Амфибия - Amphibian
| Амфибии | |
|---|---|
 | |
| По часовой стрелке сверху справа: Сеймурия, Мексиканский роющий цецилий, восточный тритон и лист зеленая квакша | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Batrachomorpha |
| Учебный класс: | Амфибия серый 1825[1] |
| Подклассы | |
| |
Амфибии находятся экзотермический, четвероногий позвоночные из учебный класс Амфибия. Все живые амфибии принадлежат к группе Лиссамфибия. Они населяют самые разные среда обитания, причем большинство видов обитает в земной, роющий, древесный или пресноводный водные экосистемы. Таким образом, амфибии обычно начинаются как личинки живут в воде, но некоторые виды развили поведенческие адаптации, чтобы обойти это.
Молодые обычно проходят метаморфоза от личинки с жабрами до взрослой дыхательной формы с легкие. Земноводные используют кожу в качестве вторичной дыхательной поверхности, а некоторые мелкие наземные саламандры и лягушки не имеют легких и полностью полагаются на свою кожу. Они внешне похожи на ящерицы но, наряду с млекопитающими и птицами, рептилии амниот и не нуждаются в водоемах для размножения. С их сложными репродуктивными потребностями и проницаемой кожей амфибии часто экологические показатели; в последние десятилетия произошли драматические сокращение популяции амфибий для многих видов по всему миру.
Самые ранние амфибии развился в Девонский период от саркоптерийский рыба с легкими и плавниками с костлявыми конечностями, которые помогли адаптироваться к суше. Они диверсифицировались и стали доминировать в Каменноугольный и Пермский период периоды, но позже были вытеснены рептилиями и другими позвоночными. Со временем амфибии уменьшились в размерах и уменьшились в разнообразии, оставив только современный подкласс Lissamphibia.
Три современных отряда амфибий: Анура (лягушки и жабы), Уродела (саламандры) и Apoda (червеобразные). Число известных видов амфибий составляет около 8000, из которых около 90% - лягушки. Самая маленькая амфибия (и позвоночное животное) в мире - лягушка из Новая Гвинея (Paedophryne amauensis ) длиной всего 7,7 мм (0,30 дюйма). Самая большая из ныне живущих амфибий - 1,8 м (5 футов 11 дюймов). Южно-китайская гигантская саламандра (Андриас слигой), но это затмевается потухшими 9 м (30 футов) Прионозух от средняя пермь Бразилии. Изучение амфибий называется батрахология, а изучение как рептилий, так и земноводных называется герпетология.
Классификация

Слово «амфибия» происходит от Древнегреческий термин ἀμφίβιος (амфибиос), что означает "оба вида жизни", ἀμφί означает "обоих видов" и βιος значение «жизнь». Первоначально этот термин использовался как общее прилагательное для животных, которые могли жить на суше или в воде, включая тюленей и выдр.[2] Традиционно к классу Amphibia относятся все позвоночные четвероногие, не являющиеся амниотами. Амфибия в самом широком смысле (Sensu lato ) был разделен на три подклассы, два из которых вымерли:[3]
- Подкласс Лепоспондили † (небольшая палеозойская группа, более родственная амниотам, чем Lissamphibia)
- Подкласс Темноспондили † (разнообразный палеозойский и раннемезозойский класс)
- Подкласс Лиссамфибия (все современные земноводные, включая лягушек, жаб, саламандр, тритонов и цецилий)
- Salientia (лягушки, жабы и родственники): от юрского до настоящего времени - 6200 современных видов в 53 семействах
- Хвостатые (саламандры, тритоны и родственники): от юрского до настоящего времени - 652 современных вида в 9 семействах
- Гимнофиона (цецилии и родственники): от юрского до настоящего времени - 192 современных вида в 10 семействах
- Allocaudata † (Albanerpetontidae ) Средняя юра - ранний плейстоцен

Фактическое количество видов в каждой группе зависит от применяемой таксономической классификации. Две наиболее распространенные системы - это классификация, принятая на веб-сайте AmphibiaWeb, Калифорнийский университет в Беркли и классификация по герпетолог Даррел Фрост и Американский музей естественной истории, доступная в виде онлайн-справочной базы данных "Amphibian Species of the World".[4] Указанное выше количество видов следует за Frost, и общее количество известных видов земноводных по состоянию на 31 марта 2019 г. составляет ровно 8000,[5] из которых почти 90% - лягушки.[6]
С филогенетический классификация, таксон Лабиринтодонтия был отброшен, поскольку это полипарафилетический группа без уникальных определяющих характеристик, кроме общие примитивные характеристики. Классификация варьируется в зависимости от предпочтительной филогении автора и от того, используют ли они на основе стебля или на основе узла классификация. Традиционно амфибии как класс определяются как все четвероногие с личиночной стадией, а группа, которая включает общих предков всех ныне живущих земноводных (лягушек, саламандр и цецилий) и всех их потомков, называется Lissamphibia. Филогения палеозойских амфибий сомнительна, и Lissamphibia, возможно, подпадают под вымершие группы, такие как Temnospondyli (традиционно относящиеся к подклассу Labyrinthodontia) или Lepospondyli, а в некоторых анализах даже к амниотам. Это означает, что сторонники филогенетическая номенклатура удалили большое количество базальный Девонские и каменноугольные группы четвероногих амфибий, ранее помещенные в Амфибии в Линнеевская таксономия, и включил их в другое место в кладистическая таксономия.[1] Если общий предок амфибий и амниот входит в состав Amphibia, он становится парафилетической группой.[7]
Все современные амфибии входят в подкласс Lissamphibia, который обычно считается клады, группа видов, которые произошли от общего предка. Три современных отряда - это Anura (лягушки и жабы), Caudata (или Urodela, саламандры) и Gymnophiona (или Apoda, цецилии).[8] Было высказано предположение, что саламандры возникли отдельно от темноспондилоподобного предка, и даже что цецилийцы являются сестринской группой продвинутых рептилиоморф амфибии, а значит, и амниот.[9] Хотя известны окаменелости нескольких более древних прото-лягушек с примитивными характеристиками, самая старая «настоящая лягушка» - Просалирус битис, от Раннеюрский период Кайентская свита Аризоны. Анатомически он очень похож на современных лягушек.[10] Самый старый из известных цецилий - еще один раннеюрский вид, Eocaecilia micropodia, также из Аризоны.[11] Самая ранняя саламандра Beiyanerpeton jianpingensis от Поздняя юра Северо-Восточного Китая.[12]
Власти расходятся во мнениях относительно того, является ли Salientia надотрядом, включающим отряд Anura, или является ли Anura подотрядом отряда Salientia. Лиссамфибию традиционно делят на три заказы, но вымершее семейство саламандр, Albanerpetontidae, теперь считается частью Лиссамфибии вместе с надотрядом Salientia. Кроме того, Salientia включает все три недавних заказа плюс триасовую прото-лягушку, Триадобатрахус.[13]
Эволюционная история


Первые крупные группы амфибий сложились в Девонский период, около 370 миллионов лет назад, с рыба с лопастными плавниками которые были похожи на современные латимерия и двоякодышащая рыба.[14] У этих древних рыб с лопастными плавниками развились многосуставные плавники, похожие на ноги, с пальцами, которые позволяли им ползать по дну моря. У некоторых рыб развились примитивные легкие, которые помогают им дышать воздухом, когда в стоячих водоемах девонских болот было мало кислорода. Они также могли использовать свои сильные ласты, чтобы подниматься из воды на сушу, если того требовали обстоятельства. В конце концов, их костлявые плавники эволюционировать в конечности, и они станут предками для всех четвероногие, включая современных амфибий, рептилий, птиц и млекопитающие. Несмотря на то, что они могли ползать по суше, многие из этих доисторических тетраподоморф рыбы по-прежнему проводили большую часть времени в воде. У них начали развиваться легкие, но они все еще дышали преимущественно жабрами.[15]
Много примеров видов, показывающих переходные особенности были обнаружены. Ихтиостега был одним из первых примитивных земноводных с ноздрями и более эффективными легкими. У него были четыре крепкие конечности, шея, хвост с плавниками и череп, очень похожий на череп с лопастными плавниками. Евстеноптерон.[14] У земноводных появились приспособления, которые позволяли им дольше оставаться вне воды. Их легкие улучшились, а их скелеты стали тяжелее и сильнее, они лучше могли выдерживать вес своих тел на суше. У них появились «руки» и «ступни» с пятью и более пальцами;[16] кожа стала более способной удерживать жидкости организма и сопротивляться высыханию.[15] Рыба гиомандибула кость в подъязычная область за жабрами уменьшилась в размерах и стала стремени уха земноводных - приспособление, необходимое для слуха на суше.[17] Родство между амфибиями и костистость рыба - это многослойная структура зубов и парная надзатылочные кости на затылке, ни одна из этих черт не встречается где-либо еще в животном мире.[18]

В конце девонского периода (360 миллионов лет назад) моря, реки и озера кишели жизнью, в то время как земля была царством ранних растений и лишена позвоночных животных.[18] хотя некоторые, такие как Ихтиостега, возможно, иногда выбирались из воды. Считается, что они, возможно, двигались передними конечностями, волоча задние конечности аналогично тому, как это использовали морской слон.[16] Рано Каменноугольный (360–345 миллионов лет назад) климат стал влажным и теплым. Обширные болота, развитые с мхи, папоротники, хвощи и каламиты. Воздуходышащий членистоногие развились и вторглись в землю, где они давали пищу для плотоядный амфибии, которые начали приспосабливаться к земной среде. Других четвероногих на суше не было, а земноводные находились на вершине пищевой цепи, занимая экологическое положение, которое в настоящее время занимает крокодил. Несмотря на то, что они обладали конечностями и могли дышать воздухом, у большинства по-прежнему было длинное сужающееся тело и сильный хвост.[18] Это были главные наземные хищники, иногда достигавшие нескольких метров в длину, охотясь на крупных насекомых того времени и многих видов рыб в воде. Им все еще нужно было вернуться в воду, чтобы отложить яйца без скорлупы, и даже у большинства современных амфибий есть полностью водная личиночная стадия с жабрами, как у их предков рыб. Это было развитие амниотический яйцо, которое предотвращает высыхание развивающегося эмбриона, что позволило рептилиям размножаться на суше и привело к их господство в последующий период.[14]
После Обрушение тропических лесов каменноугольного периода господство амфибий уступило место рептилиям,[19] и земноводные подверглись дальнейшему опустошению Пермско-триасовое вымирание.[20] Вовремя Триасовый период (250–200 миллионов лет назад) рептилии продолжали конкурировать с амфибиями, что привело к сокращению как размеров земноводных, так и их значения в биосфера. Согласно летописи окаменелостей, Лиссамфибия, который включает в себя всех современных земноводных и является единственной сохранившейся линией, возможно, произошел от вымерших групп Темноспондили и Лепоспондили в какой-то период между поздним карбоном и ранним триасом. Относительная нехватка окаменелостей исключает точное датирование,[15] но самое последнее молекулярное исследование, основанное на мультилокусная последовательность, предполагает поздний карбон /Ранняя пермь происхождение современных амфибий.[21]

