Пелагическая рыба - Pelagic fish

Пелагическая рыба жить в пелагическая зона воды океана или озера - находясь ни у дна, ни у берега - в отличие от демерсальная рыба которые живут на дне или около него, и рифовая рыба которые связаны с коралловые рифы.[1]
Морской пелагический окружающая среда является крупнейшей водной средой обитания на Земле, занимающей 1370 миллионов кубических километров (330 миллионов кубических миль), и является средой обитания для 11% известных рыбы разновидность. В океаны имеют среднюю глубину 4000 метров. Около 98% общего объема воды находится ниже 100 метров (330 футов), а 75% - ниже 1000 метров (3300 футов).[2]
Морские пелагические рыбы можно разделить на пелагических прибрежных и океанических пелагических рыб.[3] Прибрежная рыба населяют относительно мелкие и залитые солнцем воды над континентальный шельф, в то время как океанические рыбы обитают в обширных и глубоких водах за пределами континентального шельфа (хотя они также могут плавать у берега).[4]
Размеры пелагических рыб варьируются от небольших прибрежных кормовая рыба, Такие как селедка и сардины, к большому хищник вершины океанические рыбы, такие как тунец и океанический акулы.[1] Обычно это ловкие пловцы с обтекаемыми формами тела, способные выдерживать плавание на дальние дистанции. миграции. Многие пелагические рыбы плавают в школы весом в сотни тонн. Другие - одиночки, например, большой океанская солнечная рыба весом более 500 килограммов, которые иногда пассивно дрейфуют с океанскими течениями, поедая медуза.[1]
Эпипелагическая рыба


Эпипелагические рыбы обитают в эпипелагическая зона, самый верхний слой столб воды, начиная с уровень моря до 200 м (660 футов). Его также называют поверхностные воды или солнечная зона, и включает фотическая зона. Фотическая зона определяется как поверхностные воды до глубины, на которую падает солнечный свет. ослабленный до 1% от поверхностной стоимости. Эта глубина зависит от того, насколько мутный вода есть, но может достигать 200 м в чистой воде, совпадая с эпипелагиалией. Фотическая зона пропускает достаточно света для фитопланктон к фотосинтезировать.[5]
Обширная среда обитания большинства пелагических рыб, эпипелагическая зона хорошо освещена, поэтому визуальные хищники могут использовать свое зрение, обычно хорошо перемешана и насыщенный кислородом от воздействия волн и может быть хорошей средой обитания для водоросли расти. Однако это почти безликая среда обитания. Это отсутствие вариации среды обитания приводит к отсутствию видовое разнообразие, поэтому в этой зоне обитает менее 2% известных в мире видов рыб. На большей части зоны не хватает питательных веществ для поддержки рыб, поэтому эпипелагические рыбы, как правило, встречаются в прибрежных водах над уровнем моря. континентальные шельфы, куда сток земли может обеспечить питательными веществами или в тех частях океана, где апвеллинг перемещает питательные вещества в область.[5]
Эпипелагическую рыбу можно условно разделить на мелкую. кормовая рыба и больше хищная рыба которые питаются ими. Кормовая рыба школа и подача фильтра на планктон. Большинство эпипелагических рыб имеют обтекаемые формы тела, способные продолжительное плавание по миграции. В целом хищные и кормовые рыбы имеют одинаковые морфологический Особенности. Рыбы-хищники обычно веретенообразный с большими ртами, гладким телом и глубоко раздвоенным хвостом. Многие используют зрение, чтобы охотиться на зоопланктон или более мелкую рыбу, в то время как другие фильтруют корм на планктоне.

Большинство эпипелагических хищных рыб и их мелкая добыча - это затушеванный с серебристыми цветами, которые уменьшают видимость на рассеяние входящий свет.[5] Серебрение достигается с помощью световозвращающей рыбья чешуя которые функционируют как маленькие зеркала. Это может дать эффект прозрачности. На средних глубинах в море свет идет сверху, поэтому зеркало, которое ориентировано вертикально, делает животных, таких как рыбы, невидимыми сбоку.[6]
В более мелких эпипелагических водах зеркала должны отражать смесь длин волн, и, соответственно, рыба имеет стопки кристаллов с различными расстояниями между ними. Еще одна сложность для рыб с закругленными в поперечном сечении телами заключается в том, что зеркала будут неэффективными, если положить их на кожу, поскольку они не будут отражать горизонтально. Общий зеркальный эффект достигается с помощью множества небольших отражателей, все ориентированных вертикально.[6]
Хотя количество видов ограничено, эпипелагические рыбы многочисленны. Недостаток разнообразия компенсируется числом. Кормовая рыба встречается в огромных количествах, и большая рыба, которая охотится на нее, часто становится популярной. пищевая рыба. Как группа, эпипелагические рыбы составляют наиболее ценные рыболовство в мире.[5]
Многие кормовые рыбы являются факультативными хищниками, которые могут выбирать отдельные копеподы или личинок рыб из толщи воды, а затем перейти на фильтрацию, питающуюся фитопланктон когда энергетически это дает лучшие результаты. Фильтр кормления рыб обычно используют длинный мелкий жаберные тычинки для удаления мелких организмов из водной толщи. Некоторые из самых крупных эпипелагических рыб, такие как китовая акула и Китовая акула, являются фильтрующими питателями, а также некоторые из самых маленьких, например, взрослых шпроты и анчоусы.[7]
Исключительно прозрачные воды океана содержат мало пищи. Области высокой продуктивности имеют тенденцию быть несколько мутными от планктон цветет. Они привлекают фильтрующих поедателей планктона, которые, в свою очередь, привлекают более высоких хищников. Ловля тунца имеет тенденцию быть оптимальной, когда мутность воды, измеряемая максимальной глубиной a диск секки видно в солнечный день, от 15 до 35 метров.[8]
Плавающие объекты




Эпипелагические рыбы очарованы плавучими объектами. Они собираются в значительном количестве вокруг таких объектов, как дрейфующие обломки, плоты, медузы и плавающие водоросли. Кажется, что объекты создают «визуальный стимул в оптической пустоте».[9] Плавучие объекты могут предлагать убежище за молодь от хищников. Обилие дрейфующих морских водорослей или медуз может привести к значительному увеличению выживаемости некоторых молодых видов.[10]
Многие прибрежные молодые особи используют водоросли в качестве убежища и пищи, которую получают от беспозвоночных и других связанных с ними рыб. Дрейфующие водоросли, особенно пелагические Саргассум, обеспечивают нишевую среду обитания с собственным убежищем и едой и даже поддерживают свою уникальную фауну, такую как саргассовая рыба.[7] Одно исследование, проведенное недалеко от Флориды, обнаружило 54 вида из 23 семей, живущих на обломках из Саргассум коврики.[11] Медузы также используются молодыми рыбами в качестве укрытия и пищи, хотя медузы могут охотиться на мелкую рыбу.[12]
Мобильные океанические виды, такие как тунец можно запечатлеть, путешествуя на большие расстояния в больших рыболовные суда. Более простая альтернатива - использовать очарование рыбы плавающими объектами. Когда рыбаки используют такие предметы, их называют агрегаты для сбора рыбы (ФАДы). FAD - это закрепленные на якоре плоты или объекты любого типа, плавающие на поверхности или непосредственно под ней. Рыбаки в Тихом и Индийском океанах устанавливают плавучие FAD, собранные из разного рода обломков, вокруг тропических островов, а затем используют кошелек поймать привлеченную к ним рыбу.[13]
Исследование с использованием сонар во Французской Полинезии обнаружены крупные косяки молоди большеглазый тунец и желтоперый тунец агрегированные ближайшие к приборам, от 10 до 50 м. Дальше, от 50 до 150 м, была менее плотная группа более крупных желтоперых и альбакор тунец. А еще дальше, на 500 м, рассредоточилась группа различных крупных взрослых тунцов. Распределение и плотность этих групп были непостоянными и совпадающими. FAD также использовались другими рыбами, и скопления разошлись, когда было темно.[14]
Более крупная рыба, даже хищная рыба, такая как большая барракуда, часто привлекают кучу мелких рыб, которые их сопровождают стратегически безопасным образом. Skindivers которые остаются в воде на длительное время, также часто привлекают свиту рыб: более мелкие рыбы подходят близко, а более крупные рыбы наблюдают с большего расстояния. Морские черепахи, служащий передвижным укрытием для мелкой рыбы, может быть случайно пронзен мечом, пытающимся поймать рыбу.[15]
Прибрежная рыба

