Келп лес - Kelp forest
| Морские места обитания |
|---|
 Келп лес |
Ламинарии леса подводные районы с высокой плотностью ламинария, который охватывает большую часть побережья мира. Они признаны одними из самых продуктивных и динамичных. экосистемы на земле.[1] Меньшие участки прикрепленной келпа называются грядки из водорослей.
– Чарльз Дарвин, 1 июня 1834 г., Огненная Земля, Чили[2]
Леса водорослей встречаются во всем мире умеренный и полярный прибрежные океаны.[1] В 2007 г. ламинарии были обнаружены также в тропический воды рядом Эквадор.[3]
Физически образован коричневым макроводоросли, леса водорослей обеспечивают уникальную среду обитания для морских организмов[4] и являются источником понимания многих экологических процессов. За последнее столетие они стали предметом обширных исследований, особенно в трофический экология, и продолжают вызывать важные идеи, актуальные за пределами этой уникальной экосистемы. Например, леса ламинарии могут влиять на прибрежные океанографический узоры[5] и предоставить много экосистемные услуги.[6]
Однако влияние людей часто способствовало появлению ламинарии. деградация леса. Особую озабоченность вызывают эффекты перелов прибрежные экосистемы, которые могут выпускать травоядные животные от нормального регулирования популяции и приводят к чрезмерный выпас ламинарии и других водорослей.[7] Это может быстро привести к переходу на бесплодные пейзажи где сохранилось относительно немного видов.[8][9] Уже из-за комбинированного воздействия перелов и изменение климата, леса водорослей почти исчезли во многих особенно уязвимых местах, таких как Тасмания восточное побережье и побережье Северная Калифорния.[10][11] Реализация морские охраняемые территории является одной из стратегий управления, полезных для решения таких проблем, поскольку она может ограничить воздействие рыболовства и защитить экосистему от дополнительных воздействий других факторов экологического стресса.
Водоросли
Период, термин ламинария относится к морские водоросли принадлежащий к порядок Laminariales (тип: Heterokontophyta ). Хотя водоросли не считаются таксономически разнообразным отрядом, они очень разнообразны структурно и функционально.[6] Наиболее известные виды - гигантские водоросли (Макроцистис spp.), хотя многие другие роды, такие как Ламинария, Эклония, Lessonia, Алария, и Эйсения описаны.
Широкий спектр морских обитателей использует леса ламинарии для защиты или в пищу, включая рыбу. В лесах водорослей северной части Тихого океана, особенно морской окунь, и много беспозвоночные, Такие как амфиподы, креветка, морские улитки, щетинистые черви, и хрупкие звезды. Встречаются также многие морские млекопитающие и птицы, в том числе тюлени, морские львы, киты, морские выдры, чайки, крачки, снежные цапли, большие голубые цапли, и бакланы, а также некоторые береговые птицы.[12]
Часто считается экосистемный инженер, ламинария обеспечивает физический субстрат и среду обитания для лесных сообществ ламинарии.[13] В водорослях (царство Протиста ) тело отдельного организма известно как слоевище а не как растение (царство Plantae ). Морфологическое строение слоевища ламинарии определяется тремя основными структурными единицами:[8]
- В стойко держаться представляет собой корнеобразную массу, которая прикрепляет слоевище к морскому дну, хотя, в отличие от настоящих корней, не отвечает за поглощение и доставку питательных веществ к остальной части слоевища.
- В ножка аналогичен стеблю растения, идущему вертикально от опоры и обеспечивающему опорный каркас для других морфологических особенностей.
- В листья представляют собой прикрепления в виде листов или пластинок, отходящие от ножки, иногда по всей ее длине, и являются местами поглощения питательных веществ и фотосинтетической активности.
Кроме того, многие виды водорослей имеют пневматоцисты, или газонаполненные пузыри, обычно расположенные у основания листьев около ножки. Эти конструкции обеспечивают келпу необходимую плавучесть, чтобы поддерживать вертикальное положение в толще воды.
Факторы окружающей среды, необходимые для выживания ламинарии, включают твердый субстрат (обычно камень или песок), высокое содержание питательных веществ (например, азот, фосфор) и свет (минимум годовой сияние доза> 50 Е м−2[14]). Особенно продуктивные ламинарии обычно связаны с районами значительного океанографического апвеллинг, процесс, который доставляет прохладную, богатую питательными веществами воду из глубины в океан. смешанный поверхностный слой.[14] Поток воды и турбулентность способствуют усвоению питательных веществ листьями ламинарии по всей толще воды.[15] Прозрачность воды влияет на глубину, на которую может быть передано достаточное количество света. В идеальных условиях ламинария гигантская (Макроцистис spp.) может вырастать по вертикали на 30–60 см за день. Некоторые виды, такие как Nereocystis, находятся однолетники, а другие, такие как Эйсения находятся многолетние растения, живущим более 20 лет.[16] В многолетних лесах из водорослей максимальные темпы роста наблюдаются в течение месяцев апвеллинга (обычно весной и летом), а их отмирание соответствует снижению доступности питательных веществ, более коротким фотопериодам и увеличению частоты штормов.[8]
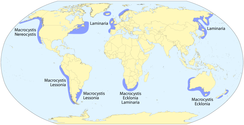
Келпы в первую очередь связаны с умеренный и Арктический воды по всему миру. Из наиболее доминирующих родов Ламинария в основном связан с обеими сторонами Атлантический океан и побережья Китай и Япония; Эклония находится в Австралия, Новая Зеландия, и Южная Африка; и Макроцистис встречается на северо-востоке и юго-востоке Тихий океан, Южный океан архипелагов, а также участками вокруг Австралии, Новой Зеландии и Южной Африки.[8] Регион с наибольшим разнообразием водорослей (более 20 видов) - северо-восток Тихого океана, к северу от Сан - Франциско, Калифорния, в Алеутские острова, Аляска.
