Биологический жизненный цикл - Biological life cycle


В биология, а биологический жизненный цикл (или просто жизненный цикл или же жизненный цикл когда биологический контекст ясен) представляет собой серию изменений формы, которые претерпевает организм, возвращаясь в исходное состояние. "Эта концепция тесно связана с историей жизни, разработка и онтогенез, но отличается от них тем, что подчеркивает обновление ».[1][2] Переход формы может включать рост, бесполое размножение, или же половое размножение.
У некоторых организмов разные «поколения» видов сменяют друг друга в течение жизненного цикла. За растения и много водоросли, есть две многоклеточные стадии, и жизненный цикл обозначается как смена поколений. Период, термин история жизни часто используется, особенно для таких организмов, как красные водоросли которые имеют три многоклеточных стадии (или больше), а не две.[3]
Жизненные циклы, включающие половое размножение, включают чередование гаплоидный (п) и диплоид (2п) этапов, т.е.смена плоидность впутан. Чтобы вернуться из диплоидной стадии в гаплоидную, мейоз должно произойти. Что касается изменений плоидность, существует 3 типа циклов:
- гаплонтический Жизненный цикл - гаплоидная стадия многоклеточная, диплоидная стадия - одиночная клетка, мейоз «зиготический».
- дипломатический жизненный цикл - диплоидная стадия многоклеточная и гаплоидная гаметы образуются, мейоз «гаметический».
- гаплодипонтический жизненный цикл (также называемый диплогаплонтический, диплобионтический, или же дибионтический жизненный цикл) - встречаются многоклеточные диплоидные и гаплоидные стадии, мейоз носит «споровой» характер.
Циклы отличаются тем, когда митоз (рост) происходит. Зиготический мейоз и гаметический мейоз имеют одну митотическую стадию: митоз происходит во время п фазы зиготического мейоза и во время 2-гоп фаза гаметного мейоза. Следовательно, зиготический и гаметический мейоз собирательно называют «гаплобионтическим» (единичная митотическая фаза, не путать с гаплонической). С другой стороны, у спорового мейоза есть митоз, состоящий из двух стадий, диплоидной и гаплоидной стадий, называемых «диплобионическим» (не путать с диплоидным).
Открытие
Изучение воспроизведение и разработка в организмах проводилась многими ботаниками и зоологами.
Вильгельм Хофмайстер продемонстрировал, что смена поколений - это свойство, объединяющее растения, и опубликовал этот результат в 1851 г. (см. сексуальность растений ).
Некоторые термины (гаплобионт и диплобионт), используемые для описания жизненных циклов, были первоначально предложены для водорослей Нильсом Сведелиусом, а затем стали использоваться для других организмов.[4][5] Другие термины (автогамия и гамонтогамия), используемые в протист Жизненные циклы были представлены Карлом Готлибом Греллем.[6] Описание сложных жизненных циклов различных организмов способствовало опровержению представлений о самозарождение в 1840-1850-х гг.[7]
Гаплонический жизненный цикл
Зиготический мейоз - это мейоз из зигота незамедлительно после кариогамия, который представляет собой слияние двух ядра клеток. Таким образом, организм завершает свою диплоидную фазу и производит несколько гаплоидных клеток. Эти клетки делятся митотически с образованием либо более крупных многоклеточных особей, либо более гаплоидных клеток. Два противоположных типа гамет (например, мужская и женская) этих особей или клеток сливаются в зиготу.
Во всем цикле зиготы - единственная диплоидная клетка; митоз происходит только в гаплоидной фазе.
Индивиды или клетки в результате митоза являются гаплонтами, поэтому этот жизненный цикл также называется гаплонтическим жизненным циклом. Гаплонты бывают:
- В архепластиданы: немного зеленые водоросли (например., Хламидомонада, Зигнема, Чара )[8]
- В страменопилы: немного золотые водоросли[8]
- В альвеолиты: много динофлагелляты, например, Ceratium, Gymnodinium, некоторые апикомплекс (например., Плазмодий )[9]
- В ризарии: немного эвглифиды,[10] аскетоспорейцы
- В раскапывает: немного парабазалиды[11]
- В амебозойные: Диктиостелиум[8]
- В опистоконцы: наиболее грибы (немного хитриды, зигомицеты, немного аскомицеты, базидиомицеты )[8][12]:15
Дипломированный жизненный цикл

В гаметном мейозе вместо немедленного деления мейотически для производства гаплоидных клеток зигота делится митотически для получения многоклеточного диплоидного индивидуума или группы более одноклеточных диплоидных клеток. Затем клетки диплоидных особей подвергаются мейозу с образованием гаплоидных клеток или гаметы. Гаплоидные клетки могут снова делиться (митозом) с образованием большего количества гаплоидных клеток, как у многих дрожжей, но гаплоидная фаза не является преобладающей фазой жизненного цикла. У большинства дипломатов митоз происходит только в диплоидной фазе, т.е. гаметы обычно образуются быстро и сливаются с образованием диплоидных зигот.