Прибрежная рыба (также называемый неритический или прибрежные рыбы) обитают в водах около морской берег и выше континентальный шельф. Поскольку континентальный шельф обычно имеет глубину менее 200 метров, из этого следует, что прибрежные рыбы, не являющиеся донными рыбами, обычно являются эпипелагическими рыбами, обитающими в освещенной солнцем эпипелагиали.[2]
Прибрежные эпипелагические рыбы - одни из самых распространенных в мире. К ним относятся кормовые рыбы, а также хищные рыбы, которые ими питаются. Кормовая рыба прекрасно себя чувствует в прибрежных водах, где высокая продуктивность является результатом апвеллинга и выноса питательных веществ с береговой линии. Некоторые из них являются частичными жителями, которые нерестятся в ручьях, устьях и заливах, но большинство завершают свой жизненный цикл в этой зоне.[7]
Океаническая рыба

Океанические рыбы (также называемые рыбами открытого океана или прибрежными водами) обитают в водах, не находящихся выше континентального шельфа. Океаническую рыбу можно противопоставить прибрежная рыба, которые живут над континентальным шельфом. Однако эти два типа не исключают друг друга, поскольку между прибрежными и океанскими регионами нет четких границ, и многие эпипелагические рыбы перемещаются между прибрежными и океаническими водами, особенно на разных стадиях своего жизненного цикла.[7]
Океанические эпипелагические рыбы могут быть настоящими жителями, частичными жителями или случайными жителями. Настоящие жители всю жизнь живут в открытом океане. Только несколько видов являются настоящими жителями, например тунец, морская рыба, летучая рыба, сайры, лоцман, Реморас, дельфин, океанские акулы и океанская солнечная рыба. Большинство этих видов мигрируют взад и вперед через открытые океаны, редко пересекая континентальные шельфы. Некоторые настоящие жители ассоциируются с дрейфующими медузами или водорослями.[7]
Частичные жители делятся на три группы: виды, обитающие в зоне только в молодом возрасте (плывущие с медузами и водорослями); виды, обитающие в зоне только взрослыми особями (лосось, летучая рыба, дельфины и китовые акулы); и глубоководные виды, которые совершают ночные миграции в поверхностные воды (например, рыба-фонарь ).[7] Случайные поселения иногда возникают, когда взрослые особи и молодые особи видов из других сред случайно переносятся в зону течениями.[7]
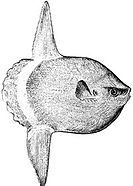
Огромный океанская солнечная рыба, настоящий житель эпипелагиали океана, иногда плывет по течению, питаясь медуза

Гигант Китовая акула, еще один житель эпипелагиали океана, фильтр питается планктон, и периодически ныряет вглубь мезопелагиали.

Фонарь частичные жители эпипелагиали океана, днем прячутся в глубоких водах, а ночью мигрируют к поверхностным водам для пропитания



Глубоководная рыба

В глубоком океане воды простираются далеко ниже эпипелагиали и поддерживают самые разные типы пелагических рыб, приспособленных к жизни в этих более глубоких зонах.[2]
В глубокой воде, морской снег это непрерывный поток органических детрит падение из верхних слоев водной толщи. Его происхождение лежит в деятельности внутри производственной световая зона. Морской снег включает мертвые или умирающие планктон, протисты (диатомеи ), фекалии, песок, сажа и прочая неорганическая пыль. «Снежинки» со временем растут и могут достигать нескольких сантиметров в диаметре, путешествуя неделями, прежде чем достичь дна океана. Однако большинство органических компонентов морского снега потребляются микробы, зоопланктон, и другие фильтрующие животные в пределах первых 1000 метров пути, то есть в пределах эпипелагической зоны. Таким образом, морской снег можно считать основой глубоководных мезопелагический и бентосный экосистемы: Поскольку солнечный свет не может достичь их, глубоководные организмы в значительной степени полагаются на морской снег как источник энергии.
Некоторые глубоководные пелагические группы, такие как рыба-фонарь, риджхед, морской топорик, и светлая рыба семьи иногда называют псевдокеанский потому что вместо того, чтобы иметь равномерное распределение в открытой воде, они встречаются в значительно большей численности вокруг структурных оазисов, особенно подводные горы, и более континентальные склоны. Это явление объясняется также обилием видов жертв, которые также привлекаются к структурам.
Рыбы в различных пелагических и глубоководных донных зонах имеют физическое строение и поведение, которое заметно отличается друг от друга. Группы сосуществующих видов в каждой зоне, похоже, действуют одинаковым образом, например, небольшой мезопелагический вертикально мигрирующий планктоноеды, батипелагические удильщик, и глубоководные бентосные гремучие хвостики.[16]
Рэй с плавниками виды с колючими плавниками редки среди глубоководных рыб, что говорит о том, что глубоководные рыбы являются древними и настолько хорошо адаптированы к окружающей среде, что вторжения более современных рыб оказались безуспешными.[17] Те немногие лучевые плавники, которые действительно существуют, в основном находятся в Beryciformes и Lampriformes, которые также являются древними формами. Большинство глубоководных пелагических рыб принадлежат к собственным отрядам, что предполагает длительную эволюцию в глубоководных условиях. Напротив, глубоководные бентосные виды находятся в отрядах, в которые входят многие родственные мелководные рыбы.[18]
Многие виды ежедневно перемещаются между зонами вертикальными миграциями. В следующей таблице они перечислены в средней или более глубокой зоне, где они регулярно встречаются.
| Зона | Виды и видовые группы включают: |
|---|---|
| Эпипелагический[5] | |
| Мезопелагический | Фонарь, опа, длинноносый ланцет, ячмень, риджхед, саблезуб, свободный стоп-сигнал, морской топорик[19] |
| Батипелагический | В основном щетинистый рот и удильщик. Также клыкастый зуб, гадюка, черный глотатель, телескопрыба, молоток, кинжал, барракудина, черные ножны, бобтейл бекас угорь, единорог, угорь, дряблый кит. |
| Бентопелагический[5] | Крысиный хвост и Brotula особенно многочисленны. |
| Бентосный | Камбала, миксина, бельдюга, зеленый глаз угорь, морской скат, пинагой, и летучая мышь[5] |
| Эпипелагический | Мезопелагический | Батипелагический | Глубокое море бентосный | |
|---|---|---|---|---|
| мышцы | мускулистые тела | слабо развитый, дряблый | ||
| скелет | крепкие окостеневшие кости | слабая, минимальная оссификация | ||
| напольные весы | да | никто | ||
| нервная система | хорошо развитый | только боковая линия и обоняние | ||
| глаза | большой и чувствительный | маленький и может не работать | переменная (от хорошо развитой до отсутствующей) | |
| фотофоры | отсутствующий | общий | общий | обычно отсутствует |
| жабры | хорошо развитый | |||
| почки | большой | маленький | ||
| сердце | большой | маленький | ||
| плавательный пузырь | у вертикально мигрирующих рыб есть плавательный пузырь | сокращен или отсутствует | переменная (от хорошо развитой до отсутствующей) | |
| размер | обычно меньше 25 см | переменная, виды крупнее одного метра не редкость |
Мезопелагическая рыба


