Кладограмма - Cladogram

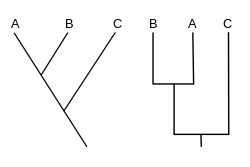
А кладограмма (от Греческий кладо "филиал" и грамма "персонаж") - диаграмма, используемая в кладистика показать отношения между организмами. Кладограмма, однако, не эволюционное дерево поскольку он не показывает, как предки связаны с потомками, и не показывает, насколько они изменились, так много различных эволюционных деревьев могут соответствовать одной кладограмме.[1][2][3][4][5] На кладограмме используются линии, которые разветвляются в разных направлениях и заканчиваются клады, группа организмов с последний общий предок. Есть много форм кладограмм, но все они имеют линии, которые отходят от других линий. Линии можно проследить до того места, где они отходят. Эти точки разветвления представляют собой гипотетического предка (а не реальную сущность), который, как можно предположить, обладал чертами, общими для терминальных таксонов над ним.[4][6] Этот гипотетический предок может затем дать подсказки о порядке эволюции различных характеристик, адаптации и других эволюционных рассказов о предках. Хотя традиционно такие кладограммы создавались в основном на основе морфологических признаков, ДНК и РНК данные секвенирования и вычислительная филогенетика в настоящее время очень часто используются при создании кладограмм либо сами по себе, либо в сочетании с морфологией.
Создание кладограммы
Эта секция нужны дополнительные цитаты для проверка. (Апрель 2016 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
Молекулярные и морфологические данные
Характеристики, используемые для создания кладограммы, можно условно разделить на две морфологические (синапсидный череп, теплокровность, нотохорда, одноклеточные и т. д.) или молекулярные (ДНК, РНК или другая генетическая информация).[7] До появления секвенирования ДНК кладистический анализ в основном использовал морфологические данные. Также могут использоваться поведенческие данные (для животных).[8]
Так как Секвенирование ДНК стало дешевле и проще, молекулярная систематика становится все более и более популярным способом вывода филогенетических гипотез.[9] Использование критерия экономности - это лишь один из нескольких методов вывода филогении на основе молекулярных данных. Такие подходы, как максимальная вероятность, которые включают явные модели эволюции последовательностей, не являются хенниговскими способами оценки данных последовательности. Еще один мощный метод реконструкции филогении - использование геномных маркеры ретротранспозонов, которые считаются менее подверженными проблеме возврат это поражает данные последовательности. Также обычно предполагается, что у них низкая частота гомоплазий, потому что когда-то считалось, что их интеграция в геном был полностью случайным; Однако иногда кажется, что это не так.

Плезиоморфии и синапоморфии
Исследователи должны решить, какие состояния характера являются «наследственными» (плезиоморфии ) и которые являются производными (синапоморфии ), потому что только синапоморфные состояния характера указывают на группировку.[10] Это определение обычно выполняется путем сравнения с состояниями символов одного или нескольких аутгруппы. Состояния, общие для внешней группы и некоторых членов внутренней группы, являются симплезиоморфиями; состояния, которые присутствуют только в подмножестве внутренней группы, являются синапоморфиями. Обратите внимание, что состояния символов, уникальные для одного терминала (аутапоморфии), не свидетельствуют о группировке. Выбор внешней группы является решающим шагом в кладистическом анализе, потому что разные внешние группы могут создавать деревья с совершенно разными топологиями.
Гомоплазии
А гомоплазия состояние характера, которое по какой-то причине разделяется двумя или более таксонами Другой чем общая родословная.[11] Двумя основными типами гомоплазии являются конвергенция (эволюция «одного и того же» персонажа по крайней мере в двух различных линиях) и реверсия (возвращение к состоянию предкового характера). Признаки, которые явно гомопластичны, такие как белый мех в различных линиях арктических млекопитающих, не должны включаться в качестве персонажа в филогенетический анализ, поскольку они ничего не вносят в наше понимание взаимоотношений. Однако гомоплазия часто не очевидна при осмотре самого признака (как, например, в последовательности ДНК), а затем обнаруживается по ее несоответствию (непродуманному распределению) на наиболее скупой кладограмме. Обратите внимание, что символы, которые являются гомопластическими, могут все еще содержать филогенетический сигнал.[12]
Хорошо известным примером гомоплазии в результате конвергентной эволюции может быть характер «наличие крыльев». Хотя крылья птиц, летучие мыши, и насекомые выполняют ту же функцию, каждое из которых развивалось независимо, как видно по их анатомия. Если бы птица, летучая мышь и крылатое насекомое оценивались по признаку «наличие крыльев», гомоплазия была бы внесена в набор данных, и это могло бы потенциально затруднить анализ, что могло бы привести к ложной гипотезе о взаимосвязях. Конечно, единственная причина, по которой гомоплазия узнаваема в первую очередь, заключается в том, что есть другие признаки, которые подразумевают паттерн взаимоотношений, раскрывающий его гомопластическое распределение.