Хотя ламинарии неизвестны в тропических поверхностных водах, некоторые виды водорослей Ламинария известно, что они встречаются исключительно в глубоких тропических водах.[17][18] Считается, что это общее отсутствие ламинарии в тропиках в основном связано с недостаточным уровнем питательных веществ, связанных с теплом, олиготрофный воды.[8] Одно недавнее исследование, наложившее необходимые физические параметры ламинарии на средние океанографические условия, позволило создать модель, предсказывающую существование подповерхностных водорослей в тропиках по всему миру на глубине до 200 м. Для точки доступа в Галапагосские острова, локальная модель была улучшена с мелкомасштабными данными и протестирована; Исследовательская группа обнаружила процветающие леса ламинарии на всех восьми выбранных участках, все из которых были предсказаны моделью, что подтвердило свой подход. Это говорит о том, что их глобальная модель на самом деле может быть довольно точной, и если это так, то ламинарии будут изобиловать тропическими подземными водами во всем мире.[3] Важность этого вклада была быстро признана в научном сообществе и подтолкнула к совершенно новой траектории исследований лесов ламинарии, особенно подчеркивая потенциал пространственного убежища от изменения климата, а также объяснения эволюционных паттернов ламинарии во всем мире.[19]
Архитектура экосистемы




Архитектура экосистемы ламинарии основана на ее физической структуре, которая влияет на связанные виды, которые определяют структуру ее сообщества. Структурно экосистема включает три гильдии водорослей и две гильдии, занятые другими водорослями:[8]
- Морские водоросли включают самые крупные виды и часто представляют собой плавающие навесы, простирающиеся до поверхности океана (например, Макроцистис и Алария).
- Келпы-гребешки обычно возвышаются на несколько метров над морским дном и могут расти плотными скоплениями (например, Эйсения и Эклония).
- Распростертые водоросли лежат рядом с морским дном и вдоль него (например, Ламинария).
- В бентосный Комплекс состоит из других видов водорослей (например, нитчатых и листовых функциональных групп, сочлененных кораллов) и сидячих организмов на дне океана.
- Инкрустация коралловых водорослей напрямую и часто широко покрывает геологический субстрат.
В лесу часто сосуществуют несколько видов водорослей; Термин «навес подлеска» относится к ножкам и распростертым водорослям. Например, Макроцистис навес может простираться на много метров над морским дном по направлению к поверхности океана, в то время как подлесок водорослей Эйсения и Птеригофора достигает вверх всего на несколько метров. Под этими водорослями может встречаться бентосное скопление лиственных красных водорослей. Плотная вертикальная инфраструктура с вышележащим пологом образует систему микроокружений, аналогичных тем, которые наблюдаются в наземном лесу, с солнечным пологом, частично затененной серединой и затемненным морским дном.[8] У каждой гильдии есть ассоциированные организмы, которые различаются по уровню зависимости от среды обитания, и совокупность этих организмов может меняться в зависимости от морфологии ламинарии.[20][21][22] Например, в Калифорнии Macrocystis pyrifera леса, голожаберный Мелибе леонина, и скелет креветки Капрелла калифорнийская тесно связаны с поверхностными навесами; окунь ламинарии Brachyistius frenatus, морской окунь Себастес spp., и многие другие рыбы водятся в подлеске прилистников; хрупкие звезды и улитки в тюрбане Тегула виды тесно связаны с водорослями, в то время как различные травоядные животные, такие как морские ежи и морские ушки, живут под распростертым пологом; многие морские звезды, гидроиды и бентосный среди донных скоплений обитают рыбы; одиночные кораллы, различные брюхоногие моллюски, и иглокожие живут над корковыми коралловыми водорослями.[20] Кроме того, пелагические рыбы и морские млекопитающие слабо связаны с лесами ламинарии, обычно взаимодействуя у краев, когда они посещают, чтобы питаться местными организмами.
