Размножение рыб - Fish reproduction

Рыбы репродуктивные органы включают яички и яичники. У большинства видов гонады находятся парные органы аналогичного размера, которые могут быть частично или полностью сплавлены.[1] Также может быть ряд вторичных органов, повышающих репродуктивную способность. В генитальный сосочек небольшая мясистая трубочка за анус у некоторых рыб, из которых сперма или яйца освобождены; пол рыбы часто можно определить по форме ее сосочка.
Анатомия
Семенники
У большинства самцов есть два яички аналогичного размера. На случай, если акулы, семенники справа обычно больше[нужна цитата ]. Примитивный рыба без челюсти имеют только один семенник, расположенный по средней линии тела, хотя даже он образуется в результате слияния парных структур эмбриона.[2]
Под жесткой мембранной оболочкой белочной оболочкой, яички некоторых костистость рыба, содержит очень тонкие спиральные трубы, называемые семенные канальцы. Канальцы выстланы слоем клеток (стволовые клетки ) что из половое созревание в старости, развиваться в сперма клетки (также известен как сперматозоиды или мужчина гаметы ). Развивающиеся сперматозоиды перемещаются по семенным канальцам к rete testis расположен в средостение яичка, в отводящие протоки, а затем в придаток яичка где созревают вновь созданные сперматозоиды (см. сперматогенез ). Сперма переходит в семявыносящий проток, и в конечном итоге изгнаны через уретра и из отверстие уретры через мышечные сокращения.
Однако у большинства рыб семенных канальцев нет. Вместо этого сперма вырабатывается в сферических структурах, называемых ампулы спермы. Это сезонные структуры, которые высвобождают свое содержимое в период размножения, а затем повторно поглощаются организмом. Перед следующим периодом размножения новые ампулы сперматозоидов начинают формироваться и созревать. В остальном ампулы по существу идентичны семенным канальцам в высшие позвоночные, включая такой же диапазон типов ячеек.[2]
С точки зрения сперматогония распределение, структура костистые кости яички имеет два типа: в большинстве случаев сперматогонии возникают на всем протяжении семенные канальцы, а у атериноморфных рыб они приурочены к дистальный часть этих структур. Рыба может быть кистозной или полукистозной. сперматогенез в отношении фазы высвобождения половых клеток в кистах в семенные канальцы просвет.[1]
Яичники

Многие функции, найденные в яичники являются общими для всех позвоночных, включая наличие фолликулярные клетки и белочной оболочкой В яичнике рыбы могут быть сотни или даже миллионы оплодотворенных икринок в любой момент времени. Свежие яйца могут развиваться из зародышевый эпителий на протяжении всей жизни. Желтые тела встречаются только у млекопитающих, а у некоторых эластожаберный рыбы; у других видов остатки фолликула быстро рассасываются яичником.[2] Яичник костистые кости часто содержит дупло, лимфа -заполненное пространство, которое открывается в яйцевод, и в который проливаются яйца.[2] Большинство нормальных самок рыб имеют два яичника. В некоторых эластожаберные ветви, только правый яичник развивается полностью. В примитивном рыба без челюсти, а у некоторых костистых имеется только один яичник, образованный в результате слияния парных органов у зародыша.[2]
Яичники рыб могут быть трех типов: гимнастические, вторичные и цистовариальные. В первом типе ооциты выпускаются прямо в целомический полости, а затем введите устье, затем через яйцевод и устраняются. Вторичная гимнастическая потеря яичников яйцеклетка в целомудрие из которого они идут прямо в яйцевод. В третьем типе ооциты выводятся наружу через яйцевод.[3] Гимноварии - примитивное состояние, обнаруженное в двоякодышащая рыба, осетр, и Bowfin. Кистоварии характерны для большинства костистых трубок, в которых просвет яичника непрерывен с яйцеводом.[1] Вторичные гимназии находятся в лососевые и несколько других костистых.
Яйца


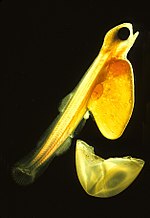
Икра рыб и амфибии желеобразные. Хрящевые рыбы (акулы, скаты, скаты, химеры) яйца оплодотворяются внутренне и демонстрируют широкий спектр как внутреннего, так и внешнего эмбрионального развития. Большинство видов рыб откладывают икру, которую оплодотворяют извне, обычно самцы осеменяют икру после того, как самка их откладывает. Эти яйца не имеют скорлупы и могут высохнуть на воздухе. Даже дышащие воздухом земноводные откладывают яйца в воде или в защитной пене, как в случае с прибрежной древесной лягушкой в пенистом гнезде, Chiromantis xerampelina.
Интромитентные органы


