Спора - Spore
эта статья нужны дополнительные цитаты для проверка. (Июль 2007 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |


В биология, а спора это единица сексуальный или бесполое размножение который может быть адаптирован для рассредоточение и для выживания, часто в течение продолжительных периодов времени, в неблагоприятных условиях. Споры входят в состав жизненные циклы из многих растения, водоросли, грибы и простейшие.[1] Бактериальные споры не являются частью полового цикла, но представляют собой устойчивые структуры, используемые для выживания в неблагоприятных условиях. Миксозойский Споры выпускают амебулы в своих хозяев для паразитарной инфекции, но также размножаются внутри хозяев за счет спаривания двух ядер внутри плазмодия, который развивается из амебулы.[2]
Споры обычно гаплоидный и одноклеточный и производятся мейоз в спорангий из диплоид спорофит. При благоприятных условиях спора может развиться в новый организм с помощью митотический подразделение, производящее многоклеточный гаметофит, который в конечном итоге производит гаметы. Две гаметы сливаются, образуя зигота который перерастает в новый спорофит. Этот цикл известен как смена поколений.
Споры семенные растения продуцируются внутри, а мегаспоры (образующиеся внутри семяпочек) и микроспоры участвуют в формировании более сложных структур, которые образуют единицы распределения, семена и пыльца зерна.
Определение
Период, термин спора происходит от древнегреческий слово σπορά спора, смысл "семя, посев », относящийся к σπόρος спор, "посев" и σπείρειν Speirein, "сеять".
В просторечии разница между «спорой» и «гамета "состоит в том, что спора прорастет и превратится в спорлинг, в то время как гамете необходимо объединиться с другой гаметой, чтобы сформировать зиготу, прежде чем развиваться дальше.
Основное различие между спорами и семенами как единицы рассредоточения состоит в том, что споры одноклеточные, первая клетка гаметофита, а семена содержат внутри себя развивающийся зародыш (многоклеточный спорофит следующего поколения), образованный слиянием мужской гаметы пыльцевой трубки с женской гаметой, образованной мегагаметофит внутри семяпочки. Споры прорастают, давая начало гаплоидным гаметофитам, а семена прорастают, давая начало диплоидным спорофитам.
Классификация спорообразующих организмов
Растения
Сосудистое растение споры всегда гаплоидный. Сосудистые растения бывают либо гомоспористый (или изоспористый) или гетероспористый. Гомоспористые растения производят споры одного размера и типа.
Гетероспористые растения, такие как семенные растения, спайкмосы, перья, и папоротники отряда Сальвиниалес производят споры двух разных размеров: более крупная спора (мегаспора) действует как «женская» спора, а меньшая (микроспора) действует как «мужская». Такие растения обычно дают начало двум видам спор из отдельных спорангиев, либо мегаспорангий который производит мегаспоры или микроспорангий который производит микроспоры. У цветковых растений эти спорангии встречаются в плодолистиках и пыльниках соответственно.
Грибы
Грибы обычно образуют споры в результате полового или бесполого размножения. Споры обычно гаплоидный и вырасти в зрелых гаплоидных особей через митотический деление клеток (Урединиоспоры и Телиоспоры среди ржавчин дикариотические). Дикариотический клетки возникают в результате слияния двух гаплоидных клеток гамет. Среди спорогенных дикариотических клеток кариогамия (слияние двух гаплоидных ядер) происходит с образованием диплоидной клетки. Диплоидные клетки подвергаются мейозу с образованием гаплоидных спор.
Классификация спор
Споры можно классифицировать несколькими способами:
По спорообразующей структуре
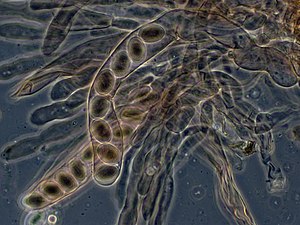