Ниже эпипелагиали условия быстро меняются. На расстоянии от 200 метров до примерно 1000 метров свет продолжает тускнеть, пока почти не исчезает. Температура падает через термоклин до температуры от 4 ° C (39 ° F) до 8 ° C (46 ° F). Это сумерки или же мезопелагический зона. Давление продолжает расти со скоростью одна атмосфера на каждые 10 метров, в то время как концентрации питательных веществ падают вместе с растворенным кислородом и скоростью, с которой циркулирует вода.[2][20]
Операторы гидролокатора, использующие технологию гидролокатора, разработанную во время Второй мировой войны, были озадачены тем, что казалось ложным морским дном глубиной 300–500 метров днем и менее глубоким ночью. Оказалось, что это произошло из-за миллионов морских организмов, в первую очередь мелких мезопелагических рыб, плавательные пузыри которых отражают сонар. Эти организмы мигрируют в более мелкие воды с наступлением сумерек, чтобы питаться планктоном. Слой более глубокий, когда нет луны, и может стать мельче, когда облака проходят над луной. Это явление получило название глубокий рассеивающий слой.[21]
Большинство мезопелагических рыб производят ежедневно вертикальные миграции, перемещаясь ночью в эпипелагиали, часто следуя аналогичным миграциям зоопланктона, а днем возвращаясь в глубину в целях безопасности.[20][2][22] Эти вертикальные миграции часто происходят на большие вертикальные расстояния и выполняются с помощью плавательный пузырь. Плавательный пузырь надувается, когда рыба хочет подняться, и, учитывая высокое давление в мессоплегической зоне, это требует значительных затрат энергии. По мере того, как рыба поднимается, давление в плавательном пузыре должно регулироваться, чтобы он не лопнул. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается.[23] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин, где температура колеблется от 10 до 20 ° C, что обеспечивает значительные допуски на изменение температуры.[24]
У этих рыб есть мускулистые тела, окостеневшие кости, чешуя, хорошо развитые жабры и центральная нервная система, а также большие сердца и почки. Мезопелагический кормушки планктона иметь маленькие рты с прекрасным жаберные тычинки, в то время как рыбоядные имеют более крупный рот и более грубые жаберные тычинки.[20][2] У вертикально мигрирующих рыб плавательные пузыри.[17]
Мезопелагические рыбы приспособлены к активной жизни в условиях низкой освещенности. Большинство из них - зрительные хищники с большими глазами. У некоторых более глубоководных рыб глаза трубчатые с большими линзами и только стержневые клетки которые смотрят вверх. Они обеспечивают бинокулярное зрение и большую чувствительность к слабым световым сигналам.[2] Эта адаптация обеспечивает улучшенное терминальное зрение за счет бокового зрения и позволяет хищнику различать Кальмар, каракатица, и более мелкие рыбы, которые вырисовываются на фоне мрака над ними.[20]
Мезопелагические рыбы обычно не имеют защитных шипов и используют цвет, чтобы камуфляж их от других рыб.[20] Засадные хищники темные, черные или красные. Поскольку более длинные красные волны света не достигают глубины моря, красный эффективно действует так же, как черный. Использование миграционных форм затушеванный серебристые цвета. На животе они часто показывают фотофоры производит низкокачественный свет. Для хищника снизу, смотрящего вверх, это биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют (недостаточный красный) окружающий свет, оставляя биолюминесценцию видимой.[25]

В Антарктический клыкач имеют большие, направленные вверх глаза, приспособленные для распознавания силуэтов хищных рыб.[26]

В свободный стоп-сигнал имеет нижняя челюсть четверть длины его тела. Челюсть не имеет пола и крепится только с помощью шарнира и модифицированной кости языка. За большими клыковидными зубами впереди следует множество мелких зубцов с зазубринами.[28][29]

В свободный стоп-сигнал также одна из немногих рыб, производящих красный биолюминесценция. Поскольку большая часть их добычи не может воспринимать красный свет, это позволяет им охотиться с помощью практически невидимого луча света.[28]




В коричневая рыба-призрак это разновидность ячмень и это единственное известное позвоночное животное, которое использует зеркало, а не линзу, чтобы сфокусировать изображение в своих глазах.[30][31]
Отбор проб глубоким траление указывает, что рыба-фонарь составляют до 65% всей глубоководной рыбы биомасса.[32] В самом деле, фонарики являются одними из самых распространенных, густонаселенных и разнообразных из всех. позвоночные, играя важную экологический роль добычи для более крупных организмов. По оценкам, глобальная биомасса фонаря составляет 550–660 миллионов человек. метрические тонны, в несколько раз выловили весь мировой промысел. Фонарь также составляет большую часть биомассы, ответственной за глубокий рассеивающий слой Мирового океана. Сонар отражается от миллионов фонарных рыб плавательные пузыри, создавая видимость ложного дна.[33]
Большеглазый тунец - это эпипелагический / мезопелагический вид, который является плотоядным и ест другую рыбу. Спутниковое мечение показало, что большеглазый тунец часто проводит длительные периоды, путешествуя глубоко под поверхностью в дневное время, иногда ныряя на глубину до 500 метров. Считается, что эти движения являются ответом на вертикальные миграции организмов-жертв в глубокий рассеивающий слой.

Длинноносый ланцет. Lancetfish - хищники из засад, часто встречающиеся в мезопелагической зоне. Это одни из самых крупных мезопелагических рыб (до 2 метров).[34]

В телескопрыба имеет большие, направленные вперед телескопические глаза с большими линзами.[35]

Большеглазый тунец круиз по эпипелагиали ночью и мезопелагиали днем.

Коллекция мезопелагических кормовых рыб, промысловых из Мексиканского залива, включая Миктофиды, личинка удильщик, щетинистый рот, а барракудина