Мужской хрящевые рыбы (акулы и скаты), а также самцы некоторых живородящих скатные плавники, имеют плавники, которые были изменены, чтобы функционировать как внутренние органы, репродуктивные придатки, которые позволяют внутреннее оплодотворение. У луко-плавниковых рыб их называют гоноподии или андроподиумы, а у хрящевых рыб их называют застежки.
Гоноподии встречаются у самцов некоторых видов в Anablepidae и Poeciliidae семьи. Это анальные плавники, которые были модифицированы для работы в качестве подвижных интромитентных органов и используются для оплодотворения самок. молока во время вязки. Третий, четвертый и пятый лучи анального плавника самца образуют трубчатую структуру, в которую выбрасывается сперма рыбы.[6] Когда гоноподий готов к спариванию, он становится прямостоячим и направлен вперед в сторону самки. Вскоре самец вставляет орган в половое отверстие самки с помощью крючкообразных приспособлений, которые позволяют рыбе хвататься за самку для обеспечения оплодотворения. Если самка остается неподвижной и ее партнер контактирует с ее желудочно-кишечным трактом своим гоноподием, она оплодотворяется. Сперма сохраняется в яйцеводе самки. Это позволяет самкам оплодотворять себя в любое время без дополнительной помощи самцов. У некоторых видов гоноподий может составлять половину общей длины тела. Иногда плавник бывает слишком длинным, чтобы его можно было использовать, как у лирохвостых пород. Xiphophorus helleri. У женщин, получавших гормональные препараты, могут развиться гоноподии. Они бесполезны для разведения.
Подобные органы с аналогичными характеристиками встречаются у других рыб, например у андроподиум в Хемирхамфодон или в Goodeidae.[7]
Застежки встречаются у мужчин хрящевые рыбы. Они представляют собой заднюю часть тазовых плавников, которые также были изменены, чтобы функционировать как интромитентные органы, и используются для направления спермы в самки. клоака во время совокупления. Акт спаривания у акул обычно включает поднятие одной из кламмеров, чтобы вода попала в сифон через определенный отверстие. Затем застежка вставляется в клоаку, где она открывается, как зонтик, чтобы закрепить ее положение. Затем сифон начинает сокращаться, вытесняя воду и сперму.[8][9]
Физиология
Оогония Развитие костистых рыб варьируется в зависимости от группы, и определение динамики оогенеза позволяет понять процессы созревания и оплодотворения. Изменения в ядро, ооплазма и окружающие ее слои характеризуют процесс созревания ооцита.[1]
Постовуляторный фолликулы структуры, образующиеся после выхода ооцитов; У них нет эндокринный функции, имеют широкий просвет неправильной формы и быстро реабсорбируются в процессе, включающем апоптоз фолликулярных клеток. Дегенеративный процесс, называемый фолликулярная атрезия реабсорбирует не рожденные вителлогенные ооциты. Этот процесс также может происходить, но реже, в ооцитах на других стадиях развития.[1]
Некоторые рыбы гермафродиты, имея как семенники, так и яичники либо на разных этапах своего жизненного цикла, либо, как в деревушки, есть их одновременно.
Репродуктивные стратегии
У рыб оплодотворение икры может быть как внешним, так и внутренним. У многих видов рыб плавники были изменены, чтобы Внутреннее оплодотворение. Точно так же развитие эмбриона может быть внешним или внутренним, хотя у некоторых видов наблюдается различие между ними на разных стадиях развития эмбриона. Тьерри Лоде описал репродуктивные стратегии с точки зрения развития зиготы и взаимоотношений с родителями; Существует пять классификаций - яйцорождение, яйцекладение, яйцеводство, гистотрофическое живорождение и гемотрофическое живорождение.[10]
Яйценоскость
Овулянтность означает, что самка откладывает неоплодотворенные яйца (яйцеклетки), которые затем необходимо оплодотворять извне.[10] Примеры яйцекладущих рыб включают: лосось, золотая рыбка, цихлиды, тунец и угри. У большинства этих видов оплодотворение происходит вне тела матери, при этом самцы и самки теряют свои гаметы в окружающую воду.
Яйцеклад
Яйцеклад - это процесс внутреннего оплодотворения, поэтому самка сбрасывает зиготы (или недавно развивающиеся эмбрионы) в воду.[10] часто с добавлением важных внешних тканей. Более 97% всех известных рыб яйцекладущий.[11] У яйцекладущих рыб внутреннее оплодотворение требует от самца использования интромитентный орган для доставки спермы в половое отверстие самки. Примеры включают яйцекладущих акул, таких как рог акулы, и яйцекладущие лучи, такие как коньки. В этих случаях самец комплектуется парой модифицированных тазовый плавники, известные как застежки.
Морская рыба может производить большое количество икры, которые часто выбрасываются в толщу открытой воды. Яйца имеют средний диаметр 1 миллиметр (0,039 дюйма). Яйца обычно окружены внеэмбриональные оболочки но не образует твердую или мягкую оболочку вокруг этих мембран. У некоторых рыб есть толстая кожистая оболочка, особенно если они должны выдерживать физическую нагрузку или высыхание. Эти яйца также могут быть очень маленькими и хрупкими.