Трофическая экология



Классические исследования экологии ламинарии в основном сосредоточены на трофических взаимодействиях (взаимоотношениях между организмами и их пищевые полотна ), особенно понимание и трофические процессы сверху вниз. Процессы снизу вверх, как правило, обусловлены абиотическими условиями, необходимыми для роста первичных производителей, такими как доступность света и питательных веществ и последующая передача энергии потребителям на более высоких трофических уровнях. Например, появление ламинарии часто коррелирует с океанографическими зонами апвеллинга, которые обеспечивают необычно высокие концентрации питательных веществ в местной окружающей среде.[23][24] Это позволяет водорослям расти и впоследствии поддерживать травоядных животных, что, в свою очередь, поддерживает потребителей на более высоком уровне. трофические уровни.[25] Напротив, в процессах сверху вниз хищники ограничивают биомассу видов на более низких трофических уровнях за счет потребления. В отсутствие хищников эти виды низшего уровня процветают, потому что ресурсы, поддерживающие их энергетические потребности, не ограничены. В хорошо изученном примере из лесов водорослей Аляски[26] морские выдры (Enhydra lutris) контрольные популяции травоядных морские ежи через хищничество. Когда каланы удаляются из экосистемы (например, в результате эксплуатации человеком), популяции ежей выходят из-под контроля хищников и резко увеличиваются. Это приводит к увеличению давления травоядных на местные водоросли. Ухудшение состояния ламинарии приводит к потере физической структуры экосистемы и, как следствие, к потере других видов, связанных с этой средой обитания. В экосистемах лесных водорослей Аляски каланы являются краеугольные камни это опосредует это трофический каскад. В Южной Калифорнии леса водорослей сохраняются без каланов, и контроль над травоядными ежами вместо этого осуществляется целым набором хищников, включая омаров и крупных рыб, таких как Калифорнийская овчарка. Эффект удаления одного хищного вида в этой системе отличается от Аляски, потому что существует избыточность на трофических уровнях, а другие хищные виды могут продолжать регулировать ежей.[21] Однако устранение нескольких хищников может эффективно освободить ежей от давления хищников и позволить системе следовать траекториям к деградации леса ламинарии.[27] Подобные примеры существуют в Новая Шотландия,[28] Южная Африка,[29] Австралия[30] и Чили.[31] Относительная важность контроля сверху вниз по сравнению с контролем снизу вверх в лесных экосистемах ламинарии и сила трофических взаимодействий по-прежнему являются предметом значительных научных исследований.[32][33][34]
Переход от макроводорослей (например, леса водорослей) к обнаженным ландшафтам, где преобладают морские ежи (или «пустоши ежей»), является широко распространенным явлением.[6][35][36][37] часто возникают в результате трофических каскадов, подобных описанным выше; две фазы рассматриваются как альтернативные стабильные состояния экосистемы.[38][39] Восстановление лесов водорослей из состояния бесплодия было документально подтверждено после драматических потрясений, таких как болезнь ежей или большие изменения температурных условий.[27][40][41] Восстановление из промежуточных состояний ухудшения менее предсказуемо и зависит от комбинации абиотических факторов и биотических взаимодействий в каждом случае.
Хотя ежи обычно являются доминирующими травоядными животными, другие существенные силы взаимодействия включают: морские звезды, изоподы, ламинария крабы, и травоядные рыбы.[8][32] Во многих случаях эти организмы питаются водорослями, которые были вытеснены из субстрата и дрейфуют у дна океана, вместо того, чтобы тратить энергию на поиск целых талломов, которыми можно было бы питаться. Когда водоросли достаточно дрейфуют, травоядные травоядные не оказывают давления на прикрепленные растения; когда дотации на дрейф недоступны, травоядные напрямую влияют на физическую структуру экосистемы.[42][43] Многие исследования в Южной Калифорнии показали, что наличие дрейфующих водорослей влияет на поведение морских ежей в поисках пищи.[44][45] Дрейфующие водоросли и твердые частицы, полученные из водорослей, также сыграли важную роль в субсидировании прилегающих мест обитания, таких как песчаные пляжи и каменистая приливная зона.[46][47][48]
Динамика патча
Еще одна важная область исследований лесов ламинарии была направлена на понимание пространственно-временных паттернов пятен ламинарии. Такая динамика не только влияет на физический ландшафт, но также влияет на виды, которые связываются с водорослями в поисках убежища или поиска пищи.[20][25] Крупномасштабные нарушения окружающей среды позволили сделать важные выводы о механизмах и экосистемах. устойчивость. Примеры нарушений окружающей среды:
- Острый и хронический загрязнение Было показано, что события влияют на леса водорослей южной Калифорнии, хотя интенсивность воздействия, похоже, зависит как от природы загрязнителей, так и от продолжительности воздействия.[49][50][51][52][53] Загрязнение может включать отложение отложений и эвтрофикация из сточных вод, промышленных побочных продуктов и загрязняющих веществ, таких как Печатные платы и тяжелые металлы (например, медь, цинк), сток органофосфаты из сельскохозяйственных угодий, химикаты против обрастания, используемые в гаванях и пристанях для яхт (например, TBT и креозот ) и наземных патогенов, таких как фекальные колиформные бактерии.