Батипелагическая рыба


Ниже мезопелагиали кромешная тьма. Это полночь или же батипелагическая зона, простирающаяся от 1000 м до дна глубокой воды бентосная зона. Если вода очень глубокая, пелагиали ниже 4000 м иногда называют нижняя полночь или же абиссопелагическая зона.
Условия в этих зонах примерно одинаковы, темнота полная, давление сокрушительное, а температура, питательные вещества и уровень растворенного кислорода низкие.[2]
У батипелагических рыб есть особые приспособления чтобы справиться с этими условиями - у них есть медленные метаболизм и неспециализированные диеты, готовность есть все, что попадается под руку. Они предпочитают сидеть и ждать еды, а не тратить энергию на ее поиски. Поведение батипелагических рыб можно противопоставить поведению мезопелагических рыб. Мезопелагические рыбы часто очень подвижны, в то время как батипелагические почти все хищники поджидают нас, обычно тратя мало энергии на движение.[41]
Доминирующие батипелагические рыбы - мелкие. щетинистый рот и удильщик; клыкастый зуб, гадюка, кинжал, и барракудина также распространены. Эти рыбки маленькие, многие около 10 сантиметров в длину и немногие длиннее 25 см. Они проводят большую часть своего времени, терпеливо ожидая в толще воды, пока не появится добыча или не соблазнится их люминофором. То немногое энергии, которое доступно в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и случайных беспозвоночных или мезопелагических рыб.[41] Около 20% пищи, происходящей из эпипелагиали, приходится на мезопелагиали,[21] но только около 5% попадает в батипелагическую зону.[37]
Батипелагические рыбы ведут оседлый образ жизни, приспособлены к выработке минимальной энергии в среде обитания с очень небольшим количеством пищи или доступной энергии, даже без солнечного света, только с биолюминесценцией. Их тела удлиненный со слабыми водянистыми мышцами и скелетный конструкции. Так как большая часть рыбы состоит из воды, они не сжимаются на таких глубинах. Часто имеют раздвижные, навесные челюсти с загнутыми зубами. Они слизистые, без напольные весы. Центральная нервная система ограничена боковой линией и обонятельной системой, глаза маленькие и могут не функционировать, и жабры, почки и сердце, и плавательные пузыри маленькие или отсутствуют.[37][42]
Это те же особенности, что и у рыб личинки, что предполагает, что в процессе эволюции батипелагические рыбы приобрели эти особенности благодаря неотения. Как и в случае с личинками, эти особенности позволяют рыбе оставаться во взвешенном состоянии в воде с небольшими затратами энергии.[43]
Несмотря на свой свирепый вид, эти глубинные твари в основном представляют собой миниатюрные рыбы со слабыми мускулами и слишком малы, чтобы представлять какую-либо угрозу для людей.
Плавательные пузыри глубоководных рыб либо отсутствуют, либо практически не функционируют, а батипелагические рыбы обычно не совершают вертикальных миграций. Заполнение баллонов при таком большом давлении требует огромных затрат энергии. У некоторых глубоководных рыб есть плавательные пузыри, которые функционируют в молодом возрасте и населяют верхнюю эпипелагическую зону, но они увядают или наполняются жиром, когда рыба перемещается в свою взрослую среду обитания.[44]
Наиболее важными сенсорными системами обычно являются внутреннее ухо, который реагирует на звук, а боковая линия, который реагирует на изменение давления воды. В обонятельный Система также может быть важна для мужчин, которые находят самок по запаху.[45]Батипелагические рыбы черные, а иногда и красные, с небольшими фотофоры. Когда фотофоры используются, они обычно используются для того, чтобы заманить добычу или привлечь партнера. Из-за нехватки пищи батипелагические хищники не избирательны в своих привычках кормления, а хватают все, что подойдет достаточно близко. Они достигают этого за счет большого рта с острыми зубами, чтобы хватать крупную добычу и перекрывать друг друга. жаберные тычинки которые не позволяют маленькой добыче, проглоченной, ускользнуть.[42]
Найти себе пару в этой зоне непросто. Некоторые виды зависят от биолюминесценция. Другие гермафродиты, что удваивает их шансы произвести яйцеклетки и сперму при встрече.[37] Самка удильщика выпускает феромоны для привлечения крошечных самцов. Когда мужчина находит ее, он кусает ее и никогда не отпускает. Когда самец удильщиков Гаплофрин моллис впивается в кожу самки, он выпускает фермент который переваривает кожу его рта и ее тело, сплавляя пару до точки соединения двух кровеносных систем. Затем самец атрофируется, превращаясь в пару гонады. Этот крайний половой диморфизм гарантирует, что когда самка будет готова к нересту, у нее сразу же появится помощник.[46]
В батипелагической зоне обитают многие виды животных, помимо рыб, такие как кальмары, большие киты, осьминоги, губки, брахиоподы, морские звезды и ехиноиды, но в этой зоне рыбам трудно жить.

В угорь использует свой рот как сеть, открывая свой большой рот и плывя к своей добыче. На конце хвоста у него есть люминесцентный орган, привлекающий добычу.

В черный глотатель, с расширяющимся желудком, отличается способностью глотать целиком, костлявые рыбы в десять раз больше его массы.[47][48]

женский Гаплофрин моллис удильщик, плывущий за прикрепленными самцами, атрофировавшимися в пару гонады, для использования, когда самка готова к нересту.

Широко распространенный клыкастый зуб имеет самые большие зубы среди всех рыб, пропорциональные размеру тела.[49] Несмотря на свирепый вид, батипелагические рыбы обычно имеют слабую мускулатуру и слишком малы, чтобы представлять какую-либо угрозу для человека.

В Гадюка Слоана могут совершать ночные миграции из батипелагических глубин в приповерхностные воды.[50]





Демерсальная рыба

Демерсальная рыба живут на дне моря или рядом с ним.[51] Придонные рыбы водятся морское дно в прибрежных районах на континентальный шельф, а в открытом океане они встречаются по внешнему континентальная окраина на континентальном склоне и на континентальном подъеме. Обычно они не встречаются в абиссопелагический или же гадопелагический глубины или на бездонная равнина. Они занимают дно моря, состоящее из ила, песка, гравия или скал.[51]
В глубоководных районах рыбы демерсальной зоны активны и относительно многочисленны по сравнению с рыбами из глубин. батипелагическая зона.[41]
Гремучие и бротулы обычны, и другие устойчивые семьи угри, бельдюги, микробы, зеленые глаза, летучие мыши, и шишки.[42]
Глубоководные тела бентосный рыбы мускулистые, с хорошо развитыми органами. В этом отношении они ближе к мезопелагическим рыбам, чем к батопелагическим рыбам. В остальном они более вариативны. Фотофоры обычно отсутствуют, глаза и плавательные пузыри варьируются от отсутствующих до хорошо развитых. Они различаются по размеру, нередки более крупные виды более одного метра.
Глубоководные донные рыбы обычно длинные и узкие. Многие из них имеют форму угрей или имеют форму угрей. Это может быть потому, что длинные тела давно боковые линии. Боковые линии улавливают низкочастотные звуки, а у некоторых донных рыб, кажется, есть мышцы, которые барабанят такие звуки, чтобы привлечь партнеров.[17] Обоняние также важно, на что указывает скорость, с которой донные рыбы находят ловушки с наживкой. приманка для рыбы.
Основной рацион глубоководных донных рыб - это глубоководные беспозвоночные. бентос и падаль. Обоняние, осязание и чувствительность по боковой линии, кажется, являются основными сенсорными устройствами для их определения.[52]
Глубоководные донные рыбы можно разделить на строго донных рыб и двунопелагических рыб. Обычно бентосные рыбы обладают отрицательной плавучестью, в то время как бентопелагические рыбы обладают нейтральной плавучестью. Строго донные рыбы постоянно контактируют с дном. Они либо лгут и ждут, как устраивать засады на хищников или активно перемещайтесь по дну в поисках еды.[52]


Бентопелагическая рыба
Бентопелагические рыбы обитают в воде чуть выше дна, питаясь бентос и бентопелагический зоопланктон.[53] Большинство дермерсальных рыб двунопелагические.[51]
Их можно разделить на дряблое и крепкое телосложение. Дряблые двупелагические рыбы похожи на батопелагических рыб, у них уменьшенная масса тела и низкий уровень метаболизма, тратя минимум энергии, пока они лежат и ждут засада добыча.[54] Пример дряблой рыбы - кусковой угорь. Acanthonus armatus,[55] хищник с огромной головой и телом, состоящим на 90% из воды. У этой рыбы самые большие уши (отолиты ) и самый маленький мозг по сравнению с размером тела среди всех известных позвоночных.[56]
Крепкие бентопелагические рыбы - мускулистые пловцы, которые активно курсируют по дну в поисках добычи. Они могут жить вокруг таких функций, как подводные горы, которые имеют сильные токи.[56] Примерами являются оранжевый грубый и Патагонский клыкач. Поскольку эти рыбы когда-то были в изобилии и их крепкие тела годятся для употребления в пищу, их вылавливают в коммерческих целях.[57][58]
Бентическая рыба
Бентические рыбы не являются пелагическими рыбами, но они кратко обсуждаются здесь для полноты и контраста.
Некоторые рыбы не попадают в приведенную выше классификацию. Например, семья почти слепых рыбы-пауки, обычные и широко распространенные, питаются бентопелагическим зоопланктоном. Тем не менее, это строго донные рыбы, так как они остаются в контакте со дном. Их плавники имеют длинные лучи, которые они используют, чтобы «стоять» на дне, когда они смотрят на поток, и захватывают зоопланктон, когда он проходит мимо.[59]
Самая глубоководная из известных рыб, строго донная Abyssobrotula galatheae, угревидный и слепой, питается донными беспозвоночными.[60][61]

Тихоокеанская миксина отдыхая на дне. Хагфиш покрывает себя и любую мертвую рыбу ядовитой слизью, которая делает их несъедобными для других видов ".