Яйцо минога

Яйцо кошачья акула (кошелек русалки )

Яйцо бычья акула

Яйцо химера

Только что вылупившееся молодняк яйцекладущих рыб называется личинки. Обычно они плохо сформированы, несут большой желточный мешок (для питания) и очень отличаются по внешнему виду от молодых и взрослых особей. Личиночный период у яйцекладущих рыб относительно короткий (обычно всего несколько недель), а личинки быстро растут и изменяют внешний вид и структуру (процесс, называемый метаморфоза ) стать несовершеннолетними. Во время этого перехода личинки должны переключиться со своего желточного мешка на питание. зоопланктон добыча, процесс, который обычно зависит от недостаточной плотности зоопланктона, в результате чего многие личинки умирают от голода.
Яйцеводство
В яйцекладущие после внутреннего оплодотворения яйца развиваются в организме матери, но получают мало или совсем не получают питания непосредственно от матери, в зависимости от запаса пищи внутри яйца, желток.[10] Каждый эмбрион развивается в собственном яйце. Знакомые примеры яйцекладущих рыб включают: гуппи, ангелы акулы, и латимерия.
Живородство
Есть два типа живородство, в зависимости от того, как потомство получает питательные вещества.
- Гистотрофное (поедание тканей) живородство означает, что эмбрионы развиваются в яйцеводах самки, но получают питательные вещества, потребляя другие ткани, такие как яйцеклетки (оофагия ) или зиготы.[10] Это наблюдалось в первую очередь среди акул, таких как короткопёрый мако и сельдевая акула, но известен также несколькими костистыми рыбками, такими как полуклюв Nomorhamphus ebrardtii.[12] Необычный способ живородения - адельфофагия или внутриутробный каннибализм, в котором самые крупные эмбрионы поедают более слабых, более мелких нерожденных братьев и сестер. Это чаще всего встречается среди акул, таких как серая акула-медсестра, но также сообщалось о Nomorhamphus ebrardtii.[12]
- Гемотрофное (поедание крови) живородство означает, что эмбрионы развиваются в яйцеводе самки (или самца), а питательные вещества поступают непосредственно от родителя, как правило, через структуру, аналогичную или аналогичную структуре матки. плацента видел в млекопитающие.[10] Примеры хемотрофных рыб включают прибой, сплитфины, лимонная акула, морские коньки и игла.
Аквариумисты обычно называют яйцеживородящих и живородящих рыб живородящие.
Гермафродитизм




