Боль в рыбе - Pain in fish

| Часть серия на |
| Права животных |
|---|
Идеи |
Связанный |
Ощущают ли рыбы боль, как у людей, или иначе - вопрос спорный. Боль это сложный психическое состояние, с отчетливым качеством восприятия, но также связанные с страдания, что является эмоциональное состояние. Из-за этой сложности наличие боль в животном или другого человека в этом отношении, нельзя однозначно определить с помощью методов наблюдения, но вывод о том, что животные испытывают боль, часто делается на основе вероятного присутствия феноменальное сознание который выводится из сравнительной физиологии мозга, а также физических и поведенческих реакций.[1][2]
Рыбы соответствуют нескольким предложенным критериям, указывающим на то, что животные, не являющиеся людьми, могут испытывать боль. Эти выполненные критерии включают подходящую нервную систему и сенсорные рецепторы, опиоидные рецепторы и снижение реакции на вредные раздражители при введении анальгетики и местные анестетики, физиологические изменения на вредные раздражители, проявление защитных двигательных реакций, проявление обучение избеганию и поиск компромиссов между избеганием вредных стимулов и другими мотивационными требованиями.
Если рыба чувствует боль, существуют этические последствия и последствия для благополучия животных, включая последствия воздействия загрязняющих веществ и практики, связанные с коммерческий и любительская рыбалка, аквакультура, в декоративная рыба и генетически модифицированная рыба и для рыбы, используемой в научное исследование.
Фон
Возможность того, что рыбы и другие животные, кроме человека, могут испытывать боль, имеет давнюю историю. Первоначально это было основано на теоретических и философских аргументах, но в последнее время превратилось в научные исследования.
Философия

Идея, что животные, не являющиеся людьми, могут не чувствовать боль восходит к французскому философу 17 века, Рене Декарт, который утверждал, что животные не испытывают боли и страданий, потому что им не хватает сознание.[3][4][5] В 1789 году британский философ и социал-реформист, Джереми Бентам, адресованный в его книге Введение в принципы морали и законодательства вопрос о нашем обращении с животными со следующими часто цитируемыми словами: «Вопрос не в том, могут ли они рассуждать? или в том, могут ли они говорить? а в том, могут ли они страдать?»[6]
Питер Сингер, биоэтик и автор Освобождение животных опубликованная в 1975 году, предположила, что сознание не обязательно является ключевым вопросом: тот факт, что животные имеют меньший мозг или «менее сознательны», чем люди, не означает, что они не способны чувствовать боль. Далее он утверждает, что мы не предполагаем, что новорожденные, люди, страдающие нейродегенеративными заболеваниями головного мозга, или люди с ограниченными возможностями обучения испытывают меньше боли, чем мы.[7]
Бернард Роллин, главный автор двух федеральных законов США, регулирующих обезболивание для животных, пишет, что исследователи до 1980-х годов оставались неуверенными в том, испытывают ли животные боль, а ветеринаров, прошедших подготовку в США до 1989 года, учили просто игнорировать боль животных.[8] Во время общения с учеными и другими ветеринарами Роллина регулярно просили «доказать», что животные находятся в сознании, и предоставить «научно приемлемые» основания для утверждения, что они чувствуют боль.[8]
В 90-е годы продолжались дискуссии о роли философии и науки в понимании познание животных и менталитет.[9] В последующие годы утверждалось, что предложение о том, что некоторые животные (скорее всего, амниот ) иметь хотя бы простые сознательные мысли и чувства[10] и что мнение, что животные чувствуют боль иначе, чем люди, теперь является точкой зрения меньшинства.[3]
Научное исследование
В 20-м и 21-м веках было проведено множество научных исследований боли у нечеловеческих животных.
Млекопитающие
В 2001 году были опубликованы исследования, показывающие, что крысы с артритом самостоятельно выбирают анальгетические опиаты.[12]В 2014 г. ветеринарная Журнал практики мелких животных опубликовал статью о признании боли, которая началась - «Способность испытывать боль универсально присуща всем млекопитающим ...»[13] а в 2015 г. об этом сообщалось в научном журнале Боль, что несколько видов млекопитающих (крыса, мышь, кролик, Кот и лошадь ) принять выражение лица в ответ на вредный раздражитель, которое согласуется с выражением боли у людей.[14]
Птицы
В то же время, что и исследования на крысах с артритом, были опубликованы исследования, показывающие, что птицы при нарушении походки самостоятельно выбирайте диету, содержащую карпрофен, человек обезболивающее.[15] В 2005 году было написано: «Птичья боль, вероятно, аналогична боли, которую испытывают большинство млекопитающих».[16] и в 2014 году «... принято, что птицы воспринимают вредные раздражители и реагируют на них, а птицы чувствуют боль»[17]
Рептилии и амфибии
Были опубликованы ветеринарные статьи о рептилиях.[18][19][20] и амфибии[21][22][23] испытывать боль аналогично людям, и что анальгетики эффективны в этих двух классы позвоночных.
Аргумент по аналогии
В 2012 году американский философ Гэри Варнер изучил литературу по исследованию боли у животных. Его выводы кратко изложены в следующей таблице.[24]
| Аргумент по аналогии[24] | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Свойство | |||||||||
| Рыбы | Амфибии | Рептилии | Птицы | Млекопитающие | |||||
| Имеет ноцицепторы | |||||||||
| Имеет мозг | |||||||||
| Ноцицепторы и мозг связаны | ?[а] / | ?[b] / | ? / | ||||||
| Имеет эндогенный опиоиды | |||||||||
| Анальгетики повлиять на ответы | ?[c] | ?[d] | |||||||
| Реакция на повреждающие раздражители, как у человека | |||||||||
Примечания
Рассуждая по аналогии, Варнер утверждает, что любое животное, проявляющее свойства, перечисленные в таблице, можно сказать, испытывает боль. На этом основании он заключает, что все позвоночные, включая рыб, вероятно, испытывают боль, но беспозвоночные, кроме головоногие моллюски наверное не испытываю боли.[24][29]
Ракообразные
Однако некоторые исследования показывают, что ракообразные действительно проявляют реакцию, соответствующую признакам боли и стресса.[30]
Испытывает боль
Хотя существует множество определений боль, почти все включают два ключевых компонента.
Первый, ноцицепция необходимо.[31] Это способность обнаруживать вредные раздражители, которые вызывают рефлекс реакция, которая быстро перемещает все животное или пораженную часть его тела от источника раздражителя. Понятие ноцицепции не подразумевает какого-либо неблагоприятного, субъективного «ощущения» - это рефлекторное действие. Примером у людей может служить быстрое отдергивание пальца, который коснулся чего-то горячего - отдергивание происходит до того, как на самом деле возникает какое-либо ощущение боли.
Второй компонент - это переживание самой «боли» или страдания - внутренняя эмоциональная интерпретация ноцицептивного опыта. Опять же, у людей это происходит, когда отведенный палец начинает болеть через несколько мгновений после отрыва. Таким образом, боль - это личное эмоциональное переживание. Боль нельзя измерить напрямую у других животных, включая других людей; ответы на предположительно болезненные стимулы можно измерить, но не сам опыт. Чтобы решить эту проблему при оценке способности других видов испытывать боль, используется аргумент по аналогии. Это основано на том принципе, что если животное реагирует на стимул так же, как и мы, у него, вероятно, был аналогичный опыт.
Ноцицепция

Ноцицепция обычно включает в себя передачу сигнала по цепочке нервные волокна от места вредного раздражителя на периферии к спинному и головному мозгу. Этот процесс вызывает рефлекторная дуга реакция, возникающая в спинном мозге и не затрагивающая головной мозг, например вздрагивание или отдергивание конечности. Ноцицепция встречается в той или иной форме у всех основных животных. таксоны.[31] Ноцицепцию можно наблюдать с помощью современных методов визуализации; и физиологический и часто можно обнаружить поведенческую реакцию на ноцицепцию. Однако ноцицептивные реакции у животных-жертв могут быть настолько тонкими, что обученные (люди) наблюдатели не могут их заметить, тогда как естественные хищники могут впоследствии нацеливаться на травмированных особей.[32]
Эмоциональная боль
Иногда различают «физическую боль» и «эмоциональную» или «эмоциональную боль».психологическая боль ". Эмоциональная боль - это боль, испытываемая при отсутствии физической травмы, например боль, которую испытывают люди после потери любимого человека или разрыва отношений. Утверждалось, что только приматы и люди могут чувствовать «эмоциональную боль», потому что они единственные животные, у которых есть неокортекс - часть коры головного мозга, считающаяся «зоной мышления». Однако исследования предоставили доказательства того, что обезьяны, собаки, кошки и птицы могут проявлять признаки эмоциональная боль и отображать поведение, связанное с депрессия во время или после болезненного опыта в частности, отсутствие мотивации, вялость, анорексия и невосприимчивость к другим животным.[7]
Физическая боль
Нервные импульсы болевой реакции могут передаваться в мозг, тем самым регистрируя местоположение, интенсивность, качество и неприятность стимула. Этот субъективный компонент боли включает сознательное осознание как ощущений, так и неприятностей (отталкивающие, отрицательные оказывать воздействие ). Мозговые процессы, лежащие в основе сознательного осознания неприятностей (страданий), изучены недостаточно.
