяют им фотосинтезировать на глубине до 268 м.[5] Хлоропласты растений отличаются от родопластов красных водорослей своей способностью синтезировать крахмал, который хранится в виде гранул в пластидах. В красных водорослях, флоридский крахмал синтезируется и хранится вне пластид в цитозоле.[10]
Наследование
Большинство растений наследуют пластиды только от одного родителя. В общем, покрытосеменные наследуют пластиды от женской гаметы, тогда как многие голосеменные наследуют пластиды от мужской пыльцы. Водоросли также наследуют пластиды только от одного родителя. Таким образом, пластидная ДНК другого родителя полностью утеряна.
При нормальных внутривидовых скрещиваниях (приводящих к появлению нормальных гибридов одного вида) наследование пластидной ДНК оказывается на 100% однородным. Однако при межвидовой гибридизации наследование пластид оказывается более неустойчивым. Хотя пластиды наследуются в основном по материнской линии при межвидовой гибридизации, имеется много сообщений о гибридах цветковых растений, которые содержат пластиды отца. Примерно 20% покрытосеменных растений, включая люцерна (Medicago sativa), обычно демонстрируют наследование пластид от двух родителей.[11]
Повреждение и восстановление ДНК
Пластид ДНК из кукуруза всходы подвержены повышенному повреждению по мере развития всходов.[12] ДНК повреждается в окислительной среде, созданной фотоокислительные реакции и фотосинтетический /дыхательный перенос электронов. Некоторые молекулы ДНК отремонтированный в то время как ДНК с неисправленными повреждениями, по-видимому, распадается на нефункциональные фрагменты.
Ремонт ДНК белки кодируются ядерным геном но могут быть перемещены в пластиды, где они поддерживают геном стабильность / целостность за счет восстановления ДНК пластиды.[13] Например, в хлоропласты из мха Physcomitrella patens, белок, используемый в репарации ошибочного спаривания ДНК (Msh1), взаимодействует с белками, используемыми в рекомбинационной репарации (RecA и RecG) для поддержания стабильности пластидного генома.[14]
Происхождение
Пластиды считаются эндосимбиотический цианобактерии. Предполагается, что это первичное эндосимбиотическое событие произошло около 1,5 миллиарда лет назад.[15] и позволил эукариотам выполнять кислородный фотосинтез.[16] С тех пор возникли три эволюционных линии, в которых пластиды названы по-разному: хлоропласты в зеленые водоросли и растения, родопласты в красных водорослях и муропласты у глаукофитов. Пластиды различаются как по пигментации, так и по ультраструктуре. Например, хлоропласты растений и зеленые водоросли потеряли все фикобилисомы, то легкие уборочные комплексы содержится в цианобактериях, красных водорослях и глаукофитах, но вместо этого содержат строму и грану тилакоиды. Пластида глаукоцистофека - в отличие от хлоропластов и родопластов - все еще окружена остатками клеточной стенки цианобактерий. Все эти первичные пластиды окружены двумя мембранами.
В отличие от первичных пластид, происходящих из первичного эндосимбиоза прокариоктических цианобактерий, сложные пластиды, происходящие из вторичных эндосимбиоз в котором эукариотический организм поглотил другой эукариотический организм, содержащий первичную пластиду. [17] Когда эукариот поглощает красную или зеленую водоросль и удерживает пластиду водорослей, эта пластида обычно окружена более чем двумя мембранами. В некоторых случаях у этих пластид может быть снижена их метаболическая и / или фотосинтетическая способность. Водоросли со сложными пластидами, образованными вторичным эндосимбиозом красной водоросли, включают гетероконты, гаптофиты, криптомонады, и большинство динофлагелляты (= родопласты). Те, которые эндосимбиозировали зеленые водоросли, включают эвглениды и хлорарахниофиты (= хлоропласты). В Apicomplexa, тип облигатных паразитарных простейших, включая возбудителей малярии (Плазмодий виды), токсоплазмоз (Toxoplasma gondii ), и многие другие болезни человека или животных также содержат сложную пластиду (хотя эта органелла была потеряна в некоторых апикомплексах, таких как Криптоспоридиум парвум, что приводит к криптоспоридиоз ). 'апикопласт 'больше не способен к фотосинтезу, но является важной органеллой и многообещающей мишенью для разработки противопаразитарных препаратов.