Гермафродитизм происходит, когда данный особь вида обладает как мужскими, так и женскими репродуктивными органами или может выбирать между обладанием сначала одним, а затем другим. Гермафродитизм часто встречается у беспозвоночных, но редко у позвоночных. Его можно противопоставить гонохоризм, где каждая особь вида - это мужчина или женщина, и остается таким на протяжении всей своей жизни. Большинство рыб - гонохористы, но известно, что гермафродитизм встречается у 14 семейств. костистость Рыбы.[13]
Обычно гермафродиты последовательный, то есть они могут сменить пол, обычно от женщины к мужчине (протогиния ). Это может произойти, если из группы самок удалить доминирующего самца. Самая крупная самка в гареме может за несколько дней сменить пол и заменить доминирующего самца.[13] Это находится среди коралловые рифы такие как групперы, рыбы-попугаи и губаны. Менее распространен переход самца на самку (протандрия ).[14]:162 Например, большинство губаны находятся первобытный гермафродиты в гаремической брачной системе.[15][16] Гермафродитизм допускает сложные брачные системы. Губаны демонстрируют три разные системы спаривания: полигинный, лекоподобный и беспорядочные брачные системы.[17] Групповой и парный нерест происходят в рамках систем спаривания. Тип нереста зависит от размера тела самца.[16] Лаброиды обычно демонстрируют нерестовой нерест, выпуская большое количество планктонных яиц, которые разносятся приливными течениями; взрослые губаны не взаимодействуют с потомством.[18] Губаны особой подгруппы семейства Labridae, Labrini, не демонстрируют широковещательного нереста.
Реже гермафродиты могут быть синхронный, что означает, что они одновременно обладают яичниками и яичками и могут одновременно функционировать как представители любого пола. Черные деревушки «по очереди выпускают сперматозоиды и яйца во время нереста. Поскольку такая торговля яйцами выгодна обоим особям, деревушки обычно моногамны на короткие периоды времени - необычная ситуация для рыб».[19] Пол многих рыб не фиксирован, но может измениться с физическими и социальными изменениями среды, в которой обитает рыба.[20]
В частности, у рыб гермафродитизм может окупиться в ситуациях, когда у одного пола больше шансов выжить и размножаться, возможно, потому, что он крупнее.[21] Анемоновые рыбы являются последовательными гермафродитами, которые рождаются мужчинами и становятся женщинами только тогда, когда становятся зрелыми. Рыбы-анемоны живут вместе моногамно в анемон, защищенный укусами анемона. Самцам не нужно соревноваться с другими самцами, а самки анемонов обычно крупнее. Когда самка умирает, туда перемещается молодь (самец) рыбы-анемона, и «постоянный самец затем превращается в самку, и репродуктивные преимущества комбинации большой самки и маленькой самки сохраняются».[22] У других рыб смена пола обратима. Например, если некоторые бычки сгруппированы по полу (мужской или женский), некоторые поменяют пол.[14]:164[21]
В мангровые заросли Криптолебиас марморатус производит яйца и сперму мейоз и обычно размножается самооплодотворением. Каждый человек гермафродит Обычно оплодотворяется само, когда яйцеклетка и сперма, произведенные внутренним органом, соединяются внутри тела рыбы.[23] В природе этот способ размножения может давать очень высокие результаты. гомозиготный линии, состоящие из особей настолько генетически однородных, что фактически идентичны друг другу.[24][25] Способность к самоопылению у этих рыб, по-видимому, сохранялась, по крайней мере, несколько сотен тысяч лет.[26]
Несмотря на то что инбридинг, особенно в крайней форме самооплодотворения, обычно считается вредным, поскольку приводит к выражению вредных рецессивных аллелей, самооплодотворение действительно приносит пользу гарантия оплодотворения (репродуктивная гарантия) в каждом поколении.[24]
Сексуальный паразитизм
Сексуальный паразитизм - это способ половое размножение, уникальный для удильщик, в котором самцы вида намного меньше самок и полагаются на самок в поисках пищи и защиты от хищников. Самцы ничего не возвращают, кроме сперма в котором женщины нуждаются, чтобы произвести следующее поколение.
Некоторые удильщики, вроде морских глубоководных цератиоид группа, используйте этот необычный метод вязки. Поскольку особи распределены очень редко, встречи также очень редки. Поэтому найти себе пару проблематично. Когда ученые впервые начали ловить цератиноидных удильщиков, они заметили, что все экземпляры были самками. Эти люди были ростом в несколько сантиметров, и почти все они выглядели как паразиты прикреплены к ним. Оказалось, что эти «паразиты» были сильно редуцированными самцами цератиоидных удильщиков. Это указывает на то, что удильщик использует полиандрический система спаривания.
Способы, с помощью которых рыба-удильщик находят партнеров, различны. У некоторых видов крошечные глаза, непригодные для определения самок, у других недоразвитые ноздри, поэтому маловероятно, что они смогут эффективно найти самок с помощью обоняния.[27] Когда самец находит самку, он кусает ее кожу и выпускает фермент который переваривает кожу его рта и ее тело, сплавляя пару до уровня кровеносных сосудов.[28] Самец становится зависимым от самки-хозяина в своем выживании, получая питательные вещества через свою теперь общую систему кровообращения, а взамен поставляет сперму самке. После слияния самцы увеличиваются в объеме и становятся намного крупнее свободноживущих самцов этого вида. Они живут и сохраняют репродуктивную функцию до тех пор, пока жива самка, и могут принимать участие в нескольких нерестах.[27] Этот крайний половой диморфизм гарантирует, что, когда самка будет готова к нересту, у нее немедленно появится помощник.[29] Несколько самцов могут быть включены в одну индивидуальную самку с до восьми самцов у некоторых видов, хотя некоторые таксоны, по-видимому, имеют правило один самец на самку.[27] В дополнение к физиологической адаптации, иммунная система изменяется, чтобы позволить соединение.[30]
