Тургор давление - Turgor pressure
Тургор давление сила внутри клетки, которая толкает плазматическая мембрана против клеточная стенка.[1]
Его еще называют гидростатическое давление, и определяется как давление, измеряемое жидкостью, измеренное в определенной точке внутри нее, когда она находится в равновесии.[2] Обычно тургорное давление вызывается осмотический поток воды и происходит в растения, грибы, и бактерии. Явление наблюдается и в протисты которые имеют клеточные стенки.[3] Эта система не наблюдается в клетках животных, поскольку отсутствие клеточной стенки может привести к лизировать когда под слишком большим давлением.[4] Давление, оказываемое осмотическим потоком воды, называется вздутием живота. Это вызвано осмотическим потоком воды через селективно проницаемая мембрана. Осмотический поток воды через полупроницаемую мембрану - это когда вода перемещается из области с низкой концентрацией растворенного вещества в область с более высокой концентрацией растворенного вещества. У растений это влечет за собой перемещение воды из растворенного вещества с низкой концентрацией вне клетки в клетку. вакуоль.[5]
Механизм

Осмос - это процесс, при котором вода течет из области с низким растворенное вещество концентрации, в соседнюю область с более высокой концентрацией растворенного вещества до тех пор, пока не будет достигнуто равновесие между двумя областями.[6] Все клетки окружены липидный бислой клеточная мембрана, которая позволяет воде входить и выходить из клетки, а также ограничивает поток растворенных веществ. В гипертонический В растворах вода вытекает из ячейки, что уменьшает ее объем. Когда в гипотонический В растворе вода проникает в мембрану и увеличивает объем клетки. Когда в изотонический раствора вода поступает в ячейку и выходит из нее с равной скоростью.[4]
Уплотнение - это точка, в которой клеточная мембрана прижимается к клеточной стенке, то есть когда тургорное давление высокое. Когда клеточная мембрана имеет низкое тургорное давление, она дряблая. У растений это проявляется в виде увядших анатомических структур. Это более конкретно известно как плазмолиз.[7]

Объем и геометрия клетки влияют на величину тургорного давления и то, как оно может влиять на пластичность клеточной стенки. Исследования показали, что более мелкие клетки испытывают более сильное эластичное изменение по сравнению с более крупными клетками.[3]
Тургорное давление также играет ключевую роль в росте растительных клеток, когда клеточная стенка подвергается необратимому расширению из-за силы тургорного давления, а также структурных изменений в клеточной стенке, которые изменяют ее растяжимость.[8]
Тургорное давление растений
Тургорное давление внутри клеток регулируется осмосом, и это также вызывает расширение клеточной стенки во время роста. Помимо размера, жесткость клетки обусловлена еще и тургорным давлением; более низкое давление приводит к увядший клеточная или растительная структура (т.е. лист, стебель). Одним из механизмов регулирования тургорного давления в растениях является полупроницаемая мембрана, которая позволяет только некоторым растворенным веществам перемещаться в клетку и из нее, а также может поддерживать минимальное давление. Другие механизмы включают испарение, что приводит к потере воды и снижению опухоли клеток.[9] Тургорное давление также является важным фактором для переноса питательных веществ по растению. Клетки одного и того же организма могут иметь разное тургорное давление по всей структуре организма. В высшие растения, тургорное давление отвечает за апикальный рост таких вещей, как кончики корней[10] и пыльцевые трубки.[11]
Рассредоточение