Было опубликовано несколько списков критериев для определения того, испытывают ли животные, кроме человека, боль, например[33][34] Вот некоторые критерии, которые могут указывать на способность других видов, в том числе рыб, чувствовать боль:[34]
- Имеет подходящий нервная система и Рецепторы чувств
- Имеет опиоидные рецепторы и демонстрирует пониженную реакцию на вредные раздражители при введении анальгетики и местные анестетики
- Физиологические изменения в вредные раздражители
- Отображает защитные двигательные реакции, которые могут включать ограниченное использование пораженной области, например хромоту, трение, удержание или аутотомия
- Показывает обучение избеганию
- Показывает компромисс между избеганием вредных стимулов и другими мотивационными требованиями.
- Высоко Познавательная способность и разумность
Адаптивное значение
В адаптивное значение ноцицепции очевидна; Организм, обнаруживший вредный раздражитель, немедленно отвлекает конечность, придаток или все тело от вредного раздражителя и тем самым избегает дальнейшего (потенциального) повреждения. Однако характерной чертой боли (по крайней мере, у млекопитающих) является то, что боль может приводить к гипералгезия (повышенная чувствительность к вредным раздражителям) и аллодиния (повышенная чувствительность к безвредным раздражителям). Когда возникает эта повышенная сенсибилизация, адаптивная ценность менее очевидна. Во-первых, боль, возникающая из-за повышенной сенсибилизации, может быть несоразмерна реальному повреждению тканей. Во-вторых, повышенная сенсибилизация также может стать хронической, сохраняясь далеко за пределами заживления тканей. Это может означать, что причиной беспокойства становится не фактическое повреждение тканей, вызывающее боль, а боль из-за повышенной сенсибилизации. Это означает, что процесс сенсибилизации иногда называют неадаптивный. Часто предполагается, что гипералгезия и аллодиния помогают организмам защитить себя во время заживления, но экспериментальных данных, подтверждающих это, недостаточно.[35][36]
В 2014 году адаптивная ценность сенсибилизации из-за травмы была протестирована с использованием хищных взаимодействий между Прибрежный кальмар (Doryteuthis pealeii) и черный морской окунь (Centropristis striata), которые являются естественными хищниками этого кальмара. Если на раненых кальмаров нападает окунь, они начинают свое защитное поведение раньше (на что указывают большие расстояния тревоги и более длинные дистанции начала полета), чем неповрежденные кальмары. Если анестетик (1% этанол и MgCl2) вводят до травмы, это предотвращает сенсибилизацию и блокирует поведенческий эффект. Авторы утверждают, что это исследование является первым экспериментальным доказательством, подтверждающим аргумент, что ноцицептивная сенсибилизация на самом деле является адаптивной реакцией на травмы.[32]
Был задан вопрос: «Если рыбы не чувствуют боли, то почему у скатов есть чисто защитные шипы на хвосте, доставляющие яд? Предковые хищники скатов - рыбы. И почему многие рыбы обладают защитными шипами плавников, а некоторые также содержат яд, вызывающий боль люди? "[37]
Результаты исследований
Периферическая нервная система
Рецепторы

Примитивные рыбы, такие как миноги (Петромизон маринус) имеют в коже свободные нервные окончания, которые реагируют на тепло и механическое давление. Однако поведенческие реакции, связанные с ноцицепцией, не зарегистрированы, и также трудно определить, действительно ли механорецепторы миноги действительно специфичны для ноцицепции или просто специфичны для давления.[38]
Радужная форель (Oncorhynchus mykiss) имеют полимодальные ноцицепторы на лице и морде, которые реагируют на механическое давление, температуры в опасном диапазоне (> 40 ° C) и 1% уксусная кислота (химический раздражитель).[39][40][41][42] Дальнейшие исследования показали, что ноцицепторы более широко распространены на теле радужной форели, а также трески и карпа. Самые чувствительные участки тела - вокруг глаз, ноздрей, мясистые части хвоста и грудной и спинные плавники.[43]
Радужная форель тоже есть роговица ноцицепторы. Из 27 рецепторов, исследованных в одном исследовании, семь были полимодальными ноцицепторами, а шесть - механотермальными ноцицепторами. Механический и тепловой пороги были ниже, чем у кожных рецепторов, что указывало на большую чувствительность роговицы.[44]
Костные рыбы обладают ноцицепторами, которые по функциям аналогичны таковым у млекопитающих.[45][46]
Нервные волокна
Есть два типа нервных волокон, связанных с болью у рыб. Нервные волокна группы C представляют собой тип сенсорных нервных волокон, в которых отсутствует миелин оболочки и имеют небольшой диаметр, а значит, они имеют низкий скорость нервной проводимости. Страдания, которые люди связывают с ожогами, зубной болью или раздавливанием, вызваны активностью С-волокон. Типичный человек кожный нерв содержит 83% нервных волокон группы С.[47] Волокна А-дельта являются еще одним типом сенсорных нервных волокон, однако они миелинизированы и поэтому передают импульсы быстрее, чем немиелинизированные С-волокна. Волокна A-дельта несут сигналы холода, давления и некоторых болевых сигналов и связаны с острой болью, которая приводит к «отстранению» от вредных раздражителей.
Костяная рыба обладают волокнами группы C и A-дельта, что составляет 38,7% (вместе взятых) волокон хвостовых нервов обыкновенного карпа и 36% тройничного нерва радужной форели. Однако только 5% и 4% из них составляют C-волокна у карпа и радужной форели соответственно.[47][48]
Некоторые виды хрящевые рыбы имеют волокна A-дельта, однако волокна C либо отсутствуют, либо встречаются в очень небольшом количестве.[47][49][50] В Агната (микробы и минога) в основном имеют волокна группы С.[51]
Центральная нервная система
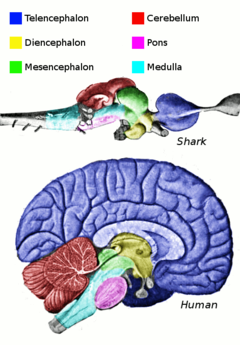
Центральная нервная система (ЦНС) рыб содержит спинной мозг, продолговатый мозг, а мозг, разделен на конечный мозг, промежуточный мозг, средний мозг и мозжечок.
У рыб, как и у других позвоночных, ноцицепция распространяется от периферических нервов вдоль спинномозговых нервов и через спинной мозг передается в нервную систему. таламус. Таламус связан с конечным мозгом множеством соединений через серое вещество. паллий, который, как было продемонстрировано, получает нервные реле для ядовитых и механических раздражителей.[51]
Основные пути, которые передают информацию о боли от периферии к мозгу, - это спиноталамический тракт (тело) и тройничный тракт (голова). Обе они были изучены на бесчеловечных, костистых и двоякожаберных рыбах (тройничный нерв у карпа, спиноталамический тракт у морского робина, Prionotus carolinus ).[52]
Мозг
Если сенсорные реакции у рыб ограничены спинным и задним мозгом, их можно рассматривать как просто рефлексивные. Однако записи спинного мозга, мозжечка, тектум и конечный мозг у форели и золотая рыбка (Карассиус ауратус) покажите, что все они реагируют на вредные раздражители. Это указывает на ноцицептивный путь от периферии к более высоким ЦНС рыб.[53]
Анализ микрочипов из экспрессия гена показывает, что мозг активен на молекулярном уровне в передний мозг, средний мозг и задний мозг карпа и радужной форели. Несколько генов, участвующих в ноцицепции у млекопитающих, например: нейротрофический фактор головного мозга (BDNF) и каннабиноид Рецептор CB1 регулируются в мозге рыб после ноцицептивного события.[54][55]
Соматосенсорные вызванные потенциалы (SEP) - это слабые электрические реакции в ЦНС после стимуляции периферических сенсорных нервов. Это также указывает на то, что существует путь от периферических ноцицепторов к более высоким областям мозга. У золотых рыбок радужная форель, Атлантический лосось (Salmo salar) и Атлантическая треска (Гадус Морхуа), было продемонстрировано, что предположительно безвредная и вредная стимуляция вызывает SEP в различных областях мозга, включая конечный мозг.[56] который может опосредовать координацию информации о боли.[46] Более того, несколько функциональная магнитно-резонансная томография Исследования (фМРТ) с несколькими видами рыб показали, что, когда они страдают от предполагаемой боли, в переднем мозге наблюдается сильная активность, которая очень напоминает ту, что наблюдается у людей, и может быть воспринята как свидетельство ощущения боли у млекопитающих.[57][58]
Следовательно, у рыб, переживающих потенциально болезненное событие, активируются «высшие» области мозга на молекулярном, физиологическом и функциональном уровнях. Снеддон заявил: «Это придает большое значение предположению о том, что рыбы испытывают некоторую форму боли, а не болевые ощущения».[59]
Опиоидная система и эффекты анальгетиков

У костистых рыб есть функциональная опиоидная система, которая включает в себя опиоидные рецепторы, аналогичные рецепторам млекопитающих.[60][61]Все четыре основных опиоидный рецептор типы (дельта, каппа, му, и NOP ) сохраняются у позвоночных, даже у примитивных бесчелюстных рыб (агнатастома).[51]
Такой же анальгетики и анестетики используется для человека и других млекопитающих, часто используется для рыб в ветеринарии. Эти химические вещества действуют на ноцицептивный пути, блокирующие сигналы в мозг, где эмоциональные реакции на сигналы далее обрабатываются определенными частями мозга, обнаруженными у амниот ("высшие позвоночные ").[62][63]
Эффекты морфина

Предварительная обработка морфий (ан обезболивающее у человека и других млекопитающих) имеет дозозависимый антиноцицептивный эффект[64] и смягчает поведенческие и вентиляционные реакции радужной форели на вредные раздражители.