Во всем цикле гаметы обычно являются единственными гаплоидными клетками, а митоз обычно происходит только в диплоидной фазе.
Диплоидная многоклеточная особь - это дипломат, поэтому гаметический мейоз также называют дипломатическим жизненным циклом. Дипломы бывают:
- В архепластиданы: немного зеленые водоросли (например., Cladophora glomerata,[13] Ацетабулярия[8])
- В страменопилы: немного бурые водоросли (в Fucales однако их жизненный цикл также можно интерпретировать как сильно гетероморфно-диплогаплонтический, с сильно редуцированной фазой гаметофитов, как в цветущие растения ),[12]:207 немного ксантофиты (например., Vaucheria ),[12]:124 наиболее диатомеи,[11] немного оомицеты (например., Сапролегния, Plasmopara viticola ),[8] опалины,[11] немного "гелиозои "(например, Актинофрис, Актинофарий )[11][14]
- В альвеолиты: инфузории [11]
- В раскапывает: немного парабазалиды [11]
- В опистоконцы: животные, немного грибы (например, некоторые аскомицеты )[8]
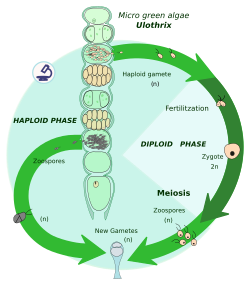
Гаплодипонтический жизненный цикл
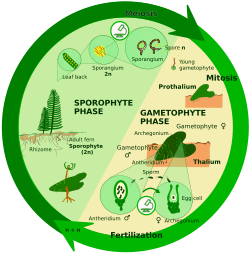
При споровом мейозе (также известном как промежуточный мейоз) зигота митотически делится с образованием многоклеточного диплоида. спорофит. Спорофит образует споры посредством мейоза, которые также затем разделите митотически продуцирующих гаплоидных особей, называемых гаметофиты. Гаметофиты производят гаметы путем митоза. У некоторых растений гаметофит не только мелкий, но и недолговечный; у других растений и многих водорослей гаметофит является «доминирующей» стадией жизненного цикла.
Гаплодиплонты бывают:
- В археопластиданы: красные водоросли (которые имеют два поколения спорофитов), некоторые зеленые водоросли (например., Ульва ), наземные растения[8]
- В страменопилы: наиболее бурые водоросли[8]
- В ризарии: много фораминиферы,[11] плазмодиофоромицеты[8]
- В амебозоа: миксогастриды
- В опистоконцы: немного грибы (немного хитриды, немного аскомицеты словно пивные дрожжи )[8]
- Другие эукариоты: гаптофиты[11]
У некоторых животных есть система определения пола называется гаплодиплоид, но это не связано с гаплодипонтическим жизненным циклом.
Вегетативный мейоз
Немного красные водоросли (Такие как Bonnemaisonia[15] и Lemanea ) и зеленые водоросли (например, Празиола ) имеют вегетативный мейоз, также называемый соматическим мейозом, что является редким явлением.[12]:82 Вегетативный мейоз может возникать как в гаплодипонтическом, так и в дипломатическом жизненных циклах. Гаметофиты остаются прикрепленными к спорофиту и являются его частью. Вегетативные (не репродуктивные) диплоидные клетки подвергаются мейозу, образуя вегетативные гаплоидные клетки. Они проходят множество митозов и продуцируют гаметы.
Другой феномен, называемый вегетативной диплоидизацией, тип апомиксис, встречается в некоторых бурые водоросли (например., Элахиста звездная).[16] Клетки в гаплоидной части растения спонтанно дублируют свои хромосомы с образованием диплоидной ткани.
Паразитический жизненный цикл
Паразиты зависят от эксплуатации одного или нескольких хозяева. Те, которые должны заразить более одного хозяина разновидность Считается, что для завершения своих жизненных циклов они имеют сложные или непрямые жизненные циклы, тогда как те, которые заражают один вид, имеют прямые жизненные циклы.