Происхождение и эволюционные отношения между тремя основными группами амфибий являются предметом споров. Молекулярная филогения 2005 г., основанная на рДНК Анализ, предполагает, что саламандры и цецилии более тесно связаны друг с другом, чем с лягушками. Также кажется, что расхождение трех групп имело место в Палеозой или рано Мезозойский (около 250 миллионов лет назад), до распада суперконтинента Пангея и вскоре после их отклонения от рыб с лопастными плавниками. Краткость этого периода и скорость, с которой происходила радиация, помогли бы объяснить относительную редкость примитивных окаменелостей амфибий.[22] Есть большие пробелы в Окаменелости, но открытие Gerobatrachus hottoni из ранней перми в Техасе в 2008 г. предоставил недостающее звено со многими характеристиками современных лягушек.[9] Молекулярный анализ предполагает, что дивергенция лягушка-саламандра произошла значительно раньше, чем палеонтологический свидетельства указывают.[9] Более новые исследования показывают, что общий предок всех лизамфибий жил около 315 миллионов лет назад, и что стереоспондилы являются ближайшими родственниками цецилий.[23]
Поскольку они произошли от выпадающих рыб, амфибиям пришлось внести определенные изменения для жизни на суше, включая необходимость разработки новых средств передвижения. В воде боковые удары хвостов толкали их вперед, но на суше требовались совсем другие механизмы. Их позвоночные столбы, конечности, пояса конечностей и мускулатура должны были быть достаточно сильными, чтобы поднимать их над землей для передвижения и кормления. Взрослые наземные животные выбросили свои боковая линия системы и адаптировали свои сенсорные системы к получению стимулов через воздух. Им нужно было разработать новые методы регулирования температуры своего тела, чтобы справляться с колебаниями температуры окружающей среды. Они выработали поведение, подходящее для воспроизводства в земной среде. Их кожа подвергалась вредному воздействию ультрафиолетовый лучи, ранее поглощенные водой. Кожа изменилась, чтобы стать более защитной и предотвратить чрезмерную потерю воды.[24]
Характеристики
Суперкласс Тетрапода делится на четыре класса позвоночных животных с четырьмя конечностями.[25] Рептилии, птицы и млекопитающие - это амниоты, яйца которых либо откладываются, либо переносятся самками и окружены несколькими оболочками, некоторые из которых непроницаемы.[26] Без этих мембран амфибиям требуются водоемы для размножения, хотя некоторые виды разработали различные стратегии для защиты или обхода уязвимой стадии водных личинок.[24] В море они не встречаются, за исключением одной или двух лягушек, которые живут в солоноватая вода в мангровые заросли болота;[27] в Саламандра андерсона Между тем встречается в озерах с солоноватой или соленой водой.[28] На суше амфибии ограничены влажными средами обитания из-за необходимости сохранять влажную кожу.[24]
Современные амфибии имеют упрощенную анатомию по сравнению с их предками из-за педоморфоз, вызванный двумя эволюционными тенденциями: миниатюризацией и необычно большим геномом, что приводит к более медленным темпам роста и развития по сравнению с другими позвоночными.[29][30] Другая причина их размера связана с их быстрой метаморфозой, которая, по-видимому, произошла только у предков лиссамфибий; во всех остальных известных направлениях развитие шло гораздо более плавно. Поскольку реконструкция устройства питания означает, что они не едят во время метаморфоза, метаморфоза должна идти быстрее, чем меньше особь, поэтому это происходит на ранней стадии, когда личинки еще маленькие. (Самые крупные виды саламандр не претерпевают метаморфоз.)[31] Амфибии, откладывающие яйца на суше, часто проходят всю метаморфозу внутри яйца. Анамниотическое наземное яйцо меньше 1 см в диаметре из-за проблем с диффузией, размер, который ограничивает количество послевлупового роста.[32]
Самая маленькая амфибия (и позвоночное животное) в мире - это микрогилид лягушка из Новая Гвинея (Paedophryne amauensis ) впервые обнаружен в 2012 году. Он имеет среднюю длину 7,7 мм (0,30 дюйма) и является частью рода, который включает четыре из десяти самых маленьких видов лягушек в мире.[33] Самая большая из ныне живущих амфибий - 1,8 м (5 футов 11 дюймов). Китайская гигантская саламандра (Андриас давидиан)[34] но это намного меньше, чем самая большая из когда-либо существовавших земноводных - вымершая 9 м (30 футов) Прионозух, похожий на крокодила темноспондил, датируемый 270 миллионами лет назад из средней перми Бразилии.[35] Самая большая лягушка - африканская Лягушка-голиаф (Конрауа голиаф), который может достигать 32 см (13 дюймов) и весить 3 кг (6,6 фунта).[34]
Амфибии - это экзотермические (хладнокровные) позвоночные, которые не поддерживают температуру своего тела за счет внутренних физиологический процессы. Их скорость метаболизма низка, и в результате их потребности в пище и энергии ограничены. Во взрослом состоянии у них есть слезные протоки и подвижные веки, а у большинства видов есть уши, которые могут улавливать колебания воздуха или земли. У них мускулистые языки, которые у многих видов могут быть выступающими. Современные амфибии полностью окостеневший позвонки с суставные отростки. Их ребра обычно короткие и могут срастаться с позвонками. Их черепа в основном широкие и короткие и часто не полностью окостенели. В их коже мало кератин и без чешуи, за исключением нескольких рыбьих чешуек у некоторых цецилий. Кожа содержит много слизистые железы а у некоторых видов ядовитые железы (разновидность зернистой железы). В сердцах амфибий три камеры, две предсердие и один желудочек. У них есть мочевой пузырь и азотные отходы выделяются в основном в виде мочевина. Большинство земноводных откладывают яйца в воде и имеют водных личинок, которые претерпевают метаморфоз, чтобы стать взрослыми наземными особями. Земноводные дышат с помощью насоса, при котором воздух сначала всасывается в щечно-глоточный область через ноздри. Затем они закрываются, и воздух нагнетается в легкие за счет сжатия горла.[36] Они дополняют это газообмен через кожу.[24]
Анура

Приказ Анура (от древнегреческого а (п) - значение "без" и ура означает «хвост») включает лягушек и жаб. У них обычно длинные задние конечности, которые складываются под ними, более короткие передние конечности, перепончатые пальцы ног без когтей, без хвостов, большие глаза и железистая влажная кожа.[8] Члены этого отряда с гладкой кожей обычно называются лягушками, а те, у кого бородавчатый шкуры известны как жабы. Таксономически это различие не является формальным, и из этого правила есть многочисленные исключения. Члены семьи Bufonidae известны как «настоящие жабы».[37] Размер лягушек колеблется от 30 сантиметров (12 дюймов). Лягушка-голиаф (Конрауа голиаф) Западной Африки[38] до 7,7 миллиметра (0,30 дюйма) Paedophryne amauensis, впервые описанный в Папуа-Новой Гвинее в 2012 году, он также является самым маленьким из известных позвоночных.[39] Хотя большинство видов связаны с водой и влажной средой обитания, некоторые из них приспособлены жить на деревьях или в пустынях. Они встречаются по всему миру, за исключением полярных областей.[40]
Анура делится на три подотряда, которые широко признаны научным сообществом, но отношения между некоторыми семьями остаются неясными. Будущее молекулярный исследования должны предоставить дальнейшее понимание их эволюционных взаимоотношений.[41] Подотряд Археобатрахия содержит четыре семейства примитивных лягушек. Это Ascaphidae, Bombinatoridae, Дискоглоссиды и Leiopelmatidae которые имеют несколько производных черт и, вероятно, являются парафилетическими по отношению к другим линиям лягушек.[42] Шесть семейств в более эволюционно развитом подотряде Мезобатрахия являются роющий Megophryidae, Pelobatidae, Pelodytidae, Scaphiopodidae и Носороги и обязательно водный Pipidae. У них есть определенные характеристики, которые занимают промежуточное положение между двумя другими подотрядами.[42] Необатрахия на сегодняшний день является самым крупным подотрядом и включает оставшиеся семейства современных лягушек, включая наиболее распространенные виды. Девяносто шесть процентов из более чем 5000 существующих видов лягушек - необатрахиане.[43]
Хвостатые

(Andrias japonicus), примитивная саламандра
Приказ Хвостатые (от латинского кауда означает «хвост») состоит из саламандр - удлиненных животных с низкой посадкой, которые по форме больше всего напоминают ящериц. Это симплезиоморфный признак и они не более тесно связаны с ящерицами, чем с млекопитающими.[44] У саламандр нет когтей, у них кожа без чешуи, гладкая или покрытая бугорки и хвосты, которые обычно приплюснуты из стороны в сторону и часто с ребрами. Их размер варьируется от Китайская гигантская саламандра (Андриас давидиан), который, как сообщается, вырастает до 1,8 метра (5 футов 11 дюймов),[45] к миниатюрному Торий пеннатул из Мексики, длина которых редко превышает 20 мм (0,8 дюйма).[46] Саламандры в основном Лауразийский распространение, присутствуя в большей части Голарктический регион северного полушария. Семья Plethodontidae также встречается в Центральной Америке и Южной Америке к северу от Бассейн Амазонки;[40] Южная Америка была захвачена из Центральной Америки примерно в начале Миоцен, 23 миллиона лет назад.[47] Уродела - это имя, иногда используемое для всех сохранившийся виды саламандр.[48] Члены нескольких семейств саламандр стали педоморфный и либо не могут завершить метаморфоз, либо сохраняют некоторые личиночные характеристики во взрослом состоянии.[49] Большинство саламандр имеют длину менее 15 см (6 дюймов). Они могут быть наземными или водными, и многие проводят часть года в каждой среде обитания. На суше они в основном проводят день, прячась под камнями или бревнами или в густой растительности, а вечером и ночью выходят на корм для червей, насекомых и других беспозвоночных.[40]

(Triturus dobrogicus), продвинутая саламандра
Подотряд Криптобранхоидея содержит примитивных саламандр. Было обнаружено несколько ископаемых криптобожаберных видов, но живых осталось только три вида: китайская гигантская саламандра (Андриас давидиан), Японская гигантская саламандра (Andrias japonicus) и адский маг (Cryptobranchus alleganiensis) из Северной Америки. Эти крупные земноводные сохраняют несколько личинок во взрослом состоянии; имеются жаберные щели и не закрытые глаза. Уникальной особенностью является их способность сосать, прижимая либо левую сторону нижней челюсти, либо правую.[50] Самцы роют гнезда, убеждают самок откладывать в них ниточки яиц и охраняют их. Они дышат не только легкими, но и через многочисленные складки своей тонкой кожи, которая капилляры близко к поверхности.[51]
Подотряд Саламандройда содержит продвинутых саламандр. Они отличаются от криптожаберных видов тем, что слились предсуставные кости в нижней челюсти и с помощью внутреннего оплодотворения. У саламандрид самец откладывает пучок спермы, сперматофор, а самка берет его и вставляет в свою клоаку, где сперма хранится до тех пор, пока не откладываются яйца.[52] Самым большим семейством в этой группе является Plethodontidae, саламандры без легких, которые включают 60% всех видов саламандр. В семья Salamandridae включает настоящих саламандр и имя "тритон "дается членам его подсемейства Pleurodelinae.[8]
Третий подотряд, Сиреноидея, содержит четыре вида сирен, которые принадлежат к одному семейству, Сирениды. Членами этого ордена являются угорь -подобные водные саламандры с сильно сокращенными передними конечностями и отсутствием задних конечностей. Некоторые из их функций являются примитивными, а другие производными.[53] Оплодотворение, вероятно, будет внешним, поскольку у сиренид отсутствуют клоакальные железы, используемые самцами саламандридов для производства сперматофоров, а у самок - клоакальные железы. сперматека для хранения спермы. Несмотря на это, яйца откладываются поодиночке, что не способствует внешнему оплодотворению.[52]
Гимнофиона

Приказ Гимнофиона (от греч. тренажерные залы что означает "голый" и офис означает «змей») или Apoda включает в себя червеобразных. Это длинные, цилиндрические, безногие животные со змеиным или червеобразный форма. Взрослые особи различаются по длине от 8 до 75 сантиметров (от 3 до 30 дюймов), за исключением Цецилия Томсона (Цецилия томпсони), который может достигать 150 сантиметров (4,9 фута). Кожа слепой кишки имеет большое количество поперечных складок и у некоторых видов содержит крошечные встроенные кожные чешуйки. У него рудиментарные глаза, покрытые кожей, которая, вероятно, ограничена различием в интенсивности света. Также есть пара коротких щупальца рядом с глазом, который можно расширить и который тактильный и обонятельный функции. Большинство цецилий живут под землей в норах во влажной почве, в гнилой древесине и под растительными остатками, но некоторые из них водные.[54] Большинство видов откладывают яйца под землей, и когда личинки вылупляются, они направляются к соседним водоемам. Другие вынашивают яйца, и личинки претерпевают метаморфоз, прежде чем вылупятся яйца. Некоторые виды рождают живых детенышей, питая их секретами желез, пока они находятся в яйцеводе.[55] Цецилийцы в основном Гондванец распространение, найденное в тропических регионах Африки, Азии и Центральной и Южной Америки.[56]
Анатомия и психология
Кожа

В покровный структура содержит некоторые типичные характеристики, общие для наземных позвоночных, такие как наличие высоко ороговевший внешние слои, периодически обновляемые в процессе линьки, контролируемые гипофиз и щитовидная железа железы. Часто встречаются локальные утолщения (часто называемые бородавками), например, на жабах. Внешняя поверхность кожи периодически сбрасывается, в основном целиком, в отличие от млекопитающих и птиц, где она сбрасывается в виде хлопьев. Земноводные часто поедают отшелушенную кожу.[40] Цецилии уникальны среди земноводных тем, что имеют минерализованные кожные чешуйки, встроенные в дерма между бороздками на коже. Их сходство с чешуей костистых рыб во многом внешнее. Ящерицы и у некоторых лягушек есть нечто похожее остеодермы формирование костных отложений в дерме, но это пример конвергентная эволюция с подобными структурами, возникшими независимо в различных линиях позвоночных.[57]
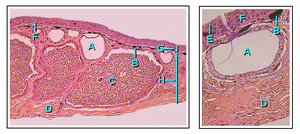
Кожа земноводных водопроницаема. Газообмен может происходить через кожу (кожное дыхание ), и это позволяет взрослым амфибиям дышать, не поднимаясь на поверхность воды, и впадать в спячку на дне прудов.[40] Чтобы компенсировать свою тонкую и нежную кожу, у земноводных развились слизистые железы, в основном на голове, спине и хвосте. Вырабатываемые ими выделения помогают сохранять кожу влажной. Кроме того, у большинства видов амфибий есть зернистые железы, выделяющие неприятные или ядовитые вещества. Некоторые токсины амфибий могут быть смертельными для человека, в то время как другие оказывают незначительное воздействие.[58] Главные ядовитые железы - паротоиды, производят нейротоксин буфотоксин и располагаются за ушами жаб, вдоль спины лягушек, за глазами саламандр и на верхней поверхности слепых червей.[59]
Цвет кожи земноводных определяется тремя слоями пигментных клеток, которые называются хроматофоры. Эти три клеточных слоя состоят из меланофоров (занимающих самый глубокий слой), гуанофоров (образующих промежуточный слой и содержащих множество гранул, производящих сине-зеленый цвет) и липофоров (желтый, самый поверхностный слой). Изменение цвета у многих видов инициируется гормоны секретируется гипофизом. В отличие от костистых рыб, нервная система не контролирует пигментные клетки напрямую, и это приводит к тому, что изменение цвета происходит медленнее, чем у рыб. Ярко окрашенная кожа обычно указывает на то, что этот вид токсичен, и является предупреждающим знаком для хищников.[60]
Скелетная система и движение