Когда уксусная кислота вводится в губы радужной форели, они проявляют аномальное поведение, такое как покачивание из стороны в сторону и трение губами по стенкам и дну резервуаров, и скорость их вентиляции увеличивается. Инъекции морфина уменьшают как аномальное поведение, связанное с вредными раздражителями, так и увеличение скорости вентиляции.[65] Когда тот же вредный раздражитель применяется к данио (Данио Рерио), они реагируют снижением активности. Как и в случае с радужной форелью, морфин, вводимый перед инъекцией кислоты, ослабляет снижение активности в зависимости от дозы.[46]
Введение уксусной кислоты в губы радужной форели снижает их естественную неофобию (боязнь новизны); это отменяется введением морфина.[43]
У золотых рыбок, которым вводили морфин или физиологический раствор, а затем подвергали воздействию неприятных температур, рыбы, которым вводили физиологический раствор, проявляли защитное поведение, указывающее на беспокойство, настороженность и страх, тогда как те, которым вводили морфин, этого не делали.[66]
Эффекты других анальгетиков
Нейромедиатор, Вещество P и анальгетический опиоид энкефалины и β-эндорфин, которые действуют как эндогенные анальгетики у млекопитающих, присутствуют в рыбе.[67]
Разные анальгетики по-разному действуют на рыбу. В исследовании эффективности трех типов анальгетиков: бупренорфин (опиоид), карпрофен (нестероидный противовоспалительный препарат) и лидокаин (местный анестетик), скорость вентиляции и время возобновления кормления использовались как индикаторы боли. Бупренорфин оказывал ограниченное влияние на реакцию рыб, карпрофен ослаблял эффекты вредной стимуляции вовремя, чтобы возобновить кормление, однако лидокаин снижал все поведенческие показатели.[68]
Трамадол также увеличивает ноцицептивный порог у рыб, что является дополнительным доказательством антиноцицептивной опиоидной системы у рыб.[43][69]
Эффекты налоксона
Налоксон является μ-опиоидный рецептор антагонист, который у млекопитающих сводит на нет обезболивающие эффекты опиоидов. И взрослые, и пятидневные личинки рыбок данио демонстрируют поведенческие реакции, указывающие на боль в ответ на введенную или разбавленную уксусную кислоту. Антиноцицептивные свойства морфина или бупренорфина меняются, если у взрослых[46] или личинки,[70] обрабатываются совместно с налоксоном. И налоксон, и пролил-лейцил-глицинамид (еще один антагонист опиатов у млекопитающих) уменьшал обезболивающее действие морфина при поражении электрическим током, полученном золотой рыбкой, что указывает на то, что они могут действовать как антагонист опиата у рыб.[71][72]
Физиологические изменения
Физиологические изменения рыб в ответ на вредные раздражители включают повышение скорости вентиляции.[54][64][65][73] и кортизол уровни.[69]
Защитные ответы



Исследования показывают, что рыбы проявляют защитные поведенческие реакции на предположительно болезненные стимулы.
Когда уксусная кислота или пчелиный яд вводятся в губы радужной форели, они демонстрируют аномальное качание из стороны в сторону. грудные плавники, потрите губами бока и дно резервуаров[74] и увеличить скорость их вентиляции.[73] Когда уксусная кислота вводится в губы рыбок данио, они реагируют снижением своей активности. Величина этой поведенческой реакции зависит от концентрации уксусной кислоты.[46]
Поведенческие реакции на вредный раздражитель различаются у разных видов рыб. Ядовито стимулированный карп (Cyprinus carpio) демонстрируют аномальное раскачивание и трутся губами о стенки резервуара, но не изменяют другое поведение или скорость вентиляции. В отличие, данио (Данио Рерио) уменьшают частоту плавания и увеличивают интенсивность вентиляции, но не проявляют аномального поведения. Радужная форель, как и данио, сокращает частоту плавания и увеличивает скорость вентиляции.[75] Нильская тилапия (Oreochromis niloticus), в ответ на хвостовой плавник клипсы, увеличивают свою плавательную активность и проводят больше времени в светлой зоне своего аквариума.[76]
Начиная с этой первоначальной работы, Снеддон и ее коллеги показали, что радужная форель, карп и рыбка данио, испытывающие ядовитую стимуляцию, демонстрируют быстрые изменения в физиологии и поведении, которые сохраняются до 6 часов и, следовательно, не являются простыми рефлексами.[52]
Пятидневные личинки рыбок данио демонстрируют зависимое от концентрации увеличение двигательной активности в ответ на различные концентрации разбавленной уксусной кислоты. Это повышение двигательной активности сопровождается увеличением мРНК cox-2, демонстрируя активацию ноцицептивных путей.[70]
Рыбы по-разному реагируют на разные вредные раздражители, даже если они кажутся похожими. Это указывает на то, что реакция является гибкой, а не просто ноцицептивным рефлексом. Атлантическая треска вводится в губу уксусной кислотой, капсаицин или прокалывание губы коммерческим рыболовным крючком, показали разные реакции на эти три типа вредной стимуляции. Треска, обработанная уксусной кислотой и капсаицином, показала более частое зависание вблизи дна аквариума и сокращение использования укрытий. Однако у пойманной трески наблюдались лишь краткие эпизоды покачивания головой.[73]
Обучение избеганию
Ранние эксперименты показали, что рыбы учатся реагировать на якобы вредные раздражители. Например, жаба (Batrachoididae ) хрюкают при поражении электрическим током, но после многократных ударов они хрюкают просто при виде электрода.[77][78] Более поздние исследования показывают, что и золотая рыбка, и форель учатся избегать мест, в которых они получают электрический ток. Кроме того, это обучение избеганию является гибким и связано с интенсивностью стимула.[69][79][80]
Компромиссы в мотивации

Болезненный опыт может изменить мотивацию нормальных поведенческих реакций.