Если паразиту необходимо заразить данного хозяина, чтобы завершить свой жизненный цикл, то он считается облигатный паразит этого хозяина; иногда инфекция бывает факультативный - паразит может выжить и завершить свой жизненный цикл, не заразив этот конкретный вид хозяина. Иногда паразиты заражают хозяев, у которых они не могут завершить свой жизненный цикл; это случайные хозяева.
Хозяин, в котором паразиты размножаются половым путем, известен как окончательный, конечный или основной хозяин. У промежуточных хозяев паразиты либо не размножаются, либо размножаются бесполым путем, но паразит всегда развивается до новой стадии у этого типа хозяина. В некоторых случаях паразит заражает хозяина, но не развивается, эти хозяева известны как паратенический[17] или транспортных хозяев. Паратенический хозяин может быть полезен для повышения шансов передачи паразита окончательному хозяину. Например, кошачий легочный червь (Aelurostrongylus abstrusus) в качестве промежуточного хозяина использует слизня или улитку; личинка первой стадии попадает в моллюска и развивается до личинки третьей стадии, которая заразна для окончательного хозяина - кошки. Если мышь съест слизня, личинка третьей стадии войдет в ткани мыши, но не будет развиваться.

Эволюция
В примитивном типе жизненного цикла, вероятно, были гаплоидные особи с бесполым размножением.[11] Бактерии и археи демонстрируют такой жизненный цикл, и некоторые эукариоты, по-видимому, тоже (например, Криптофита, Чоанофлагеллата, много Эвгленозоа, много Амебозоа, немного красных водорослей, немного зеленые водоросли, то несовершенные грибы, немного коловратки и многие другие группы, не обязательно гаплоидные).[18] Однако эти эукариоты, вероятно, не являются примитивно бесполыми, но утратили половое размножение, или этого просто еще не наблюдали.[19][20] Многие эукариоты (включая животных и растения) проявляют бесполое размножение, которые могут быть факультативными или обязательными в жизненном цикле, с более или менее частым половым размножением.[21]
Смотрите также
- Смена поколений - Репродуктивный цикл растений и водорослей
- Апомиксис - Замена нормального полового размножения бесполым, без оплодотворения
- Гаплодиплоидия - Биологическая система, в которой пол определяется количеством наборов хромосом
- Парасексуальный цикл - Несексуальный механизм передачи генетического материала без мейоза
- Партеногенез - Естественная форма бесполого размножения, при которой рост и развитие эмбрионов происходит без оплодотворения.
- Репродуктивная биология - Отделение биологии репродукции изучает
- Митотическая рекомбинация
Рекомендации
- ^ Грэм Белл и Василики Куфопану (1991). «Архитектура жизненного цикла малых организмов». Философские труды: биологические науки. 332 (1262): 81–89. Дои:10.1098 / рстб.1991.0035. JSTOR 55494.
- ^ Родригес, Джулиани Кола Фернандес; Годиньо, Хосеана Лима Прадо; де Соуза, Вандерли (2014). "Биология патогенных трипаносоматид человека: эпидемиология, жизненный цикл и ультраструктура". Белки и протеомика лейшмании и трипаносомы. Субклеточная биохимия. Springer Нидерланды. 74: 1–42. Дои:10.1007/978-94-007-7305-9_1. ISBN 978-94-007-7304-2. PMID 24264239.
- ^ Диксон, П.С. 1973 г. Биология родофиты. Оливер и Бойд. ISBN 0 05 002485 Х
- ^ К. Скоттсберг (1961), «Нильс Эберхард Сведелиус. 1873-1960», Биографические воспоминания членов Королевского общества, 7: 294–312, Дои:10.1098 / рсбм.1961.0023
- ^ Сведелиус, Н. 1931. Ядерные фазы и чередование у Rhodophyceae. В: Beihefte zum Botanischen Centralblatt. Группа 48/1: 38-59.
- ^ Л. Маргулис (1996), «Архейно-эубактериальные слияния в происхождении эукарии: филогенетическая классификация жизни», PNAS, 93 (3): 1071–1076, Дои:10.1073 / pnas.93.3.1071, ЧВК 40032, PMID 8577716
- ^ Мозелио Шехтер (2009). Энциклопедия микробиологии. Академическая пресса. Том 4, с. 85.