- Катастрофические штормы могут разрушать поверхностные водоросли из-за активности волн, но обычно оставляют водоросли нетронутыми; они также могут удалять ежей, когда у них мало пространственного убежища.[38][43] Перемежающиеся поляны навеса создают мозаику морского пейзажа, где солнечный свет проникает глубже в лес водорослей, и виды, которые обычно ограничены светом в подлеске, могут процветать. Точно так же субстрат, очищенный от опор ламинарии, может предоставить место для других сидячих видов, чтобы они обосновались и заняли морское дно, иногда напрямую конкурируя с молодыми водорослями и даже препятствуя их расселению.[54]
- Эль-Ниньо - Южное колебание (ENSO) явления включают понижение океанографических термоклинов, резкое сокращение поступления питательных веществ и изменение характера штормов.[38][55] Стресс из-за теплой воды и истощение питательных веществ может повысить восприимчивость ламинарии к ураганам и травоядным выпасам, иногда даже вызывая фазовые сдвиги в ландшафты, где преобладают ежи.[41][44][56] В целом, океанографические условия (то есть температура воды, течения) влияют на успех пополнения ламинарии и ее конкурентов, что явно влияет на последующие взаимодействия видов и динамику леса ламинарии.[38][57]
- Перелов более высоких трофических уровней, которые естественным образом регулируют популяции травоядных, также признан важным фактором стресса в лесах ламинарии.[7][34][58] Как описано в предыдущем разделе, движущие силы и результаты трофических каскадов важны для понимания пространственно-временных моделей лесов ламинарии.[26][27][32]
В дополнение к экологическому мониторингу лесов ламинарии до, во время и после таких нарушений, ученые пытаются разобраться в тонкостях динамики ламинарии с помощью экспериментальных манипуляций. Работая в более мелких пространственно-временных масштабах, они могут контролировать наличие или отсутствие определенных биотических и абиотических факторов, чтобы обнаружить действующие механизмы. Например, в южной Австралии манипуляции с типами полога ламинарии показали, что относительное количество Ecklonia radiata в пологе может использоваться для предсказания сообществ подлеска; следовательно, доля E. radiata может использоваться как индикатор других видов, встречающихся в окружающей среде.[59]
Человеческое использование
Леса водорослей были важны для существования человека на протяжении тысяч лет.[60] В самом деле, многие сейчас предполагают, что первая колонизация Америки была вызвана рыболовными сообществами, которые следовали за лесами водорослей Тихого океана во время последнего ледникового периода. Одна из теорий утверждает, что леса из водорослей, которые простирались бы от Северо-Восточной Азии до американского побережья Тихого океана, принесли бы много пользы древним лодочникам. Леса водорослей предоставили бы много возможностей к существованию, а также выступили в качестве своего рода буфера от бурной воды. Помимо этих преимуществ, исследователи полагают, что леса из ламинарии могли помочь первым яхтсменам ориентироваться, действуя как своего рода «шоссе из ламинарии». Теоретики также предполагают, что леса водорослей помогли этим древним колонистам, обеспечив им стабильный образ жизни и не давая им адаптироваться к новым экосистемам и разрабатывать новые методы выживания, даже когда они преодолевали тысячи миль.[61] Современная экономика основана на рыболовство видов, связанных с водорослями, таких как Омар и морской окунь. Люди также могут собирать водоросли непосредственно для кормления аквакультурных видов, таких как морское ушко и для экстракции соединения альгиновая кислота, который используется в таких продуктах, как зубная паста и антациды.[62][63] Леса водорослей ценятся для рекреационных мероприятий, таких как подводное плавание с аквалангом и каякинг; Отрасли, поддерживающие эти виды спорта, представляют одно преимущество, связанное с экосистемой, а удовольствие, получаемое от этих видов деятельности, - другое. Все это примеры экосистемные услуги обеспечивается специально лесами ламинарии.