Рыба-тренога (Батиптероевый граллатор ), вид паучьей рыбы, использует свои плавники, чтобы «стоять» на дне.[62]

В пятнистый луч веернохвоста Питается донной рыбой, двустворчатыми моллюсками, крабами и креветками.[63]



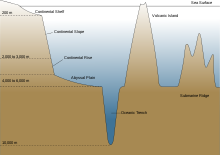
На больших глубинах нехватка пищи и экстремальное давление ограничивают выживаемость рыбы. Самая глубокая точка океана составляет около 11000 метров. Батипелагические рыбы обычно не встречаются ниже 3000 метров. Наибольшая зафиксированная глубина для придонной рыбы составляет 8 370 м.[64] Возможно, чрезмерное давление нарушает основные функции ферментов.[37]
Бентические рыбы более разнообразны и, вероятно, встречаются на континентальный склон, где есть разнообразие местообитаний и часто запасы пищи. Примерно 40% дна океана состоит из абиссальные равнины, но эти плоские безликие области покрыты осадок и в основном лишены бентоса (бентос ). Глубоководные донные рыбы чаще ассоциируются с каньонами или выходами скал на равнинах, где обосновались сообщества беспозвоночных. Подводные горы (подводные горы ) может перехватывать глубоководные течения и вызывать продуктивные апвеллинги, поддерживающие донную рыбу. Подводные горные хребты могут разделять подводные регионы на разные экосистемы.[16]
Пелагическое рыболовство
Кормовая рыба
| ||||||||||||||||||||||||||||||||||||||||||||||||
Основные морские промыслы в дикой природе |
Мелкие пелагические рыбы обычно кормовая рыба на которые охотятся более крупные пелагические рыбы и другие хищники. Кормовая рыба подача фильтра на планктон и обычно имеют длину менее 10 сантиметров. Они часто остаются вместе в школы и может мигрировать большие расстояния между нерестилищами и местами нагула. Особенно они встречаются в апвеллинг регионы вокруг северо-восточной Атлантики, у побережья Японии и у западных берегов Африки и Америки. Кормовые рыбы, как правило, недолговечны, и их акции заметно колеблются с годами.[65]
сельдь находятся в Северное море и Североатлантический на глубинах до 200 метров. Важные промыслы сельди существовали в этих районах на протяжении веков. Сельдь разного размера и скорости роста принадлежит к разным популяциям, каждая из которых имеет свои пути миграции. При нересте самка откладывает от 20 000 до 50 000 яиц. После нереста сельдь теряет жир и возвращается в районы нагула, богатые планктоном.[66] В районе Исландии традиционно вылавливали три отдельные популяции сельди. Эти запасы рухнули в конце 1960-х годов, хотя с тех пор два из них восстановились. После распада Исландия превратилась в мойва, которые в настоящее время составляют около половины общего улова Исландии.[67]
Путассу находятся в открытом океане и над континентальный склон на глубинах от 100 до 1000 метров. Они следят за вертикальными миграциями зоопланктон Днем они питаются со дна, а ночью - на поверхность.[66][68]
Традиционное рыболовство для анчоусы и сардины также работали в Тихом океане, Средиземном море и юго-восточной Атлантике.[69] Мировой годовой улов кормовой рыбы в последние годы составлял примерно 22 миллиона тонн, или четверть всего мирового улова.

Эти учеба в школе Тихоокеанские сардины находятся кормовая рыба

Перуанская анчовета





Рыба-хищник
Пелагические рыбы среднего размера включают: Trevally, барракуда, летучая рыба, бонито, Махи Махи, и прибрежная скумбрия.[1] Многие из этих рыб охотятся на кормовых рыб, но на них, в свою очередь, охотятся еще более крупные пелагические рыбы. Почти вся рыба в какой-то мере является хищной рыбой, и, за исключением высших хищников, различие между хищной рыбой и хищной или кормовой рыбой является несколько искусственным.[70]
По всей Европе есть три популяции прибрежных скумбрия. Одна популяция мигрирует в Северное море, другая остается в водах ирландское море, а третья популяция мигрирует на юг вдоль западного побережья Шотландии и Ирландии. Крейсерская скорость скумбрии составляет впечатляющие 10 километров в час.[66][71]
Многие крупные пелагические рыбы - это океанические кочевые виды, которые совершают длительные миграции в море. Питаются мелкой пелагической кормовой рыбой, а также пелагической рыбой среднего размера. Иногда они следуют за своей стайной добычей, и многие виды сами образуют стайки.
Примеры более крупных пелагических рыб: тунец, морская рыба, королевская макрель, акулы и большие лучи.
В частности, тунец имеет большое значение для коммерческого рыболовства. Хотя тунцы мигрируют через океаны, попытки их найти не являются обычным подходом. Тунцы, как правило, собираются в районах с обильным кормом, вдоль границ течений, вокруг островов, у подводных гор и в некоторых районах апвеллинга вдоль континентальных склонов. Ловят тунца несколькими способами: кошельковые невода оградить всю наземную школу специальными сетками, полюса и линейные суда которые используют удочки с наживкой с другой мелкой пелагической рыбой в качестве приманка, и плоты назывались агрегаты для сбора рыбы создаются, потому что тунец, как и некоторые другие пелагические рыбы, имеют тенденцию скапливаться под плавучими объектами.[1]
Другие крупные пелагические рыбы занимают первое место дичь, особенно марлин и рыба-меч.

Желтоперый тунец вылавливаются в качестве замены ныне в значительной степени истощенных Южный голубой тунец

Королевская макрель круиз на дальние перелеты со скоростью 10 километров в час [66][71]






Продуктивность
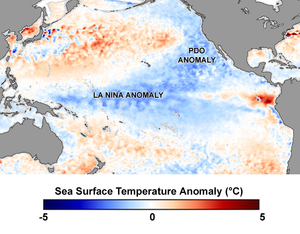
Апвеллинг происходит как вдоль береговых линий, так и в середине океана при столкновении глубоких Океанские течения выводит на поверхность холодную воду, богатую питательными веществами. Эти апвеллинги поддерживают цветение фитопланктона, который, в свою очередь, производит зоопланктон и поддерживает многие из основных промыслов в мире. Если апвеллинг не удастся, значит, промысел в этом районе не состоится.[13]
В 1960-е гг. Перуанская анчовета рыболовство было крупнейшим промыслом в мире. Популяция анчоусов значительно сократилась в 1972 г. Эль-Ниньо событие, когда теплая вода течет по холодной Гумбольдтовское течение, как часть 50-летнего цикла, снижая глубину термоклин. Апвеллинг остановился и фитопланктон производство резко упало, как и популяция анчоусов, и миллионы морские птицы, зависимый от анчоуса, погиб.[72] С середины 1980-х годов апвеллинг возобновился, и уровни вылова перуанской анчоусы вернулись к уровням 1960-х годов.
У берегов Японии столкновение Оясио Текущий с Куросио Текущий производит богатые питательными веществами апвеллинги. Циклическое изменение этих токов привело к снижению сардина Sardinops melanosticta населения. Уловы рыболовства снизились с 5 миллионов тонн в 1988 году до 280 тысяч тонн в 1998 году. Тихоокеанский голубой тунец перестал переезжать в регион кормиться.[73][74]
Океанские течения могут влиять на распределение рыб, как концентрируя, так и рассеивая их. Соседние океанические течения могут определять четкие границы, если они меняются. Эти границы могут быть даже видимыми, но обычно их присутствие отмечается быстрыми изменениями солености, температуры и мутности.[13]
Например, в азиатской северной части Тихого океана альбакор заключены между двумя текущими системами. Северная граница определяется холодным Северо-Тихоокеанское течение а южная граница определяется Северное экваториальное течение. Чтобы усложнить ситуацию, их распределение далее изменяется в пределах области, определяемой двумя текущими системами, другим течением, Куросио Текущий, потоки которых изменяются сезонно.[75]
Часто эпипелагические рыбы порождать в районе, где яйца и личинки дрейфуют вниз по течению в подходящие места кормления и, в конечном итоге, попадают в районы кормления взрослых особей.[13]
Острова и банки могут взаимодействовать с течениями и апвеллингами, что приводит к образованию районов с высокой продуктивностью океана. Большие водовороты могут образовывать нисходящие или подветренные потоки от островов, концентрируя планктон.[76] Берега и рифы могут перехватывать восходящие течения.[13]
Далеко мигрирующие виды