Что не является кладограммой
Кладограмма - это схематический результат анализа, в котором таксоны группируются только на основе синапоморфий. Есть много других филогенетических алгоритмов, которые обрабатывают данные несколько иначе, и в результате получаются филогенетические деревья, которые выглядят как кладограммы, но не являются кладограммами. Например, фенетические алгоритмы, такие как UPGMA и Neighbor-Joining, группируются по общему сходству и рассматривают как синапоморфии, так и симплезиоморфии как свидетельство группировки. Полученные диаграммы являются фенограммами, а не кладограммами. Правдоподобный или байесовский подходы), которые учитывают как порядок ветвления, так и «длину ветвления», учитывают как синапоморфии, так и аутапоморфии как доказательства за или против группировки. Диаграммы, полученные в результате такого рода анализа, также не являются кладограммами.[13]
Подбор кладограммы
Есть несколько алгоритмы доступны для определения «лучшей» кладограммы.[14] Большинство алгоритмов используют метрика для измерения согласованности кладограммы кандидата с данными. Большинство алгоритмов кладограмм используют математические методы оптимизация и минимизация.
В общем, алгоритмы генерации кладограмм должны быть реализованы в виде компьютерных программ, хотя некоторые алгоритмы могут выполняться вручную, когда наборы данных скромны (например, всего несколько видов и пара характеристик).
Некоторые алгоритмы полезны только тогда, когда характерные данные являются молекулярными (ДНК, РНК); другие алгоритмы полезны только тогда, когда характеристические данные являются морфологическими. Другие алгоритмы могут использоваться, когда характеристические данные включают как молекулярные, так и морфологические данные.
Алгоритмы для кладограмм или других типов филогенетических деревьев включают: наименьших квадратов, присоединение к соседу, скупость, максимальная вероятность, и Байесовский вывод.
Биологи иногда используют термин скупость для определенного типа алгоритма генерации кладограмм, а иногда и как общий термин для всех филогенетических алгоритмов.[15]
Алгоритмы, выполняющие задачи оптимизации (например, построение кладограмм), могут быть чувствительны к порядку, в котором представлены входные данные (список видов и их характеристики). Ввод данных в различном порядке может привести к тому, что один и тот же алгоритм даст разные «лучшие» кладограммы. В таких ситуациях пользователь должен ввести данные в различном порядке и сравнить результаты.
Использование разных алгоритмов на одном наборе данных может иногда давать разные «лучшие» кладограммы, потому что каждый алгоритм может иметь уникальное определение того, что является «лучшим».
Из-за астрономического количества возможных кладограмм алгоритмы не могут гарантировать, что решение является лучшим в целом. Неоптимальная кладограмма будет выбрана, если программа ориентирована на локальный минимум, а не на желаемый глобальный минимум.[16] Чтобы решить эту проблему, многие алгоритмы кладограмм используют имитация отжига подход к увеличению вероятности того, что выбранная кладограмма является оптимальной.[17]
В базальное положение - это направление основания (или корня) укорененного филогенетического дерева или кладограммы. Базальная клада - это самая ранняя клада (данного таксономического ранга [a]), которая разветвляется внутри более крупной клады.
Статистика
Тест на несоответствие длины (или тест на однородность раздела)
Тест разницы длины неконгруэнтности (ILD) - это измерение того, как комбинация различных наборов данных (например, морфологических и молекулярных, пластидных и ядерных генов) способствует получению более длинного дерева. Он измеряется путем расчета общей длины дерева каждого раздела и их суммирования. Затем создаются реплики путем создания произвольно собранных разделов, состоящих из исходных разделов. Длины суммируются. Значение p, равное 0,01, получается для 100 повторов, если 99 реплик имеют более длинную объединенную длину дерева.