- ^ а б c d е ж грамм час я j k Диас Гонсалес, Т.Э., К. Фернандес-Карвахаль Альварес и Х.А. Фернандес Прието. (2004). Curso de Botánica. Хихон: Треа. Интернет-материалы: Ботаника: Ciclos biológicos de Vegetales. (Жизненные циклы растений, на испанском языке). Departamento de Biología de Organismos y Sistemas, Universidad de Oviedo.
- ^ Sinden, R.E .; Хартли, Р. Х. "Идентификация мейотического отдела малярийных паразитов". Журнал протозоологии.
- ^ Лар DJ, Парфри Л.В., Митчелл Е.А., Кац Л.А., Лара Е. (июль 2011 г.). «Целомудрие амеб: переоценка доказательств секса у амебоидных организмов». Proc. Биол. Наука. 278 (1715): 2083–6. Дои:10.1098 / rspb.2011.0289. ЧВК 3107637. PMID 21429931.
- ^ а б c d е ж грамм час я Рупперт, Э. Э., Фокс, Р. С., и Барнс, Р. Д. (2004). Зоология беспозвоночных: функциональный эволюционный подход. Бельмонт, Калифорния: Томас-Брукс / Коул, стр. 26.
- ^ а б c d ван ден Хук, К., Д.Г. Манн, Х. Янс (1995). Водоросли: введение в фикологию. Издательство Кембриджского университета (623 стр.).
- ^ О. П. Шарма. Учебник водорослей, стр. 189
- ^ Белл, Г. (1989). Секс и смерть у простейших. Издательство Кембриджского университета, стр. 11, [1].
- ^ Сальвадор Солер, Ноэми; Гомес Гаррета, Амелия; Антония Рибера Сигуан, М. (1 августа 2009 г.). «Соматический мейоз в истории жизни Bonnemaisonia asparagoides и Bonnemaisonia clavata (Bonnemaisoniales, Rhodophyta) с Пиренейского полуострова». Европейский журнал психологии. 44 (3): 381–393. Дои:10.1080/09670260902780782.
- ^ Льюис, Р.Дж. (1996). «Хромосомы бурых водорослей». Phycologia. 35 (1): 19–40. Дои:10.2216 / i0031-8884-35-1-19.1.
- ^ Шмидт и Робертс. 1985. Основы паразитологии 3-е изд. Times Mirror / Mosby College Publishing
- ^ Heywood, P .; Маги, П. (1976). «Мейоз у протистов. Некоторые структурные и физиологические аспекты мейоза у водорослей, грибов и простейших». Бактериологические обзоры. 40 (1): 190–240. Дои:10.1128 / ммбр.40.1.190-240.1976. ЧВК 413949. PMID 773364.
- ^ Шехре-Бану Малик; Артур У. Пайтлинг; Лорен М. Стефаниак; Эндрю М. Шурко и Джон М. Логсдон-младший (2008). "Расширенный перечень консервативных мейотических генов предоставляет доказательства секса в Влагалищная трихомонада". PLoS ONE. 3 (8): e2879. Bibcode:2008PLoSO ... 3,2879 млн. Дои:10.1371 / journal.pone.0002879. ЧВК 2488364. PMID 18663385.
- ^ Speijer, D .; Lukeš, J .; Элиаш, М. (2015). «Секс - это вездесущий, древний и неотъемлемый атрибут эукариотической жизни». PNAS. 112 (29): 8827–8834. Дои:10.1073 / pnas.1501725112. ЧВК 4517231. PMID 26195746.
- ^ Schön, I .; Martens, K .; ван Дейк, П. (2009). Утраченный секс: эволюционная биология партеногенеза. Springer. ISBN 9789048127702.
дальнейшее чтение
- Жизненные циклы: размышления эволюционного биолога. Джон Тайлер Боннер. Princeton University Press, Принстон (Нью-Джерси).
- Валеро, М. (1992). «Эволюция чередования гаплоидной и диплоидной фаз в жизненных циклах». Тенденции в экологии и эволюции. 7 (1): 25–29. Дои:10.1016 / 0169-5347 (92) 90195-ч. PMID 21235940.
- Mable, B .; Отто, С. (1998). «Эволюция жизненных циклов с гаплоидной и диплоидной фазой» (PDF). BioEssays. 20 (6): 453–462. Дои:10.1002 / (sici) 1521-1878 (199806) 20: 6 <453 :: aid-bies3> 3.0.co; 2-n.