Угрозы и управление

Учитывая сложность лесов ламинарии - их изменчивую структуру, географию и взаимодействие - они представляют собой серьезную проблему для менеджеров по охране окружающей среды. Экстраполировать даже хорошо изученные тенденции в будущее сложно, потому что взаимодействия внутри экосистемы будут меняться в различных условиях, не все взаимосвязи в экосистеме понятны, а нелинейные пороги переходов еще не признаны.[64] Что касается лесов водорослей, основные проблемы, вызывающие беспокойство, включают загрязнение моря и качество воды, сбор ламинарии и рыболовство, инвазивные виды,[6] и изменение климата.[65] Самой серьезной угрозой сохранению лесов водорослей может быть чрезмерный вылов прибрежных экосистем, что за счет устранения более высоких трофических уровней способствует их переходу в бедные пустоши ежей.[7] Поддержание биоразнообразие считается способом общей стабилизации экосистем и их услуг с помощью таких механизмов, как функциональная компенсация и снижение восприимчивости к инвазиям чужеродных видов.[66][67][68][69]
Во многих местах менеджеры решили регулировать сбор ламинарии.[24][70] и / или вылов лесных водорослей рыбным промыслом.[6][58] Хотя они могут быть эффективными в определенном смысле, они не обязательно защищают всю экосистему. Морские охраняемые территории (MPA) предлагают уникальное решение, охватывающее не только целевые виды для промысла, но также взаимодействия, окружающие их и местную среду в целом.[71][72] Прямые выгоды от МОР для рыболовства (например, побочные эффекты) хорошо документированы во всем мире.[7][73][74][75] Косвенные преимущества также были продемонстрированы в нескольких случаях среди таких видов, как морские ушки и рыбы в Центральной Калифорнии.[76][77] Что наиболее важно, МОР могут быть эффективными для защиты существующих экосистем ламинарии, а также могут способствовать восстановлению тех, которые были затронуты.[38][78][79]
Рекомендации
- ^ а б Манн, К. 1973. Морские водоросли: их продуктивность и стратегия роста. Наука 182: 975-981.
- ^ Дарвин, С. 1909. Путешествие Бигля. Гарвардская классика, том 29. Нью-Йорк, США: П.Ф. Компания Collier & Son.
- ^ а б Грэм, M.H., B.P. Кинлан, Л. Дрюль, Л. Гарске и С. Бэнкс. 2007. Глубоководные рефугиумы водорослей как потенциальные горячие точки тропического морского разнообразия и продуктивности. Труды Национальной академии наук 104: 16576-16580.
- ^ Кристи, Х., Йоргенсен, Н.М., Нордерхауг, К.М., Вааге-Нильсен, Э., 2003. Распределение видов и использование среды обитания фауны, связанной с водорослями (Laminaria hyperborea) вдоль норвежского побережья. Журнал Морской биологической ассоциации Великобритании 83, 687-699.
- ^ Джексон, Г.А. и C.D. Winant. 1983. Влияние ламинарии на прибрежные течения. Отчет о континентальном шельфе 2: 75-80.
- ^ а б c d е Steneck, R.S., M.H. Грэм, Б.Дж. Бурк, Д. Корбетт, Дж. М. Эрландсон, J.A. Estes и М.Дж. Тегнер. 2002. Лесные экосистемы водорослей: биоразнообразие, стабильность, устойчивость и будущее. Охрана окружающей среды 29: 436-459.
- ^ а б c d Сала, Э., К.Ф. Бурдуреск и М. Хармелин-Вивьен. 1998. Рыбалка, трофические каскады и структура сообществ водорослей: оценка старой, но непроверенной парадигмы. Ойкос 82: 425-439.
- ^ а б c d е ж грамм час Дейтон, П.К. 1985a. Экология сообществ ламинарии. Ежегодный обзор экологии и систематики 16: 215-245.
- ^ Нордерхауг, К.М., Кристи, Х., 2009. Выпас морских ежей и возобновление растительности водорослей в северо-восточной части Атлантического океана. Исследования морской биологии 5, 515-528
- ^ Мортон, Адам; Корделл, Марни; Фаннер, Дэвид; Болл, Энди; Эвершед, Ник. «Мертвое море: подводные леса Тасмании исчезают при нашей жизни». хранитель. Получено 2020-10-22.
- ^ Штейнбауэр, Джеймс. «Что потребуется, чтобы вернуть лес водорослей? - журнал Bay Nature». Залив природа. Получено 2020-10-22.
- ^ Леса водорослей являются средой обитания для множества беспозвоночных, рыб, морских млекопитающих и птиц. NOAA. Обновлено 11 января 2013 г. Проверено 15 января 2014 г.
- ^ Джонс, К.Г., Дж. Х. Лоутон и М. Шахак. 1997. Положительные и отрицательные эффекты организмов как инженеров физических экосистем. Экология 78: 1946-1957.
- ^ а б Дрюль, Л. 1981. Распространение ламинариевых в северной части Тихого океана со ссылкой на влияние окружающей среды. Труды Международного конгресса по систематической эволюции и биологии 2: 248-256.
- ^ Уиллер, В.Н. 1980. Влияние переноса пограничного слоя на фиксацию углерода гигантскими водорослями. Macrocystis pyrifera. Морская биология 56: 103-110.
- ^ Стенек, Р. и М. Детье. 1994. Функционально-групповой подход к структуре сообществ с преобладанием водорослей. Ойкос 69: 476-498.
- ^ Жоли, А. и E.C. Oliveira Filho. 1967. Два бразильца Ламинарии. Instituto de Pesquisas da Marinha 4: 1-7.
- ^ Петров Ю.Е., М.В. Суховеева, Г.В. Авдеев. 1973 г. Новые виды рода. Ламинария из Филиппинского моря. Nov Sistem. Низч. Раст. 10: 59-61.