У амфибий скелетная система структурно гомологичный другим четвероногим, хотя и с рядом вариаций. У всех есть четыре конечности, за исключением безногих слепых и нескольких видов саламандр с уменьшенными конечностями или без них. Кости полые и легкие. Скелетно-мышечная система сильна, что позволяет ей поддерживать голову и тело. Кости полностью окостеневший и позвонки сцепляются друг с другом посредством перекрывающихся отростков. В грудной пояс поддерживается мышцами, а хорошо развитые тазовый пояс прикрепляется к позвоночнику парой крестцовых ребер. В подвздошная кость наклоняется вперед, и тело держится ближе к земле, чем у млекопитающих.[61]
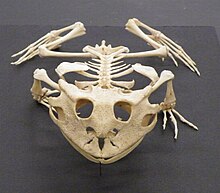
(Ceratophrys cornuta)
У большинства земноводных четыре пальца на передней лапе и пять на задней, но ни на одной из них нет когтей. У некоторых саламандр меньше пальцев и амфиумы похожи на угря с крошечными короткими ножками. В сирены водные саламандры с короткими передними конечностями и без задних конечностей. Цецилии без конечностей. Они зарываются, как дождевые черви, с движущимися по телу зонами мышечных сокращений. По поверхности земли или в воде они передвигаются, волнообразно покачиваясь из стороны в сторону.[62]
У лягушек задние лапы больше передних, особенно у тех видов, которые передвигаются в основном путем прыжков или плавания. У ходоков и бегунов задние конечности не такие большие, а норы в основном имеют короткие конечности и широкое тело. У ступней есть приспособления к образу жизни, с перепонками между пальцами для плавания, широкими липкими подушечками пальцев для лазания и ороговевшими бугорками на задних лапах для копания (лягушки обычно копают землю задом наперед). У большинства саламандр конечности короткие, примерно одинаковой длины и выступают под прямым углом к телу. Передвижение на суше - это ходьба, и хвост часто раскачивается из стороны в сторону или используется в качестве опоры, особенно при лазании. В их обычной походке только одна нога продвигается вперед, как это было у их предков, рыб с лопастными плавниками.[61] Некоторые саламандры в роду Анеиды и некоторые полнокровие лазать по деревьям и иметь длинные конечности, большие пальцы ног и цепкий хвост.[52] У водных саламандр и головастиков-лягушек хвост имеет спинной и вентральный плавников и перемещается из стороны в сторону как средство движения. У взрослых лягушек нет хвостов, а у цецилий только очень короткие.[62]

Саламандры используют свои хвосты для защиты, и некоторые готовы выбросить их за борт, чтобы спасти свои жизни, в процессе, известном как аутотомия. Некоторые виды Plethodontidae имеют слабую зону у основания хвоста и легко используют эту стратегию. Хвост часто продолжает подергиваться после отделения, что может отвлечь нападающего и позволить саламандре сбежать. И хвосты, и конечности можно регенерировать.[63] Взрослые лягушки не могут отрастить конечности, но головастики могут.[62]
Сердечно-сосудистая система


1 - Внутренние жабры, где происходит реоксигенация крови
2 - Точка, в которой кровь истощает кислород и возвращается к сердцу по венам.
3 - Двухкамерное сердце.
Красный цвет означает кровь, насыщенную кислородом, а синий - кровь с низким содержанием кислорода.
У амфибий есть ювенильная стадия и взрослая стадия, и кровеносные системы этих двух различны. На стадии молоди (или головастика) кровообращение похоже на кровообращение рыбы; двухкамерное сердце качает кровь через жабры, где она насыщается кислородом, и распространяется по всему телу и обратно к сердцу в виде единой петли. Во взрослом возрасте земноводные (особенно лягушки) теряют жабры и развивают легкие. У них есть сердце, состоящее из одного желудочка и двух предсердий. Когда желудочек начинает сокращаться, дезоксигенированная кровь перекачивается через легочная артерия в легкие. Продолжая сокращение, насыщенная кислородом кровь перекачивается по всему телу. Смешивание двух кровотоков сводится к минимуму анатомией камер.[64]
Нервная и сенсорная системы
В нервная система в основном такая же, как и у других позвоночных, с центральным мозгом, спинным мозгом и нервами по всему телу. Мозг земноводных менее развит, чем у рептилий, птиц и млекопитающих, но по морфологии и функциям похож на мозг рыб. Считается, что земноводные способны воспринимать боль. Мозг состоит из равных частей, головной мозг, средний мозг и мозжечок. Различные части головного мозга обрабатывают сенсорную информацию, такую как обоняние в обонятельной доле и зрение в зрительной доле, и, кроме того, это центр поведения и обучения. Мозжечок - центр мышечной координации и продолговатый мозг контролирует некоторые функции органов, включая сердцебиение и дыхание. Мозг посылает сигналы через спинной мозг и нервы, чтобы регулировать активность остальных частей тела. В шишковидное тело, который, как известно, регулирует режим сна у людей, считается, что он производит гормоны, участвующие в спячка и праздник у земноводных.[65]
Головастики сохраняют систему боковых линий своих предковых рыб, но у взрослых земноводных она отсутствует. Некоторые цецилийцы обладают электрорецепторы которые позволяют им находить объекты вокруг себя, когда они погружены в воду. У лягушек хорошо развиты уши. Наружного уха нет, но большое круглое барабанная перепонка лежит на поверхности головы сразу за глазом. Это вибрирует, и звук передается через единственную кость, стремени, к внутреннему уху. Таким образом слышны только высокочастотные звуки, такие как брачные призывы, но низкочастотные шумы можно обнаружить с помощью другого механизма.[61] Есть участок специализированных волосковых клеток, называемый сосочек амфибиорума, во внутреннем ухе способный улавливать более глубокие звуки. Другой особенностью, уникальной для лягушек и саламандр, является комплекс колумелла-покрышка, примыкающий к слуховой капсуле, который участвует в передаче как воздушных, так и сейсмических сигналов.[66] Уши саламандр и цецилий менее развиты, чем у лягушек, поскольку они обычно не общаются друг с другом посредством звука.[67]
У головастиков нет век, но при метаморфозе роговица становится более куполообразной, линза становится более плоским, и веки и связанные железы и протоки развиваются.[61] Глаза взрослых являются улучшением по сравнению с глазами беспозвоночных и явились первым шагом в развитии более продвинутых глаз позвоночных. Они обеспечивают цветовое зрение и глубину резкости. В сетчатке есть зеленые палочки, которые воспринимают широкий диапазон длин волн.[67]
Пищеварительная и выделительная системы

Многие земноводные ловят свою добычу, высовывая удлиненный язык с липким кончиком и втягивая его обратно в рот, прежде чем схватить предмет своими челюстями. Some use inertial feeding to help them swallow the prey, repeatedly thrusting their head forward sharply causing the food to move backwards in their mouth by инерция. Most amphibians swallow their prey whole without much chewing so they possess voluminous stomachs. Короткая пищевод выложен реснички that help to move the food to the stomach and слизь produced by glands in the mouth and глотка eases its passage. Фермент хитиназа produced in the stomach helps digest the chitinous cuticle of arthropod prey.[68]
Amphibians possess a поджелудочная железа, печень и желчный пузырь. The liver is usually large with two lobes. Its size is determined by its function as a гликоген and fat storage unit, and may change with the seasons as these reserves are built or used up. Жировая ткань is another important means of storing energy and this occurs in the abdomen (in internal structures called fat bodies), under the skin and, in some salamanders, in the tail.[69]
Есть два почки located dorsally, near the roof of the body cavity. Their job is to filter the blood of metabolic waste and transport the urine via ureters to the urinary bladder where it is stored before being passed out periodically through the cloacal vent. Larvae and most aquatic adult amphibians excrete the nitrogen as ammonia in large quantities of dilute urine, while terrestrial species, with a greater need to conserve water, excrete the less toxic product urea. Some tree frogs with limited access to water excrete most of their metabolic waste as uric acid.[70]
Дыхательная система

The lungs in amphibians are primitive compared to those of amniotes, possessing few internal септа и большой альвеолы, and consequently having a comparatively slow diffusion rate for oxygen entering the blood. Ventilation is accomplished by buccal pumping.[71] Most amphibians, however, are able to exchange gases with the water or air via their skin. To enable sufficient кожное дыхание, the surface of their highly vascularised skin must remain moist to allow the oxygen to diffuse at a sufficiently high rate.[68] Because oxygen concentration in the water increases at both low temperatures and high flow rates, aquatic amphibians in these situations can rely primarily on cutaneous respiration, as in the Водяная лягушка Титикака и hellbender salamander. In air, where oxygen is more concentrated, some small species can rely solely on cutaneous gas exchange, most famously the plethodontid salamanders, which have neither lungs nor gills. Many aquatic salamanders and all tadpoles have gills in their larval stage, with some (such as the axolotl ) retaining gills as aquatic adults.[68]
Размножение

С целью воспроизведение most amphibians require пресная вода although some lay their eggs on land and have developed various means of keeping them moist. A few (e.g. Fejervarya raja ) can inhabit brackish water, but there are no true морской амфибии.[72] There are reports, however, of particular amphibian populations unexpectedly invading marine waters. Such was the case with the Черное море invasion of the natural hybrid Pelophylax esculentus reported in 2010.[73]
Several hundred frog species in адаптивные излучения (например., Eleutherodactylus, the Pacific Платимантида, the Australo-Papuan microhylids, and many other tropical frogs), however, do not need any water for breeding in the wild. They reproduce via direct development, an ecological and evolutionary adaptation that has allowed them to be completely independent from free-standing water. Almost all of these frogs live in wet тропические леса and their eggs hatch directly into miniature versions of the adult, passing through the tadpole stage within the egg. Reproductive success of many amphibians is dependent not only on the quantity of rainfall, but the seasonal timing.[74]
In the tropics, many amphibians breed continuously or at any time of year. In temperate regions, breeding is mostly seasonal, usually in the spring, and is triggered by increasing day length, rising temperatures or rainfall. Experiments have shown the importance of temperature, but the trigger event, especially in arid regions, is often a storm. In anurans, males usually arrive at the breeding sites before females and the vocal chorus they produce may stimulate ovulation in females and the endocrine activity of males that are not yet reproductively active.[75]
In caecilians, fertilisation is internal, the male extruding an intromittent organ, то phallodeum, and inserting it into the female cloaca. The paired Müllerian glands inside the male cloaca secrete a fluid which resembles that produced by mammalian предстательная железа glands and which may transport and nourish the sperm. Fertilisation probably takes place in the oviduct.[76]
The majority of salamanders also engage in внутреннее оплодотворение. In most of these, the male deposits a spermatophore, a small packet of sperm on top of a gelatinous cone, on the субстрат either on land or in the water. The female takes up the sperm packet by grasping it with the lips of the cloaca and pushing it into the vent. The spermatozoa move to the spermatheca in the roof of the cloaca where they remain until ovulation which may be many months later. Courtship rituals and methods of transfer of the spermatophore vary between species. In some, the spermatophore may be placed directly into the female cloaca while in others, the female may be guided to the spermatophore or restrained with an embrace called Amplexus. Certain primitive salamanders in the families Sirenidae, Hynobiidae и Cryptobranchidae practice external fertilisation in a similar manner to frogs, with the female laying the eggs in water and the male releasing sperm onto the egg mass.[76]
With a few exceptions, frogs use external fertilisation. The male grasps the female tightly with his forelimbs either behind the arms or in front of the back legs, or in the case of Epipedobates tricolor, around the neck. They remain in amplexus with their cloacae positioned close together while the female lays the eggs and the male covers them with sperm. Roughened nuptial pads on the male's hands aid in retaining grip. Often the male collects and retains the egg mass, forming a sort of basket with the hind feet. Исключением является granular poison frog (Oophaga granulifera) where the male and female place their cloacae in close proximity while facing in opposite directions and then release eggs and sperm simultaneously. В tailed frog (Ascaphus truei) exhibits internal fertilisation. The "tail" is only possessed by the male and is an extension of the cloaca and used to inseminate the female. This frog lives in fast-flowing streams and internal fertilisation prevents the sperm from being washed away before fertilisation occurs.[77] The sperm may be retained in storage tubes attached to the oviduct until the following spring.[78]
Most frogs can be classified as either prolonged or explosive breeders. Typically, prolonged breeders congregate at a breeding site, the males usually arriving first, calling and setting up territories. Other satellite males remain quietly nearby, waiting for their opportunity to take over a territory. The females arrive sporadically, mate selection takes place and eggs are laid. The females depart and territories may change hands. More females appear and in due course, the breeding season comes to an end. Explosive breeders on the other hand are found where temporary pools appear in dry regions after rainfall. These frogs are typically роющий species that emerge after heavy rains and congregate at a breeding site. They are attracted there by the calling of the first male to find a suitable place, perhaps a pool that forms in the same place each rainy season. The assembled frogs may call in unison and frenzied activity ensues, the males scrambling to mate with the usually smaller number of females.[77]