Эпипелагические рыбы обычно перемещаются на большие расстояния между районами нагула и нереста или в ответ на изменения в океане. Крупные океанические хищники, такие как лосось и тунец, могут мигрировать на тысячи километров, пересекая океаны.[78]
В исследовании 2001 г. движение Атлантический голубой тунец из области за пределами Северной Каролины были изучены с помощью специальных всплывающих тегов. Прикрепленные к тунцу, эти метки отслеживали движения тунца около года, затем отделялись и всплывали на поверхность, где передавали свою информацию на спутник. Исследование показало, что у тунца было четыре разных способа миграции. Одна группа на год прижилась к западной части Атлантического океана. Другая группа также оставалась в основном в западной части Атлантического океана, но мигрировала в Мексиканский залив для нереста. Третья группа пересекла Атлантический океан и обратно. Четвертая группа перешла в восточную Атлантику, а затем перебралась на нерест в Средиземное море. Исследование показывает, что, несмотря на некоторую дифференциацию по нерестилищам, по сути, существует только одна популяция атлантического синего тунца, смешанные группы, которые используют всю северную часть Атлантического океана, Мексиканский залив и Средиземное море.[79]
Период, термин далеко мигрирующие виды (HMS) - это юридический термин, который берет свое начало в статье 64 Конвенция Организации Объединенных Наций по морскому праву (ЮНКЛОС).[80]
К далеко мигрирующим видам относятся: тунец и тунцеобразные виды (альбакор, Атлантический голубой тунец, большеглазый тунец, скипджек, желтоперый, черный плавник, маленький тунец, Тихоокеанский голубой плавник, южный голубой плавник и пуля ), помфрет, марлин, парусник, рыба-меч, сайра и океан акулы, а также млекопитающих, таких как дельфины, и другие китообразные.
По сути, далеко мигрирующие виды совпадают с более крупными из «крупных пелагических рыб», обсуждавшихся в предыдущем разделе, если добавлены китообразные и некоторые коммерчески неважные рыбы, такие как солнечная рыба, исключены. Это высокие трофический уровень виды, которые совершают миграции на значительные, но изменчивые расстояния через океаны для кормления, часто кормовой рыбой или размножения, а также имеют широкое географическое распространение. Таким образом, эти виды встречаются в пределах 200 морских миль (370 км). исключительные экономические зоны и в открытое море вне этих зон. Они есть пелагический видов, что означает, что они в основном живут в открытом океане и не живут у морского дна, хотя они могут проводить часть своего жизненного цикла в прибрежные воды.[81]
Производство захвата
Согласно Продовольственная и сельскохозяйственная организация (ФАО), мировой урожай в 2005 г. состояла из 93,2 млн. тонны захвачен коммерческое рыболовство в дикий рыбный промысел.[82] Из этого общего количества около 45% составляли пелагические рыбы. В следующей таблице показано мировое производство отлова в тонны.[83]
| Производство улова по группам видов в тоннах | ||||||||
|---|---|---|---|---|---|---|---|---|
| Тип | Группа | 1999 | 2000 | 2001 | 2002 | 2003 | 2004 | 2005 |
| Мелкая пелагическая рыба | Сельдь, сардины, анчоусы | 22 671 427 | 24 919 239 | 20 640 734 | 22 289 332 | 18 840 389 | 23 047 541 | 22 404 769 |
| Крупная пелагическая рыба | Тунцы, бонито, рыбки | 5 943 593 | 5 816 647 | 5 782 841 | 6 138 999 | 6 197 087 | 6 160 868 | 6 243 122 |
| Другие пелагические рыбы | 10 712 994 | 10 654 041 | 12 332 170 | 11 772 320 | 11 525 390 | 11 181 871 | 11 179 641 | |
| Хрящевые рыбы | Акулы, лучи, химеры | 858 007 | 870 455 | 845 854 | 845 820 | 880 785 | 819 012 | 771 105 |
Угрожаемые виды
В 2009 г. Международный союз охраны природы (МСОП) произвел первый красный список для угрожаемых океанических акул и скатов. Они утверждают, что примерно одна треть акул и скатов открытого океана находится под угроза исчезновения.[84] В списке 64 вида океанических акул и скатов, в том числе молоты, гигант дьявольские лучи, и сельдевая акула.[85]
Океанические акулы попал случайно меч-рыба и тунец открытое море рыболовство. В прошлом было мало рынков для акул, которые считались бесполезными. прилов. Теперь акулы становятся все более нацеленными на поставки на развивающиеся азиатские рынки, особенно для акульи плавники, которые используются в суп из акульих плавников.[85]
По оценкам, популяция акул в северо-западной части Атлантического океана сократилась на 50% с начала 1970-х годов. Океанические акулы уязвимы, потому что они не производят много детенышей, а для созревания молоди могут потребоваться десятилетия.[85]

В зубчатый молот классифицируется как находящийся под угрозой исчезновения

В океаническая белоперая акула снизился на 99% в Мексиканский залив[85]

