Эндосперм - Endosperm
В эндосперм ткань, произведенная внутри семена из большинства цветущие растения после оплодотворения. это триплоид (имеется в виду три набора хромосом на ядро) у большинства видов.[1] Он окружает эмбрион и обеспечивает питание в виде крахмал, хотя он также может содержать масла и белок. Это может сделать эндосперм источником питания в рационе животных. Например, пшеница эндосперм измельчают в муку для хлеб (остальное зерно также входит в цельнозерновая мука ), в то время как ячмень эндосперм является основным источником сахаров для пиво производство. Другими примерами эндосперма, составляющего основную часть съедобной части, являются: кокос «мясо» и кокосовая «вода»,[2] и кукуруза. Некоторые растения, такие как орхидеи, отсутствие эндосперма в их семена.
Происхождение эндосперма
У предковых цветковых растений есть семена с маленькими зародышами и обильным эндоспермом, а эволюционное развитие цветущих растений имеет тенденцию к появлению растений со зрелыми семенами с небольшим количеством эндосперма или без него. У более производных цветковых растений зародыш занимает большую часть семян, а эндосперм не развивается или потребляется до созревания семян.[3][4]
Двойное оплодотворение
Эндосперм образуется, когда два сперма ядра внутри пыльца зерна достигают внутренней части женского гаметофита (иногда называемого зародышевый мешок ). Одно ядро спермы оплодотворяет Клетка яйца, образуя зигота, в то время как другое ядро сперматозоида обычно сливается с двуядерной центральной клеткой, образуя первичную клетку эндосперма (его ядро часто называют ядро тройного слияния). Эта ячейка, созданная в процессе двойное оплодотворение развивается в эндосперм. Поскольку он образуется в результате отдельного оплодотворения, эндосперм представляет собой организм, отдельный от растущего эмбриона.
Около 70% видов покрытосеменных имеют клетки эндосперма, которые полиплоид.[5] Обычно это триплоид (содержащий три набора хромосомы ), но может значительно отличаться от диплоид (2н) до 15н.[6]
Один вид цветущего растения, Nuphar полисепала, было показано, что эндосперм является диплоидным, возникающим в результате слияния ядра пыльцы с одним, а не двумя материнскими ядрами. То же можно сказать и о некоторых других базальных покрытосеменных.[7] Считается, что на раннем этапе развития линий покрытосеменных в этом способе размножения имело место дупликация с образованием семиклеточных / восьмиядерных женских гаметофитов и триплоидных эндоспермов с соотношением материнского генома к отцовскому геному 2: 1.[8]
Двойное оплодотворение - характерная черта покрытосеменные.
Он используется во многих биотехнологические процессы
Формирование эндосперма
Существует три типа развития эндосперма:
Формирование ядерного эндосперма - где имеют место повторные свободные ядерные деления; если клеточная стенка образуется, она образуется после делений свободных ядер. Обычно называется жидким эндоспермом. Кокосовая вода является примером этого.
Формирование клеточного эндосперма - где клеточная стенка формирование совпадает с ядерными подразделениями. Кокосовое мясо - это клеточный эндосперм. Acoraceae имеет клеточный эндосперм, в то время как другие однодольные являются гелобиальными.
Образование гелобиального эндосперма - Когда клеточная стенка закладывается между первыми двумя ядрами, после чего одна половина развивает эндосперм по клеточному образцу, а другая половина - по ядерному образцу.
Эволюционное происхождение
Эволюционное происхождение двойного оплодотворения и эндосперма неясно, что привлекает внимание исследователей уже более века. Есть две основные гипотезы:[6]
- Изначально двойное оплодотворение использовалось для получения двух идентичных, независимых эмбрионов («близнецов»). Позже эти зародыши приобрели разные роли: один врастал в зрелый организм, а другой просто поддерживал его. Таким образом, ранний эндосперм, вероятно, был диплоидным, как и эмбрион. Немного голосеменные, такие как Эфедра, может произвести двойные эмбрионы при двойном оплодотворении. Любой из этих двух эмбрионов способен заполнить семя, но обычно только один развивается дальше (второй в конечном итоге прерывается). Кроме того, большинство базальных покрытосеменных все еще содержат четырехклеточный зародышевый мешок и продуцируют диплоидные эндоспермы.