There is a direct competition between males to win the attention of the females in salamanders and newts, with elaborate courtship displays to keep the female's attention long enough to get her interested in choosing him to приятель с.[79] Some species store сперма through long breeding seasons, as the extra time may allow for interactions with rival sperm.[80]
Жизненный цикл
Most amphibians go through метаморфоза, a process of significant morphological change after birth. In typical amphibian development, eggs are laid in water and larvae are adapted to an aquatic lifestyle. Frogs, toads and salamanders all hatch from the egg as larvae with external gills. Metamorphosis in amphibians is regulated by тироксин concentration in the blood, which stimulates metamorphosis, and пролактин, which counteracts thyroxine's effect. Specific events are dependent on threshold values for different tissues.[81] Because most embryonic development is outside the parental body, it is subject to many adaptations due to specific environmental circumstances. For this reason tadpoles can have horny ridges instead of teeth, whisker-like skin extensions or fins. They also make use of a sensory lateral line organ similar to that of fish. After metamorphosis, these organs become redundant and will be reabsorbed by controlled cell death, called апоптоз. The variety of adaptations to specific environmental circumstances among amphibians is wide, with many discoveries still being made.[82]
Яйца

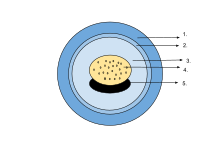
The egg of an amphibian is typically surrounded by a transparent gelatinous covering secreted by the oviducts and containing мукопротеины и мукополисахариды. This capsule is permeable to water and gases, and swells considerably as it absorbs water. The ovum is at first rigidly held, but in fertilised eggs the innermost layer liquefies and allows the эмбрион to move freely. This also happens in salamander eggs, even when they are unfertilised. Eggs of some salamanders and frogs contain unicellular green algae. These penetrate the jelly envelope after the eggs are laid and may increase the supply of oxygen to the embryo through photosynthesis. They seem to both speed up the development of the larvae and reduce mortality.[83] Most eggs contain the pigment меланин which raises their temperature through the absorption of light and also protects them against ultraviolet radiation. Caecilians, some полнокровие salamanders and certain frogs lay eggs underground that are unpigmented. в деревянная лягушка (Rana sylvatica), the interior of the globular egg cluster has been found to be up to 6 °C (11 °F) warmer than its surroundings, which is an advantage in its cool northern habitat.[84]
The eggs may be deposited singly or in small groups, or may take the form of spherical egg masses, rafts or long strings. In terrestrial caecilians, the eggs are laid in grape-like clusters in burrows near streams. The amphibious salamander Ensatina attaches its similar clusters by stalks to underwater stems and roots. В greenhouse frog (Eleutherodactylus planirostris) lays eggs in small groups in the soil where they develop in about two weeks directly into juvenile frogs without an intervening larval stage.[85] В tungara frog (Physalaemus pustulosus) builds a floating nest from foam to protect its eggs. First a raft is built, then eggs are laid in the centre, and finally a foam cap is overlaid. The foam has anti-microbial properties. Он не содержит моющие средства but is created by whipping up белки и lectins secreted by the female.[86][87]
Личинки

The eggs of amphibians are typically laid in water and hatch into free-living larvae that complete their development in water and later transform into either aquatic or terrestrial adults. In many species of frog and in most lungless salamanders (Plethodontidae), direct development takes place, the larvae growing within the eggs and emerging as miniature adults. Many caecilians and some other amphibians lay their eggs on land, and the newly hatched larvae wriggle or are transported to water bodies. Some caecilians, the альпийская саламандра (Salamandra atra) and some of the African live-bearing toads (Nectophrynoides spp.) находятся живородящий. Their larvae feed on glandular secretions and develop within the female's oviduct, often for long periods. Other amphibians, but not caecilians, are ovoviviparous. The eggs are retained in or on the parent's body, but the larvae subsist on the yolks of their eggs and receive no nourishment from the adult. The larvae emerge at varying stages of their growth, either before or after metamorphosis, according to their species.[88] The toad genus Нектофриноиды exhibits all of these developmental patterns among its dozen or so members.[6]
Лягушки
Frog larvae are known as tadpoles and typically have oval bodies and long, vertically flattened tails with fins. The free-living larvae are normally fully aquatic, but the tadpoles of some species (such as Nannophrys ceylonensis ) are semi-terrestrial and live among wet rocks.[89] Tadpoles have cartilaginous skeletons, gills for respiration (external gills at first, internal gills later), lateral line systems and large tails that they use for swimming.[90] Newly hatched tadpoles soon develop gill pouches that cover the gills. The lungs develop early and are used as accessory breathing organs, the tadpoles rising to the water surface to gulp air. Some species complete their development inside the egg and hatch directly into small frogs. These larvae do not have gills but instead have specialised areas of skin through which respiration takes place. While tadpoles do not have true teeth, in most species, the jaws have long, parallel rows of small keratinized structures called keradonts surrounded by a horny beak.[91] Front legs are formed under the gill sac and hind legs become visible a few days later.
Iodine and T4 (over stimulate the spectacular апоптоз [programmed cell death] of the cells of the larval gills, tail and fins) also stimulate the эволюция нервной системы transforming the aquatic, vegetarian tadpole into the terrestrial, carnivorous frog with better neurological, visuospatial, olfactory and cognitive abilities for hunting.[92][93]
In fact, tadpoles developing in ponds and streams are typically травоядный. Pond tadpoles tend to have deep bodies, large caudal fins and small mouths; they swim in the quiet waters feeding on growing or loose fragments of vegetation. Stream dwellers mostly have larger mouths, shallow bodies and caudal fins; they attach themselves to plants and stones and feed on the surface films of водоросли и бактерии.[94] Они также питаются диатомеи, filtered from the water through the жабры, and stir up the sediment at bottom of the pond, ingesting edible fragments. They have a relatively long, spiral-shaped gut to enable them to digest this diet.[94] Some species are carnivorous at the tadpole stage, eating insects, smaller tadpoles and fish. Young of the Кубинская древесная лягушка (Остеопилус септентрионалис) can occasionally be каннибалистический, the younger tadpoles attacking a larger, more developed tadpole when it is undergoing metamorphosis.[95]
At metamorphosis, rapid changes in the body take place as the lifestyle of the frog changes completely. The spiral‐shaped mouth with horny tooth ridges is reabsorbed together with the spiral gut. The animal develops a large jaw, and its gills disappear along with its gill sac. Eyes and legs grow quickly, and a tongue is formed. There are associated changes in the neural networks such as development of stereoscopic vision and loss of the lateral line system. All this can happen in about a day. A few days later, the tail is reabsorbed, due to the higher thyroxine concentration required for this to take place.[94]
Саламандры

(Макродактилум амбистомы)

(Макродактилум амбистомы)

(Ichthyosaura alpestris)
At hatching, a typical salamander larva has eyes without lids, teeth in both upper and lower jaws, three pairs of feathery external gills, a somewhat laterally flattened body and a long tail with спинной и вентральный плавники. The forelimbs may be partially developed and the hind limbs are rudimentary in pond-living species but may be rather more developed in species that reproduce in moving water. Pond-type larvae often have a pair of balancers, rod-like structures on either side of the head that may prevent the gills from becoming clogged up with sediment. Some members of the genera Амбистома и Dicamptodon have larvae that never fully develop into the adult form, but this varies with species and with populations. В northwestern salamander (Ambystoma gracile) is one of these and, depending on environmental factors, either remains permanently in the larval state, a condition known as neoteny, or transforms into an adult.[96] Both of these are able to breed.[97] Neoteny occurs when the animal's growth rate is very low and is usually linked to adverse conditions such as low water temperatures that may change the response of the tissues to the hormone thyroxine.[98] Other factors that may inhibit metamorphosis include lack of food, lack of trace elements and competition from сородичи. В тигровая саламандра (Тигринум амбистома) also sometimes behaves in this way and may grow particularly large in the process. The adult tiger salamander is terrestrial, but the larva is aquatic and able to breed while still in the larval state. When conditions are particularly inhospitable on land, larval breeding may allow continuation of a population that would otherwise die out. There are fifteen species of обязать neotenic salamanders, including species of Нектур, Протей и Амфиума, and many examples of факультативный ones that adopt this strategy under appropriate environmental circumstances.[99]
Lungless salamanders in the family Plethodontidae are terrestrial and lay a small number of unpigmented eggs in a cluster among damp leaf litter. Each egg has a large yolk sac and the larva feeds on this while it develops inside the egg, emerging fully formed as a juvenile salamander. The female salamander often broods the eggs. In the genus Ensatinas, the female has been observed to coil around them and press her throat area against them, effectively massaging them with a mucous secretion.[100]
In newts and salamanders, metamorphosis is less dramatic than in frogs. This is because the larvae are already carnivorous and continue to feed as predators when they are adults so few changes are needed to their digestive systems. Their lungs are functional early, but the larvae do not make as much use of them as do tadpoles. Their gills are never covered by gill sacs and are reabsorbed just before the animals leave the water. Other changes include the reduction in size or loss of tail fins, the closure of gill slits, thickening of the skin, the development of eyelids, and certain changes in dentition and tongue structure. Salamanders are at their most vulnerable at metamorphosis as swimming speeds are reduced and transforming tails are encumbrances on land.[101] Adult salamanders often have an aquatic phase in spring and summer, and a land phase in winter. For adaptation to a water phase, prolactin is the required hormone, and for adaptation to the land phase, thyroxine. External gills do not return in subsequent aquatic phases because these are completely absorbed upon leaving the water for the first time.[102]
Цецилийцы

Most terrestrial caecilians that lay eggs do so in burrows or moist places on land near bodies of water. The development of the young of Ichthyophis glutinosus, a species from Sri Lanka, has been much studied. The eel-like larvae hatch out of the eggs and make their way to water. They have three pairs of external red feathery gills, a blunt head with two rudimentary eyes, a lateral line system and a short tail with fins. They swim by undulating their body from side to side. They are mostly active at night, soon lose their gills and make sorties onto land. Metamorphosis is gradual. By the age of about ten months they have developed a pointed head with sensory tentacles near the mouth and lost their eyes, lateral line systems and tails. The skin thickens, embedded scales develop and the body divides into segments. By this time, the caecilian has constructed a burrow and is living on land.[103]

In the majority of species of caecilians, the young are produced by viviparity. Typhlonectes compressicauda, a species from South America, is typical of these. Up to nine larvae can develop in the oviduct at any one time. They are elongated and have paired sac-like gills, small eyes and specialised scraping teeth. At first, they feed on the yolks of the eggs, but as this source of nourishment declines they begin to rasp at the ciliated epithelial cells that line the oviduct. This stimulates the secretion of fluids rich in липиды and mucoproteins on which they feed along with scrapings from the oviduct wall. They may increase their length sixfold and be two-fifths as long as their mother before being born. By this time they have undergone metamorphosis, lost their eyes and gills, developed a thicker skin and mouth tentacles, and reabsorbed their teeth. A permanent set of teeth grow through soon after birth.[104][105]
The ringed caecilian (Siphonops annulatus ) has developed a unique adaptation for the purposes of reproduction. The progeny feed on a skin layer that is specially developed by the adult in a phenomenon known as maternal dermatophagy. The brood feed as a batch for about seven minutes at intervals of approximately three days which gives the skin an opportunity to regenerate. Meanwhile, they have been observed to ingest fluid exuded from the maternal cloaca.[106]
Родительская забота

The care of offspring among amphibians has been little studied but, in general, the larger the number of eggs in a batch, the less likely it is that any degree of parental care takes place. Nevertheless, it is estimated that in up to 20% of amphibian species, one or both adults play some role in the care of the young.[107] Those species that breed in smaller water bodies or other specialised habitats tend to have complex patterns of behaviour in the care of their young.[108]
Many woodland salamanders lay clutches of eggs under dead logs or stones on land. В black mountain salamander (Desmognathus welteri) does this, the mother brooding the eggs and guarding them from predation as the embryos feed on the yolks of their eggs. When fully developed, they break their way out of the egg capsules and disperse as juvenile salamanders.[109] The male hellbender, a primitive salamander, excavates an underwater nest and encourages females to lay there. The male then guards the site for the two or three months before the eggs hatch, using body undulations to fan the eggs and increase their supply of oxygen.[51]