Транспортные белки закачку растворенных веществ в клетку можно регулировать давлением тургора клетки. Более низкие значения позволяют увеличить перекачку растворенных веществ; что, в свою очередь, увеличивает осмотическое давление. Эта функция важна как реакция растений в условиях засухи.[12] (видя, как поддерживается тургорное давление), и для клеток, которым необходимо накапливать растворенные вещества (т.е. фрукты ).[13]
Цветущие и репродуктивные органы
Было зафиксировано, что лепестки Gentiana kochiana и Каланхоэ цветоносное цветение за счет изменчивого тургорного давления клеток на растения адаксиальный поверхность.[11] Во время таких процессов, как пыльник расхождение, было замечено, что сушка эндотеций клетки вызывают силу изгиба наружу, которая приводит к выбросу пыльцы. Это означает, что в этих структурах наблюдаются более низкие тургорные давления из-за того, что они обезвожены. Пыльцевые трубки клетки, которые удлиняются, когда пыльца приземляется на клеймо, на кончике запястья. Эти клетки довольно быстро растут за счет повышения тургорного давления. Эти клетки подвергаются росту на кончиках. Пыльцевая трубка лилий в процессе роста может иметь тургорное давление 0–21 МПа.[14]
Распространение семян

Во фруктах, таких как Impatiens parviflora, Оксалия ацетоселла и Экбаллий элатериум, тургорное давление - способ диспергирования семян.[15] В Экбаллий элатериум, или брызгающий огурец, тургорное давление плодов нарастает до такой степени, что они агрессивно отделяются от стебля, а семена и вода разбрызгиваются повсюду, когда плод падает на землю. Тургорное давление плодов колеблется от 0,003 до 1,0 МПа.[16]
Рост
Обычно считается, что действие тургорного давления на растяжимые клеточные стенки является движущей силой роста внутри клетки.[17] Повышение тургорного давления вызывает расширение клеток и расширение апикальных клеток, пыльцевых трубок и других структур растений, таких как кончики корней. Расширение клеток и повышение тургорного давления происходит из-за попадания внутрь распространение воды в клетку, а тургорное давление увеличивается за счет увеличения объема вакуолярный сок. Давление тургора растущей клетки корня может достигать 0,6 МПа, что более чем в три раза превышает давление в автомобильной шине. Эпидермальный клетки в лист может иметь давление от 1,5 до 2,0 МПа.[18] Поскольку растения могут работать при таком высоком давлении, это может объяснить, почему они могут расти через асфальт и другие твердые поверхности.[17]
Опухлость
Вздутие наблюдается в клетке, где клеточная мембрана прижимается к клеточной стенке. У некоторых растений их клеточные стенки разрыхляются быстрее, чем вода может пересечь мембрану, что приводит к клетке с более низким тургорным давлением.[3]
Устьица

Тургорное давление внутри устьиц регулирует, когда устьица могут открываться и закрываться, что влияет на скорость транспирации растения. Это также важно, потому что эта функция регулирует потерю воды внутри растения. Более низкое тургорное давление может означать, что в клетке низкая концентрация воды, и закрытие устьиц поможет сохранить воду. Благодаря высокому тургорному давлению устьица открыты для газообмена, необходимого для фотосинтеза.[9]
Мимоза стыдливая

Сделан вывод, что потеря тургорного давления в листьях Мимоза стыдливая отвечает за реакцию растения при прикосновении. Другие факторы, такие как изменение осмотического давления, протоплазматический сокращение и увеличение клеточного проницаемость наблюдалось влияние на этот ответ. Также зафиксировано, что тургорное давление различается в верхнем и нижнем Pulvinar клетки растения, а движение ионов калия и кальция по клеткам вызывает повышение тургорного давления. При прикосновении пульвинус активируется и источает сократительный белки, что в свою очередь увеличивает тургорное давление и закрывает листья растения.[19]
Функция в других таксонах
Как указывалось ранее, тургорное давление может быть обнаружено и у других организмов, помимо растений, и может играть большую роль в развитии, движении и природе этих организмов.