В исследовании 2007 года золотых рыбок учили кормиться в аквариуме, где впоследствии их ударили током. Количество попыток кормления и время, проведенное в зоне кормления / шока, уменьшались с увеличением интенсивности шока, а с увеличением отсутствия пищи количество и продолжительность попыток кормления увеличивались, как и реакции побега при входе в эту зону. Исследователи предположили, что золотые рыбки идут на компромисс между мотивацией кормления и мотивацией избегать острого вредного раздражителя.[80]
Радужная форель, естественно, избегает новизны (т.е. неофобский ). Виктория Брейтуэйт описывает исследование, в котором ярко окрашенный конструктор Лего кирпич кладется в емкость с радужной форелью. Форель, которой вводили в губу небольшое количество физиологического раствора, сильно избегала кирпичей Lego, однако форель, которым вводили уксусную кислоту, проводила значительно больше времени рядом с блоком Lego. Когда исследование было повторено, но рыбам также давали морфин, реакция избегания возвращалась у тех рыб, которым вводили уксусную кислоту, и ее нельзя было отличить от ответов рыб, которым вводили физиологический раствор.[43][81]
Чтобы изучить возможность компромисса между реакцией на вредный раздражитель и хищником, исследователи представили радужной форели конкурирующий стимул - сигнал хищника. Ядовито стимулированные рыбы перестают проявлять реакцию против хищников, что указывает на то, что боль становится их основной мотивацией. В том же исследовании изучался потенциальный компромисс между реакцией на вредный стимул и социальным статусом. Реакция на ядовито обработанную форель варьировалась в зависимости от того, с какой рыбой они были знакомы. Исследователи предположили, что результаты мотивационных изменений и компромиссов служат доказательством центральной обработки боли, а не просто демонстрацией ноцицептивного рефлекса.[81][82]
Плата за обезболивание
Рыбкам данио предоставлен доступ в бесплодную, ярко освещенную камеру или обогащенный Камерные предпочитают обогащенную площадь. Когда этим рыбам вводят уксусную кислоту или физиологический раствор в качестве контроля, они все равно выбирают ту же обогащенную камеру. Однако, если анальгетик растворяется в бесплодной, менее предпочтительной камере, рыбки данио, которым вводят ядовитую кислоту, теряют свое предпочтение и проводят большую часть своего времени в ранее менее благоприятной бесплодной камере. Это предполагает компромисс в мотивации, и, кроме того, они готовы заплатить определенную цену, чтобы войти в менее предпочтительную среду для получения облегчения боли.[33]
Познавательные способности и разум
Способность рыб к обучению, продемонстрированная в ряде исследований, указывает на сложность когнитивные процессы которые более сложные, чем простые ассоциативное обучение. Примеры включают способность узнавать социальных товарищей, избегать (в течение нескольких месяцев или лет) мест, где они встретили хищника или были пойманы на крючок, и формирование ментальные карты.[67]
Утверждалось, что, хотя высокая когнитивная способность может указывать на большую вероятность переживания боли, она также дает этим животным большую способность справляться с этим, оставляя животных с более низкими когнитивными способностями более серьезной проблемой для преодоления боли.[83]
Критерии восприятия боли
Ученые также предположили, что в сочетании с аргументацией по аналогии критерии физиологии или поведенческих реакций могут использоваться для оценки возможности восприятия боли животными, не являющимися людьми. Ниже приводится таблица критериев, предложенных Снеддоном. и другие.[33]
| Критерии приема боли у рыб | ||||
|---|---|---|---|---|
| Критерии | ||||
| Рыба без челюсти
| Хрящевые рыбы
| Костяная рыба
| Рыба с лопастными плавниками
| |
| Имеет ноцицепторы | ? | ? | ? | |
| Пути к центральной нервной системе | ? | ? | ? | |
| Центральная обработка в мозгу | ? | ? | ? | |
| Рецепторы анальгетических препаратов | ? | ? | ? | |
| Физиологические реакции | ? | ? | ? | |
| Уход от вредных раздражителей | ? | ? | ? | |
| Поведенческие изменения от нормы | ? | ? | ? | |
| Защитное поведение | ? | ? | ? | |
| Ответы на обезболивающие уменьшаются | ? | ? | ? | |
| Самостоятельное обезболивание | ? | ? | ? | |
| Ответы с более высоким приоритетом по сравнению с другими стимулами | ? | ? | ? | |
| Плата за доступ к анальгезии | ? | ? | ? | |
| Измененные поведенческие предпочтения / предпочтения | ? | ? | ? | |
| Облегчение обучения | ? | ? | ? | |
| Трение, хромота или охрана | ? | ? | ? | |
| Платить за то, чтобы избежать стимулов | ? | ? | ? | |
| Компромиссы с другими требованиями | ? | ? | ? | |



В таблице, ![]() указывает на положительные доказательства и? означает, что он не был протестирован или доказательств недостаточно.
указывает на положительные доказательства и? означает, что он не был протестирован или доказательств недостаточно.
Социальные последствия

Учитывая, что некоторые интерпретировали существующую научную информацию, предполагающую, что рыба может чувствовать боль,[84] было высказано предположение, что принципы предосторожности следует применять к коммерческому рыболовству, что может иметь множество последствий.[84]
И ученые, и защитники животных выразили обеспокоенность по поводу возможных страданий (боли и страха) рыб, вызванных рыбалка.[85][86][87]
Другие социальные последствия испытываемой рыбой боли включают острое и хроническое воздействие загрязнителей, коммерческое и спортивное рыболовство (например, травмы во время траления, мечение / обрезание плавников во время оценки запасов, повреждение тканей, физическое истощение и серьезный дефицит кислорода во время отлова, боль и стресс во время убоя , использование живой наживки), аквакультура (например, мечение / стрижка плавников, высокая плотность посадки, приводящая к повышенной агрессии, лишение пищи для лечения болезней или перед промыслом, удаление из воды для обычного разведения, боли во время убоя), декоративная рыба (например, отлов сублетальное отравление, постоянные неблагоприятные физические состояния из-за селекционного разведения), научные исследования (например, генетическая модификация) могут иметь пагубные последствия для благосостояния, преднамеренно навязанные неблагоприятные физические, физиологические и поведенческие состояния, электроловушка, мечение, обрезка плавников или иная маркировка рыба, обращение с которой может привести к травмам.[38][88]
Browman et al.[89] предполагают, что если нормативно-правовая среда будет продолжать двигаться по текущей траектории (добавление большего количества таксонов водных животных к уже регулируемым), деятельность в некоторых секторах может быть серьезно ограничена или даже запрещена. Они также утверждают, что распространение правовой защиты на водных животных является выбором общества, но подчеркивают, что этот выбор не следует приписывать сильной поддержке со стороны исследования, которое еще не существует и может никогда не существовать, и последствиями принятия такого решения. необходимо тщательно взвесить.
Законодательство
В Великобритании закон о защите животных во время научных исследований, «Закон о животных (научных процедурах) 1986 года», защищает рыбу с того момента, как они становятся способными к самостоятельному питанию.[90] Законодательством, защищающим животных в большинстве других обстоятельств в Великобритании, является «Закон о благополучии животных 2006 года», в котором говорится, что в Законе «животное» означает позвоночное животное, отличное от человека »,[91] явно включая рыбу.
В США законом, защищающим животных во время научных исследований, является «Закон о защите животных».[92] Это исключает защиту «хладнокровных» животных, в том числе рыб.[93]
В норвежском законе о правах животных 1974 года говорится, что это касается млекопитающих, птиц, лягушек, саламандр, рептилий, рыб и ракообразных.[94]
Статья Ховарда Броумана и его коллег за 2018 год представляет собой обзор того, что означают различные точки зрения в отношении боли и благополучия рыб в контексте аквакультуры, коммерческого рыболовства, любительского рыболовства и исследований.[89]
Полемика
Нервная система
Рецепторы и нервные волокна
Утверждалось, что рыбы не чувствуют боли, потому что у них нет достаточной плотности соответствующих нервных волокон. Типичный кожный нерв человека содержит 83% группы C нервные волокна,[47] однако те же нервы у людей с врожденная нечувствительность к боли имеют только 24–28% волокон С-типа.[47] Основываясь на этом, Джеймс Роуз из Университета Вайоминга утверждал, что отсутствие волокон С-типа у хрящевых акул и скатов указывает на то, что передача сигналов, ведущих к восприятию боли, вероятно, невозможна, а низкие числа для костистых рыб (например, 5% для карпа и форели) указывают на то, что это также маловероятно для этих рыб.[47] Роуз приходит к выводу, что существует мало доказательств того, что акулы и скаты обладают ноцицепторами, необходимыми для инициирования обнаружения боли в мозгу, и что, хотя костлявые рыбы способны бессознательно учиться избегать вредных раздражителей, они не намного чаще испытывают сознательную боль, чем акулы.[нужна цитата ]