Немного динофлагелляты и морские слизни, в частности из рода Элизия, употребляйте водоросли в пищу и сохраняйте пластиду переваренных водорослей, чтобы получать выгоду от фотосинтеза; через некоторое время перевариваются и пластиды. Этот процесс известен как клептопластика, от греческого, клептес, вор.
Цикл развития пластид

В 1977 году Дж. М. Уатли предложил цикл разработки пластид, в котором говорилось, что развитие пластид не всегда однонаправлено, а является циклическим процессом несколько раз. Проплатиды являются предшественниками более дифференцированных форм пластид, как показано на изображенной диаграмме.[18]
Паулинелла хроматофора
Паулинелла имеет похожую органеллу, которая не принадлежит к пластидам, обсуждаемым в этом тексте. Это хроматофор, который представляет собой недавно приобретенную эндосимбиотическую β-цианобактерию.[19] Это событие эндосимбиоза произошло намного позже и является лишь вторым известным первичным событием эндосимбиоза цианобактерий.
Смотрите также
использованная литература
- ^ Сато Н. (2006). «Происхождение и эволюция пластидов: геномный взгляд на объединение и разнообразие пластидов». В Wise RR, Hoober JK (ред.). Строение и функции пластидов. Достижения в фотосинтезе и дыхании. 23. Springer Нидерланды. С. 75–102. Дои:10.1007/978-1-4020-4061-0_4. ISBN 978-1-4020-4060-3.
- ^ Мур KR, Magnabosco C, Momper L, Gold DA, Bosak T, Fournier GP (2019). «Расширенная рибосомная филогения цианобактерий поддерживает глубокое размещение пластидов». Границы микробиологии. 10: 1612. Дои:10.3389 / fmicb.2019.01612. ЧВК 6640209. PMID 31354692.
- ^ Врис, Ян де; Гулд, Свен Б. (15.01.2018). «Монопластидное узкое место в эволюции водорослей и растений». Журнал клеточной науки. 131 (2): jcs203414. Дои:10.1242 / jcs.203414. ISSN 0021-9533. PMID 28893840.
- ^ Колаттукуды, П. (1996) «Биосинтетические пути кутина и восков и их чувствительность к стрессам окружающей среды», стр. 83–108 в: Кутикула растений. Г. Керстиенс (редактор), BIOS Scientific publishers Ltd., Оксфорд
- ^ а б Мудрый, Роберт Р. (2006). «1. Разнообразие форм и функций пластидов». Достижения в области фотосинтеза и дыхания. 23. Springer. С. 3–26. Дои:10.1007/978-1-4020-4061-0_1. ISBN 978-1-4020-4060-3.
- ^ Wicke, S; Schneeweiss, GM; деПамфилис, CW; Мюллер, К.Ф .; Квандт, Д. (2011). «Эволюция пластидной хромосомы у наземных растений: содержание генов, порядок генов, функции генов». Молекулярная биология растений. 76 (3–5): 273–297. Дои:10.1007 / s11103-011-9762-4. PMID 21424877.
- ^ Wicke, S; Науманн, Дж (2018). «Молекулярная эволюция пластидных геномов паразитических цветковых растений». Достижения в ботанических исследованиях. 85: 315–347. Дои:10.1016 / bs.abr.2017.11.014.
- ^ а б «Растения без пластидных геномов». Ученый. Получено 2015-09-26.
- ^ Барбрук А.С., Хоу С.Дж., Пертон С. (февраль 2006 г.). «Почему пластидные геномы сохраняются у нефотосинтезирующих организмов?». Тенденции в растениеводстве. 11 (2): 101–8. Дои:10.1016 / j.tplants.2005.12.004. PMID 16406301.
- ^ Виола Р., Ниваль П., Педерсен М. (июль 2001 г.). «Уникальные особенности метаболизма крахмала в красных водорослях». Ход работы. Биологические науки. 268 (1474): 1417–22. Дои:10.1098 / rspb.2001.1644. ЧВК 1088757. PMID 11429143.
- ^ Чжан Кью (март 2010 г.). «Почему у покрытосеменных возрождается двуродительская пластидная наследственность?». Журнал исследований растений. 123 (2): 201–6. Дои:10.1007 / s10265-009-0291-z. PMID 20052516. S2CID 5108244.
- ^ Кумар Р.А., Ольденбургский ди-джей, Бендич А.Дж. (декабрь 2014 г.). «Изменения в повреждении ДНК, молекулярной целостности и количестве копий пластидной ДНК и митохондриальной ДНК в процессе развития кукурузы». Журнал экспериментальной ботаники. 65 (22): 6425–39. Дои:10.1093 / jxb / eru359. ЧВК 4246179. PMID 25261192.