Грибы

У грибов тургорное давление рассматривается как важный фактор субстрат проникновение. У таких видов, как Saprolegnia ferax, Magnaporthe grisea и Aspergillus oryzae, наблюдается огромное давление тургора в их гифы. Исследование показало, что они могут проникать в такие вещества, как клетки растений и синтетические материалы, такие как поливинил хлорид.[20] При наблюдении за этим явлением отмечается, что инвазивный рост гиф происходит из-за тургорного давления вместе с коферментами, секретируемыми грибами для вторжения в указанные субстраты.[21] Рост гиф напрямую связан с тургорным давлением, и рост замедляется по мере снижения тургорного давления. В Magnaporthe grisea наблюдается давление до 8 МПа.[22]
Протисты
Некоторые протисты не имеют клеточных стенок и не могут испытывать тургорное давление. Эти несколько протистов используют свою сократительную вакуоль для регулирования количества воды в клетке. Клетки простейших избегают лизирования в растворах, используя вакуоль, которая выкачивает воду из клеток для поддержания осмотического равновесия.[23]
Животные
Тургорное давление не наблюдается в животное клетки потому что у них отсутствует клеточная стенка. В организмах с клеточными стенками клеточная стенка препятствует лизису клетки при высоких давлениях.[1]
Диатомеи
У диатомовых водорослей Heterokontophyta имеют полифилетический тургорустойчивые клеточные стенки. На протяжении жизненного цикла этих организмов тщательно контролируемое тургорное давление отвечает за расширение клеток и выделение сперматозоидов, но не за такие вещи, как щетинка рост.[24]
Цианобактерии
Газ-пылесос[проверять орфографию ] цианобактерии те, кто обычно отвечает за цветение воды. Они обладают способностью плавать из-за скопления газов внутри их вакуолей, а роль тургорного давления и его влияние на емкость этих вакуолей наблюдалась в различных научных работах.[25][26] Отмечено, что чем выше тургорное давление, тем меньше емкость газовых вакуолей у разных цианобактерий. Эксперименты, использованные для корреляции осмоса и тургорного давления в прокариоты были использованы, чтобы показать, как диффузия растворенных веществ в клетку влияет на тургорное давление внутри клетки.[27]
Измерения
При измерении тургорного давления растений необходимо учитывать многие вещи. Обычно утверждается, что полностью набухшие клетки имеют значение тургорного давления, равное значению тургорного давления клетки, а вялые клетки имеют значение, равное нулю или близкое к нему. Другие принимаемые во внимание клеточные механизмы включают: протопласт растворенные вещества в протопласте (растворенный потенциал), испарение нормы растения и напряжение клеточных стенок. Измерения ограничены в зависимости от используемого метода, некоторые из которых рассматриваются и объясняются ниже. Не все методы могут быть использованы для всех организмов из-за размера и других свойств. Например, диатомовые водоросли не имеет тех же свойств, что и растение, которое может ограничить то, что можно использовать для определения тургорного давления.[28]
Единицы
Единицы измерения тургорного давления не зависят от мер, используемых для его определения. Общие единицы включают бары, МПа, или ньютоны за квадратный метр. 1 бар равен 0,1 МПа.[29]
Методы
Уравнение водного потенциала
Тургорное давление можно определить по общему водный потенциал, Ψш, и осмотический потенциал, Ψs, известны в уравнении водного потенциала.[30] Эти уравнения используются для измерения общего водного потенциала растения с использованием таких переменных, как матричный потенциал, осмотический потенциал, потенциал давления, гравитационные эффекты и тургорное давление.[31] Взяв разницу между Ψs и Ψш, приводится значение тургорного давления. При использовании этого метода гравитационный и матричный потенциал считаются незначительными, поскольку их значения обычно либо отрицательны, либо близки к нулю.[30]
Техника под давлением
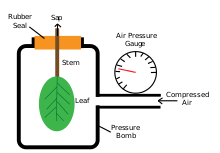
В бомба высокого давления Методика была разработана Шоландером и др., рассмотренная Тайри и Хаммелем в их публикации 1972 года, для проверки движения воды через растения. Этот прибор используется для измерения тургорного давления путем помещения листа (с прикрепленным стержнем) в закрытую камеру, куда добавляется газ под давлением. Измерения проводятся при ксилемный сок появляется из-за поверхности разреза и в месте, где он не накапливается и не отступает обратно в поверхность разреза.[32]
Атомно-силовой микроскоп
Атомно-силовые микроскопы использовать тип сканирующая зондовая микроскопия (SPM). Маленькие зонды вводятся в интересующую область, а пружина внутри зонда измеряет значения посредством смещения.[33] Этот метод можно использовать для измерения тургорного давления организмов. При использовании этого метода дополнительная информация, такая как уравнения механики сплошной среды, кривые глубины с одной силой и геометрию ячеек можно использовать для количественной оценки тургорного давления в заданной области (обычно в ячейке).