Волокна A-дельта-типа, которые, как считается, вызывают реакцию избегания, обычны у костистых рыб, хотя они не были обнаружены у акул или скатов.[47]
Роуз приходит к выводу, что рыбы хорошо выжили в эволюционном смысле без полного спектра болевых ощущений, типичных для людей или других млекопитающих.[47]
Мозг
В 2002 году Роуз опубликовал обзоры, в которых утверждалось, что рыбы не могут чувствовать боль, потому что у них нет неокортекс в мозгу.[95][96] Этот аргумент также исключает восприятие боли у большинства млекопитающих, птиц и рептилий.[57] Однако в 2003 году исследовательская группа под руководством Линн Снеддон пришла к выводу, что мозг радужной форели активирует нейроны так же, как человеческий мозг, когда испытывает боль.[97][98] Роуз раскритиковал исследование, заявив, что оно было некорректным, главным образом потому, что оно не предоставило доказательств того, что рыбы обладают «сознательной осведомленностью, особенно такой же, как у нас».[99]
Роуз, а совсем недавно Брайан Ки[2][100] из Университета Квинсленда, утверждают, что, поскольку мозг рыб очень отличается от человеческого, рыбы, вероятно, не обладают сознанием, как люди, и хотя рыбы могут реагировать аналогично тому, как люди реагируют на боль, реакции в У рыбы есть и другие причины. По словам Роуз, исследования, показывающие, что рыба может чувствовать боль, путали ноцицепцию с ощущением боли. «Боль основывается на осознании. Ключевым вопросом является различие между ноцицепцией и болью. Человек, находящийся под наркозом в операционной, все равно будет физически реагировать на внешний раздражитель, но он или она не будет чувствовать боли».[101] Согласно Роуз и Кей, литература, касающаяся вопроса о сознании рыб, склонна к антропоморфизму, и необходимо проявлять осторожность, чтобы избежать ошибочного приписывания рыбам человеческих способностей.[102] Снеддон предполагает, что вполне возможно, что виды с другим эволюционным путем могли развить разные нейронные системы для выполнения тех же функций (т. Е. конвергентная эволюция ), как показали исследования мозга птиц.[103] Ки соглашается с тем, что феноменальное сознание может иметь место у млекопитающих и птиц, но не у рыб.[2] Бихевиорист Темпл Грандин утверждает, что рыбы все еще могут иметь сознание без неокортекса, потому что «разные виды могут использовать разные структуры и системы мозга для выполнения одних и тех же функций».[98] Снеддон предполагает, что предположение, что функция внезапно возникает без примитивной формы, противоречит законам эволюции.[нужна цитата ]
Другие исследователи также считают, что сознание животных не требует неокортекса, но может возникать из гомологичный подкорковый сети мозга.[11] Было высказано предположение, что мозговой ствол цепи могут вызвать боль. Это включает исследования с анэнцефальный дети, которые, несмотря на отсутствие значительных участков коры головного мозга, выражают эмоции. Имеются также данные исследований активации, показывающие, что у нормальных людей ощущения, опосредованные стволом мозга, и реакции отмены плода на вредную стимуляцию, но до развития коры головного мозга.[104]
В статьях, опубликованных в 2017 и 2018 годах, Майкл Вудрафф[105][106] обобщил значительное количество исследовательских статей, которые, в отличие от выводов Роуза и Ки, решительно поддерживают гипотезу о том, что нейроанатомическая организация паллиума рыб и его связи с субпаллиальными структурами, особенно с прегломерулярным ядром и тектумом, являются сложными. достаточно, чтобы быть аналогом схем коры и таламуса, которые, как предполагается, лежат в основе чувствительности у млекопитающих. Он добавил к этим анатомическим наблюдениям нейрофизиологические и поведенческие данные, которые также подтверждают гипотезу о том, что паллий является важной частью иерархической сети, предложенной Файнбергом и Маллаттом, лежащей в основе сознания у рыб.[107]
Защитные ответы
Первоначальная работа Снеддон и ее сотрудников характеризовала поведенческие реакции у радужной форели, карпа и рыбок данио.[52] Однако, когда эти эксперименты были повторены Ньюби и Стивенсом без анестезии, покачивания и трения не наблюдалось, что позволяет предположить, что некоторые из предполагаемых болевых реакций, наблюдаемых Снеддоном и его сотрудниками, вероятно, были вызваны выздоровлением рыбы после наркоза.[108][109][110]
Некоторые исследователи спорят об определении боли, используемом в поведенческих исследованиях, поскольку записанные наблюдения были противоречивыми, не подтвержденными и не повторяемыми другими исследователями.[39][47] В 2012 году Роуз утверждал, что рыбы возобновляют «нормальное питание и активность сразу или вскоре после операции».[47]
Нордгрин сказал, что различия в поведении, которые они обнаружили в ответ на неудобную температуру, показали, что рыбы чувствуют как рефлексивную, так и когнитивную боль.[111] «Эксперимент показывает, что рыбы не только реагируют на болезненные раздражители рефлексами, но и меняют свое поведение после происшествия», - сказал Нордгрин. «Вместе с тем, что мы знаем из экспериментов, проведенных другими группами, это указывает на то, что рыбы сознательно воспринимают тестовую ситуацию как болезненную и переключаются на поведение, указывающее на то, что они прошли через неприятный опыт».[111] В 2012 году Роуз и другие проанализировали это и дальнейшие исследования, которые пришли к выводу, что у рыб была обнаружена боль. Они пришли к выводу, что результаты такого исследования связаны с плохим дизайном и неправильной интерпретацией, и что исследователи не смогли отличить бессознательное обнаружение повреждающих стимулов (ноцицепцию) от осознанной боли.[47]
Смотрите также
Рекомендации
- ^ Эбботт, Ф.В., Франклин, К.Б.Дж. и Вестбрук Р.Ф. (1995). «Формалиновый тест: оценка свойств первой и второй фаз болевого ответа у крыс». Боль. 60 (1): 91–102. Дои:10.1016 / 0304-3959 (94) 00095-В. PMID 7715946. S2CID 35448280.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c Ки, Б. (2015). «Рыбы не чувствуют боли и ее значение для понимания феноменального сознания». Биология и философия. 30 (2): 149–165. Дои:10.1007 / s10539-014-9469-4. ЧВК 4356734. PMID 25798021.
- ^ а б Карбоне, Л. (2004). Чего хотят животные: опыт и пропаганда политики защиты лабораторных животных. Издательство Оксфордского университета. п. 149. ISBN 9780195161960.
- ^ Раднер, Д. и Раднер, М. (1989). Сознание животных. Книги Прометея: Буффало.
- ^ Харрисон, П. (1992). «Декарт о животных». Философский квартал. 42 (167): 219–227. Дои:10.2307/2220217. JSTOR 2220217.
- ^ "Бентам, Дж. (1879). Введение в принципы морали и законодательства. Кларендон Пресс.
- ^ а б Снеддон, Л.У. "Могут ли животные чувствовать боль?". Добро пожаловать в доверие. Архивировано из оригинал 13 апреля 2012 г.. Получено 24 сентября, 2015.
- ^ а б Роллин, Б. (1989). Незаметный крик: сознание животных, боль животных и наука. Oxford University Press, стр. Xii, 117-118, процитировано в Carbone 2004, стр. 150.
- ^ Аллен, К. (1998). «Оценка познания животных: этологические и философские перспективы». Журнал зоотехники. 76 (1): 42–47. Дои:10.2527 / 1998.76142x. PMID 9464883.
- ^ Гриффин, Д. И Спек, Г. (2004). «Новое свидетельство животного сознания». Познание животных. 7 (1): 5–18. Дои:10.1007 / s10071-003-0203-х. PMID 14658059. S2CID 8650837.
- ^ а б Лоу, П. (7 июля 2012 г.). Яак Панксепп; Дайана Рейсс; Дэвид Эдельман; Бруно Ван Суиндерен; Филип Лоу; Кристоф Кох (ред.). «Кембриджская декларация о сознании» (PDF). Кембриджский университет.
- ^ Кольпаерт, ФК, Тарайр, Дж. П., Аллиага, М., Слот. Л.А.Б., Аттал Н. и Кук В. (2001). «Самостоятельное введение опиатов как мера хронической ноцицептивной боли у крыс с артритом». Боль. 91 (1–2): 33–45. Дои:10.1016 / s0304-3959 (00) 00413-9. PMID 11240076. S2CID 24858615.CS1 maint: несколько имен: список авторов (связь)
- ^ Мэтьюз, К., Кронен, П.В., Ласселлес, Д., Нолан, А., Робертсон, С., Стигалл, П.В., Райт, Б. и Ямасита, К. (2014). «Рекомендации по распознаванию, оценке и лечению боли». Журнал практики мелких животных. 55 (6): E10 – E68. Дои:10.1111 / jsap.12200. PMID 24841489.CS1 maint: несколько имен: список авторов (связь)
- ^ Чемберс, К. и Могил, Дж. (2015). «Онтогенез и филогения лицевого выражения боли». Боль. 156 (5): 798–799. Дои:10.1097 / j.pain.0000000000000133. PMID 25887392. S2CID 2060896.CS1 maint: несколько имен: список авторов (связь)
- ^ Дэнбери, Т.К., Уикс, К.А., Чемберс, Дж. П., Уотерман-Пирсон, А.Э. и Кестин, С.С. (2000). «Самовыбор обезболивающего препарата карпрофен хромыми цыплятами-бройлерами». Ветеринарная карта. 146 (11): 307–311. Дои:10.1136 / vr.146.11.307. PMID 10766114. S2CID 35062797.CS1 maint: несколько имен: список авторов (связь)
- ^ Мачин, К. (2005). «Птичий обезболивающий». Семинары по медицине птиц и экзотических животных. 14 (4): 236–242. Дои:10.1053 / j.saep.2005.09.004.
- ^ Пол-Мерфи, Дж. И Хокинс, М. (2014). «Глава 26 - Особенности птиц: распознавание боли у домашних птиц». В Gaynor, J.S. & Muir III, W. W. (ред.). Справочник по ветеринарному лечению боли. Elsevier Health Sciences.
- ^ Мосли, К.А. (2005). «Анестезия и анальгезия у рептилий». Семинары по медицине птиц и экзотических животных. 14 (4): 243–262. Дои:10.1053 / j.saep.2005.09.005.