Мужчина Colostethus subpunctatus, a tiny frog, protects the egg cluster which is hidden under a stone or log. When the eggs hatch, the male transports the tadpoles on his back, stuck there by a mucous secretion, to a temporary pool where he dips himself into the water and the tadpoles drop off.[110] Мужчина акушерка жаба (Alytes obstetricans) winds egg strings round his thighs and carries the eggs around for up to eight weeks. He keeps them moist and when they are ready to hatch, he visits a pond or ditch and releases the tadpoles.[111] Женщина gastric-brooding frog (Rheobatrachus spp.) reared larvae in her stomach after swallowing either the eggs or hatchlings; however, this stage was never observed before the species became extinct. The tadpoles secrete a hormone that inhibits digestion in the mother whilst they develop by consuming their very large yolk supply.[112] В pouched frog (Assa darlingtoni) lays eggs on the ground. When they hatch, the male carries the tadpoles around in brood pouches on his hind legs.[113] The aquatic Суринамская жаба (Пипа пипа) raises its young in pores on its back where they remain until metamorphosis.[114] The granular poison frog (Oophaga granulifera) is typical of a number of tree frogs in the poison dart frog family Dendrobatidae. Its eggs are laid on the forest floor and when they hatch, the tadpoles are carried one by one on the back of an adult to a suitable water-filled crevice such as the пазуха of a leaf or the розетка из бромелия. The female visits the nursery sites regularly and deposits unfertilised eggs in the water and these are consumed by the tadpoles.[115]
Feeding and diet

(Ambystoma gracile) eating a worm
With a few exceptions, adult amphibians are хищники, feeding on virtually anything that moves that they can swallow. The diet mostly consists of small prey that do not move too fast such as beetles, caterpillars, earthworms and spiders. The sirens (Siren spp. ) often ingest aquatic plant material with the invertebrates on which they feed[116] and a Brazilian tree frog (Xenohyla truncata ) includes a large quantity of fruit in its diet.[117] В Мексиканская роющая жаба (Носорог спинной) has a specially adapted tongue for picking up ants and termites. It projects it with the tip foremost whereas other frogs flick out the rear part first, their tongues being hinged at the front.[118]
Food is mostly selected by sight, even in conditions of dim light. Movement of the prey triggers a feeding response. Frogs have been caught on fish hooks baited with red flannel and green frogs (Рана кламитанс) have been found with stomachs full of elm seeds that they had seen floating past.[119] Toads, salamanders and caecilians also use smell to detect prey. This response is mostly secondary because salamanders have been observed to remain stationary near odoriferous prey but only feed if it moves. Cave-dwelling amphibians normally hunt by smell. Some salamanders seem to have learned to recognize immobile prey when it has no smell, even in complete darkness.[120]
Amphibians usually swallow food whole but may chew it lightly first to subdue it.[40] They typically have small hinged pedicellate teeth, a feature unique to amphibians. The base and crown of these are composed of dentine разделены uncalcified layer and they are replaced at intervals. Salamanders, caecilians and some frogs have one or two rows of teeth in both jaws, but some frogs (Rana spp.) lack teeth in the lower jaw, and toads (Bufo spp.) have no teeth. In many amphibians there are also сошковые зубы attached to a facial bone in the roof of the mouth.[121]

В тигровая саламандра (Тигринум амбистома) is typical of the frogs and salamanders that hide under cover ready to ambush unwary invertebrates. Others amphibians, such as the Bufo spp. toads, actively search for prey, while the Argentine horned frog (Ceratophrys ornata) lures inquisitive prey closer by raising its hind feet over its back and vibrating its yellow toes.[122] Among leaf litter frogs in Panama, frogs that actively hunt prey have narrow mouths and are slim, often brightly coloured and toxic, while ambushers have wide mouths and are broad and well-camouflaged.[123] Caecilians do not flick their tongues, but catch their prey by grabbing it with their slightly backward-pointing teeth. The struggles of the prey and further jaw movements work it inwards and the caecilian usually retreats into its burrow. The subdued prey is gulped down whole.[124]
When they are newly hatched, frog larvae feed on the yolk of the egg. When this is exhausted some move on to feed on bacteria, algal crusts, detritus and raspings from submerged plants. Water is drawn in through their mouths, which are usually at the bottom of their heads, and passes through branchial food traps between their mouths and their gills where fine particles are trapped in mucus and filtered out. Others have specialised mouthparts consisting of a horny beak edged by several rows of labial teeth. They scrape and bite food of many kinds as well as stirring up the bottom sediment, filtering out larger particles with the papillae around their mouths. Some, such as the spadefoot toads, have strong biting jaws and are carnivorous or even cannibalistic.[125]
Вокализация

The calls made by caecilians and salamanders are limited to occasional soft squeaks, grunts or hisses and have not been much studied. A clicking sound sometimes produced by caecilians may be a means of orientation, as in bats, or a form of communication. Most salamanders are considered voiceless, but the Калифорнийская гигантская саламандра (Dicamptodon ensatus) has vocal cords and can produce a rattling or barking sound. Some species of salamander emit a quiet squeak or yelp if attacked.[126]

Frogs are much more vocal, especially during the breeding season when they use their voices to attract mates. The presence of a particular species in an area may be more easily discerned by its characteristic call than by a fleeting glimpse of the animal itself. In most species, the sound is produced by expelling air from the lungs over the vocal cords into an air sac or sacs in the throat or at the corner of the mouth. This may distend like a balloon and acts as a resonator, helping to transfer the sound to the atmosphere, or the water at times when the animal is submerged.[126] The main vocalisation is the male's loud advertisement call which seeks to both encourage a female to approach and discourage other males from intruding on its territory. This call is modified to a quieter courtship call on the approach of a female or to a more aggressive version if a male intruder draws near. Calling carries the risk of attracting predators and involves the expenditure of much energy.[127] Other calls include those given by a female in response to the advertisement call and a release call given by a male or female during unwanted attempts at amplexus. When a frog is attacked, a distress or fright call is emitted, often resembling a scream.[128] The usually nocturnal Cuban tree frog (Остеопилус септентрионалис) produces a rain call when there is rainfall during daylight hours.[129]
Территориальное поведение
Little is known of the territorial behaviour of caecilians, but some frogs and salamanders defend home ranges. These are usually feeding, breeding or sheltering sites. Males normally exhibit such behaviour though in some species, females and even juveniles are also involved. Although in many frog species, females are larger than males, this is not the case in most species where males are actively involved in territorial defence. Some of these have specific adaptations such as enlarged teeth for biting or spines on the chest, arms or thumbs.[130]

In salamanders, defence of a territory involves adopting an aggressive posture and if necessary attacking the intruder. This may involve snapping, chasing and sometimes biting, occasionally causing the loss of a tail. The behaviour of red back salamanders (Plethodon cinereus) has been much studied. 91% of marked individuals that were later recaptured were within a metre (yard) of their original daytime retreat under a log or rock.[131] A similar proportion, when moved experimentally a distance of 30 metres (98 ft), found their way back to their home base.[131] The salamanders left odour marks around their territories which averaged 0.16 to 0.33 square metres (1.7 to 3.6 sq ft) in size and were sometimes inhabited by a male and female pair.[132] These deterred the intrusion of others and delineated the boundaries between neighbouring areas. Much of their behaviour seemed stereotyped and did not involve any actual contact between individuals. An aggressive posture involved raising the body off the ground and glaring at the opponent who often turned away submissively. If the intruder persisted, a biting lunge was usually launched at either the tail region or the naso-labial grooves. Damage to either of these areas can reduce the fitness of the rival, either because of the need to regenerate tissue or because it impairs its ability to detect food.[131]
In frogs, male territorial behaviour is often observed at breeding locations; calling is both an announcement of ownership of part of this resource and an advertisement call to potential mates. In general, a deeper voice represents a heavier and more powerful individual, and this may be sufficient to prevent intrusion by smaller males. Much energy is used in the vocalization and it takes a toll on the territory holder who may be displaced by a fitter rival if he tires. There is a tendency for males to tolerate the holders of neighbouring territories while vigorously attacking unknown intruders. Holders of territories have a "home advantage" and usually come off better in an encounter between two similar-sized frogs. If threats are insufficient, chest to chest tussles may take place. Fighting methods include pushing and shoving, deflating the opponent's vocal sac, seizing him by the head, jumping on his back, biting, chasing, splashing, and ducking him under the water.[133]
Защитные механизмы

Amphibians have soft bodies with thin skins, and lack claws, defensive armour, or spines. Nevertheless, they have evolved various defence mechanisms to keep themselves alive. The first line of defence in salamanders and frogs is the mucous secretion that they produce. This keeps their skin moist and makes them slippery and difficult to grip. The secretion is often sticky and distasteful or toxic.[134] Snakes have been observed yawning and gaping when trying to swallow Африканские когтистые лягушки (Xenopus laevis), which gives the frogs an opportunity to escape.[134][135] Caecilians have been little studied in this respect, but the Cayenne caecilian (Typhlonectes compressicauda) produces toxic mucus that has killed predatory fish in a feeding experiment in Brazil.[136] In some salamanders, the skin is poisonous. В шершавый тритон (Taricha granulosa) from North America and other members of its genus contain the neurotoxin тетродотоксин (TTX), the most toxic non-protein substance known and almost identical to that produced by рыба фугу. Handling the newts does not cause harm, but ingestion of even the most minute amounts of the skin is deadly. In feeding trials, fish, frogs, reptiles, birds and mammals were all found to be susceptible.[137][138] The only predators with some tolerance to the poison are certain populations of обычная подвязка змея (Тамнофис сирталис).In locations where both snake and salamander co-exist, the snakes have developed immunity through genetic changes and they feed on the amphibians with impunity.[139] Коэволюция occurs with the newt increasing its toxic capabilities at the same rate as the snake further develops its immunity.[138] Some frogs and toads are toxic, the main poison glands being at the side of the neck and under the warts on the back. These regions are presented to the attacking animal and their secretions may be foul-tasting or cause various physical or neurological symptoms. Altogether, over 200 toxins have been isolated from the limited number of amphibian species that have been investigated.[140]


Poisonous species often use bright colouring to warn potential predators of their toxicity. These warning colours tend to be red or yellow combined with black, with the огненная саламандра (Саламандра саламандра) being an example. Once a predator has sampled one of these, it is likely to remember the colouration next time it encounters a similar animal. У некоторых видов, таких как fire-bellied toad (Bombina spp.), the warning colouration is on the belly and these animals adopt a defensive pose when attacked, exhibiting their bright colours to the predator. The frog Allobates zaparo is not poisonous, but имитирует the appearance of other toxic species in its locality, a strategy that may deceive predators.[142]
Многие амфибии ведут ночной образ жизни и днем прячутся, избегая дневных хищников, которые охотятся визуально. Другие амфибии используют камуфляж чтобы избежать обнаружения. Они имеют различные цвета, такие как пятнистые коричневые, серые и оливковые цвета, которые сливаются с фоном. Некоторые саламандры принимают защитные позы, когда сталкиваются с потенциальным хищником, таким как североамериканский северная короткохвостая бурозубка (Blarina brevicauda). Их тела корчатся, они поднимают и хлестают хвостом, что мешает хищнику избежать контакта с их производящими яд зернистыми железами.[143] Некоторые саламандры при атаке автоматизируют свой хвост, жертвуя этой частью своей анатомии, чтобы позволить им сбежать. У основания хвоста может быть сужение, чтобы его можно было легко отсоединить. Хвост регенерируется позже, но затраты энергии животного на его замену значительны.[63]Некоторые лягушки и жабы надуваются, чтобы выглядеть большими и свирепыми, а некоторые лопатоногие жабы (Pelobates spp) кричать и прыгать к нападающему.[40] Гигантские саламандры из рода Андриас, а также Цератофрин и Пиксицефал лягушки обладают острыми зубами и способны кровоточить при оборонительном укусе. В чернобрюхая саламандра (Desmognathus quadramaculatus) может укусить нападающую обыкновенную подвязочную змею (Тамнофис сирталис) в два или три раза больше его размера на голове, и часто ему удается убежать.[144]
Познание
У амфибий есть свидетельства привыкание, ассоциативное обучение через оба классический и инструментальное обучение, и способности различения.[145]
В одном эксперименте, когда предлагали живым плодовым мушкам (Дрозофила вирилис), саламандры выбрали большее из 1 против 2 и 2 против 3. Лягушки могут различать низкие числа (1 против 2, 2 против 3, но не 3 против 4) и большие числа (3 против 6, 4 против 8, но не 4 против 6) добычи. Это не зависит от других характеристик, например площади поверхности, объема, веса и движения, хотя различие между большими числами может быть основано на площади поверхности.[146]
Сохранение

Резкое сокращение популяции амфибий, включая катастрофы популяции и массовые локализованные вымирание, были отмечены с конца 1980-х годов во всем мире, и сокращение численности амфибий, таким образом, считается одной из наиболее серьезных угроз глобальному развитию. биоразнообразие.[147] В 2004 г. Международный союз охраны природы (МСОП) сообщил, что в настоящее время птицы,[148] темпы исчезновения млекопитающих и земноводных были как минимум в 48 раз выше, чем темпы естественного вымирания - возможно, в 1024 раза выше. В 2006 году считалось, что 4 035 видов земноводных зависели от воды на определенном этапе своего жизненного цикла. Из них 1356 (33,6%) считались находящимися под угрозой исчезновения, и эта цифра, вероятно, будет заниженной, поскольку не включает 1427 видов, по которым не было достаточных данных для оценки их статуса.[149] Предполагается, что это связано с рядом причин, в том числе разрушение среды обитания и модификация, чрезмерная эксплуатация, загрязнение, интродуцированные виды, глобальное потепление, загрязнители, нарушающие работу эндокринной системы, разрушение озоновый слой (ультрафиолетовая радиация особенно опасны для кожи, глаз и яиц земноводных), а также таких заболеваний, как хитридиомикоз. Однако многие причины сокращения численности амфибий до сих пор плохо изучены и являются предметом постоянных дискуссий.[150]