- ^ Сантеличе Б. 2007. Обнаружение лесов ламинарии в глубоководных местообитаниях тропических регионов. Труды Национальной академии наук Авана Риака 104: 19163-19164.
- ^ а б c Фостер, М. и Д. Schiel. 1985. Экология гигантских лесов водорослей в Калифорнии: профиль сообщества. Отчет Службы рыболовства и дикой природы США 85: 1-152.
- ^ а б Грэм, М. 2004. Влияние местной вырубки лесов на разнообразие и структуру пищевых сетей гигантских лесных водорослей Южной Калифорнии. Экосистемы 7: 341-357.
- ^ Фаулер-Уокер, М.Дж., Б.М. Гилландерс, С.Д. Коннелл и А.Д. Ирвинг. 2005. Паттерны ассоциации между морфологией полога и сообществами подлеска в Австралии с умеренным климатом. Эстуарии, прибрежные районы и шельфовые науки 63: 133-141.
- ^ Джексон, Г.А. 1977 г. Питательные вещества и производство гигантских водорослей, Macrocystis pyrifera, у южной Калифорнии. Лимнология и океанография 22: 979-995.
- ^ а б Дейтон, П.К. М.Дж. Тегнер, П. Эдвардс и К. Подъемник. 1999. Временные и пространственные масштабы демографии водорослей: роль океанографического климата. Экологические монографии 69: 219-250.
- ^ а б Карр, М. 1994. Влияние динамики макроводорослей на пополнение рифовых рыб умеренного пояса. Экология 75: 1320-1333.
- ^ а б Эстес, Дж. и делать. Даггинс. 1995. Морские каланы и леса водорослей на Аляске: общность и разнообразие экологической парадигмы сообщества. Экологические монографии 65: 75-100.
- ^ а б c Пирс, Дж. и А.Х. Хайнс. 1987 г.Расширение лесов водорослей в центральной Калифорнии после массовой гибели морских ежей. Морская биология 51: 83-91.
- ^ Scheibiling, R.E. и А. Хеннигар. 1997. Периодические вспышки болезней морских ежей. Strongylocentrotus droebachiensis в Новой Шотландии: свидетельство связи с крупномасштабными метеорологическими и океанографическими событиями. Серия «Прогресс морской экологии» 152: 155-165.
- ^ Велимиров Б., Ж. Филд, C.L. Гриффитс и П. Зутендык. 1977 г. Экология сообществ водорослей в бенгельской системе апвеллинга. Морские исследования Гельголанда 30: 495-518.
- ^ Андрей, Н. 1993. Пространственная неоднородность, выпас морских ежей и структура местообитаний рифов в Австралии с умеренным климатом. Экология 74: 292-302.
- ^ Дейтон, П.К. 1985b. Структура и регуляция некоторых южноамериканских сообществ ламинарии. Экологические монографии 55: 447-468.
- ^ а б c Сала, Э. и М.Х. Грэм. 2002. Распределение силы взаимодействия хищник-жертва в лесах ламинарии в масштабе сообщества. Слушания Национальной академии наук 99: 3678-3683.
- ^ Бирнс, Дж., Дж. Дж. Дж. Стахович, К. Халтгрен, А. Хьюз, С.В. Олярник и К.С.Торнбер. 2006. Разнообразие хищников усиливает трофические каскады в лесах ламинарии, изменяя поведение травоядных. Письма по экологии 9: 61-71.
- ^ а б Халперн, Б.С., К. Коттени, Б.Р. Бройтман. 2006. Сильный нисходящий контроль в экосистемах лесных водорослей Южной Калифорнии. Наука 312: 1230-1232.
- ^ Лоуренс, Дж. М. 1975. О взаимоотношениях между морскими растениями и морскими ежами. Ежегодный обзор океанографии и морской биологии. 13: 213-286.
- ^ Хьюз, Т. 1994. Катастрофы, фазовые сдвиги и крупномасштабная деградация Карибского кораллового рифа. Наука 265: 1547-1551.
- ^ Сиверстен, К. 2006. Чрезмерный выпас грядок водорослей вдоль побережья Норвегии. Журнал прикладной психологии 18: 599-610.
- ^ а б c d е Дейтон, П.К., М.Дж. Тегнер, П.Е. Парнелл и П. Эдвардс. 1992. Временные и пространственные закономерности нарушения и восстановления в сообществе ламинарии. Экологические монографии 62: 421-445.
- ^ Пирс, Дж. 2006. Экологическая роль пурпурных морских ежей. Наука 314: 940-941.
- ^ Лафферти, К. 2004. Промысел омаров косвенно увеличивает эпидемии морских ежей. Экологические приложения 14: 1566-1573.
- ^ а б Васкес, Дж. А., Дж. М. Алонсо Вега и А. Х. Бушманн. 2006. Долгосрочная изменчивость в структуре сообществ ламинарии на севере Чили и в ЭНСО 1997-98 гг. Журнал прикладной психологии 18: 505-519.