- ^ Мосли, К. (2011). «Боль и ноцицепция у рептилий». Ветеринарные клиники Северной Америки: практика экзотических животных. 14 (1): 45–60. Дои:10.1016 / j.cvex.2010.09.009. PMID 21074702.
- ^ Сладкий, К. И Манс, К. (2012). «Клиническая анальгезия у рептилий». Журнал экзотической медицины для домашних животных. 21 (2): 158–167. Дои:10.1053 / j.jepm.2012.02.012.
- ^ Мачин, К. (1999). «Амфибийная боль и обезболивание». Журнал медицины зоопарков и дикой природы. 30 (1): 2–10. JSTOR 20095815. PMID 10367638.
- ^ Мачин, К. (2001). «Обезболивание рыб, амфибий и рептилий». Ветеринарные клиники Северной Америки. Экзотические животные. 4 (1): 19–33. Дои:10.1016 / S1094-9194 (17) 30048-8. PMID 11217460.
- ^ Стивенс, C.W. (2011). «Обезболивание у амфибий: доклинические исследования и клиническое применение». Ветеринарные клиники Северной Америки: практика экзотических животных. 14 (1): 33–44. Дои:10.1016 / j.cvex.2010.09.007. ЧВК 3056481. PMID 21074701.
- ^ а б c Варнер, Гэри Э. (2012) "Какие животные разумны?" Глава 5 в: Личность, этика и познание животных: расположение животных в двухуровневом утилитаризме Зайца, Oxford University Press. ISBN 9780199758784. Дои:10.1093 / acprof: oso / 9780199758784.001.0001 Таблица в статье основана на таблице 5.2, стр. 113.
- ^ Генетт С.А., Жиру М.С. и Вашон, П. (2013). «Восприятие боли и анестезия у исследовательских лягушек». Экспериментальные животные. 62 (2): 87–92. Дои:10.1538 / expim.62.87. PMID 23615302.CS1 maint: несколько имен: список авторов (связь)
- ^ Мосли, К. (2006). «Боль, ноцицепция и обезболивание у рептилий: когда ваша змея уходит!'" (PDF). Североамериканская ветеринарная конференция. 20: 1652–1653.
- ^ Кобл, Д.Дж., Тейлор, Д.К. и Мук Д. (2011). «Обезболивающие эффекты мелоксикама, сульфата морфина, флуниксина меглумина и ксилазина гидрохлорида у африканских когтистых лягушек (Xenopus laevis)». Журнал Американской ассоциации лабораторных исследований животных. 50 (3): 355–60. ЧВК 3103286. PMID 21640031.CS1 maint: несколько имен: список авторов (связь)
- ^ Бейкер Б.Б., Сладкий К.К. и Джонсон, С. (2011). «Оценка обезболивающих эффектов перорального и подкожного введения трамадола у красноухих черепах-слайдеров». Журнал Американской ветеринарной медицинской ассоциации. 238 (2): 220–227. Дои:10.2460 / javma.238.2.220. ЧВК 3158493. PMID 21235376.CS1 maint: несколько имен: список авторов (связь)
- ^ Эндрюс, Кристин (2014) Животный разум: введение в философию познания животных раздел 3.6.2, Routledge. ISBN 9781317676751.
- ^ «Каков самый гуманный способ умерщвления ракообразных для употребления в пищу человеком? - База знаний RSPCA». Получено 2020-09-20.
- ^ а б Снеддон, Л.У. (2004). «Эволюция ноцицепции у позвоночных: сравнительный анализ низших позвоночных». Обзоры исследований мозга. 46 (2): 123–130. Дои:10.1016 / j.brainresrev.2004.07.007. PMID 15464201. S2CID 16056461.
- ^ а б Крук, Р.Дж., Диксон, К., Хэнлон, Р.Т. и Уолтерс, Э. (2014). «Ноцицептивная сенсибилизация снижает риск нападения хищников». Текущая биология. 24 (10): 1121–1125. Дои:10.1016 / j.cub.2014.03.043. PMID 24814149.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c Снеддон, Л.У., Элвуд, Р.В., Адамо, С.А. и Лич, М.С. (2014). «Определение и оценка боли животных». Поведение животных. 97: 201–212. Дои:10.1016 / j.anbehav.2014.09.007. S2CID 53194458.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Элвуд, Р.В., Барр, С., Паттерсон, Л. (2009). «Боль и стресс у ракообразных?». Прикладная наука о поведении животных. 118 (3): 128–136. Дои:10.1016 / j.applanim.2009.02.018.CS1 maint: несколько имен: список авторов (связь)
- ^ Прайс, T.J. И Дуссор, Г. (2014). «Эволюция: преимущество« неадаптивной »пластичности боли». Текущая биология. 24 (10): R384 – R386. Дои:10.1016 / j.cub.2014.04.011. ЧВК 4295114. PMID 24845663.
- ^ «Неадаптивная боль». Оксфордский справочник. Получено 16 мая, 2016.
- ^ Сафина, К. (2016). «Рыбная боль: болезненная тема». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 41.
- ^ а б Хантингфорд, Ф.А., Адамс, К., Брейтуэйт, В.А., Кадри, С., Поттинджер, Т.Г., Сандё, П. и Тернбулл, Дж. Ф. (2006). «Обзорный доклад: Актуальные проблемы благополучия рыб» (PDF). Журнал биологии рыб. 68 (2): 332–372. Дои:10.1111 / j.0022-1112.2006.001046.x.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Снеддон, Л.У., Брейтуэйт, В.А. и Джентл, М.Дж. (2003). «Есть ли у рыб ноцицепторы: свидетельства эволюции сенсорной системы позвоночных». Труды Королевского общества B: биологические науки. 270 (1520): 1115–1121. Дои:10.1098 / rspb.2003.2349. ЧВК 1691351. PMID 12816648.CS1 maint: несколько имен: список авторов (связь)
- ^ Снеддон Л.У. (2003). «Соматосенсорная иннервация тройничного нерва головы радужной форели с особым упором на ноцицепцию». Исследование мозга. 972 (1–2): 44–52. Дои:10.1016 / с0006-8993 (03) 02483-1. PMID 12711077. S2CID 14616224.
- ^ Эшли П.Дж., Снеддон Л.У. и McCrohan C.R. (2007). «Ноцицепция у рыб: стимульно-ответные свойства рецепторов на голове форели Oncorhynchus mykiss». Исследование мозга. 1166: 47–54. Дои:10.1016 / j.brainres.2007.07.011. PMID 17673186. S2CID 15837167.CS1 maint: несколько имен: список авторов (связь)
- ^ Меттам Дж. Дж., МакКрохан С. Р. и Снеддон Л. У. (2011). «Характеристика хемосенсорных рецепторов тройничного нерва у радужной форели (Oncorhynchus mykiss): реакция на раздражители и углекислый газ». Журнал экспериментальной биологии. 215 (4): 685–693. Дои:10.1242 / jeb.060350. PMID 22279076.
- ^ а б c d Брейтуэйт, В. (2010). Чувствуют ли рыбы боль?. Издательство Оксфордского университета.
- ^ Эшли П.Дж., Снеддон Л.У. и McCrohan, C.R. (2006). «Свойства рецепторов роговицы костистых рыб». Письма о неврологии. 410 (3): 165–168. Дои:10.1016 / j.neulet.2006.08.047. PMID 17101221. S2CID 14375428.CS1 maint: несколько имен: список авторов (связь)
- ^ Снеддон, Л.У. (2015). «Боль у водных животных». Журнал экспериментальной биологии. 218 (7): 967–976. Дои:10.1242 / jeb.088823. PMID 25833131.
- ^ а б c d е Коррейя, А.Д., Кунья, С.Р., Шольце, М., Стивенс, Е.Д. (2011). «Новая поведенческая модель ноцицепции рыб для тестирования анальгетиков». Фармацевтические препараты. 4 (4): 665–680. Дои:10.3390 / ph4040665. ЧВК 4055884.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c d е ж грамм час я j k Rose, J.D .; Arlinghaus, R .; Cooke, S.J .; Diggles, B.K .; Sawynok, W .; Стивенс, E.D .; Wynne, C.D.L. (2012). "Может ли рыба действительно чувствовать боль?" (PDF). Рыба и рыболовство. 15 (1): 97–133. Дои:10.1111 / faf.12010.
- ^ Снеддон, Л.У. (2002). «Анатомо-электрофизиологический анализ тройничного нерва радужной форели Onchorynchus mykiss». Письма о неврологии. 319 (3): 167–171. Дои:10.1016 / S0304-3940 (01) 02584-8. PMID 11834319. S2CID 14807046.