Датчик давления
Изначально эта машина использовалась для измерения индивидуальных водоросль клеток, но теперь может использоваться на образцах с более крупными клетками. Обычно используется на высшее растение ткани, но не использовались для измерения тургорного давления, пока Хюскен и Циммерман не усовершенствовали метод.[34] Датчики давления измеряют тургорное давление по смещению. Стеклянная микрокапиллярная трубка вставляется в ячейку, и все, что ячейка выделяет в трубку, наблюдается через микроскоп. Присоединенное устройство затем измеряет, какое давление требуется, чтобы выбросить излучение обратно в ячейку.[32]
Микроманипуляционный зонд
Они используются для точной количественной оценки более мелких ячеек. В эксперименте Вебера, Смита и его коллег отдельные клетки томата были сжаты между зондом для микроманипуляций и стеклом, чтобы позволить микрокапилляру зонда давления определить тургорное давление клетки.[35]
Теоретические рассуждения
Отрицательное тургорное давление
Было замечено, что значение Ψш уменьшается по мере обезвоживания клетки,[30] но ученые предположили, будет ли это значение продолжать уменьшаться, но никогда не упадет до нуля, или значение может быть меньше нуля. Были исследования[36][37] которые показывают, что отрицательное давление в ячейке может существовать в ксерофитный растений, но в статье М. Т. Тайри исследуется, возможно ли это, или делается вывод, основанный на неверно истолкованных данных. В своей статье он приходит к выводу, что, неправильно классифицируя «связанную» и «свободную» воду в клетке, исследователи, которые утверждали, что обнаружили отрицательные значения тургорного давления, оказались неверными. Анализируя изотермы апопластической и симпластной воды, он показывает, что отрицательное тургорное давление не может присутствовать в засушливых растениях из-за чистой потери воды образцом во время засухи. Несмотря на его анализ и интерпретацию данных, отрицательные значения тургорного давления все еще используются в научном сообществе.[38]
Рост кончиков у высших растений
Гипотеза, сформированная М. Гарольдом и его коллегами, предполагает, что рост кончиков на высших планах имеет амебную природу и не вызван тургорным давлением, как это широко распространено, а это означает, что расширение вызывается актиновым цитоскелетом в этих растительных клетках. Подразумевается, что регуляция роста клеток вызвана: цитоплазматический микротрубочки, которые контролируют ориентацию фибрилл целлюлозы, которые откладываются в соседней клеточной стенке и приводят к росту. У растений клетки окружены клеточными стенками и нитевидными белками, которые сохраняют и регулируют рост и форму растительной клетки. Как объясняется в статье, низшие растения растут за счет апикального роста, который отличается, поскольку клеточная стенка расширяется только на одном конце клетки.[39]
использованная литература
- ^ а б Причард, Джереми (2001). «Тургорное давление». Энциклопедия наук о жизни. Американское онкологическое общество. Дои:10.1038 / npg.els.0001687. ISBN 9780470015902. Отсутствует или пусто
| название =(Помогите) - ^ Фрике, Виланд (январь 2017 г.). «Тургорное давление». Энциклопедия наук о жизни: 1–6. Дои:10.1002 / 9780470015902.a0001687.pub2. ISBN 9780470015902.
- ^ а б c Steudle, Эрнст (февраль 1977 г.). «Влияние тургорного давления и размера клеток на эластичность стенок растительных клеток». Физиология растений. 59 (2): 285–9. Дои:10.1104 / стр.59.2.285. ЧВК 542383. PMID 16659835.
- ^ а б «Ханская академия». Ханская академия. Получено 2017-04-27.
- ^ «Осмос (клеточный) - Осмос в клетках растений». science.jrank.org. Получено 2017-04-27.
- ^ "GCSE Bitesize: Осмос в клетках". BBC.
- ^ «Плазмолиз в растительных клетках Elodea - Science NetLinks». sciencenetlinks.com. Получено 2017-04-27.
- ^ Джордан, Б.М., и Дюмэ, Дж. (2010). «Биомеханика роста растительных клеток». Энциклопедия наук о жизни.
- ^ а б Wagoner, Paul E .; Зелич, Израиль (1965-12-10). «Транспирация и устьица листьев». Наука. 150 (3702): 1413–1420. Bibcode:1965Sci ... 150.1413W. Дои:10.1126 / science.150.3702.1413. PMID 17782290.
- ^ Симадзаки, Юми; Оокава, Тайитиро; Хирасава, Тадаши (1 сентября 2005 г.). «Верхушка корня и ускоряющая область подавляют удлинение замедляющейся области без какого-либо воздействия на тургор клеток в первичных корнях кукурузы в условиях водного стресса». Физиология растений. 139 (1): 458–465. Дои:10.1104 / стр.105.062091. ЧВК 1203394. PMID 16100358.