- ^ Cowen, R.K. 1983. Эффект овчарки (Семикоссифус пульчер) хищничество на красного морского ежа (Strongylocentrotus franciscanus) популяции: экспериментальный анализ. Oecologia 58: 249-255.
- ^ а б Эбелинг, A.W., D.R. Лаур и Р.Дж. Роули. 1985. Сильный шторм и изменение структуры сообщества в лесу водорослей южной Калифорнии. Морская биология 84: 287-294.
- ^ а б Дейтон, П.К. и М.Дж. Тегнер. 1984. Катастрофические штормы, Эль-Ниньо и стабильность пятен в сообществе водорослей южной Калифорнии. Наука 224: 283-285.
- ^ Харролд, К. и Д.К. Рид. 1985. Наличие пищи, выпас морских ежей и структура лесных сообществ ламинарии. Экология 66: 1160-1169.
- ^ Куп, К., Р.С. Ньюэлл и М. Лукас. 1982. Биодеградация и поток углерода на основе водорослей (Эклония максима) мусор в микромире песчаного пляжа. Серия «Прогресс морской экологии» 7: 315-326.
- ^ Бустаманте, Р.Х., Г.М. Бранч и С. Ихаут. 1995. Поддержание исключительной биомассы приливных травоядных животных в Южной Африке: субсидия сублиторальных водорослей. Экология 76: 2314-2329.
- ^ Kaehler, S., E.A. Пахомов, Р. Калин и С. Дэвис. 2006. Трофическая важность взвешенных твердых частиц, полученных из водорослей, в субантарктической системе. Серия «Прогресс морской экологии» 316: 17-22.
- ^ Григг, Р.В. и Р.С. Кивала. 1970. Некоторые экологические последствия сбрасываемых отходов на морскую жизнь. Калифорнийский департамент рыбы и дичи 56: 145-155.
- ^ Стулл, Дж. 1989. Загрязняющие вещества в отложениях вблизи крупного морского сброса: история, последствия и будущее. OCEANS ’89 Proceedings 2: 481-484.
- ^ Норт, W.J., D.E. Джеймс и Л. Джонс. 1993. История клумбы из водорослей (Макроцистис) в округах Ориндж и Сан-Диего, Калифорния. Hydrobiologia 260/261: 277-283.
- ^ Тегнер, М.Дж., П.К. Дейтон, П. Эдвардс, К. Райзер, Д. Чедвик, Т. Дин и Л. Дейшеры. 1995. Влияние большого разлива сточных вод на сообщества ламинарии: катастрофа или нарушение? Исследования морской среды 40: 181-224.
- ^ Карпентер, С.Р., Р.Ф. Карако, Д.Ф. Корнелл, Р.В. Ховарт, А.Н. Шарпли и В. Смит. 1998. Неточечное загрязнение поверхностных вод фосфором и азотом. Экологические приложения 8: 559-568.
- ^ Кеннелли, С.Дж. 1987. Физические нарушения в сообществе австралийских водорослей. I. Временные эффекты. Серия «Прогресс морской экологии» 40: 145-153.
- ^ Макфаден, М.Дж. 1999. Происхождение и эволюция Эль-Ниньо 1997–1998 годов. Наука 283: 950-954.
- ^ Эдвардс, М. и Г. Эрнандес-Кармона. 2005. Задержка восстановления гигантских водорослей вблизи южной границы ареала в северной части Тихого океана после Эль-Ниньо. Морская биология 147: 273-279.
- ^ Даггинс, Д.О., Дж.Э. Экман, А.Т. Сьюэлл. 1990. Экология среды обитания подлеска ламинарии. II. Влияние ламинарии на пополнение донных беспозвоночных. Журнал экспериментальной морской биологии и экологии 143: 27-45.
- ^ а б Джексон, J.B.C, M.X. Кирби, W.H. Бергер, К. Бьорндал, Л. Ботсфорд, Б.Дж.Бурк, Р.Х. Брэдбери, Р.Кук, Дж. Эрландсон, J.A. Estes, Т. Хьюз, С. Кидвелл, С.Б. Ланге, Х.С. Ленихан, Дж. М. Пандольфи, К. Х. Петерсон, Р. Стенек, М.Дж. Тегнер и Р.Р. Уорнер. 2002. Исторический перелов и недавний крах прибрежных экосистем. Наука 293: 629-638.
- ^ Ирвинг, А.Д. и С.Д. Коннелл. 2006. Прогнозирование структуры подлеска по наличию и составу пологов: правило сборки морских водорослей. Oecologia 148: 491-502.
- ^ Сименстад, К.А., J.A. Estes и К. Кеньон. 1978. Алеуты, каланы и альтернативные стабильно-государственные сообщества. Наука 200: 403-411.
- ^ Pringle Колонизировали ли люди мир с помощью лодок?