- ^ Сноу П.Дж., Плендерлейт М.Б. и Райт Л.Л. (1993). «Количественное исследование популяций первичных сенсорных нейронов трех видов пластиножаберных рыб». Журнал сравнительной неврологии. 334 (1): 97–103. Дои:10.1002 / cne.903340108. PMID 8408762. S2CID 32762031.CS1 maint: несколько имен: список авторов (связь)
- ^ Брейтуэйт, В. И Булкотт П. (2007). «Восприятие боли, отвращение и страх у рыб» (PDF). Болезни водных организмов. 75 (2): 131–138. Дои:10.3354 / dao075131. PMID 17578252.
- ^ а б c Вебер, Э. (2011). «Рыбная анальгезия: боль, стресс, отвращение к страху или ноцицепция?». Ветеринарные клиники Северной Америки: практика экзотических животных. 14 (1): 21–32. Дои:10.1016 / j.cvex.2010.09.002. PMID 21074700.
- ^ а б c Снеддон, Л.У. (2009). «Восприятие боли у рыб: индикаторы и конечные точки». Журнал ILAR. 50 (4): 338–342. Дои:10.1093 / ilar.50.4.338. PMID 19949250.
- ^ Данлоп Р. и Лэминг П. (2005). «Механорецептивные и ноцицептивные реакции в центральной нервной системе золотой рыбки (Carassius auratus) и форели (Oncorhynchus mykiss)». Журнал боли. 6 (9): 561–568. Дои:10.1016 / j.jpain.2005.02.010. PMID 16139775.
- ^ а б Рейли, С.С., Куинн, Дж. П., Коссинс, А. и Снеддон Л.У. (2008). «Новые гены-кандидаты, идентифицированные в мозге во время ноцицепции у карпа (Cyprinus carpio) и радужной форели (Oncorhynchus mykiss)». Письма о неврологии. 437 (2): 135–138. Дои:10.1016 / j.neulet.2008.03.075. PMID 18440145. S2CID 18763423.CS1 maint: несколько имен: список авторов (связь)
- ^ Познер, Л.П. (2009). «Боль и страдания у рыб: обзор доказательств». Журнал ILAR. 50 (4): 327–328. Дои:10.1093 / ilar.50.4.327. PMID 19949248.
- ^ Людвигсен, С., Стенклев, Н.К., Йонсен, Х.К., Лаукли, Э., Матре, Д. и Аас-Хансен, Ø. (2014). «Вызванные потенциалы у атлантической трески после якобы безобидной и якобы вредной электростимуляции: минимально инвазивный подход». Физиология и биохимия рыб. 40 (1): 173–181. Дои:10.1007 / s10695-013-9834-2. ЧВК 3901938. PMID 23896862.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Браун, К. (2015). «Рыбный интеллект, разум и этика». Познание животных. 18 (1): 1–17. Дои:10.1007 / s10071-014-0761-0. PMID 24942105. S2CID 207050888.
- ^ Sneddon, L.U .; Лич, М.С. (2016). «Антропоморфное отрицание боли рыб». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 28.
- ^ Снеддон, Л.У. (2011). «Восприятие боли у рыб и последствия употребления рыбы». Журнал исследований сознания. 18 (9): 209–229.
- ^ Буатти, М. И Пастернак, Г. (1981). «Множественные опиатные рецепторы: филогенетические различия». Исследование мозга. 218 (1–2): 400–405. Дои:10.1016/0006-8993(81)91319-6. PMID 6268247. S2CID 6870252.
- ^ Веласко Э.М.Ф., Закон П.Ю. и Родригес, Р. (2009). «Опиоидный рецептор Mu из рыбок данио проявляет функциональные характеристики, как и опиоидный рецептор Mu млекопитающих». Данио. 6 (3): 259–268. Дои:10.1089 / zeb.2009.0594. PMID 19761379.CS1 maint: несколько имен: список авторов (связь)
- ^ Виньуэла-Фернандес И., Джонс Э., Валлийский Э.М., Флитвуд-Уокер С.М. (сентябрь 2007 г.). «Механизмы боли и их значение для лечения боли у сельскохозяйственных и домашних животных». Вет. J. 174 (2): 227–39. Дои:10.1016 / j.tvjl.2007.02.002. PMID 17553712.
- ^ Снеддон, Л.У. (2012). «Клиническая анестезия и анальгезия у рыб». Журнал экзотической медицины для домашних животных. 21: 32–43. Дои:10.1053 / j.jepm.2011.11.009.
- ^ а б Джонс, С.Г., Камунде, К., Лемке, К., Стивенс, Э.Д. (2012). «Отношение доза-ответ для антиноцицептивного эффекта морфина у рыб, радужной форели». Журнал ветеринарной фармакологической терапии. 35 (6): 563–570. Дои:10.1111 / j.1365-2885.2011.01363.x. PMID 22229842.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Снеддон, Л.У. (2003). «Доказательства боли у рыб: использование морфина в качестве анальгетика». Прикладная наука о поведении животных. 83 (2): 153–162. Дои:10.1016 / s0168-1591 (03) 00113-8.
- ^ Нордгрин, Дж., Джозеф, П., Гарнер, Дж. П., Янчак, А. М., Ранхейм, Б., Мюир, В. М. и Хорсберг, Т. (2009). «Термоноцицепция у рыб: влияние двух различных доз морфина на тепловой порог и поведение после тестирования у золотой рыбки (Carassius auratus)». Прикладная наука о поведении животных. 119 (1–2): 101–107. Дои:10.1016 / j.applanim.2009.03.015.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Веник, Д. (2007). «Когнитивные способности и разум: каких водных животных следует защищать?» (PDF). Болезни водных организмов. 75 (2): 99–108. Дои:10.3354 / dao075099. PMID 17578249.
- ^ Меттам, Дж. Дж., Оултон, Л. Дж., МакКрохан, К. Р. и Снеддон, Л. У. (2011). «Эффективность трех типов анальгетиков в уменьшении боли у радужной форели Oncorhynchus mykiss». Прикладная наука о поведении животных. 133 (3): 265–274. Дои:10.1016 / j.applanim.2011.06.009.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c Волкерс, К.П.Б., Джуниор, Б.А., Менескаль-де-Оливейра, Л. и Хоффманн, А. (2013). «Стресс-индуцированная антиноцицепция у рыб, отмененная налоксоном». PLOS ONE. 8 (7): e71175. Bibcode:2013PLoSO ... 871175W. Дои:10.1371 / journal.pone.0071175. ЧВК 3728202. PMID 23936261.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Стинберген П.Дж. и Бардин Н. (2014). «Антиноцицептивные эффекты бупренорфина у личинок рыбок данио: альтернатива для моделей грызунов для изучения боли и ноцицепции?». Прикладная наука о поведении животных. 152: 92–99. Дои:10.1016 / j.applanim.2013.12.001.
- ^ Эренсинг, Р.Х., Мичелл, Г.Ф. и Кастин, А.Дж. (1982). «Аналогичный антагонизм морфиновой анальгезии MIF-1 и налоксоном у Carassius auratus». Фармакология, биохимия и поведение. 17 (4): 757–761. Дои:10.1016/0091-3057(82)90358-6. PMID 6129644. S2CID 31113845.CS1 maint: несколько имен: список авторов (связь)
- ^ Червова, Л. И Лапшин, Д.Н. (2000). «Опиоидная модуляция болевого порога у рыб». Доклады биологических наук. 375 (1): 590–591. Дои:10.1023 / а: 1026681519613. PMID 11211504. S2CID 1180288.
- ^ а б c Экрот, Дж. Р., Аас-Хансен, О., Снеддон, Л. У., Бичао, Х. и Дёвинг, К.Б. (2014). «Психологические и поведенческие реакции на вредные раздражители у атлантической трески (Gadus morhua)». PLOS ONE. 9 (6): e100150. Bibcode:2014PLoSO ... 9j0150E. Дои:10.1371 / journal.pone.0100150. ЧВК 4061104. PMID 24936652.CS1 maint: использует параметр авторов (связь)
- ^ Грандин, Т. (2015). «Глава 2 - Важность измерения для улучшения благополучия домашнего скота, птицы и рыбы». В Грандин Т. (ред.). Улучшение благополучия животных: практический подход.
- ^ Рейли, С.С., Куинн, Дж. П., Коссинс, А. и Снеддон, Л.У. (2008). «Поведенческий анализ ноцицептивного события у рыб: сравнение трех видов демонстрирует специфические реакции». Прикладная наука о поведении животных. 114 (1): 248–259. Дои:10.1016 / j.applanim.2008.01.016.CS1 maint: несколько имен: список авторов (связь)
- ^ Рокес, J.A.C., Аббинк, В., Геерд, Ф., ван де Вис, Х. и Флик, Г. (2010). «Стрижка хвостового плавника, болезненная процедура: исследования нильской тилапии и карпа». Физиология и поведение. 101 (4): 533–540. Дои:10.1016 / j.physbeh.2010.08.001. PMID 20705079. S2CID 25859917.CS1 maint: несколько имен: список авторов (связь)
- ^ Дунайер, Джоан, "Рыба: чувствительность вне досягаемости похитителя", The Animals 'Agenda, июль / август 1991 г., стр. 12-18.