- ^ Ольденбургский ди-джей, Бендич AJ (2015). «Поддержание ДНК в пластидах и митохондриях растений». Границы науки о растениях. 6: 883. Дои:10.3389 / fpls.2015.00883. ЧВК 4624840. PMID 26579143.
- ^ Одахара М., Кишита Ю., Секин Ю. (август 2017 г.). «MSH1 поддерживает стабильность генома органелл и генетически взаимодействует с RECA и RECG в мхе Physcomitrella patens». Журнал растений. 91 (3): 455–465. Дои:10.1111 / tpj.13573. PMID 28407383.
- ^ Очоа де Альда Дж. А., Эстебан Р., Диаго М. Л., Хумард Дж. (Сентябрь 2014 г.). «Пластидный предок произошел от одной из основных ветвей цианобактерий». Nature Communications. 5: 4937. Bibcode:2014 НатКо ... 5.4937O. Дои:10.1038 / ncomms5937. PMID 25222494.
- ^ Хеджес С.Б., Блэр Дж. Э., Вентури М.Л., Шу Дж. Л. (январь 2004 г.). «Молекулярная шкала времени эволюции эукариотов и возникновения сложной многоклеточной жизни». BMC Эволюционная биология. 4: 2. Дои:10.1186/1471-2148-4-2. ЧВК 341452. PMID 15005799.
- ^ Чан CX, Bhattachary D (2010). «Происхождение пластидов». Природное образование. 3 (9): 84.
- ^ Уотли, Джин М. (1978). «Предлагаемый цикл взаимоотношений развития пластид». Новый Фитолог. 80 (3): 489–502. Дои:10.1111 / j.1469-8137.1978.tb01581.x. ISSN 0028-646X. JSTOR 2431207.
- ^ Марин, Биргер; Новак, Ева СМ; Глёкнер, Гернот; Мелконян, Майкл (2007). «Предок хроматофора Paulinella получил карбоксисомный оперон путем горизонтального переноса гена от Nitrococcus-like γ-proteobacterium». BMC Эволюционная биология. 7 (1): 85. Дои:10.1186/1471-2148-7-85. ЧВК 1904183. PMID 17550603.
дальнейшее чтение
- Hanson MR, Köhler RH. «Новый взгляд на структуру хлоропласта». Физиология растений онлайн. Архивировано из оригинал на 2005-06-14.
- Wycliffe P, Sitbon F, Wernersson J, Ezcurra I., Ellerström M, Rask L (октябрь 2005 г.). «Непрерывная экспрессия в листьях табака гомолога Brassica napus PEND блокирует дифференцировку пластид и развитие палисадных клеток». Журнал растений. 44 (1): 1–15. Дои:10.1111 / j.1365-313X.2005.02482.x. PMID 16167891.
- Бирки CW (2001). «Наследование генов в митохондриях и хлоропластах: законы, механизмы и модели» (PDF). Ежегодный обзор генетики. 35: 125–48. Дои:10.1146 / annurev.genet.35.102401.090231. PMID 11700280. Архивировано из оригинал (PDF) на 2010-06-22. Получено 2009-03-01.
- Чан С.Х., Бхаттачарья Д. (2010). «Истоки пластид». Природное образование. 3 (9): 84.
- Бхаттачарья Д., изд. (1997). Происхождение водорослей и их пластидов. Нью-Йорк: Springer-Verlag / Wein. ISBN 978-3-211-83036-9.
- Гулд С.Б., Валлер РФ, Макфадден Г.И. (2008). «Пластидная эволюция». Ежегодный обзор биологии растений. 59: 491–517. Дои:10.1146 / annurev.arplant.59.032607.092915. PMID 18315522. S2CID 30458113.
- Килинг П.Дж. (март 2010 г.). «Эндосимбиотическое происхождение, разнообразие и судьба пластид». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 365 (1541): 729–48. Дои:10.1098 / rstb.2009.0103. ЧВК 2817223. PMID 20124341.
внешние ссылки
- Транспластомные растения для биосдерживания (биологическое удержание трансгенов) - Дополнительный исследовательский проект по сосуществованию и отслеживанию цепочек поставок ГМ и не ГМ
- Древо жизни эукариоты
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||
| Авторитетный контроль |
|---|