- ^ А. Гутьеррес, Т. Корреа, В. Муньос, А. Сантибаньес, Р. Маркос, К. Касерес и А. Х. Бушманн. 2006. Выращивание гигантских водорослей. Macrocystis pyrifera на юге Чили для разработки новых продуктов питания. Журнал прикладной психологии 18: 259-267.
- ^ Ортис, М. и В. Стоц. 2007. Экологические и эко-социальные модели интродукции морского ушка. Haliotis Discus hannai в бентосные системы северо-центральной части Чили: оценка устойчивости. Сохранение водных ресурсов: морские и пресноводные экосистемы 17: 89-105.
- ^ Scheffer, M., S. Carpenter, J.A. Фоли, К. Фолк и Б. Уолтер. 2001. Катастрофические сдвиги в экосистемах. Nature 413: 591-596.
- ^ Макдональд, Люси (06.02.2019). «95% гигантских водорослей Тасмании исчезли, ученые пытаются спасти то, что осталось». ABC News. Получено 2020-02-09.
- ^ Фрост Т.М., С.Р. Карпентер, А. Айвз, Т. Кратц. 1995. "Видовая компенсация и взаимодополняемость в функции экосистемы". В: К. Джонс и Дж. Лоутон, редакторы. Связывание видов и экосистем. Чепмен и Холл, Лондон. 387pp.
- ^ Тильман, Д., К. Lehman и C.E. Bristow. 1998. Взаимосвязь разнообразия и стабильности: статистическая неизбежность или экологические последствия? Американский натуралист 151: 277-282.
- ^ Стахович, Дж. Дж., Р. Б. Уитлатч и Р. В. Осман. 1999. Видовое разнообразие и устойчивость к инвазии в морской экосистеме. Наука 286: 1577-1579.
- ^ Элмквист Т., К. Фольке, М. Нистрем, Г. Петерсон, Дж. Бенгтссон, Б. Уокер и Дж. Норберг. 2003. Разнообразие ответных мер, изменение экосистемы и устойчивость. Границы экологии и окружающей среды 1: 488-494.
- ^ Стеколл, M.S., L.E. Дейшер и М. Гесс. 2006. Дистанционный подход к оценке биомассы ламинарии. Журнал прикладной психологии 18: 323-334.
- ^ Эллисон, Г.А., Дж. Любченко и М.Х. Карр. 1998. Морские заповедники необходимы, но недостаточны для сохранения морской среды. Экологические приложения 8: S79-S92.
- ^ Айраме, С., Дж. Э. Дуган, К. Д. Лафферти, Х. Лесли, Д.А. Макардл и Р.Р.Уорнер. 2003. Применение экологических критериев к проектированию морских заповедников: пример из Калифорнийских Нормандских островов. Экологические приложения 13: S170-S184.
- ^ Bohnsack, J.A. 1998. Применение морских заповедников в управлении рифовым рыболовством. Австралийский журнал экологии 23: 298-304.
- ^ Гелл, Ф. и К. Робертс. 2003. Выгоды за пределами границ: рыбные последствия морских заповедников. Тенденции в экологии и эволюции 18: 448-455.
- ^ Уиллис, Т.Дж., Р.Б. Миллар, Р.С. Бэбкок. 2003. Защита эксплуатируемой рыбы в регионах с умеренным климатом: высокая плотность и биомасса луциана. Pagrus auratus (Sparidae) в морских заповедниках на севере Новой Зеландии. Журнал прикладной экологии 40: 214-227.
- ^ Paddack, M.J. и J.A. Estes. 2000. Популяции лесных водорослей в морских заповедниках и прилегающих эксплуатируемых районах Центральной Калифорнии. Экологические приложения 10: 855-870.
- ^ Роджерс-Беннетт, Л. и Дж. Пирс. 2001. Косвенные преимущества охраняемых районов моря для молоди морского морского ушка. Биология сохранения 15: 642-647.
- ^ Бэбкок, Р.С., С. Келли, Н.Т. Ножницы, J.W. Уокер и Т.Дж. Уиллис. 1999. Изменения в структуре сообществ в морских заповедниках умеренного пояса. Серия «Прогресс морской экологии» 189: 125-134.
- ^ Халперн, Б. и Р.Р. Уорнер. 2002. Морские запасы оказывают быстрое и длительное воздействие. Письма по экологии 5: 361-366.
внешняя ссылка
- "Лес водорослей и скалистые сублиторальные места обитания". noaa.gov. Архивировано из оригинал 22 марта 2007 г.
- "Часы водорослей". tas.gov.au. Тасмания, Австралия: Департамент сырьевых отраслей, водных ресурсов и окружающей среды. Архивировано из оригинал на 2004-12-04. Отличная общая информация о лесах ламинарии, а также конкретная информация о лесах ламинарии Тасмании.
- "Камеру водорослей в аквариуме Монтерей-Бей". mbayaq.org. Аквариум Монтерей Бэй. Архивировано из оригинал на 1999-11-28. Посмотрите прямую трансляцию с выставки ламинарии.


| Авторитетный контроль |
|---|