Грибы
У грибов и грибоподобных организмов споры часто классифицируются по структуре, в которой происходит мейоз и образование спор. Поскольку грибы часто классифицируют по их споровым структурам, эти споры часто характерны для определенных таксон из грибы.
- Спорангиоспоры: споры, продуцируемые спорангий во многих грибах, таких как зигомицеты.
- Зигоспоры: споры, продуцируемые зигоспорангий, характерная для зигомицеты.
- Аскоспоры: споры, продуцируемые аск, характерная для аскомицеты.
- Базидиоспоры: споры, продуцируемые базидиум, характерная для базидиомицеты.
- Эциоспоры: споры, продуцируемые эций в некоторых грибах, таких как ржавеет или грязь.
- Урединиоспоры: споры, продуцируемые урединиум в некоторых грибах, таких как ржавеет или грязь.
- Телиоспоры: споры, продуцируемые телий в некоторых грибах, таких как ржавеет или грязь.
- Ооспоры: споры, продуцируемые оогоний, характерная для оомицеты.
Красные водоросли
- Карпоспоры: споры, продуцируемые карпоспорофит, характерная для красные водоросли.
- Тетраспоры: споры, продуцируемые тетраспорофит, характерная для красные водоросли.
По функциям
- Хламидоспоры: покоящиеся толстостенные споры грибы произведены, чтобы выжить в неблагоприятных условиях.
- Паразитарные споры грибов можно разделить на внутренние споры, которые прорастают внутри хозяина, и внешние споры, также называемые спорами окружающей среды, высвобождаемые хозяином для заражения других хозяев.[3]
По происхождению в течение жизненного цикла
- Мейоспоры: споры, продуцируемые мейоз; они таким образом гаплоидный, и дают начало гаплоидным дочерним клеткам или гаплоидным индивидуумам. Примеры - клетки-предшественники гаметофиты из семенные растения нашел в цветы (покрытосеменные ) или шишки (голосеменные ), а зооспоры образуется в результате мейоза спорофитов водорослей, таких как Ульва.
- Микроспоры: мейоспоры, дающие начало самцу гаметофит, (пыльца в семенниках).
- Мегаспоры (или макроспоры): мейоспоры, которые дают начало самке гаметофит, (у семенных растений гаметофит образуется в яйцеклетка ).
- Митоспоры (или конидии, конидиоспоры): споры, продуцируемые митоз; они характерны для Аскомицеты. Грибы, в которых обнаружены только митоспоры, называются «митоспорическими грибами» или «анаморфными грибами» и ранее классифицируются как таксон Deuteromycota (Увидеть Телеоморф, анаморф и голоморф ).
По мобильности
Споры можно различить по тому, могут ли они шаг или нет.
- Зооспоры: подвижные споры, которые перемещаются с помощью одного или нескольких жгутики, и его можно найти в некоторых водоросли и грибы.
- Апланоспоры: неподвижные споры, которые, тем не менее, могут потенциально разрастать жгутики.
- Автоспоры: неподвижные споры, которые не могут развить жгутики.
- Баллистоспоры: споры, которые принудительно выводятся или выбрасываются из грибка плодовое тело в результате действия внутренней силы, например повышения давления. Наиболее базидиоспоры также являются баллистоспорами, и еще одним ярким примером являются споры рода Пилоболус.
- Статизмоспоры: споры, которые выходят из плодового тела грибов в результате воздействия внешней силы, например, капель дождя или проходящего мимо животного. Примеры клубнички.
Внешняя анатомия


Под высоким увеличение, споры часто имеют сложный рисунок или орнамент на своей внешней поверхности. Для описания особенностей таких паттернов была разработана специальная терминология. Некоторые отметки представляют собой отверстия, места, где жесткая внешняя оболочка спор может проникнуть во время прорастания. Споры можно разделить на категории в зависимости от положения и количества этих отметин и отверстий. Споры Alete не показывать линий. В монолетные споры, на споре имеется единственная узкая линия (laesura).[4] Это указывает на то, что материнская спора разделена на четыре части по вертикальной оси.[нужна цитата ] В трилете споры, каждая спора показывает три узкие линии, исходящие от центрального полюса.[4] Это показывает, что четыре споры имели общее происхождение и первоначально находились в контакте друг с другом, образуя тетраэдр.[нужна цитата ] Более широкое отверстие в форме канавки можно назвать кольп.[4] По количеству кольпи различают основные группы растений. Eudicots имеют трехслойный споры (то есть споры с тремя кольпи).[5]
Споры тетрады и трилете споры
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Август 2008 г.) |
Тетрады спор в оболочке считаются самым ранним свидетельством существования растений на суше.[6] датируется серединой ордовика (ранний Лланвирн, ~470 миллион лет назад), период, из которого еще не были извлечены макрофоссилии.[7]Отдельные споры трилете, напоминающие споры современных криптогамный растения впервые появились в летописи окаменелостей в конце ордовикского периода.[8]
Рассредоточение

У грибов как бесполые, так и половые споры или спорангиоспоры многих видов грибов активно распространяются путем насильственного выброса из их репродуктивных структур. Этот выброс обеспечивает выход спор из репродуктивных структур, а также их перемещение по воздуху на большие расстояния. Таким образом, многие грибы обладают специализированными механическими и физиологическими механизмами, а также поверхностными структурами спор, такими как гидрофобины, для выброса спор. Эти механизмы включают, например, насильственное удаление аскоспор, обусловленное структурой асков, и накопление осмолиты в жидкостях аска, что приводит к взрывному выбросу аскоспор в воздух.[9]
Насильственное удаление отдельных спор, называемое баллистоспоры предполагает образование небольшой капли воды (Капля Буллера ), который при контакте со спорой приводит к высвобождению ее снаряда с начальным ускорением более 10000 г.[10] Другие грибы полагаются на альтернативные механизмы высвобождения спор, такие как внешние механические силы, примером которых является клубнички. Привлечение насекомых, таких как мухи, к плодоносящим структурам, благодаря их яркой окраске и гнилостному запаху, для распространения спор грибов - еще одна стратегия, наиболее широко используемая вонючие рога.
В мохе обыкновенном Smoothcap (Atrichum undulatum ), было показано, что вибрация спорофита является важным механизмом высвобождения спор.[11]
В случае распространения спор сосудистые растения такие как папоротники, распространение очень легких спор ветром обеспечивает большую способность к распространению. Кроме того, споры менее подвержены хищничеству животных, чем семена, потому что они почти не содержат запасов пищи; однако они более подвержены хищничеству грибков и бактерий. Их главное преимущество состоит в том, что из всех форм потомства для образования спор требуется меньше всего энергии и материалов.
В мохе Селагинелла лепидофилла, рассредоточение частично достигается за счет необычного типа диаспора, а перекати-поле.[12]
Галерея