С их сложными репродуктивными потребностями и проницаемой кожей земноводных часто считают экологические показатели.[151] Во многих наземных экосистемах они составляют одну из самых больших частей биомассы позвоночных. Любое сокращение численности амфибий повлияет на характер хищничества. Исчезновение хищных видов в верхней части пищевой цепи нарушит хрупкий баланс экосистемы и может вызвать резкое увеличение числа условно-патогенных видов. На Ближнем Востоке растущий аппетит к поеданию лягушачьих лапок и последующий сбор их для еды был связан с увеличением комары.[152] Хищники, питающиеся амфибиями, страдают от их упадка. В Западная земная подвязка змея (Thamnophis elegans) в Калифорнии в основном водный и сильно зависит от двух видов лягушек, численность которых сокращается: Йосемитская жаба (Bufo Canorus) и горная желтоногая лягушка (Rana muscosa), ставя под угрозу будущее змеи. Если змея станет редкостью, это повлияет на хищных птиц и других хищников, которые ею питаются.[153] Между тем, в прудах и озерах меньше лягушек - меньше головастиков. Обычно они играют важную роль в контроле роста водорослей, а также в кормлении детрит который накапливается в виде осадка на дне. Уменьшение количества головастиков может привести к чрезмерному росту водорослей, что приведет к истощению кислорода в воде, когда водоросли позже погибнут и разлагаются. В этом случае водные беспозвоночные и рыбы могут погибнуть, что приведет к непредсказуемым экологическим последствиям.[154]
А глобальная стратегия Чтобы остановить кризис, был выпущен в 2005 году План действий по сохранению амфибий. В этом призыве к действию, разработанном более чем восьмидесятью ведущими специалистами в этой области, подробно описывается, что потребуется для ограничения сокращения и исчезновения амфибий в течение следующих пяти лет и сколько это будет стоить. Группа специалистов по амфибиям МСОП возглавляет усилия по реализации всеобъемлющей глобальной стратегии сохранения амфибий.[155] Амфибия Ковчег - это организация, которая была создана для выполнения рекомендаций этого плана по сохранению ex-situ, и они работали с зоопарками и аквариумами по всему миру, поощряя их к созданию гарантированных колоний амфибий, находящихся под угрозой исчезновения.[155] Одним из таких проектов является Панамский проект по спасению и сохранению амфибий, основанный на существующих в Панаме усилиях по сохранению, чтобы создать общенациональный ответ на угрозу хитридиомикоза.[156]
Смотрите также
- Список амфибий
- Список родов амфибий
- Список находящихся под угрозой исчезновения рептилий и амфибий Соединенных Штатов
Рекомендации
- ^ а б Блэкберн, Д. С .; Уэйк, Д. Б. (2011). «Класс Amphibia Gray, 1825. В: Zhang, Z.-Q. (Ed.) Биоразнообразие животных: план классификации более высокого уровня и обзор таксономического богатства» (PDF). Zootaxa. 3148: 39–55. Дои:10.11646 / zootaxa.3148.1.8.
- ^ Скит, Уолтер В. (1897). Краткий этимологический словарь английского языка. Кларендон Пресс. п. 39.
- ^ Бэрд, Дональд (май 1965 г.). «Палеозойские лепоспондильные амфибии». Интегративная и сравнительная биология. 5 (2): 287–294. Дои:10.1093 / icb / 5.2.287.
- ^ Фрост, Даррел (2013). "Американский музей естественной истории: виды земноводных в мире 5.6, справочная информация в Интернете". Американский музей естественной истории. Получено 24 октября, 2013.
- ^ http://amphibiaweb.org:8000/lists/index.shtml
- ^ а б Крамп, Марта Л. (2009). «Разнообразие амфибий и история жизни» (PDF). Экология и сохранение амфибий. Справочник по методам: 3–20. Архивировано из оригинал (PDF) 15 июля 2011 г.
- ^ Speer, B.W .; Ваггонер, Бен (1995). «Амфибия: Систематика». Музей палеонтологии Калифорнийского университета. Получено 13 декабря, 2012.
- ^ а б c Стеббинс и Коэн 1995, п. 3.
- ^ а б c Андерсон, Дж .; Reisz, R .; Scott, D .; Fröbisch, N .; Сумида, С. (2008). «Стеблевой батрахиан из ранней перми Техаса и происхождение лягушек и саламандр». Природа. 453 (7194): 515–518. Bibcode:2008Натура.453..515A. Дои:10.1038 / природа06865. PMID 18497824. S2CID 205212809.
- ^ Рочек, З. (2000). «14. Мезозойские амфибии» (PDF). В Heatwole, H .; Кэрролл, Р. Л. (ред.). Биология амфибий: палеонтология: эволюционная история амфибий. 4. Суррей Битти и сыновья. С. 1295–1331. ISBN 978-0-949324-87-0.
- ^ Дженкинс, Фариш А. мл .; Уолш, Денис М .; Кэрролл, Роберт Л. (2007). "Анатомия Eocaecilia micropodia, конечностный цецилий ранней юры ». Вестник Музея сравнительной зоологии. 158 (6): 285–365. Дои:10.3099 / 0027-4100 (2007) 158 [285: AOEMAL] 2.0.CO; 2.
- ^ Гаоа, Кэ-Цинь; Шубин, Нил Х. (2012). «Позднеюрский саламандроид из западного Ляонина, Китай». Труды Национальной академии наук Соединенных Штатов Америки. 109 (15): 5767–5772. Bibcode:2012PNAS..109.5767G. Дои:10.1073 / pnas.1009828109. ЧВК 3326464. PMID 22411790.
- ^ Каннателла, Дэвид (2008). "Салиенция". Веб-проект "Древо жизни". Получено 31 августа, 2012.
- ^ а б c «Эволюция амфибий». Университет Вайкато: эволюция растений и животных. Получено 30 сентября, 2012.
- ^ а б c Кэрролл, Роберт Л. (1977). Халлам, Энтони (ред.). Модели эволюции, как показано в летописи окаменелостей. Эльзевир. С. 405–420. ISBN 978-0-444-41142-6.
- ^ а б Клак, Дженнифер А. (2006). "Ихтиостега". Веб-проект "Древо жизни". Получено 29 сентября, 2012.
- ^ Lombard, R.E .; Болт, Дж. Р. (1979). «Эволюция уха четвероногих: анализ и переосмысление». Биологический журнал Линнеевского общества. 11 (1): 19–76. Дои:10.1111 / j.1095-8312.1979.tb00027.x.
- ^ а б c Spoczynska, J.O.I. (1971). Ископаемые: исследование эволюции. Фредерик Мюллер Лтд., Стр. 120–125. ISBN 978-0-584-10093-8.
- ^ Sahney, S .; Бентон, М.Дж. и Ферри, П.А. (2010). «Связи между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше». Письма о биологии. 6 (4): 544–547. Дои:10.1098 / рсбл.2009.1024. ЧВК 2936204. PMID 20106856.
- ^ Sahney, S .; Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен». Труды Королевского общества B: биологические науки. 275 (1636): 759–65. Дои:10.1098 / rspb.2007.1370. ЧВК 2596898. PMID 18198148.
- ^ Сан-Мауро, Д. (2010). «Мультилокусная шкала времени происхождения современных амфибий». Молекулярная филогенетика и эволюция. 56 (2): 554–561. Дои:10.1016 / j.ympev.2010.04.019. PMID 20399871.
- ^ Сан-Мауро, Диего; Vences, Мигель; Алькобендас, Марина; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие живых земноводных предшествовало распаду Пангеи». Американский натуралист. 165 (5): 590–599. Дои:10.1086/429523. PMID 15795855. S2CID 17021360.
- ^ Крошечные окаменелости раскрывают предысторию самой загадочной из ныне живущих амфибий
- ^ а б c d Дорит, Уокер и Барнс 1991 С. 843–859.
- ^ Лорен, Мишель (2011). «Наземные позвоночные». Веб-проект "Древо жизни". Получено 16 сентября, 2012.
- ^ Лорин, Мишель; Готье, Жак А. (2012). «Амниота». Веб-проект "Древо жизни". Получено 16 сентября, 2012.
- ^ Sumich, James L .; Моррисси, Джон Ф. (2004). Введение в биологию морской жизни. Джонс и Бартлетт Обучение. п. 171. ISBN 978-0-7637-3313-1.
- ^ Брэд Шаффер; Оскар Флорес-Виллела; Габриэла Парра-Олеа; Дэвид Уэйк (2004). «Ambystoma andersoni». Красный список исчезающих видов МСОП. Версия 2013.2. Международный союз охраны природы
- ^ Экология и поведение амфибий
- ^ Проблемы биологического масштабирования и решения у земноводных - NCBI
- ^ Эволюция амфибий: жизнь ранних наземных позвоночных
- ^ Мишель Лорен (2004). «Эволюция размеров тела, правило Коупа и происхождение амниот». Систематическая биология. 53 (4): 594–622. Дои:10.1080/10635150490445706. PMID 15371249.
- ^ Rittmeyer, Eric N .; Эллисон, Аллен; Грюндлер, Майкл С .; Томпсон, Деррик К .; Остин, Кристофер С. (2012). «Эволюция экологической гильдии и открытие самого маленького позвоночного в мире». PLOS ONE. 7 (1): e29797. Bibcode:2012PLoSO ... 729797R. Дои:10.1371 / journal.pone.0029797. ЧВК 3256195. PMID 22253785.
- ^ а б Нгуен, Брент; Каваньяро, Джон (июль 2012 г.). "Факты об амфибиях". АмфибияВеб. Получено 9 ноября, 2012.
- ^ Прайс, Л. И. (1948). "Um anfibio Labirinthodonte da formacao Pedra de Fogo, Estado do Maranhao". Болетим. Ministryio da Agricultura, Departamento Nacional da Producao Inral Divisao de Geologia e Mineralogia. 24: 7–32.
- ^ Стеббинс и Коэн 1995 С. 24–25.
- ^ Каннателла, Дэвид; Грейбил, Анна (2008). "Bufonidae, настоящие жабы". Веб-проект "Древо жизни". Получено 1 декабря, 2012.
- ^ "Забавные факты о лягушках". Американский музей естественной истории. 12 января 2010 г.. Получено 29 августа, 2012.
- ^ Челленджер, Дэвид (12 января 2012 г.). «Самая маленькая лягушка в мире обнаружена в Папуа-Новой Гвинее». CNN. Архивировано из оригинал 20 апреля 2012 г.. Получено 29 августа, 2012.
- ^ а б c d е ж грамм Арнольд, Николас; Овенден, Денис (2002). Рептилии и амфибии Британии и Европы. Издательство Харпер Коллинз. С. 13–18. ISBN 978-0-00-219318-4.
- ^ Файвович, Дж .; Haddad, C.F.B .; Garcia, P.CA .; Frost, D. R .; Кэмпбелл, Дж. А .; Уиллер, В. К. (2005). «Систематический обзор семейства лягушек Hylidae, со специальной ссылкой на Hylinae: филогенетический анализ и пересмотр». Бюллетень Американского музея естественной истории. 294: 1–240. CiteSeerX 10.1.1.470.2967. Дои:10.1206 / 0003-0090 (2005) 294 [0001: SROTFF] 2.0.CO; 2.
- ^ а б Ford, L. S .; Каннателла, Д. К. (1993). «Основные клады лягушек». Герпетологические монографии. 7: 94–117. Дои:10.2307/1466954. JSTOR 1466954.
- ^ Сан-Мауро, Диего; Vences, Мигель; Алькобендас, Марина; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие живых земноводных предшествовало распаду Пангеи». Американский натуралист. 165 (5): 590–599. Дои:10.1086/429523. PMID 15795855. S2CID 17021360.
- ^ Баум, Дэвид (2008). «Эволюция черт на филогенетическом дереве: родство, сходство и миф об эволюционном развитии». Природное образование. Получено 1 декабря, 2012.
- ^ Спарребум, Макс (7 февраля 2000 г.). "Андриас давидиан Китайская гигантская саламандра ". АмфибияВеб. Получено 1 декабря, 2012.
- ^ Уэйк, Дэвид Б. (8 ноября 2000 г.). "Торий пеннатул". АмфибияВеб. Получено 25 августа, 2012.