Измерение гомоплазии
Некоторые меры пытаются измерить количество гомоплазии в наборе данных со ссылкой на дерево,[18] хотя не всегда ясно, какое свойство эти меры стремятся количественно оценить[19]
Индекс согласованности
Индекс согласованности (CI) измеряет согласованность дерева набору данных - меру минимального количества гомоплазии, подразумеваемого деревом.[20] Он рассчитывается путем подсчета минимального количества изменений в наборе данных и деления его на фактическое количество изменений, необходимых для кладограммы.[20] Индекс согласованности также можно рассчитать для отдельного персонажа. я, обозначается cя.
Помимо отражения количества гомоплазии, этот показатель также отражает количество таксонов в наборе данных,[21] (в меньшей степени) количество символов в наборе данных,[22] степень, в которой каждый персонаж несет филогенетическую информацию,[23] и способ кодирования дополнительных символов, что делает его непригодным для использования.[24]
cя занимает диапазон от 1 до 1 / [n.taxa/ 2] в двоичных символах с четным распределением состояний; его минимальное значение больше, когда состояния распределены неравномерно.[23][18] Как правило, для двоичного или недвоичного символа с , cя занимает диапазон от 1 до .[23]


Индекс удержания
Индекс удержания (RI) был предложен как улучшение CI «для определенных приложений».[25] Этот показатель также предназначен для измерения степени гомоплазии, но также измеряет, насколько хорошо синапоморфии объясняют дерево. Он рассчитывается путем деления (максимальное количество изменений в дереве минус количество изменений в дереве) и деления на (максимальное количество изменений в дереве минус минимальное количество изменений в наборе данных).
Индекс согласованности с измененным масштабом (RC) получается путем умножения CI на RI; по сути, это расширяет диапазон CI таким образом, что его минимальное теоретически достижимое значение масштабируется до 0, а максимальное остается равным 1.[18][25] Индекс гомоплазии (HI) равен 1 - CI.
Коэффициент избытка гомоплазии
Это измеряет количество гомоплазии, наблюдаемой на дереве, относительно максимального количества гомоплазии, которое теоретически может присутствовать - 1 - (наблюдаемый избыток гомоплазии) / (максимальный избыток гомоплазии).[22] Значение 1 указывает на отсутствие гомоплазии; 0 представляет столько гомоплазии, сколько было бы в полностью случайном наборе данных, а отрицательные значения указывают на еще большую гомоплазию (и имеют тенденцию встречаться только в надуманных примерах).[22] HER представлен как лучший доступный в настоящее время показатель гомоплазии.[18][26]
Смотрите также
использованная литература
- ^ Майр, Эрнст (2009). «Кладистический анализ или кладистическая классификация?». Журнал зоологической систематики и эволюционных исследований. 12: 94–128. Дои:10.1111 / j.1439-0469.1974.tb00160.x.
- ^ Фут, Майк (весна 1996 г.). «О вероятности появления предков в летописи окаменелостей». Палеобиология. 22 (2): 141–51. Дои:10.1017 / S0094837300016146. JSTOR 2401114.
- ^ Дайрат, Бенуа (лето 2005 г.). «Отношения предков-потомков и реконструкция древа жизни». Палеобиология. 31 (3): 347–53. Дои:10.1666 / 0094-8373 (2005) 031 [0347: aratro] 2.0.co; 2. JSTOR 4096939.
- ^ а б Посада, Давид; Крэндалл, Кейт А. (2001). «Внутривидовые генеалогии: деревья в сети». Тенденции в экологии и эволюции. 16: 37–45. Дои:10.1016 / S0169-5347 (00) 02026-7.
- ^ Подани, Янош (2013). «Древовидное мышление, время и топология: комментарии к интерпретации древовидных диаграмм в эволюционной / филогенетической систематике» (PDF). Кладистика. 29 (3): 315–327. Дои:10.1111 / j.1096-0031.2012.00423.x.
- ^ Schuh, Рэндалл Т. (2000). Биологическая систематика: принципы и применение. ISBN 978-0-8014-3675-8.[страница нужна ]
- ^ ДеСалл, Роб (2002). Методы молекулярной систематики и эволюции. Бирхаузер. ISBN 978-3-7643-6257-7.[страница нужна ]
- ^ Венцель, Джон В. (1992). «Поведенческая гомология и филогения». Анну. Rev. Ecol. Syst. 23: 361–381. Дои:10.1146 / annurev.es.23.110192.002045.
- ^ Хиллис, Дэвид (1996). Молекулярная систематика. Синаур. ISBN 978-0-87893-282-5.[страница нужна ]
- ^ Хенниг, Вилли (1966). Филогенетическая систематика. Университет Иллинойса Press.