Одно объяснение эволюция Полового паразитизма заключается в том, что относительно низкая плотность самок в глубоководной среде оставляет мало возможностей для выбора партнера среди удильщиков. Самки остаются крупными, чтобы приспособиться плодовитость, о чем свидетельствуют их крупные яичники и яйца. Ожидается, что самцы уменьшатся в размерах, чтобы снизить метаболические затраты в условиях ограниченных ресурсов и разовьют узкоспециализированные способности к поиску самок. Если мужчине удастся найти женскую паразитическую привязанность, то, в конечном итоге, он с большей вероятностью улучшит жизнеспособность на протяжении всей жизни по сравнению со свободной жизнью, особенно когда шансы найти будущих помощников невелики. Дополнительным преимуществом паразитизма является то, что сперму самца можно использовать при множественных оплодотворениях, так как он всегда доступен самке для спаривания. Более высокая плотность встреч самцов и самок может коррелировать с видами, которые демонстрируют факультативный паразитизм или просто использовать более традиционные временные контактные вязки.[31]
Партеногенез
Партеногенез это форма бесполое размножение в котором рост и развитие эмбрионы происходить без оплодотворение. У животных партеногенез означает развитие эмбриона из неоплодотворенной Клетка яйца. Первое полностью женское (однополое) размножение в г. позвоночные был описан в Амазонка Молли в 1932 г.[32] С тех пор было описано не менее 50 видов однополых позвоночных, в том числе не менее 20 рыб, 25 ящериц, один вид змей, лягушек и саламандр.[33] Как и в случае со всеми типами бесполого размножения, партеногенез связан как с затратами (низкое генетическое разнообразие и, следовательно, подверженностью неблагоприятным мутациям), так и с преимуществами (размножение без потребности в самце).
Партеногенез у акул подтвержден в капот[34] и зебра акула.[35] Другие, обычно половые виды, могут иногда воспроизводиться партеногенетически, и молот и черноперые акулы[36] являются недавним дополнением к известному списку факультативных партеногенетических позвоночных.
Частным случаем партеногенеза является гиногенез. При этом типе воспроизводства потомство производится по тому же механизму, что и при партеногенезе, однако развитие яйца стимулируется просто за счет присутствие из сперма - сперматозоиды не передают потомству никакого генетического материала. Поскольку все гиногенетические виды - самки, активация их яиц требует спаривания с самцами близкородственных видов для получения необходимого стимула. В Амазонка Молли, (на фото), воспроизводится путем гиногенеза.
Другие
Скульпин лосося (Alcichthys elongatus ) - морская костистая кость с уникальным репродуктивным способом, называемым «внутренняя гаметическая ассоциация». Сперма вводится в яичник путем совокупления, а затем попадает в микропилярный канал овулированных яиц в полости яичника. Однако фактическое слияние сперматозоидов и яйцеклеток не происходит, пока яйца не будут выпущены в морскую воду.[37]
Инбридинг
Инбридинговая депрессия
Эффект инбридинг по репродуктивному поведению изучалась в Poeciliid рыбы Heterandria formosa.[38] Было обнаружено, что одно поколение полных сибсов снижает репродуктивную способность и вероятный репродуктивный успех мужского потомства. Другие отображаемые черты инбридинговая депрессия - жизнеспособность потомства и время созревания как самцов, так и самок.
Воздействие химического агента окружающей среды на рыб данио, аналогичное тому, которое вызвано антропогенным загрязнением, усиливало влияние инбридинга на ключевые репродуктивные черты.[39] Жизнеспособность эмбрионов была значительно снижена у инбредных рыб, подвергшихся воздействию, и у инбредных самцов наблюдалась тенденция давать меньше потомства.
Поведение несовершеннолетних Кижуч с низким или средним инбридингом сравнивали в парных соревнованиях.[40] Рыбы с низким уровнем инбридинга показали почти вдвое более агрессивное преследование при защите территории, чем рыбы со средним инбридингом, и, кроме того, имели более высокую удельную скорость роста. Значительный эффект от инбридинговая депрессия на выживаемость молоди, но только в конкурентной среде с высокой плотностью, что позволяет предположить, что внутривидовая конкуренция может усиливать пагубные последствия инбридинга.
Избегание инбридинга
Инбридинг обычно имеет негативные последствия для приспособленности (инбридинговая депрессия), и в результате виды выработали механизмы, позволяющие избегать инбридинга. Многочисленные избегание инбридинга описаны механизмы, действующие до спаривания. Однако механизмы предотвращения инбридинга, которые действуют после совокупления, менее известны. У гуппи происходит пост-копулятивный механизм избегания инбридинга, основанный на конкуренции между сперматозоидами соперничающих самцов за достижение оплодотворение.[41] В соревнованиях между спермой от неродственного мужчины и от полноправного брата или сестры наблюдалась значительная предвзятость в отцовстве в отношении неродственного мужчины.[41]
Считается, что инбридинговая депрессия во многом обусловлена выражением гомозиготный вредные рецессивные мутации.[42] Ауткроссинг между неродственными особями приводит к полезной маскировке вредных рецессивных мутаций в потомстве.[43]
Сексуальные стратегии
Стратегии нереста
Нерестилища
Примеры