- ^ Elmer, K. R .; Bonett, R.M .; Уэйк, Д. Б .; Lougheed, S. C. (4 марта 2013 г.). «Раннемиоценовое происхождение и загадочное разнообразие южноамериканских саламандр». BMC Эволюционная биология. 13 (1): 59. Дои:10.1186/1471-2148-13-59. ЧВК 3602097. PMID 23497060.
- ^ Larson, A .; Диммик, В. (1993). «Филогенетические отношения семейств саламандр: анализ соответствия между морфологическими и молекулярными признаками». Герпетологические монографии. 7 (7): 77–93. Дои:10.2307/1466953. JSTOR 1466953.
- ^ Дорит, Уокер и Барнс 1991, п. 852.
- ^ Эй, Хизер (2003). "Cryptobranchidae". Сеть разнообразия животных. университет Мичигана. Получено 25 августа, 2012.
- ^ а б Mayasich, J .; Grandmaison, D .; Филлипс, К. (1 июня 2003 г.). «Отчет об оценке статуса Eastern Hellbender» (PDF). Служба рыболовства и дикой природы США. Получено 25 августа, 2012.
- ^ а б c Уэйк, Дэвид Б. «Хвостатые». Энциклопедия Britannica Online. Британская энциклопедия. Получено 25 августа, 2012.
- ^ Коггер, Х. Г. (1998). Цвайфель, Р. Г. (ред.). Энциклопедия рептилий и амфибий. Академическая пресса. С. 69–70. ISBN 978-0-12-178560-4.
- ^ Стеббинс и Коэн 1995, п. 4.
- ^ Дорит, Уокер и Барнс 1991, п. 858.
- ^ Duellman, Уильям Э. "Гимнофиона". Энциклопедия Britannica Online. Британская энциклопедия. Получено 30 сентября, 2012.
- ^ Зильберберг, Луиза; Уэйк, Марвали Х. (1990). «Структура весов Дермофис и Микроцецилии (Amphibia: Gymnophiona) и сравнение с кожными окостенениями других позвоночных ". Журнал морфологии. 206 (1): 25–43. Дои:10.1002 / jmor.1052060104. PMID 29865751. S2CID 46929507.
- ^ Институт биоразнообразия Онтарио; Хеберт, Пол Д. Н. (12 октября 2008 г.). «Морфология и размножение амфибий». Энциклопедия Земли. Получено 15 августа, 2012.CS1 maint: использует параметр авторов (связь)
- ^ Стеббинс и Коэн 1995 С. 10–11.
- ^ Спирмен Р. И. (1973). Покровы: Учебник биологии кожи. Издательство Кембриджского университета. п.81. ISBN 978-0-521-20048-6.
Цвет кожи амфибии.
- ^ а б c d Дорит, Уокер и Барнс 1991, п. 846.
- ^ а б c Стеббинс и Коэн 1995 С. 26–36.
- ^ а б Бенески, Джон Т. младший (сентябрь 1989 г.). «Адаптивное значение хвостовой аутотомии у саламандры, Ensatina". Журнал герпетологии. 23 (3): 322–324. Дои:10.2307/1564465. JSTOR 1564465.
- ^ Дорит, Уокер и Барнс 1991, п. 306.
- ^ Стеббинс и Коэн 1995, п. 100.
- ^ Стеббинс и Коэн 1995, п. 69.
- ^ а б Duellman, William E .; Цуг, Джордж Р. (2012). "Амфибия". Энциклопедия Britannica Online. Британская энциклопедия. Получено 27 марта, 2012.
- ^ а б c Дорит, Уокер и Барнс 1991, п. 847.
- ^ Стеббинс и Коэн 1995, п. 66.
- ^ Дорит, Уокер и Барнс 1991, п. 849.
- ^ Брейнерд, Э. Л. (1999). «Новые взгляды на эволюцию механизмов вентиляции легких у позвоночных». Экспериментальная биология онлайн. 4 (2): 1–28. Дои:10.1007 / s00898-999-0002-1. S2CID 35368264.
- ^ Хопкинс Гарет Р .; Броди Эдмунд Д. младший (2015). «Появление амфибий в засоленных местообитаниях: обзор и эволюционная перспектива». Герпетологические монографии. 29 (1): 1–27. Дои:10.1655 / HERPMONOGRAPHS-D-14-00006. S2CID 83659304.
- ^ Натчев, Николай; Цанков Николай; Джем, Ричард (2011). «Нашествие зеленой лягушки в Черном море: экология обитания Pelophylax esculentus комплексная (анура, амфибия) популяция в районе лагуны Шабленска Тузла в Болгарии » (PDF). Заметки герпетологии. 4: 347–351.
- ^ Хоган, К. Майкл (31 июля 2010 г.). «Абиотический фактор». Энциклопедия Земли. Национальный совет по науке и окружающей среде. Архивировано из оригинал 8 июня 2013 г.. Получено 30 сентября, 2012.
- ^ Стеббинс и Коэн 1995 С. 140–141.
- ^ а б Duellman, Willia E .; Труб, Линда (1994). Биология амфибий. JHU Press. С. 77–79. ISBN 978-0-8018-4780-6.
- ^ а б Стеббинс и Коэн 1995 С. 154–162.
- ^ Адамс, Майкл Дж .; Перл, Кристофер А. (2005). "Ascaphus truei". АмфибияВеб. Получено 23 ноября, 2012.
- ^ Романо, Антонио; Бруни, Джакомо (2011). "Ухаживание, брачный период и сексуальное вмешательство самцов в Salamandrina perspicillata". Амфибия-Рептилии. 32 (1): 63–76. Дои:10.1163 / 017353710X541878.
- ^ Адамс, Эрика М .; Jones, A.G .; Арнольд, С. Дж. (2005). «Множественное отцовство в естественной популяции саламандры с длительным хранением спермы». Молекулярная экология. 14 (6): 1803–1810. Дои:10.1111 / j.1365-294X.2005.02539.x. PMID 15836651. S2CID 18899524.
- ^ Кикуяма, Сакаэ; Кавамура, Коусукэ; Танака, Сигэясу; Ямамото, Какутоши (1993). «Аспекты метаморфоза амфибий: гормональный контроль». Международный обзор цитологии: обзор клеточной биологии. Академическая пресса. С. 105–126. ISBN 978-0-12-364548-7.
- ^ Ньюман, Роберт А. (1992). «Адаптивная пластичность при метаморфозе амфибий». Бионаука. 42 (9): 671–678. Дои:10.2307/1312173. JSTOR 1312173.
- ^ Гилберт, Перри В. (1942). "Наблюдения за яйцами Пятнистая амбистома с особым упором на зеленые водоросли, обнаруженные внутри яичных конвертов ". Экология. 23 (2): 215–227. Дои:10.2307/1931088. JSTOR 1931088.
- ^ Уолдман, Брюс; Райан, Майкл Дж. (1983). «Тепловые преимущества коллективного откладывания яичной массы у древесных лягушек (Rana sylvatica)". Журнал герпетологии. 17 (1): 70–72. Дои:10.2307/1563783. JSTOR 1563783.
- ^ Мешака, Уолтер Э. мл. "Eleutherodactylus planirostris". АмфибияВеб. Получено 12 декабря, 2012.
- ^ Дэлгетти, Лаура; Кеннеди, Малкольм В. (2010). «Строительство дома из пенопласта: архитектура пенного гнезда для лягушек тунгара и трехэтапный процесс строительства». Письма о биологии. 6 (3): 293–296. Дои:10.1098 / rsbl.2009.0934. ЧВК 2880057. PMID 20106853.
- ^ «Белки лягушачьих пенных гнезд». Школа естественных наук Университета Глазго. Получено 24 августа, 2012.
- ^ Стеббинс и Коэн 1995, стр. 6–9.
- ^ Янзен, Питер (10 мая 2005 г.). "Nannophrys ceylonensis". АмфибияВеб. Получено 20 июля, 2012.
- ^ Duellman, W. E .; Цуг, Г. «Анура: от головастика до взрослой особи». Энциклопедия Britannica Online. Получено 13 июля, 2012.
- ^ Стеббинс и Коэн 1995 С. 179–181.
- ^ Вентури, Себастьяно (2011). «Эволюционное значение йода». Современная химическая биология. 5 (3): 155–162. Дои:10.2174/187231311796765012. ISSN 1872-3136.
- ^ Вентури, Себастьяно (2014). «Йод, ПНЖК и йодолипиды в здоровье и болезнях: эволюционная перспектива». Эволюция человека-. 29 (1–3): 185–205. ISSN 0393-9375.
- ^ а б c Duellman, William E .; Цуг, Джордж Р. (2012). "Анура". Энциклопедия Britannica Online. Британская энциклопедия. Получено 26 марта, 2012.
- ^ Крамп, Марта Л. (1986). «Каннибализм молодых головастиков: еще одна опасность метаморфоза». Копея. 1986 (4): 1007–1009. Дои:10.2307/1445301. JSTOR 1445301.
- ^ Валентин, Барри Д .; Деннис, Дэвид М. (1964). "Сравнение жаберной дуги и плавников трех родов личиночных саламандр, Риакотритон, Гиринофил, и Амбистома". Копея. 1964 (1): 196–201. Дои:10.2307/1440850. JSTOR 1440850.
- ^ Шаффер, Х. Брэдли (2005). "Амбистома изящная". АмфибияВеб. Получено Двадцать первое ноября, 2012.
- ^ Киёнага, Робин Р. «Метаморфозы против неотении (педоморфоза) у саламандр (Caudata)». Получено Двадцать первое ноября, 2012.
- ^ Duellman, William E .; Труб, Линда (1994). Биология амфибий. JHU Press. С. 191–192. ISBN 978-0-8018-4780-6.
- ^ Стеббинс и Коэн 1995, п. 196.
- ^ Шаффер, Х. Брэдли; Austin, C.C .; Хьюи, Р. Б. (1991). «Последствия метаморфозы на саламандре (Амбистома) двигательная способность » (PDF). Физиологическая зоология. 64 (1): 212–231. Дои:10.1086 / Physzool.64.1.30158520. JSTOR 30158520. S2CID 87191067.
- ^ Уэйк, Дэвид Б. (2012). «Хвостатые». Энциклопедия Britannica Online. Британская энциклопедия. Получено 26 марта, 2012.
- ^ Breckenridge, W. R .; Nathanael, S .; Перейра, Л. (1987). «Некоторые аспекты биологии и развития Ichthyophis glutinosus". Журнал зоологии. 211 (3): 437–449. Дои:10.1111 / jzo.1987.211.3.437.
- ^ Уэйк, Марвали Х. (1977). «Сохранение плода и его эволюционное значение в Amphibia: Gymnophiona». Журнал герпетологии. 11 (4): 379–386. Дои:10.2307/1562719. JSTOR 1562719.
- ^ Дуэллман, Уильям Э. (2012). "Гимнофиона". Энциклопедия Britannica Online. Британская энциклопедия. Получено 26 марта, 2012.
- ^ Уилкинсон, Марк; Купфер, Александр; Маркиз-Порту, Рафаэль; Джеффкинс, Хилари; Antoniazzi, Marta M .; Джаред, Карлос (2008). «Сто миллионов лет кожного питания? Расширенная родительская забота в неотропической цецилии (Amphibia: Gymnophiona)». Письма о биологии. 4 (4): 358–361. Дои:10.1098 / рсбл.2008.0217. ЧВК 2610157. PMID 18547909.
- ^ Крамп, Марта Л. (1996). «Родительская забота среди амфибий». Родительская забота: эволюция, механизмы и адаптивное значение. Достижения в изучении поведения. 25. С. 109–144. Дои:10.1016 / S0065-3454 (08) 60331-9. ISBN 978-0-12-004525-9.
- ^ Brown, J. L .; Morales, V .; Саммерс, К. (2010). «Ключевая экологическая черта стимулировала эволюцию двупородной заботы и моногамии у амфибии». Американский натуралист. 175 (4): 436–446. Дои:10.1086/650727. PMID 20180700. S2CID 20270737.
- ^ Дорит, Уокер и Барнс 1991 С. 853–854.
- ^ Фандиньо, Мария Клаудиа; Люддеке, Хорст; Амезкуита, Адольфо (1997). «Вокализация и личиночная транспортировка самца. Colostethus subpunctatus (Анура: Dendrobatidae) ". Амфибия-Рептилии. 18 (1): 39–48. Дои:10.1163 / 156853897X00297.
- ^ ван дер Мейден, Арье (18 января 2010 г.). "Алитес акушеры". АмфибияВеб. Получено 29 ноября, 2012.
- ^ Семейн, Э. (2002). "Реобатрахус силус". Сеть разнообразия животных. Музей зоологии Мичиганского университета. Получено 5 августа, 2012.
- ^ Герой, Жан-Марк; Кларк, Джон; Мейер, Эд (2004). "Асса Дарлингтони". Красный список исчезающих видов МСОП. Версия 2012.2. Получено Второе октября, 2019.CS1 maint: использует параметр авторов (связь)
- ^ Ла Марка, Энрике; Азеведо-Рамос, Клаудиа; Сильвано, Дебора; Coloma, Luis A .; Рон, Сантьяго; Харди, Джерри; Байер, Манфред (2010). "Пипа пипа (Суринамская жаба) ". Красный список исчезающих видов МСОП. Версия 2012.1. Получено Второе октября, 2019.
- ^ ван Вейнгаарден, Рене; Боланьос, Федерико (1992). "Родительская забота в Dendrobates granuliferus (Anura: Dendrobatidae), с описанием головастика ". Журнал герпетологии. 26 (1): 102–105. Дои:10.2307/1565037. JSTOR 1565037.