- ^ Вест-Эберхард, Мэри Джейн (2003). Пластичность развития и эволюция. Oxford Univ. Нажмите. стр.353 –376. ISBN 978-0-19-512235-0.
- ^ Калерсьо, Мари; Альберт, Виктор А .; Фаррис, Джеймс С. (1999). «Гомоплазия увеличивает филогенетическую структуру». Кладистика. 15: 91–93. Дои:10.1111 / j.1096-0031.1999.tb00400.x.
- ^ Брауэр, Эндрю В.З. (2016). «Что такое кладограмма, а что нет?». Кладистика. 32 (5): 573–576. Дои:10.1111 / cla.12144.
- ^ Китчинг, Ян (1998). Кладистика: теория и практика анализа экономии. Издательство Оксфордского университета. ISBN 978-0-19-850138-1.[страница нужна ]
- ^ Стюарт, Каро-Бет (1993). «Силы и подводные камни бережливости». Природа. 361 (6413): 603–7. Bibcode:1993Натура.361..603С. Дои:10.1038 / 361603a0. PMID 8437621.
- ^ Фоли, Питер (1993). Кладистика: практический курс систематики. Oxford Univ. Нажмите. п.66. ISBN 978-0-19-857766-9.
- ^ Никсон, Кевин С. (1999). "Трещотка экономичности, новый метод быстрого анализа экономии". Кладистика. 15 (4): 407–414. Дои:10.1111 / j.1096-0031.1999.tb00277.x.
- ^ а б c d рассмотрено в Арчи, Джеймс У. (1996). «Меры гомоплазии». В Sanderson, Michael J .; Хафффорд, Ларри (ред.). Гомоплазия. стр.153 –188. Дои:10.1016 / B978-012618030-5 / 50008-3. ISBN 9780126180305.
- ^ Чанг, Джозеф Т .; Ким, Чунхён (1996). «Измерение гомоплазии: стохастический взгляд». Гомоплазия. С. 189–203. Дои:10.1016 / b978-012618030-5 / 50009-5. ISBN 9780126180305.
- ^ а б Kluge, A. G .; Фаррис, Дж. С. (1969). «Количественная филетика и эволюция бесхвостых животных». Систематическая зоология. 18 (1): 1–32. Дои:10.2307/2412407. JSTOR 2412407.
- ^ Archie, J. W .; Фельзенштейн, Дж. (1993). «Число эволюционных шагов на деревьях случайной и минимальной длины для случайных эволюционных данных». Теоретическая популяционная биология. 43: 52–79. Дои:10.1006 / tpbi.1993.1003.
- ^ а б c Арчи, Дж. У. (1989). «КОЭФФИЦИЕНТЫ ПРЕВЫШЕНИЯ ГОМОПЛАЗИИ: НОВЫЕ ПОКАЗАТЕЛИ ДЛЯ ИЗМЕРЕНИЯ УРОВНЕЙ ГОМОПЛАЗИИ В ФИЛОГЕНЕТИЧЕСКОЙ СИСТЕМАТИКЕ И КРИТИК ИНДЕКСА СООТВЕТСТВИЯ». Систематическая зоология. 38 (3): 253–269. Дои:10.2307/2992286. JSTOR 2992286.
- ^ а б c Хойал Катхилл, Дженнифер Ф .; Брэдди, Саймон Дж .; Донохью, Филип С. Дж. (2010). «Формула для максимально возможных шагов в мультистатических символах: изолирующие эффекты параметра матрицы на меры эволюционной конвергенции». Кладистика. 26: 98–102. Дои:10.1111 / j.1096-0031.2009.00270.x.
- ^ Сандерсон, М. Дж .; Донохью, М. Дж. (1989). «Паттерны вариации уровней гомоплазии». Эволюция. 43 (8): 1781–1795. Дои:10.2307/2409392. JSTOR 2409392.
- ^ а б Фаррис, Дж. С. (1989). «Индекс удержания и измененный индекс согласованности». Кладистика. 5 (4): 417–419. Дои:10.1111 / j.1096-0031.1989.tb00573.x.
- ^ Хойал Катхилл, Дженнифер (2015). «Размер пространства состояний персонажа влияет на возникновение и обнаружение гомоплазии: моделирование вероятности несовместимости для неупорядоченных филогенетических символов». Журнал теоретической биологии. 366: 24–32. Дои:10.1016 / j.jtbi.2014.10.033. PMID 25451518.
внешние ссылки
 СМИ, связанные с Кладограммы в Wikimedia Commons
СМИ, связанные с Кладограммы в Wikimedia Commons
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||