- Золотая рыбка
Золотая рыбка, как все карповые, являются яйцекладушками. Обычно они начинают размножаться после значительного изменения температуры, часто весной. Самцы преследуют самок, заставляя их выпустить яйца, толкая их. Когда золотая рыбка-самка откладывает икру, самец ее оплодотворяет. Их яйца липкие и прикрепляются к водной растительности. Яйца вылупляются в течение 48-72 часов. Примерно через неделю мальки начинают принимать свою окончательную форму, хотя может пройти год, прежде чем они приобретут зрелый цвет золотой рыбки; до тех пор они имеют металлический коричневый цвет, как и их дикие предки. В первые недели жизни мальки быстро растут - адаптация, порожденная высоким риском быть съеденными взрослой золотой рыбкой.
- Карп
Карп нерестится в период с апреля по август, что в значительной степени зависит от климата и условий, в которых они живут. Уровень кислорода в воде, доступность корма, размер каждой рыбы, возраст, количество нерестов рыбы раньше и температура воды - все это факторы, влияющие на то, когда и сколько яиц каждый карп нерестит одновременно.[44]
- Сиамские бойцовые рыбы
До нереста самец Сиамские бойцовые рыбы строить пузырьковые гнезда разного размера на поверхности воды. Когда самец начинает интересоваться самкой, он раздувает свои жабры, скручивает тело и расправляет плавники. Самка темнеет в цвете и изгибает свое тело взад и вперед. Акт нереста происходит в «брачных объятиях», когда самец обертывает свое тело вокруг самки, каждое объятие приводит к высвобождению 10-40 яиц, пока самка не исчерпает свои яйца. Самец со своей стороны отпускает молока в воду и оплодотворение происходит извне. Во время и после нереста самец использует свой рот, чтобы извлечь тонущие яйца и положить их в гнездо пузыря (во время спаривания самка иногда помогает своему партнеру, но чаще она просто съедает все яйца, которые ей удается поймать).Как только самка выпустит все свои яйца, ее прогоняют с территории самца, так как вполне вероятно, что она съест яйца из-за голода.[45] Затем яйца остаются на попечении самца. Он держит их в гнезде с пузырьками, следя за тем, чтобы никто не упал на дно, и ремонтирует гнездо по мере необходимости. Инкубация длится 24–36 часов, и только что вылупившиеся личинки остаются в гнезде в течение следующих 2–3 дней, пока их желточные мешки полностью не рассосутся. После этого мальки покидают гнездо и начинается стадия свободного плавания.[46]

Сиамские бойцовые рыбы строить пузырьковые гнезда разных размеров.

Пара сиамских бойцовых рыб нерестится под своим пузырьковым гнездом.