- ^ Габбард, Джесси (2000). "Сирена промежуточная: Малая сирена ". Сеть разнообразия животных. Музей зоологии Мичиганского университета. Получено 11 августа, 2012.
- ^ Да Силва, Х. Р .; Де Бритто-Перейра, М. К. (2006). «Сколько фруктов едят лягушки-плоды?» Исследование диеты Xenohyla truncata (Лиссамфибия: Anura: Hylidae) ". Журнал зоологии. 270 (4): 692–698. Дои:10.1111 / j.1469-7998.2006.00192.x.
- ^ Труб, Линда; Ганс, Карл (1983). "Специализации кормления мексиканской роющей жабы, Носорог спинной (Anura: Rhinophrynidae) " (PDF). Журнал зоологии. 199 (2): 189–208. Дои:10.1111 / j.1469-7998.1983.tb02090.x. HDL:2027.42/74489.
- ^ Гамильтон, У. Дж. Младший (1948). "Пища и пищевое поведение зеленой лягушки, Рана кламитанс Латрей, штат Нью-Йорк ". Копея. Американское общество ихтиологов и герпетологов. 1948 (3): 203–207. Дои:10.2307/1438455. JSTOR 1438455.
- ^ Стеббинс и Коэн 1995, п. 56.
- ^ Стеббинс и Коэн 1995 С. 57–58.
- ^ Рэдклифф, Чарльз У .; Чисар, Давид; Эстеп, Карен; Мерфи, Джеймс Б.; Смит, Хобарт М. (1986). «Наблюдения за приманкой педалей и движениями педалей у лягушек лептодактилид». Журнал герпетологии. 20 (3): 300–306. Дои:10.2307/1564496. JSTOR 1564496.
- ^ Тофт, Кэтрин А. (1981). «Экология питания панамских бесхвостых особей помета: закономерности в рационе и режиме кормления». Журнал герпетологии. 15 (2): 139–144. Дои:10.2307/1563372. JSTOR 1563372.
- ^ Bemis, W. E .; Schwenk, K .; Уэйк, М. Х. (1983). «Морфология и функции питающего аппарата в Dermophis mexicanus (Амфибия: Gymnophiona) ". Зоологический журнал Линнеевского общества. 77 (1): 75–96. Дои:10.1111 / j.1096-3642.1983.tb01722.x.
- ^ Стеббинс и Коэн 1995 С. 181–185.
- ^ а б Стеббинс и Коэн 1995 С. 76–77.
- ^ Салливан, Брайан К. (1992). «Половой отбор и призывное поведение у американской жабы (Буфо американский)". Копея. 1992 (1): 1–7. Дои:10.2307/1446530. JSTOR 1446530.
- ^ Толедо, Л. Ф .; Хаддад, К. Ф. Б. (2007). "Capitulo 4" (PDF). Когда лягушки кричат! Обзор защитных вокализаций ануранов (Тезис). Instituto de Biociências, Сан-Паулу.
- ^ Джонсон, Стив А. (2010). "Кубинская древесная лягушка (Остеопилус септентрионалис) во Флориде". EDIS. Университет Флориды. Получено 13 августа, 2012.
- ^ Шайн, Ричард (1979). «Половой отбор и половой диморфизм у амфибий». Копея. 1979 (2): 297–306. Дои:10.2307/1443418. JSTOR 1443418.
- ^ а б c Gergits, W. F .; Джегер, Р. Г. (1990). "Приложение к сайту красной саламандрой, Plethodon cinereus". Журнал герпетологии. 24 (1): 91–93. Дои:10.2307/1564297. JSTOR 1564297.
- ^ Каспер, Гэри С. "Plethodon cinereus". АмфибияВеб. Получено 25 сентября, 2012.
- ^ Уэллс, К. Д. (1977). «Территориальность и успешность спаривания самцов зеленой лягушки (Рана кламитанс)". Экология. 58 (4): 750–762. Дои:10.2307/1936211. JSTOR 1936211.
- ^ а б Barthalmus, G.T .; Зелински В. Дж. (1988). "Xenopus кожная слизь вызывает дискинезию полости рта, которая способствует бегству от змей ». Фармакология, биохимия и поведение. 30 (4): 957–959. Дои:10.1016/0091-3057(88)90126-8. PMID 3227042. S2CID 25434883.
- ^ Crayon, Джон Дж. "Xenopus laevis". АмфибияВеб. Получено 8 октября, 2012.
- ^ Муди, Г. Э. (1978). "Наблюдения за жизненным циклом червеца Компрессионный тифлонект (Думерил и Биброн) в бассейне Амазонки ». Канадский журнал зоологии. 56 (4): 1005–1008. Дои:10.1139 / z78-141.
- ^ Броди, Эдмунд Д. младший (1968). «Исследования кожного токсина взрослого тритона с шероховатой кожей, Taricha granulosa". Копея. 1968 (2): 307–313. Дои:10.2307/1441757. JSTOR 1441757.
- ^ а б Ханифин, Чарльз Т .; Йоцу-Ямасита, Мари; Ясумото, Такеши; Броди, Эдмунд Д.; Броди, Эдмунд Д. мл. (1999). «Токсичность опасной добычи: колебания уровней тетродотоксина внутри популяций тритона и между ними. Taricha granulosa". Журнал химической экологии. 25 (9): 2161–2175. Дои:10.1023 / А: 1021049125805. S2CID 543221.
- ^ Geffeney, Shana L .; Фудзимото, Эстер; Броди, Эдмунд Д.; Броди, Эдмунд Д. Младший; Рубен, Питер К. (2005). «Эволюционная диверсификация ТТХ-резистентных натриевых каналов при взаимодействии хищник – жертва». Природа. 434 (7034): 759–763. Bibcode:2005Натура.434..759Г. Дои:10.1038 / природа03444. PMID 15815629. S2CID 4426708.
- ^ Стеббинс и Коэн 1995, п. 110.
- ^ Паточка, Иржи; Вульф, Крауфф; Паломеке, Мария Виктория (1999). "Ядовитые лягушки-дротики и их токсины". Информационный бюллетень ASA. 5 (75). ISSN 1057-9419. Получено 29 января, 2013.
- ^ Darst, Catherine R .; Каммингс, Молли Э. (9 марта 2006 г.). «Обучение хищников способствует имитации менее токсичной модели ядовитых лягушек». Природа. 440 (7081): 208–211. Bibcode:2006Натура 440..208D. Дои:10.1038 / природа04297. PMID 16525472.
- ^ Броди, Эдмунд Д. Младший; Новак, Роберт Т .; Харви, Уильям Р. (1979). «Выделения хищников и поведение отдельных саламандр против землероек». Копея. 1979 (2): 270–274. Дои:10.2307/1443413. JSTOR 1443413.
- ^ Броди, Э. Д. мл. (1978). «Укусы и вокализация как механизмы борьбы с хищниками у наземных саламандр». Копея. 1978 (1): 127–129. Дои:10.2307/1443832. JSTOR 1443832.
- ^ Хлох, А. (2010). Что помнит саламандра после зимы? (PDF). Венский университет. Fakultät für Lebenswissenschaften.
- ^ Stancher, G .; Rugani, R .; Regolin, L .; Валлортигара, Г. (2015). «Численное различение лягушек (Bombina orientalis)». Познание животных. 18 (1): 219–229. Дои:10.1007 / s10071-014-0791-7. PMID 25108417. S2CID 16499583.
- ^ Маккаллум, М. Л. (2007). «Упадок амфибий или вымирание? Текущее снижение фоновой скорости вымирания карликов». Журнал герпетологии. 41 (3): 483–491. Дои:10.1670 / 0022-1511 (2007) 41 [483: ADOECD] 2.0.CO; 2.
- ^ "Что значит быть человеком?". Смитсоновский национальный музей естественной истории. Получено 19 ноября, 2013.
- ^ Hoekstra, J.M .; Molnar, J. L .; Jennings, M .; Revenga, C .; Spalding, M.D .; Баучер, Т. М .; Robertson, J.C .; Heibel, T. J .; Эллисон, К. (2010). «Количество видов земноводных, находящихся под угрозой исчезновения в пресноводном экорегионе». Атлас глобальной охраны природы: изменения, проблемы и возможности изменить мир к лучшему. Охрана природы. Получено 5 сентября, 2012.
- ^ «Группа специалистов-амфибий». Группа специалистов по амфибиям МСОП. Получено 30 марта, 2012.
- ^ Уоддл, Джеймс Хардин (2006). Использование амфибий как видов-индикаторов экосистемы (PDF) (Кандидат наук.). Университет Флориды.
- ^ Regier, Генри А .; Баскервиль, Гордон, Л. (1996). «Вопросы устойчивости для менеджеров ресурсов». Устойчивое восстановление региональных экосистем, деградированных в результате эксплуатации.. Издательство ДИАНА. С. 36–38. ISBN 978-0-7881-4699-2.
- ^ Дженнингс, У. Брайан; Брэдфорд, Дэвид Ф .; Джонсон, Дейл Ф. (1992). "Зависимость от подвязки змейки" Thamnophis elegans на амфибиях в Сьерра-Неваде в Калифорнии ». Журнал герпетологии. 26 (4): 503–505. Дои:10.2307/1565132. JSTOR 1565132.
- ^ Стеббинс и Коэн 1995, п. 249.
- ^ а б «План действий по сохранению амфибий». МСОП. Архивировано из оригинал 27 апреля 2012 г.. Получено 30 марта, 2012.
- ^ «Панамский проект по спасению и сохранению амфибий». Амфибия Ковчег. Архивировано с оригинал 14 июня 2010 г.. Получено 30 марта, 2012.
Цитированные тексты
- Dorit, R.L .; Уокер, В. Ф .; Барнс, Р. Д. (1991). Зоология. Издательство колледжа Сондерс. ISBN 978-0-03-030504-7.
- Стеббинс, Роберт С.; Коэн, Натан В. (1995). Естественная история амфибий. Издательство Принстонского университета. ISBN 978-0-691-03281-8.
дальнейшее чтение
- Кэрролл, Роберт Л. (1988). Палеонтология и эволюция позвоночных. В. Х. Фриман. ISBN 978-0-7167-1822-2.
- Кэрролл, Роберт Л. (2009). Восхождение земноводных: 365 миллионов лет эволюции. Издательство Университета Джона Хопкинса. ISBN 978-0-8018-9140-3.
- Duellman, William E .; Линда Труб (1994). Биология амфибий. Издательство Университета Джона Хопкинса. ISBN 978-0-8018-4780-6.
- Frost, Darrel R .; Грант, Таран; Файвович, Хулиан; Bain, Raoul H .; Хаас, Александр; Haddad, Célio F.B .; Де Са, Рафаэль О .; Ченнинг, Алан; Уилкинсон, Марк; Доннеллан, Стивен С.; Раксуорти, Кристофер Дж .; Кэмпбелл, Джонатан А .; Блотто, Борис Л .; Молер, Пол; Drewes, Роберт С.; Nussbaum, Ronald A .; Линч, Джон Д .; Грин, Дэвид М .; Уилер, Уорд С. (2006). «Амфибия Древо Жизни». Бюллетень Американского музея естественной истории. 297: 1–291. Дои:10.1206 / 0003-0090 (2006) 297 [0001: TATOL] 2.0.CO; 2. HDL:2246/5781.
- Паундс, Дж. Алан; Bustamante, Martín R .; Coloma, Luis A .; Консуэгра, Джейми А .; Фогден, Майкл П. Л .; Foster, Pru N .; Ла Марка, Энрике; Мастерс, Карен Л .; Мерино-Витери, Андрес; Пушендорф, Роберт; Рон, Сантьяго Р .; Санчес-Азофейфа, Г. Артуро; Тем не менее, Кристофер Дж .; Янг, Брюс Э. (2006). «Широко распространенное вымирание земноводных в результате эпидемических заболеваний, вызванных глобальным потеплением». Природа. 439 (7073): 161–167. Bibcode:2006Натура 439..161А. Дои:10.1038 / природа04246. PMID 16407945. S2CID 4430672.
- Стюарт, Саймон Н .; Шансон, Дженис С .; Cox, Neil A .; Янг, Брюс Э .; Rodrigues, Ana S.L .; Fischman, Debra L .; Уоллер, Роберт В. (2004). «Состояние и тенденции сокращения и исчезновения амфибий во всем мире». Наука. 306 (5702): 1783–1786. Bibcode:2004Наука ... 306.1783S. CiteSeerX 10.1.1.225.9620. Дои:10.1126 / science.1103538. PMID 15486254. S2CID 86238651.
- Стюарт, С. Н .; Hoffmann, M .; Chanson, J. S .; Cox, N.A .; Berridge, R.J .; Ramani, P .; Янг, Б. Э., ред. (2008). Земноводные мира, находящиеся под угрозой исчезновения. Опубликовано Lynx Edicions, в связи с МСОП - Всемирный союз охраны природы, Conservation International и Природа. ISBN 978-84-96553-41-5. Архивировано из оригинал 30 октября 2014 г.. Получено 30 октября, 2014.
внешняя ссылка
- Амфибии - AnimalSpot.net
- ArchéoZooThèque: рисунки скелетов амфибий : доступен в векторном, графическом и PDF форматах.
- Группа специалистов по амфибиям
- Амфибия Ковчег
- АмфибияВеб
- Глобальная оценка амфибий
- Вокализации амфибий на архивных звукозаписях







| Авторитетный контроль |
|---|