В дьявольская рыба, большой луч, находится под угрозой

В белоснежная акула находится под угрозой




В некоторых частях мира зубчатый молот акулы сократились на 99% с конца 1970-х годов. Его статус в красном списке означает, что он находится под угрозой исчезновения во всем мире, что означает, что он находится на грани исчезновения.[85]
Смотрите также
|
|
Рекомендации
Примечания
- ^ а б c d е Lal, Brij V .; Удача, Кейт (2000). Острова Тихого океана: энциклопедия. Гавайский университет Press. п. 8. ISBN 978-0-8248-2265-1.
- ^ а б c d е ж грамм час Мойл и Чех, п. 585
- ^ McLintock, AH (ред.) «Рыба, морская». Те Ара - Энциклопедия Новой Зеландии. Обновлено 18 сентября 2007 г.
- ^ Валронд, Карл. «Океаническая рыба». Энциклопедия Новой Зеландии. Обновлено 21 сентября 2007 г.
- ^ а б c d е ж грамм Мойл и Чех, п. 571
- ^ а б Селедка, Питер (2002). Биология глубокого океана. Oxford University Press. С. 192–95. ISBN 978-0-19-854956-7.
- ^ а б c d е ж грамм Мойл и Чех, п. 572
- ^ Блэкберн (1965). «Океанография и экология тунца». Океанография и морская биология: ежегодный обзор. 3: 299–322.
- ^ Хантер, младший; Митчелл CT (1966). «Ассоциация рыб с обломками в прибрежных водах Центральной Америки». Бюллетень рыболовства. 66: 13–29.
- ^ Кингсфорд MJ (1993). «Биотическая и абиотическая структура в пелагической среде: важность для мелких рыб». Бюллетень морской науки. 53 (2): 393–415.
- ^ Дули Дж. К. (1972). «Рыбы, связанные с пелагическим комплексом саргасса, с обсуждением сообщества саргассов». Вклад в морскую науку. 16: 1–32.
- ^ Мойл и Чех, п. 576
- ^ а б c d е Мойл и Чех, стр. 574–575
- ^ Хосе, Э. (2000). "Typologie et comportement des agrégations thonières autour de dispositifs de focus de poissons à partir de prospections acoustiques en Polynésie française". Водные живые ресурсы. 13 (4): 183–192. Дои:10.1016 / S0990-7440 (00) 00051-6.
- ^ Frazier, J. G .; Fierstine, H.L .; Beavers, S.C .; Achaval, F .; Suganuma, H .; Pitman, R.L .; Yamaguchi, Y .; Приджони, К. М. (1994). «Прокол морских черепах (Reptitia, Chelonia: Cheloniidae и Dermochelyidae) морскими рыбами (Osteichthyes, Perciformes: Istiophoridae и Xiphiidae)». Экологическая биология рыб. 39: 85–96. Дои:10.1007 / BF00004759. S2CID 23551149.
- ^ а б Мойл и Чех, п. 591
- ^ а б c Хэдрих, Р. Л. (1996). «Глубоководные рыбы: эволюция и адаптация в самых больших жизненных пространствах Земли». Журнал биологии рыб. 49: 40–53. Дои:10.1111 / j.1095-8649.1996.tb06066.x.
- ^ Мойл и Чех, п. 586
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Argyropelecus aculeatus" в FishBase. Версия от августа 2009 г.
- ^ а б c d е Salvanes, A.G.V .; Кристофферсен, Дж. Б. (2001). «Мезопелагические рыбы» (PDF). Энциклопедия наук об океане. 3. Получено 4 ноября 2020.
- ^ а б Райан П. «Глубоководные существа: мезопелагическая зона» Те Ара - Энциклопедия Новой Зеландии. Обновлено 21 сентября 2007 г.
- ^ Кость и Мур, п. 38.
- ^ Дуглас, Э .; Friedl, W .; Пиквелл, Г. (1976). «Рыбы в зонах минимума кислорода: характеристики оксигенации крови». Наука. 191 (4230): 957–9. Bibcode:1976Научный ... 191..957D. Дои:10.1126 / science.1251208. PMID 1251208.
- ^ Мойл и Чех, п. 590
- ^ Мунц, В. Р. А. (2009). «О желтых линзах у мезопелагических животных». Журнал Морской биологической ассоциации Соединенного Королевства. 56 (4): 963–976. Дои:10.1017 / S0025315400021019.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Dissostichus mawsoni" в FishBase. Версия от августа 2009 г.
- ^ Тайна глубоководной рыбы с трубчатыми глазами и прозрачной головой раскрыта ScienceDaily, 24 февраля 2009 г.
- ^ а б Kenaley, C.P (2007). "Пересмотр семейства Stoplight Loosejaw" Малакостей (Teleostei: Stomiidae: Malacosteinae), с описанием нового вида из умеренного южного полушария и Индийского океана ». Копея. 2007 (4): 886–900. Дои:10.1643 / 0045-8511 (2007) 7 [886: ROTSLG] 2.0.CO; 2.
- ^ Саттон, Т.Т. (ноябрь 2005 г.). «Трофическая экология глубоководных рыб. Malacosteus niger (Рыбы: Stomiidae): Загадочная экология кормления, обеспечивающая уникальную зрительную систему? ". Deep-Sea Research Part I: Oceanographic Research Papers. 52 (11): 2065–2076. Bibcode:2005DSRI ... 52.2065S. Дои:10.1016 / j.dsr.2005.06.011.
- ^ Вагнер, Х.Дж., Дуглас, Р.Х., Франк, Т.М., Робертс, штат Нью-Йорк, и Партридж, Дж. К. (27 января 2009 г.). «Новый глаз позвоночных животных, использующий как рефракционную, так и отражающую оптику». Текущая биология. 19 (2): 108–114. Дои:10.1016 / j.cub.2008.11.061. PMID 19110427. S2CID 18680315.CS1 maint: несколько имен: список авторов (связь)
- ^ Смит, Л. (8 января 2009 г.). «Рыба с четырьмя глазами может видеть сквозь мрак глубокого моря». Times Online. Times Newspapers Ltd., получено 14 марта 2009 г.
- ^ Халли, П. Александр (1998). Paxton, J.R .; Эшмейер, W.N. (ред.). Энциклопедия рыб. Сан-Диего: Academic Press. С. 127–128. ISBN 978-0-12-547665-2.
- ^ Cornejo, R .; Коппельманн, Р. и Саттон, Т. (2006). «Разнообразие глубоководных рыб и экология в придонном пограничном слое».
- ^ Мойл и Чех, п. 336
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Гигантура чуни" в FishBase. Версия от октября 2010 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Anotopterus pharao" в FishBase. Версия от апреля 2010 г.
- ^ а б c d е Райан П. «Глубоководные существа: Батипелагическая зона» Те Ара - Энциклопедия Новой Зеландии. Обновлено 21 сентября 2007 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2006). "Гоностома батифильная" в FishBase. Версия от января 2006 г.
- ^ Froese, Rainer и Daniel Pauly, ред. (2009). «Гоностома» в FishBase. Версия от августа 2009 г.
- ^ Шмид, Рэндольф Э. (22 января 2009 г.). «Ученые разгадывают тайну: все 3 рыбы одинаковые». Ассошиэйтед Пресс.
- ^ а б c Мойл и Чех, п. 594
- ^ а б c Мойл и Чех, п. 587
- ^ Маршалл (1984) «Прогенетические тенденции у глубоководных рыб», стр. 91–101 в Potts GW и Wootton RJ (ред.) (1984) Размножение рыб: стратегии и тактика Общество рыболовства Британских островов.
- ^ Хорн MH (1970). «Плавательный пузырь как ювенильный орган строматеидных рыб». Breviora. 359: 1–9.
- ^ Джемпер, Дж .; Бэрд, Р. К. (1991). "Расположение по обонянию: модель и приложение к проблеме спаривания у глубоководных топориков" Argyropelecus hemigymnus". Американский натуралист. 138 (6): 1431. Дои:10.1086/285295. JSTOR 2462555.
- ^ Пич, Т. В. (1975). "Преждевременный половой паразитизм в глубоководных цератиоидных удильщиках, Cryptopsaras couesi Джилл ». Природа. 256 (5512): 38–40. Bibcode:1975Натура.256 ... 38П. Дои:10.1038 / 256038a0. S2CID 4226567.
- ^ Джордан, Д.С. (1905). Руководство по изучению рыб. Х. Холт и компания.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Chiasmodon niger" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Anoplogaster cornuta" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Chauliodus sloani" в FishBase. Версия от апреля 2010 г.