- ^ а б Бозами, Лена; Накаяма, Наоми; Будауд, Арезки (01.11.2014). «Цветы под давлением: тонкости регуляции тургора в развитии». Анналы ботаники. 114 (7): 1517–1533. Дои:10.1093 / aob / mcu187. ЧВК 4204789. PMID 25288632.
- ^ Фишер, Дональд Б .; Кэш-Кларк, Кора Э. (27 апреля 2017 г.). «Градиенты водного потенциала и тургорного давления на пути транслокации при заполнении зерновых в нормально поливаемых и подверженных водному стрессу растений пшенице». Физиология растений. 123 (1): 139–148. Дои:10.1104 / стр.123.1.139. ЧВК 58989. PMID 10806232.
- ^ Келлер, Маркус; Шреста, Прадип М. (2014). «Накопление растворенного вещества отличается в вакуолях и апопласте созревающих ягод винограда». Planta. 239 (3): 633–642. Дои:10.1007 / s00425-013-2004-z. PMID 24310282. S2CID 15443543.
- ^ Бенкерт, Райнер; Обермейер, Герхард; Бентруп, Фридрих-Вильгельм (1 марта 1997 г.). «Тургорное давление растущих пыльцевых трубок лилии». Протоплазма. 198 (1–2): 1–8. Дои:10.1007 / BF01282125. S2CID 23911884.
- ^ Hayashi, M .; Feilich, K. L .; Эллерби, Д. Дж. (2009-05-01). "Механика взрывного рассеивания семян оранжевого драгоценного камня (Impatiens capensis)". Журнал экспериментальной ботаники. 60 (7): 2045–2053. Дои:10.1093 / jxb / erp070. ЧВК 2682495. PMID 19321647.
- ^ Козловский, Т.Т. (2012). Биология семян: важность, развитие и прорастание. 1. Академическая пресса. С. 195–196.
- ^ а б Kroeger, Jens H .; Зерзур, Рабах; Гайтманн, Аня (25 апреля 2011 г.). «Регулятор или движущая сила? Роль тургорного давления в колебательном росте растительных клеток». PLOS ONE. 6 (4): e18549. Bibcode:2011PLoSO ... 618549K. Дои:10.1371 / journal.pone.0018549. ЧВК 3081820. PMID 21541026.
- ^ Серпе, Марсело Д .; Мэтьюз, Марк А. (1 января 1994 г.). «Рост, давление и напряжение стенки в эпидермальных клетках листьев бегонии argenteo- guttata L. во время развития». Международный журнал наук о растениях. 155 (3): 291–301. Дои:10.1086/297168. JSTOR 2475182. S2CID 84209016.
- ^ Аллен, Роберт Д. (1969-08-01). «Механизм сейсмонастической реакции у Mimosa pudica1». Физиология растений. 44 (8): 1101–1107. Дои:10.1104 / стр.44.8.1101. ЧВК 396223. PMID 16657174.
- ^ Ховард, Ричард (декабрь 1991 г.). «Проникновение грибка в твердые субстраты с огромным тургорным давлением». Proc. Natl. Акад. Наука. 88 (24): 11281–11284. Bibcode:1991PNAS ... 8811281H. Дои:10.1073 / пнас.88.24.11281. ЧВК 53118. PMID 1837147.
- ^ Жерве, Патрик; Абади, Кристоф и Молин, Пол (1999). «Тургорное давление грибковых клеток: теоретический подход и измерение». Журнал научных и промышленных исследований. 58 (9): 671–677.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Деньги, Николас П. (1995-12-31). «Тургорное давление и механика проникновения грибков». Канадский журнал ботаники. 73 (S1): 96–102. Дои:10.1139 / b95-231.
- ^ "Пирсон - место биологии". www.phschool.com. Получено 2017-04-27.
- ^ Raven, J. A .; Уэйт, А. М. (2004-04-01). «Эволюция окварцевания диатомовых водорослей: неизбежное опускание и опускание как побег?». Новый Фитолог. 162 (1): 45–61. Дои:10.1111 / j.1469-8137.2004.01022.x.
- ^ Родственник, Р. (январь 1991 г.). «Коллапс газовых пузырьков под действием тургорного давления и его роль в регулировании плавучести с помощью Anabaena flos-aquae". Журнал общей микробиологии. 143 (3): 1171–1178. Дои:10.1099/00221287-137-5-1171.