- Эндосперм - это эволюционный остаток настоящего гаметофит, подобный сложным многоклеточным гаметофитам голосеменных растений. В этом случае получение дополнительного ядра от сперматозоида является более поздним этапом эволюции. Это ядро может обеспечивать родительский (не только материнский) организм некоторым контролем над развитием эндосперма. Превращение в триплоид или полиплоид - это более поздние этапы эволюции этого «первичного гаметофита». Нецветущие семенные растения (хвойные деревья, саговники, гинкго, эфедра) образуют большой гомозиготный женский гаметофит, который питает зародыш внутри семени.[9]
Роль эндосперма в развитии семян
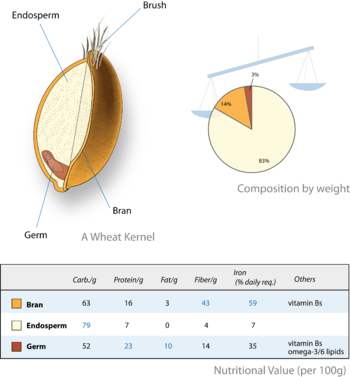
В некоторых группах (например, зерна семейства Poaceae ) эндосперм сохраняется до стадии зрелого семени в качестве запасающей ткани, и в этом случае семена называют «белковыми» или «эндоспермными», а в других случаях он абсорбируется во время развития эмбриона (например, большинство членов семейства Fabaceae, в том числе фасоль, Phaseolus vulgaris), и в этом случае семена называют «экзальбуминозными» или «семядольными», а функцию запасающей ткани выполняет увеличенная семядоли («семенные листья»). У некоторых видов (например, кукурузы, Zea Mays); функция хранения распределяется как между эндоспермом, так и между эмбрионом. Некоторые зрелые ткани эндосперма хранят жиры (например, клещевина, Ricinus communis) и другие (включая зерновые, такие как пшеница и кукуруза) хранят в основном крахмалы.
Пыльные семена орхидеи не имеют эндосперма. Саженцы орхидей микогетеротрофный в их раннем развитии. У некоторых других видов, таких как кофе, эндосперм также не развивается.[10] Вместо этого нуцеллус производит питательную ткань, называемую «перисперм». Эндосперм некоторых видов отвечает за покой семян.[11] Ткань эндосперма также опосредует перенос питательных веществ от материнского растения к эмбриону, она действует как место для импринтинга генов и отвечает за абортирование семян, полученных от генетически несовместимых родителей.[5] У покрытосеменных эндосперм содержит такие гормоны, как цитокинины, которые регулируют клеточную дифференцировку и формирование эмбриональных органов.[12]
Злаки
Крупа культуры выращиваются для их съедобных фрукты (зерна или зерновки ), которые в первую очередь являются эндоспермом. У зерновки тонкая стенка плода срастается с семенной оболочкой. Следовательно, питательной частью зерна является семя и его эндосперм. В некоторых случаях (например, пшеница, рис) эндосперм избирательно задерживается при переработке пищевых продуктов (обычно это называется белая мука ), а эмбрион (зародыш ) и семенной оболочки (отруби ) удалено. Переработанное зерно имеет более низкую питательную ценность. Таким образом, эндосперм играет важную роль в рационе человека во всем мире.