- ^ «Познание животных». 2015-04-22. Получено 15 сентября, 2015.
- ^ Данлоп Р., Миллсопп С. и Лэминг П. (2006). «Обучение избеганию у золотой рыбки (Carassius auratus) и форели (Oncorhynchus mykiss) и последствия для восприятия боли». Прикладная наука о поведении животных. 97 (2): 255–271. Дои:10.1016 / j.applanim.2005.06.018.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Миллсопп, С. и Лэминг, П. (2007). «Компромисс между кормлением и избеганием шока у золотой рыбки (Carassius auratus)». Прикладная наука о поведении животных. 113 (1–3): 247–254. Дои:10.1016 / j.applanim.2007.11.004.
- ^ а б Снеддон Л.У., Брейтуэйт В.А. И Джентл М.Дж. (2003). «Тест нового объекта: исследование боли и страха у радужной форели». Журнал боли. 4 (8): 431–440. Дои:10.1067 / S1526-5900 (03) 00717-X. PMID 14622663.
- ^ Эшли П.Дж., Рингроуз С., Эдвардс К.Л., Уоллингтон Э., МакКрохан К.Р. и Снеддон Л.У. (2009). «Влияние вредной стимуляции на реакцию борьбы с хищниками и статус доминирования радужной форели». Поведение животных. 77 (2): 403–410. Дои:10.1016 / j.anbehav.2008.10.015. S2CID 19225428.CS1 maint: несколько имен: список авторов (связь)
- ^ Веник, Д. (2001). «Эволюция боли» (PDF). Vlaams Diergeneeskundig Tijdschrift. 70 (1): 17–21.
- ^ а б Браун, Кулум (2016). «Рыбная боль: неудобная правда». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 32.
- ^ Кук, С.Дж. И Снеддон Л.У. (2007). «Перспективы защиты животных при любительской рыбалке». Прикладная наука о поведении животных. 104 (3–4): 176–198. CiteSeerX 10.1.1.630.459. Дои:10.1016 / j.applanim.2006.09.002.
- ^ Лик, Дж. (14 марта 2004 г.). «Рыболовы встретятся с проверкой RSPCA». Санди Таймс. Получено 15 сентября, 2015.
- ^ Дигглз, Б. (2016). «Рыбная боль: изменит ли это текущую передовую практику в реальном мире?». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 35.
- ^ Снеддон, Л.У. (2006). «Этика и благополучие: восприятие боли у рыб». Бык. Евро. Доц. Рыбы. Патол. 26 (1): 6–10.
- ^ а б Скифтесвик, Энн Берит; Бровман, Говард I; Уотсон, Крейг А; Арлингхаус, Роберт; Кук, Стивен Дж; Cowx, Ian G; Дербишир, Стюарт В. Г.; Касумян, Александр; Ки, Брайан; Роуз, Джеймс Д; Шваб, Александр; Стивенс, Э. Дон (2018). «Благополучие водных животных: где они находятся, куда они направляются и что это значит для научных исследований, аквакультуры, любительской рыбалки и коммерческого рыболовства». Журнал морских наук ICES. 76 (1): 82–92. Дои:10.1093 / icesjms / fsy067. ISSN 1054-3139.
- ^ "Закон 1986 года о животных (научных процедурах)" (PDF). Домашний офис (Великобритания). Получено 23 сентября, 2015.
- ^ «Закон о защите животных 2006 г.». Правительство Великобритании. 2006 г.. Получено 25 сентября, 2015.
- ^ «Раздел 7 США - СЕЛЬСКОЕ ХОЗЯЙСТВО».
- ^ «Животные в исследованиях». neavs. Архивировано из оригинал 18 сентября 2015 г.. Получено 25 сентября, 2015.
- ^ Хенриксен, С., Ваагланд, Х., Сундт-Хансен, Л., Мэй, Р. и Фьельхейм, А. (2003). «Последствия восприятия боли у рыб при вылове и выпуске, аквакультуре и промысловом рыболовстве» (PDF).CS1 maint: несколько имен: список авторов (связь)
- ^ Роуз, JD (2002). «Нейроповеденческая природа рыб и вопрос осознания и боли» (PDF). Обзоры в науке о рыболовстве. 10 (1): 1–38. CiteSeerX 10.1.1.598.8119. Дои:10.1080/20026491051668. S2CID 16220451. Архивировано из оригинал (PDF) на 2012-10-10.
- ^ Роуз, JD (2002). "Рыба чувствует боль?". Архивировано из оригинал 20 января 2013 г.. Получено 27 сентября, 2007.
- ^ "Рыбы действительно чувствуют боль, - говорят ученые". Новости BBC. 30 апреля 2003 г.. Получено 20 мая 2010.
- ^ а б Грандин, Т. и Джонсон, К. (2005). Животные в переводе. Нью-Йорк: Скрибнер. стр.183–184. ISBN 978-0-7432-4769-6.
- ^ Роуз, JD (2003) Критика статьи: «Есть ли у рыб ноцицепторы: свидетельства эволюции сенсорной системы позвоночных» В архиве 2008-11-19 на Wayback Machine В: Информационные ресурсы о благополучии рыб, 1970–2003, Информационные ресурсы о благополучии животных № 20. Х. Э. Эриксон, Эд. Департамент сельского хозяйства США, Белтсвилл, Мэриленд. Стр. 49-51.
- ^ Ки, Б. (2016). «Почему рыбы не чувствуют боли». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 1.
- ^ «У рыб не хватает мозгов, чтобы чувствовать боль, - утверждает последняя школа мысли».. Телеграф. 10 февраля 2003 г.
- ^ Роуз, Дж. Д. (2007). «Антропоморфизм и« психическое благополучие »рыб». Болезни водных организмов. 75 (2): 139–154. Дои:10.3354 / dao075139. PMID 17578253.
- ^ Снеддон, Л.У. (28 августа 2012 г.). «Восприятие боли рыбами: почему критики не могут принять научные доказательства боли у рыб» (PDF). Цитировать журнал требует
| журнал =(помощь) - ^ Дербишир, С.В. (2016). «Рыбам не хватает мозгов и психологии для боли». Чувство животных: междисциплинарный журнал о чувствах животных. 1 (3): 18.
- ^ Вудрафф, М. (2017). «Сознание у костистых: есть что-то вроде рыбы». Чувство животных: междисциплинарный журнал о чувствах животных. 2 (13): 1.
- ^ Вудрафф, М. (2018). «Чувствительность рыб: подробнее о доказательствах». Чувство животных: междисциплинарный журнал о чувствах животных. 3 (13): 16.
- ^ Файнберг, Т. Э. и Маллатт, Дж. М. (2016). Древние истоки сознания: как мозг создавал опыт. Кембридж, Массачусетс: MIT Press.
- ^ Ньюби, Н.С. и Стивенс, Е.Д. (2008). «Влияние« болевого »теста уксусной кислоты на кормление, плавание и респираторные реакции радужной форели (Oncorhynchus mykiss)». Прикладная наука о поведении животных. 114 (1): 260–269. Дои:10.1016 / j.applanim.2007.12.006.
- ^ Снеддон, Л.У. (2009). «Влияние« болевого »теста уксусной кислоты на кормление, плавание и респираторные реакции радужной форели (Oncorhynchus mykiss): критика Newby and Stevens (2008)». Прикладная наука о поведении животных. 116 (1): 96–97. Дои:10.1016 / j.applanim.2008.07.006.
- ^ Ньюби, Н.С. и Стивенс, Э.Д. (2009). «Влияние« болевого »теста уксусной кислоты на кормление, плавание и респираторные реакции радужной форели (Oncorhynchus mykiss): критика Newby and Stevens (2008) - реакция». Прикладная наука о поведении животных. 116 (1): 97–99. Дои:10.1016 / j.applanim.2008.07.009.
- ^ а б «Рыбы действительно могут чувствовать боль и реагировать на нее так же, как люди». Университет Пердью. 29 апреля 2009 г.
внешняя ссылка
- Рыбная боль
- "Наука показывает, что рыбы чувствуют боль". Марк Бекофф. Huffington Post. 2015-12-29.
- «Чувствуют ли рыбы боль? Дебаты продолжаются». Марк Лалланилья. LiveScience.
- Как наиболее гуманно убивать ракообразных для употребления в пищу человеком? База знаний RSPCA. RSPCA Австралия
| Познание | |
|---|---|
| Интеллект | |
| Боль | |
| Отношение к мозгу | |
| |
| О рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||