Однодневные личинки сиамских бойцовых рыб в пузырчатом гнезде - их желточные мешки еще не рассосаны

Пятнадцатидневный свободно плавающий мальк сиамской бойцовой рыбы




Смотрите также
Рекомендации
- ^ а б c d е Гимарайнш-Крус, Родриго Дж., Родриго Дж .; Сантос, Хосе Э. дос; Сантос, Гилмар Б. (2005). «Строение гонад и гаметогенез Лорикария лентигинозная Исбрюкер (Рыбы, Teleostei, Siluriformes) ". Rev. Bras. Zool. 22 (3): 556–564. Дои:10.1590 / S0101-81752005000300005. ISSN 0101-8175.
- ^ а б c d е Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного. Филадельфия, Пенсильвания: Holt-Saunders International. С. 385–386. ISBN 978-0-03-910284-5.
- ^ Brito, M.F.G .; Баццоли, Н. (2003). «Размножение сома сурубима (Pisces, Pimelodidae) в реке Сан-Франциско, регион Пирапора, Минас-Жерайс, Бразилия». Arquivo Brasileiro de Medicina Veterinária e Zootecnia. 55 (5): 624–633. Дои:10.1590 / S0102-09352003000500018. ISSN 0102-0935.
- ^ Мастерсон, Дж. "Gambusia affinis". Смитсоновский институт. Получено 21 октября 2011.
- ^ Кунц, Альберт (1913). "Заметки о привычках, морфологии репродуктивных органов и эмбриологии живородящих рыб Gambusia affinis". Бюллетень Бюро рыболовства США. 33: 181–190.
- ^ Капур Б.Г. и Кханна Б. (2004) Справочник по ихтиологии С. 497–498, Springer Science & Business Media. ISBN 9783540428541.
- ^ Хелфман Джи, Коллетт БиБи, Фэйси Д.Х. и Боуэн Б.В. (2009) Разнообразие рыб: биология, эволюция и экология п. 35, Вили-Блэквелл. ISBN 978-1-4051-2494-2.
- ^ «Системный глоссарий». FishBase. Получено 2013-02-15.
- ^ Heinicke, Matthew P .; Naylor, Gavin J. P .; Хеджес, С. Блэр (2009). Древо жизни: хрящевые рыбы (Chondrichthyes). Издательство Оксфордского университета. п. 320. ISBN 978-0191560156.
- ^ а б c d е ж Лоде, Т. (2012). «Яйцо или живорождение? Вот в чем вопрос…». Репродуктивная биология. 12 (3): 259–264. Дои:10.1016 / j.repbio.2012.09.001. PMID 23153695.
- ^ Питер Скотт: Живородящие рыбы, п. 13. Тетра Пресс 1997. ISBN 1-56465-193-2
- ^ а б Мейснер, А; Бернс, Дж (1997). "Живородство в родах полуклювов Дермогенис и Nomorhamphus (Teleostei: Hemiramphidae) ". Журнал морфологии. 234 (3): 295–317. Дои:10.1002 / (SICI) 1097-4687 (199712) 234: 3 <295 :: AID-JMOR7> 3.0.CO; 2-8. PMID 29852651.
- ^ а б Шапиро Д.Ю. (1984) «Процессы смены пола и социодемографические процессы у рыб коралловых рифов» Страницы 103–116 в GW Potts and RK Wootoon, eds., Размножение рыб: стратегии и тактика, Academic Press.
- ^ а б Мойл ПБ и Чех Джей Джей (2004) Рыбы, Введение в ихтиологию. 5-е изд., Бенджамин Каммингс. ISBN 978-0-13-100847-2
- ^ Робертсон, Д.Р .; Р. Р. Уорнер (1978). "Половые модели лаброидных рыб Западной части Карибского бассейна II: рыбы-попугаи (Scaridae)". Вклад Смитсоновского института в зоологию. 255 (255): 1–26. Дои:10.5479 / si.00810282.255.
- ^ а б Kazancioglu, E .; S.H. Алонзо (август 2010). «Сравнительный анализ смены пола у Labridae подтверждает гипотезу преимущества в размере». Эволюция. 64 (8): 2254–2264. Дои:10.1111 / j.1558-5646.2010.01016.x. PMID 20394662.
- ^ Colin, P.L .; Л. Дж. Белл (1992). «Аспекты нереста лабридных и скаридных рыб (Pisces, Labroidi) на атолле Эниветак, Маршалловы острова с примечаниями о других семьях (исправленная перепечатка)».. Экологическая биология рыб. 33 (3): 330–345. Дои:10.1007 / BF00005881.
- ^ Hanel, R .; М. В. Вестнит; К. Штурбауэр (декабрь 2002 г.). «Филогенетические отношения, эволюция поведения по уходу за потомством и географическое видообразование в племени губанов Labrini». Журнал молекулярной эволюции. 55 (6): 776–789. Bibcode:2002JMolE..55..776H. Дои:10.1007 / s00239-002-2373-6. PMID 12486536. S2CID 3002410.
- ^ Фишер Э.А., Петерсон Ч.В. (1987). «Эволюция сексуальности морских окуней». Бионаука. 37 (7): 482–489. Дои:10.2307/1310420. JSTOR 1310420.
- ^ Чан СТХ и Йунг WSB (1983) «Контроль пола и изменение пола у рыб в естественных условиях». Страницы 171–222 в WS Hoar, DJ Randall and EM Donaldson, eds., Физиология рыб 9B: Воспроизводство, поведение и контроль фертильности. Академическая пресса.
- ^ а б Кувамура, Тецуо; Накашима, Ясухиро (1998). «Новые аспекты смены пола среди рифовых рыб: недавние исследования в Японии». Экологическая биология рыб. 52: 125–135. Дои:10.1023 / А: 1007389519702. S2CID 30244987.
- ^ Фрике, Ганс; Фрике, Симона (1977). «Моногамия и смена пола путем агрессивного доминирования у рыб коралловых рифов». Природа. 266 (5605): 830–832. Bibcode:1977Натура.266..830F. Дои:10.1038 / 266830a0. PMID 865603. S2CID 4177265.
- ^ Sakakura, Y .; Сояно, К .; Noakes, D.L.G .; Хагивара, А. (2006). «Морфология гонад у самоопыляющихся мангровых грибов Kryptolebias marmoratus». Ихтиологические исследования. 53 (4): 427–430. Дои:10.1007 / s10228-006-0362-2. HDL:10069/35713. S2CID 9474211.
- ^ а б Avise JC, Татаренков А (2012). «Аргумент Алларда против утверждения Бейкера об адаптивном значении самоопыления у рыб-гермафродитов». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 109 (46): 18862–7. Bibcode:2012PNAS..10918862A. Дои:10.1073 / pnas.1217202109. ЧВК 3503157. PMID 23112206.
- ^ Эрли Р.Л., Ханнинен А.Ф., Фуллер А., Гарсия М.Дж., Ли Е.А. (2012). «Фенотипическая пластичность и интеграция в мангровых зарослях (Kryptolebias marmoratus): проспект». Интегр. Комп. Биол. 52 (6): 814–27. Дои:10.1093 / icb / ics118. ЧВК 3501102. PMID 22990587.
- ^ Татаренков А, Лима С.М., Тейлор Д.С., Avise JC (2009). «Долгосрочное сохранение самооплодотворения в кладе рыб». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 106 (34): 14456–9. Bibcode:2009ПНАС..10614456Т. Дои:10.1073 / pnas.0907852106. ЧВК 2732792. PMID 19706532.
- ^ а б c Питч, Теодор В. (25 августа 2005 г.). «Диморфизм, паразитизм и пол снова: способы размножения среди глубоководных цератиоидных удильщиков (Teleostei: Lophiiformes)». Ихтиологические исследования. 52 (3): 207–236. Дои:10.1007 / s10228-005-0286-2. S2CID 24768783.
- ^ Гулд, Стивен Джей (1983). Куриные зубы и конские пальцы. Нью-Йорк: W. W. Norton & Company. ISBN 978-0-393-01716-8.