В алейрон представляет собой внешний слой клеток эндосперма, присутствующий во всех мелких зернах и сохраняющийся во многих двудольных с временным эндоспермом. Зерновой алейрон служит как для хранения, так и для пищеварения. Во время прорастания выделяет амилаза фермент, который расщепляет крахмал эндосперма на сахара для питания растущих проростков.[13][14]
Смотрите также
использованная литература
- ^ Стюарт-Кокс Дж. А., Бриттон Н. Ф., Моги М (август 2004 г.). «Триплоидия эндосперма имеет избирательное преимущество во время продолжающегося родительского конфликта посредством импринтинга». Ход работы. Биологические науки. 271 (1549): 1737–43. Дои:10.1098 / rspb.2004.2783. ЧВК 1691787. PMID 15306295.
- ^ «Съедобные пальмовые фрукты». Слово Уэйна: онлайн-учебник естественной истории. Получено 14 июля 2010.
- ^ "Место биологии семян - покой семян". Seedbiology.de. Получено 2014-02-05.
- ^ Фридман В.Е. (1998), «Эволюция двойного оплодотворения и эндосперма:« историческая »перспектива», Половое размножение растений, 11: 6, Дои:10.1007 / s004970050114, S2CID 19785565
- ^ а б Олсен О. (2007). Эндосперм: эволюционная и молекулярная биология. ISBN 9783540712350.
- ^ а б Baroux C, Spillane C, Grossniklaus U (август 2002 г.). «Эволюционное происхождение эндосперма цветковых растений». Геномная биология. 3 (9): отзывы1026. Дои:10.1186 / gb-2002-3-9-reviews1026. ЧВК 139410. PMID 12225592.
- ^ Уильямс Дж. Х., Фридман В. Е. (январь 2002 г.). «Идентификация диплоидного эндосперма в ранней линии покрытосеменных». Природа. 415 (6871): 522–6. Bibcode:2002Натурал.415..522Вт. Дои:10.1038 / 415522a. PMID 11823859. S2CID 4396197.
- ^ Фридман В.Е., Уильямс Дж. Х. (февраль 2003 г.). «Модульность женского гаметофита покрытосеменных и ее влияние на раннюю эволюцию эндосперма у цветковых растений». Эволюция; Международный журнал органической эволюции. 57 (2): 216–30. Дои:10.1111 / j.0014-3820.2003.tb00257.x. PMID 12683519. S2CID 24303275.
- ^ Фридман В.Е. (апрель 1995 г.). «Органическое дублирование, инклюзивная теория приспособленности и альтруизм: понимание эволюции эндосперма и репродуктивного синдрома покрытосеменных». Труды Национальной академии наук Соединенных Штатов Америки. 92 (9): 3913–7. Bibcode:1995PNAS ... 92.3913F. Дои:10.1073 / пнас.92.9.3913. ЧВК 42072. PMID 11607532.
- ^ Хук WG (1938). «Эндосперм и перисперм кофе с заметками о морфологии семяпочки и развитии семян». Американский журнал ботаники. 25 (1): 56–61. Дои:10.2307/2436631. JSTOR 2436631.
- ^ Басра А.С. (1994). Механизмы роста растений и повышение урожайности: современные подходы. Нью-Йорк: М. Деккер. ISBN 978-0-8247-9192-6.
- ^ Пирсон LC (1995). Разнообразие и эволюция растений. Бока-Ратон: CRC Press. п. 547. ISBN 978-0-8493-2483-3.
- ^ Becraft PW, Yi G (март 2011 г.). «Регуляция развития алейронов в зерновых культурах». Журнал экспериментальной ботаники. 62 (5): 1669–75. Дои:10.1093 / jxb / erq372. PMID 21109580.
- ^ Бекрафт П.В., Гутьеррес-Маркос Дж. (2012). «Развитие эндосперма: динамические процессы и клеточные инновации, лежащие в основе альтруизма братьев и сестер». Междисциплинарные обзоры Wiley: биология развития. 1 (4): 579–93. Дои:10.1002 / wdev.31. PMID 23801534. S2CID 5752973.
внешние ссылки
- Бич, Чендлер Б., изд. (1914). . . Чикаго: F. E. Compton and Co.
- Эндосперм: стержень полового конфликта у цветковых растений в Earthling Nature
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||