- ^ Reed, R.H .; Уолсби, А. Э. (1985-12-01). «Изменения тургорного давления в ответ на увеличение внешней концентрации NaCl в газо-вакуолизированной цианобактерии. Микроцистис sp ". Архив микробиологии. 143 (3): 290–296. Дои:10.1007 / BF00411252. S2CID 25006411.
- ^ Оливер, Родерик Льюис (1994-04-01). «Плавающие и тонущие в цианобактериях с газовым вакуумом1». Журнал психологии. 30 (2): 161–173. Дои:10.1111 / j.0022-3646.1994.00161.x. S2CID 83747596.
- ^ Tomos, A.D .; Leigh, R.A .; Shaw, C.A .; Джонс, Р. Г. У. (1984-11-01). «Сравнение методов измерения тургорного и осмотического давления клеток ткани хранения красной свеклы». Журнал экспериментальной ботаники. 35 (11): 1675–1683. Дои:10.1093 / jxb / 35.11.1675.
- ^ «Что такое единица давления« бар »(б)». www.aqua-calc.com. Получено 2017-04-27.
- ^ а б c Крамер, Пол (2012). Водные отношения растений. Elsevier Science. ISBN 978-0124250406. OCLC 897023594.
- ^ Безграничный (2016-05-26). «Давление, гравитация и матричный потенциал». Безграничный.
- ^ а б Тайри, М. Т .; Хаммель, Х. Т. (1972). «Измерение давления тургора и водные отношения растений под давлением бомбы Technique». Журнал экспериментальной ботаники. 23 (1): 267–282. Дои:10.1093 / jxb / 23.1.267.
- ^ Боузамы, Лена (май 2015). «Количественная оценка гидростатического давления в клетках растений с помощью индентирования с помощью атомно-силового микроскопа». Биофизический журнал. 108 (10): 2448–2456. Bibcode:2015BpJ ... 108.2448B. Дои:10.1016 / j.bpj.2015.03.035. ЧВК 4457008. PMID 25992723.
- ^ Хюскен, Дитер; Steudle, Эрнст; Циммерманн, Ульрих (1978-02-01). «Техника датчика давления для измерения водного отношения клеток высших растений». Физиология растений. 61 (2): 158–163. Дои:10.1104 / стр.61.2.158. ЧВК 1091824. PMID 16660252.
- ^ Вебер, Ален; Брейбрук, Шивон; Хуфлейт, Михал; Моска, Габриэлла; Рутье-Кежковская, Анн-Лиз; Смит, Ричард С. (01.06.2015). «Измерение механических свойств растительных клеток путем сочетания микровыступов с осмотической обработкой». Журнал экспериментальной ботаники. 66 (11): 3229–3241. Дои:10.1093 / jxb / erv135. ЧВК 4449541. PMID 25873663.
- ^ Ян, Дунмэй; Ли, Цзюньхуэй; Дин, Итин; Тайри, Мелвин Т. (2017-03-01). «Экспериментальные доказательства отрицательного тургорного давления в маленьких клетках листа Robinia pseudoacacia L по сравнению с большими клетками Metasequoia glyptostroboides Hu et W.C. Cheng. 2. Диаграммы Хефлера ниже нулевого тургора и теоретическое значение для кривых давление-объем живых клеток». Растения, клетки и окружающая среда. 40 (3): 340–350. Дои:10.1111 / шт.12860. PMID 27861986.
- ^ Oertli, J.J. (Июль 1986 г.). «Влияние размера клеток на коллапс клеток при отрицательном тургорном давлении». Журнал физиологии растений. 124 (3–4): 365–370. Дои:10.1016 / S0176-1617 (86) 80048-7.
- ^ Тайри, М. (январь 1976 г.). «Отрицательное тургорное давление в растительных клетках: факт или заблуждение?». Канадский журнал ботаники. 54 (23): 2738–2746. Дои:10.1139 / b76-294.
- ^ Пикетт-Хипс, Дж. Д., Кляйн, А. Г. (1998). «Рост кончиков в растительных клетках может быть амебовидным и не вызван тургорным давлением». Труды: Биологические науки.. 265 (1404): 1453–1459. Дои:10.1098 / rspb.1998.0457. ЧВК 1689221.CS1 maint: несколько имен: список авторов (ссылка на сайт)
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||