- ^ Теодор В. Питч (июль 1975 г.). «Преждевременный половой паразитизм в глубоководных цератиоидных удильщиках, Cryptopsaras couesi Gill». Природа. 256 (5512): 38–40. Bibcode:1975Натура.256 ... 38П. Дои:10.1038 / 256038a0. S2CID 4226567.
- ^ Суонн, Джереми Б. (30 июля 2020 г.). «Иммуногенетика полового паразитизма». Наука: eaaz9445. Дои:10.1126 / science.aaz9445. HDL:21.11116 / 0000-0006-CE67-F. PMID 32732279.
- ^ Мия, Масаки; Питч, Теодор В.; Орр, Джеймс В.; Арнольд, Рэйчел Дж; Сато, Такаши П.; Шедлок, Эндрю М; Хо, Сюань-Цзин; Симадзаки, Мицуоми; Ябэ, Мамору; Нисида, Муцуми (1 января 2010 г.). «Эволюционная история удильщиков (Teleostei: Lophiiformes): митогеномная перспектива». BMC Эволюционная биология. 10 (1): 58. Дои:10.1186/1471-2148-10-58. ЧВК 2836326. PMID 20178642.
- ^ Hubbs, C.L .; Хаббс, Л. К. (1932). «Видимый партеногенез в природе у рыб гибридного происхождения». Наука. 76 (1983): 628–630. Bibcode:1932Sci .... 76..628H. Дои:10.1126 / science.76.1983.628. PMID 17730035.
- ^ Vrijenhoek, R.C., R.M. Доули, Си Джей Коул и Дж. П. Богарт. 1989. Список известных однополых позвоночных, стр. 19-23. в: Эволюция и экология однополых позвоночных. Р.М. Доули и Дж. П. Богарт (ред.) Бюллетень 466, Государственный музей Нью-Йорка, Олбани, Нью-Йорк
- ^ Chapman, Demian D .; Shivji, Mahmood S .; Луи, Эд; Соммер, Джули; Флетчер, Хью; Продёль, Пауло А. (2007). "Девственное рождение у акулы-молота". Письма о биологии. 3 (4): 425–427. Дои:10.1098 / rsbl.2007.0189. ЧВК 2390672. PMID 17519185.
- ^ Робинсон, Д. П .; Baverstock, W .; Аль-Джару, А .; Hyland, K .; Хазанехдари, К. А. (2011). "Ежегодно повторяющийся партеногенез у акулы-зебры Стегостома фасциатум". Журнал биологии рыб. 79 (5): 1376–1382. Дои:10.1111 / j.1095-8649.2011.03110.x. PMID 22026614.
- ^ Chapman, D. D .; Firchau, B .; Шивджи, М. С. (2008). «Партеногенез у крупнотелой акулы-реквиема, черноголовки». Журнал биологии рыб. 73 (6): 1473–1477. Дои:10.1111 / j.1095-8649.2008.02018.x.
- ^ Коя, Ю., Мунехара, Х., Такано, К. (2002). «Хранение и подвижность сперматозоидов в яичнике морского бычка Alcichthys alcicornis (Teleostei: Scorpaeniformes) с внутренней гаметической ассоциацией». Журнал экспериментальной зоологии. 292 (2): 145–155. Дои:10.1002 / jez.1150. PMID 11754030.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Ала-Хонкола О., Уддстрём А., Паули Б.Д., Линдстрем К. (2009). «Сильная инбридинговая депрессия в брачном поведении самцов поецилиидной рыбы». J. Evol. Биол. 22 (7): 1396–406. Дои:10.1111 / j.1420-9101.2009.01765.x. PMID 19486236.
- ^ Бикли Л.К., Браун А.Р., Хоскен Д.Д., Гамильтон П.Б., Ле Пейдж Г., Пол Г.К., Оуэн С.Ф., Тайлер С.Р. (2013). «Интерактивные эффекты инбридинга и эндокринных нарушений на воспроизводство у модельной лабораторной рыбы». Evol Appl. 6 (2): 279–89. Дои:10.1111 / j.1752-4571.2012.00288.x. ЧВК 3689353. PMID 23798977.
- ^ Галлардо Дж. А., Нейра Р. (2005). «Экологическая зависимость инбридинговой депрессии у культивируемого кижуча (Oncorhynchus kisutch): агрессивность, доминирование и внутривидовая конкуренция». Наследственность (Edinb). 95 (6): 449–56. Дои:10.1038 / sj.hdy.6800741. PMID 16189545.
- ^ а б Фитцпатрик Дж. Л., Эванс Дж. П. (2014). «Избегание посткопулятивного инбридинга у гуппи» (PDF). J. Evol. Биол. 27 (12): 2585–94. Дои:10.1111 / jeb.12545. PMID 25387854.
- ^ Чарльзуорт Д., Уиллис Дж. Х. (2009). «Генетика инбридинговой депрессии». Nat. Преподобный Жене. 10 (11): 783–96. Дои:10.1038 / nrg2664. PMID 19834483. S2CID 771357.
- ^ Бернштейн Х., Хопф Ф.А., Мишод РЭ (1987). «Молекулярные основы эволюции пола». Молекулярная генетика развития. Adv. Genet. Успехи в генетике. 24. С. 323–70. Дои:10.1016 / с0065-2660 (08) 60012-7. ISBN 9780120176243. PMID 3324702.
- ^ http://www.carp.me.uk В архиве 2011-03-21 на Wayback Machine Информация о нересте карпа
- ^ Леонг, Пол (2004). «Архивная копия». Архивировано из оригинал на 2009-03-02. Получено 2009-03-13.CS1 maint: заархивированная копия как заголовок (ссылка на сайт). Проверено 13 марта, 2009.
- ^ Рейнвейт Флорида и Миллер Э. Дж. (1967) «Ухаживание и репродуктивное поведение сиамских бойцовых рыб, Betta splendens Regan" Слушания Академии наук Оклахомы, Государственный университет Оклахомы.
Дальнейшие ссылки
- Агарвал Н.К. (2008) Размножение рыб Издательство APH. ISBN 9788131303573.
- Бабин П.Дж., Серда Дж. И Любценс Э. (редакторы) (2007) Ооцит рыбы: от фундаментальных исследований до биотехнологического применения Springer. ISBN 9781402062339.
- Кость Кью и Мур Р. (2008) Биология рыб Глава 8: Размножение и истории жизни, стр. 217–255. Тейлор и Фрэнсис. ISBN 9781134186310.
- Cabrita E, Robles V и Paz Herraez P (редакторы) (2008) Методы репродуктивной аквакультуры: морские и пресноводные виды CRC Press. ISBN 9780849380549.
- Коул, Кэтлин Сабина (Эд) (2010) Размножение и сексуальность морских рыб: закономерности и процессы Калифорнийский университет Press. ISBN 9780520264335.
- Хоар WS, Рэндалл DJ и Дональдсон EM (редакторы) (1983) Физиология рыб: Том 9: Воспроизводство Часть A: Эндокринные ткани и гормоны. Академическая пресса. ISBN 9780080585291.
- Хоар WS, Рэндалл DJ и Дональдсон EM (редакторы) (1983) Физиология рыб: Том 9: Воспроизводство Часть B: Поведение и контроль фертильности. Академическая пресса. ISBN 9780080585307.
- Якобсен Т., Фогарти М.Дж., Мегрей Б.А. и Мокснесс Э. (редакторы) (2009) Репродуктивная биология рыб: значение для оценки и управления Джон Вили и сыновья. ISBN 9781444312126.
- Меламед П. и Шервуд Н. (редакторы) (2005) Гормоны и их рецепторы в воспроизводстве рыб World Scientific. ISBN 9789812569189.
- Поттс Г.В., Вуттон Р.Дж. и Вуттон Р.Дж. (редакторы) (1984) Размножение рыб: стратегии и тактика Академическая пресса. ISBN 9780125636605.
- Роча MJ, Arukwe A и Kapoor BG (ред.) (2008) Размножение рыб CRC Press. ISBN 9781578083312.
внешняя ссылка
| Около рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||
| Режимы |
|  |
|---|---|---|
| Удобрение | ||
| Родительская забота | ||
| похожие темы | ||