Споры мох Bartramia ithyphylla. (вид под микроскопом, 400x)

Расщепленные спорангии папоротника. (вид под микроскопом, спор не видно)

Споры и ловушки хвоща. (Equisetum, микроскопический вид)

Ископаемые споры растений (Сциласпора ) из силурийских отложений Швеции.

Плесень плода со спорами и заметным клеточным ростом. (2000x)

Скопления спор, образующиеся внутри спорангиев слизистой плесени. Ретикулярия оливковая, из сосновых лесов восточной Украина.

Внутренняя поверхность перидиум слизи Tubifera dudkae со спорами.






Смотрите также
использованная литература
- ^ Интернет-проект "Древо жизни". В архиве из оригинала 5 февраля 2018 г.. Получено 5 февраля 2018.
- ^ "Myxozoa В архиве 16 февраля 2015 г. Wayback Machine ". Интернет-проект" Древо жизни ". Иван Фиала, 10 июля 2008 г., Интернет. 14 января 2014 г.
- ^ "Microsporidia (Protozoa): Справочник по биологии и методам исследования". Архивировано 26 июня 2008 года.. Получено 8 июля 2007.CS1 maint: BOT: статус исходного URL-адреса неизвестен (ссылка на сайт). modares.ac.ir
- ^ а б c Punt, W .; Hoen, P.P .; Blackmore, S .; Нильссон, С. & Ле Томас, А. (2007). «Словарь терминов по пыльце и спорам». Обзор палеоботаники и палинологии. 143 (1): 1–81. Дои:10.1016 / j.revpalbo.2006.06.008.
- ^ Джадд, Уолтер С. и Олмстед, Ричард Г. (2004). «Обзор филогенетических взаимоотношений триколпата (эвдикота)». Американский журнал ботаники. 91 (10): 1627–44. Дои:10.3732 / ajb.91.10.1627. PMID 21652313.
- ^ Gray, J .; Chaloner, W. G .; Вестолл, Т. С. (1985). «Летопись микрофоссилий ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984». Философские труды Королевского общества B. 309 (1138): 167–195. Bibcode:1985РСПТБ.309..167Г. Дои:10.1098 / рстб.1985.0077. JSTOR 2396358.
- ^ Веллман CH, Грей J (2000). «Летопись микрофоссилий ранних наземных растений». Философские труды Королевского общества B. 355 (1398): 717–732. Дои:10.1098 / rstb.2000.0612. ЧВК 1692785. PMID 10905606.
- ^ Steemans, P .; Herisse, A. L .; Мелвин, Дж .; Миллер, М. А .; Paris, F .; Verniers, J .; Веллман, К. Х. (2009). «Происхождение и радиация древнейших сосудистых растений суши» (PDF). Наука. 324 (5925): 353. Bibcode:2009Sci ... 324..353S. Дои:10.1126 / science.1169659. ISSN 0036-8075. PMID 19372423. S2CID 206518080. В архиве (PDF) из оригинала 22 сентября 2017 г.. Получено 1 ноября 2017.
- ^ Трейл Ф. (2007). «Грибковые пушки: взрывные разряды спор в Ascomycota». Письма о микробиологии FEMS. 276 (1): 12–8. Дои:10.1111 / j.1574-6968.2007.00900.x. PMID 17784861.
- ^ Прингл А., Патек С. Н., Фишер М., Штольце Дж., Деньги Н. П. (2005). «Захваченный запуск баллистоспоры». Микология. 97 (4): 866–71. Дои:10.3852 / mycologia.97.4.866. PMID 16457355.
- ^ Йоханссон, Лённелл, Сандберг и Хиландер (2014) Пороги выброса спор мха: важность турбулентности и длины спорофита. Журнал экологии, н / д-н / д.
- ^ "Ложная роза Иерихона - Selaginella lepidophyllaЛожная роза Иерихона - Selaginella lepidophylla". Гид по растениям и цветам. Февраль 2009 г. В архиве из оригинала 15 июля 2011 г.. Получено 1 февраля 2010.
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||