- ^ а б c Уолронд С Карл. «Прибрежная рыба - Рыба дна открытого моря» Те Ара - Энциклопедия Новой Зеландии. Обновлено 2 марта 2009 г.
- ^ а б Мойл и Чех, п. 588
- ^ Mauchline J; Гордон JDM (1986). «Стратегии добычи глубоководной рыбы». Mar. Ecol. Прог. Сер. 27: 227–238. Bibcode:1986МЭПС ... 27..227М. Дои:10.3354 / meps027227.
- ^ Кослоу, Дж. А. (1996). «Образцы энергии и жизненного цикла глубоководных донных, бентопелагических рыб и рыб, связанных с подводными горами». Журнал биологии рыб. 49: 54–74. Дои:10.1111 / j.1095-8649.1996.tb06067.x.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Acanthonus armatus" в FishBase. Версия от августа 2009 г.
- ^ а б Хорошо, M. L .; Хорн, М. Х .; Кокс, Б. (1987). "Acanthonus armatus, глубоководная костистая рыба с коротким мозгом и большими ушами ". Труды Королевского общества B: биологические науки. 230 (1259): 257–65. Bibcode:1987RSPSB.230..257F. Дои:10.1098 / rspb.1987.0018. JSTOR 36061. PMID 2884671. S2CID 19183523.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Hoplostethus atlanticus" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Dissostichus eleginoides" в FishBase. Версия от августа 2009 г.
- ^ Сулак KJ. «Систематика и биология Батиптерой (Pisces, Chlorophthalmidae) с пересмотренной классификацией донных миктофоформных рыб ». Галатея Реп. 14.
- ^ Нильсен Дж. Г. (1977). "Самая глубокая из ныне живущих рыб Abyssobrotula galatheae: новый род и вид яйцекладущих офидиоидов (Pisces, Brotulidae) ». Отчет Галатеи. 14: 41–48.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Abyssobrotula galatheae" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Батиптероевый граллатор" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Taeniura Meyeni" в FishBase. Версия от августа 2009 г.
- ^ Нильсен, Дж. (1977). "Самая глубокая из ныне живущих рыб Abyssobrotula galatheae: новый род и вид яйцекладущих офидиоидов (Pisces, Brotulidae) ». Отчет Галатеи. 14: 41–48.
- ^ Checkley D, Alheit J и Oozeki Y (2009) Изменение климата и мелкие пелагические рыбы, Издательство Кембриджского университета. ISBN 0-521-88482-9.
- ^ а б c d Пелагические виды В архиве 2012-02-11 в Wayback Machine Пелагическая ассоциация морозильников-траулеров. Проверено 22 июля 2009 года.
- ^ Пелагические рыбы Исландское рыболовство. Проверено 24 июля 2009 года.
- ^ Путассу Институт морских исследований. Проверено 23 июля 2009 года.
- ^ Кость и Мур, п. 443
- ^ ФАО: Проект LAPE Кормовые виды Рим. Обновлено 28 ноября 2008 г.
- ^ а б Скумбрия Институт морских исследований. Проверено 23 июля 2009 года.
- ^ Chavez, F. P .; Райан, Джон; Луч-Кота, Сальвадор, Э .; Чикен С., Мигель (2003). «От анчоусов до сардин и обратно: многолетние изменения в Тихом океане». Наука. 299 (5604): 217–21. Bibcode:2003Наука ... 299..217C. Дои:10.1126 / science.1075880. PMID 12522241. S2CID 37990897.
- ^ Половина, Дж. Дж. (1996). "Десятилетняя изменчивость транс-Тихоокеанской миграции северного синего тунца (Thunnus tynnus) в соответствии с изменением численности добычи, вызванным климатом ». Океанография рыболовства. 5 (2): 114–119. Дои:10.1111 / j.1365-2419.1996.tb00110.x.
- ^ ФАО: Информационные бюллетени по видам: Sardinops melanostictus (Шлегель, 1846 г.) Рим. Проверено 18 августа 2009 года.
- ^ Накамура, Хироши (1969). Распространение и миграция тунца. Новости рыбалки.
- ^ Блэкберн М (1965). «Океанография и экология тунца» (PDF). Океанография и морская биология: ежегодный обзор. 3: 299–322.
- ^ Casey, J. G .; Колер, Н. Э. (1992). "Исследования по мечению короткоперой акулы-мако (Isurus oxyrinchus) в западной части Северной Атлантики ». Морские и пресноводные исследования. 43: 45. Дои:10.1071 / MF9920045.
- ^ Мойл и Чех, п. 578
- ^ Блок, Б. А .; Дьюара, H; Blackwell, S. B .; Уильямс, Т. Д .; Prince, E.D .; Farwell, C.J .; Бустани, А; Teo, S. L .; Зейтц, А; Валли, А; Фадж, Д. (2001). «Миграционные движения, предпочтения глубины и термобиология атлантического синего тунца» (PDF). Наука. 293 (5533): 1310–4. Bibcode:2001Sci ... 293.1310B. Дои:10.1126 / science.1061197. PMID 11509729. S2CID 32126319.
- ^ Объединенные Нации Конвенция о Закон моря: Текст
- ^ Тихоокеанский совет по управлению рыболовством: Справочная информация: далеко мигрирующие виды В архиве 2009-07-12 в Wayback Machine
- ^ Рыболовство и аквакультура. ФАО. Проверено 1 мая 2015.
- ^ ФАО (2007) Состояние мирового рыболовства и аквакультуры, 2006 г. Департамент рыболовства и аквакультуры. ISBN 978-92-5-105568-7
- ^ Третья часть акул открытого океана находится под угрозой исчезновения МСОП. 25 июня 2009 г.
- ^ а б c d е Рыбная ловля подвергает риску исчезновения треть всех видов океанических акул guardian.co.uk, 26 июня 2009 г.
Библиография
- Кость, Квентин; Мур, Ричард (2008). Биология рыб. Наука о гирляндах. ISBN 978-0-203-88522-2.
- Мойл, П. Б. и Чех, Дж. Дж. (2004). Рыбы, Введение в ихтиологию (5-е изд.). Бенджамин Каммингс. ISBN 978-0-13-100847-2.
дальнейшее чтение
- Коллетт, BB (2010) «Размножение и развитие эпипелагических рыб» В: Кэтлин С. Коул, Размножение и сексуальность морских рыб: закономерности и процессы, стр. 21–64, Калифорнийский университет Press. ISBN 978-0-520-26433-5.
- Фреон, Пьер (1998) Динамика распределения и поведения пелагических рыб: влияние на промысел и оценку запасов, Wiley-Blackwell. ISBN 978-0-85238-241-7.
- Йонсен, S (2003). «Снимая покров невидимости: влияние изменения оптических условий на пелагический крипсис1». Интегративная и сравнительная биология. 43 (4): 580–590. Дои:10.1093 / icb / 43.4.580. PMID 21680466.
- Макрис, N; Ratilal, P; Джаганнатан, S; Гонг, Z; Эндрюс, М; Бертсатос, I; Годо, штат Орегон; Неро, RW; Jech, JM (2009). «Критическая плотность населения вызывает быстрое образование обширных косяков океанических рыб». Наука. 323 (5922): 1734–1737. Bibcode:2009Научный ... 323.1734M. Дои:10.1126 / science.1169441. PMID 19325116. S2CID 6478019.
- Пепперелл Дж (2011) Рыбы открытого океана: естествознание и иллюстрированный справочник Издательство Университета Нового Южного Уэльса, ISBN 978-1-74223-267-6.
- Salvanesa AGV и Kristoffersen JB «Мезопелагические рыбы» В: Энциклопедия наук об океанеС. 1711–1717. Дои:10.1006 / rwos.2001.0012
- Ученые установили генезис моделей поведения животных PhysOrg.com, 26 марта 2009 г.
- Одна рыба, две рыбы: новый датчик MIT улучшает подсчет рыбы PhysOrg.com, 2 февраля 2006 г.
внешняя ссылка
- Пелагическая рыба – Институт морских исследований
- Яркая жизнь в подводном мире Видео TED от Эдит Виддер
- Открытый океан MarineBio.org. MarineBio.org. Обновлено 28 августа 2011 г. Видео TED от Эдит Виддер
- Открытый океан MarineBio.org. MarineBio.org. Обновлено 28 августа 2011 г.
- Пелагический региональный консультативный совет из Европейская комиссия


| О рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||
| |||||||||||||||||||||
| |||||||||||||||||||||

