Морские беспозвоночные - Marine invertebrates

Морские беспозвоночные являются беспозвоночные которые живут в морская среда обитания. Беспозвоночные - это общий срок это включает в себя все животные отдельно от позвоночное животное члены хордовый тип. Беспозвоночные не хватает позвоночник, а некоторые разработали ракушка или жесткий экзоскелет. Как на суше, так и в воздухе, морские беспозвоночные обладают большим разнообразием. планы тела, и были отнесены к более чем 30 категориям тип. Они составляют большую часть макроскопической жизни в океанах.
Эволюция
Раннее животные были морскими беспозвоночные, то есть, позвоночные пришел позже. Животные многоклеточный эукариоты,[примечание 1] и отличаются от растений, водорослей и грибов отсутствием клеточные стенки.[1] Морские беспозвоночные - это животные, населяющие морской окружающей среды, кроме позвоночных членов хордовый тип; беспозвоночные не имеют позвоночник. Некоторые разработали ракушка или жесткий экзоскелет.
Самые ранние животные могут принадлежать к роду Дикинсония,[2] От 571 миллиона до 541 миллиона лет назад.[3] Индивидуальный Дикинсония обычно напоминают двусторонне симметричный ребристый овал. Они продолжали расти, пока не покрылись осадком или не умерли иным образом,[4] и провели большую часть своей жизни, твердо прикрепив свои тела к отложениям.[5] Их таксономическое сходство в настоящее время неизвестны, но их способ роста соответствует двухсторонний близость.[6]
Помимо Дикинсония, самые ранние общепринятые окаменелости животных выглядят довольно современно. книдарийцы (группа, в которую входят медуза, морские анемоны и Гидра ), возможно, откуда-то 580 Ма[7] В Ediacara биота, который процветал последние 40 миллионов лет до начала Кембрийский,[8] были первыми животными длиной более нескольких сантиметров. Нравиться Дикинсония, многие из них были плоскими с «стеганым» видом и казались настолько странными, что было предложено отнести их к отдельным Королевство, Vendozoa.[9] Другие, однако, были истолкованы еще моллюски (Кимберелла[10][11]), иглокожие (Аркаруа[12]), и членистоногие (Spriggina,[13] Парванкорина[14]). До сих пор ведутся споры о классификации этих образцов, главным образом потому, что диагностические признаки, позволяющие систематикам классифицировать более современные организмы, такие как сходство с живыми организмами, как правило, отсутствуют у эдиакарцев. Однако мало сомнений в том, что Кимберелла был по крайней мере триплобластический bilaterian животное, другими словами, животное значительно более сложное, чем книдарии.[15]
В фауна малых ракушек представляют собой очень смешанную коллекцию окаменелостей, найденных между поздним эдиакарским периодом и Средний кембрий периоды. Раннее, Cloudina, показывает признаки успешной защиты от хищников и может указывать на начало эволюционная гонка вооружений. Некоторые крошечные ракушки раннего кембрия почти наверняка принадлежали моллюскам, а владельцы некоторых «броневых пластин» - Халкирия и Микродиктион, были в конечном итоге идентифицированы, когда в кембрийском периоде были найдены более полные образцы. lagerstätten сохранившие мягкотелые животные.[16]

Дикинсония может быть самым ранним животным. Они появляются в летописи окаменелостей 571-541 миллион лет назад.

Кимберелла, ранний моллюск, важный для понимания Кембрийский взрыв. Беспозвоночные сгруппированы в разные тип (планы тела ).

Опабиния, вымерший стволовая группа членистоногие появился в Средний кембрий[17]:124–136



В 1970-х годах уже велись дебаты о том, было ли появление современных типов «взрывным» или постепенным, но скрытым нехваткой Докембрийский окаменелости животных.[16] Повторный анализ окаменелостей из Burgess Shale lagerstätte повысил интерес к этому вопросу, когда раскрыл животных, таких как Опабиния, которые не укладывались ни в один известный филюм. В то время это было истолковано как свидетельство того, что современные типы эволюционировали очень быстро во время кембрийского взрыва и что «странные чудеса» сланцев Берджесс показали, что ранний кембрий был уникальным экспериментальным периодом эволюции животных.[17] Более поздние открытия похожих животных и развитие новых теоретических подходов привели к выводу, что многие из «странных чудес» были эволюционными «тетками» или «кузенами» современных групп.[18]- например, что Опабиния был членом лобоногие, группа, которая включает предков членистоногих, и что она, возможно, была тесно связана с современными тихоходки.[19] Тем не менее, до сих пор ведется много споров о том, был ли кембрийский взрыв действительно взрывным, и если да, то как и почему он произошел и почему он кажется уникальным в истории животных.[20]
Классификация

Беспозвоночные сгруппированы в разные тип. Неформально типы можно рассматривать как способ группировки организмов в соответствии с их план тела.[21][22]:33 План тела относится к плану, который описывает форму или морфология организма, например, его симметрия, сегментация и расположение его придатки. Идея планов тела возникла с позвоночные, которые были сгруппированы в один тип. Но план тела позвоночных - лишь один из многих, а беспозвоночные состоят из множества типов или планов тела. Историю открытия планов тела можно рассматривать как движение от мировоззрения, сосредоточенного на позвоночных, к рассмотрению позвоночных как одного строения тела среди многих. Среди новаторских зоологи Линней выделил два строения тела за пределами позвоночных; Кювье выделил троих; у Геккеля было четыре, а у Протиста - еще восемь, всего двенадцать. Для сравнения, количество типов, признанных современными зоологами, составляет поднялся до 35.[22]
Исторически считалось, что планы тела быстро развивались в течение Кембрийский взрыв,[23] но более детальное понимание эволюции животных предполагает постепенное развитие планов тела на раннем этапе Палеозойский и дальше.[24] В более общем смысле тип может быть определен двумя способами: как описано выше, как группа организмов с определенной степенью морфологического сходства или сходства в развитии ( фенетический определение), или группа организмов с определенной степенью эволюционного родства ( филогенетический определение).[24]
Как на суше, так и в воздухе, беспозвоночные составляют подавляющее большинство всей макроскопической жизни, поскольку позвоночные составляют подтип одного из более чем 30 известных типов животных, что делает этот термин почти бессмысленным для таксономический цель. Морская жизнь беспозвоночных включает следующие группы, некоторые из которых относятся к типам:


- Acoela, среди самых примитивных двусторонние животные;
- Аннелида, (полихеты и море пиявки );
- Брахиоподы, морские животные, имеющие твердые «створки» (раковины) на верхней и нижней поверхностях;
- Мшанки, также известные как моховые животные или морские циновки;
- Chaetognatha, широко известные как черви-стрелы, представляют собой тип хищных морских червей, которые являются основным компонентом планктона;
- Головнохордовые представлен в современных океанах ланцетники (также известный как Амфиоксус);
- Книдария, Такие как медуза, морские анемоны, и кораллы;
- Ракообразные, включая лобстеры, крабы, креветка, рак, ракушки, раки-отшельники, креветки-богомолы, и копеподы;
- Гребневик, также известные как гребешки, крупнейшие животные, плавающие с помощью ресничек;
- Иглокожие, включая морские звезды, хрупкие звезды, морские ежи, песочные доллары, морские огурцы, морские лилии, и морские ромашки;
- Эчиура, также известные как ложные черви;
- Гнатостомулиды тонкие до нитевидных червей с прозрачным телом, обитающие в песке и иле под мелкими прибрежными водами;
- Гастротрича, часто называемые волосатыми спинками, обнаруживаются в основном между частицами осадка;
- Hemichordata, включает желудевые черви, одиночные червеобразные организмы;
- Kamptozoa сидячие водные животные чашевидной формы с относительно длинными стеблями и «короной» из твердых щупалец, также называемые Entoprocta;
- Киноринча сегментированные животные без конечностей, широко распространенные в грязи или песке на любой глубине, также называемые грязевыми драконами;
- Лорицифера от очень маленьких до микроскопических морских обитающих в донных отложениях животных, обнаруженных только в 1983 году;
- Моллюска, включая моллюски, Кальмар, осьминог, щенячки, Наутилус, каракатица, голожаберники, гребешки, морские улитки, Аплакофора, Каудофовеата, Моноплакофора, Полиплакофора, и Scaphopoda;
- Мизостомида таксономическая группа мелких морских червей, паразитирующих на морских лилиях или «морских лилиях»;
- Немертинея, также известные как «ленточные черви» или «хоботковые черви»;
- Ортонектида, небольшой тип малоизвестных паразитов морских беспозвоночных, которые относятся к числу простейших многоклеточных организмов;
- Phoronida, тип морских животных, которые фильтруют корм с помощью лофофора («корона» из щупалец) и строят вертикальные трубки из хитина, чтобы поддерживать и защищать свои мягкие тела;
- Placozoa, маленькие, плоские, многоклеточные животные диаметром около 1 миллиметра и простейшие по строению. У них нет правильного очертания, хотя нижняя поверхность несколько вогнута, а верхняя всегда уплощена;
- Porifera (губки), многоклеточные организмы, тела которых имеют поры и каналы, позволяющие воде циркулировать через них;
- Приапулида, или черви пениса, представляют собой тип морских червей, которые живут в морской грязи. Они названы в честь их расширяемого шиповатого хоботка, который у некоторых видов может иметь форму, подобную форме человеческого пениса;
- Пикногонида, также называемые морскими пауками, не связаны с пауками или даже с паукообразными, на которых они похожи;
- Сипункулида, также называемые арахисовыми червями, представляют собой группу, содержащую 144–320 видов (по разным оценкам) двусторонне-симметричных несегментированных морских червей;
- Туниката, также известные как морские брызги или морская свинина, представляют собой фильтрующие устройства, прикрепленные к камням или аналогичным подходящим поверхностям на дне океана;
- Немного плоские черви классов Турбеллария и Моногенея;
- Ксенотурбелла род билатерианских животных, который включает только два морских червеобразных вида;
- Ксифосура, включает большое количество вымерших линий и только четыре современных вида в семействе Limulidae, которые включают подковообразные крабы.
Членистоногие всего около 1113000 описанных современных видов, моллюски около 85000 и хордовые около 52000.[25]
Морские губки

Губки животные из филюм Porifera (современная латынь для несущие поры [26]). Это многоклеточные организмы, тела которых заполнены порами и каналами, позволяющими воде циркулировать через них, состоящие из желеобразных мезохил зажатый между двумя тонкими слоями клетки. У них есть неспециализированные ячейки, которые могут превращаться в другие типы и которые часто мигрируют между основными клеточными слоями и мезохилом в процессе. Губки не имеют нервный, пищеварительный или же системы кровообращения. Вместо этого большинство из них полагается на поддержание постоянного потока воды через свое тело для получения пищи и кислорода и удаления отходов.
Губки похожи на других животных в том, что они многоклеточный, гетеротрофный, недостаток клеточные стенки и производить сперматозоиды. В отличие от других животных, им не хватает истинного ткани и органы, и нет симметрия тела. Формы их тел адаптированы для максимальной эффективности потока воды через центральную полость, где она откладывает питательные вещества и выходит через отверстие, называемое оскулюм. Многие губки имеют внутренние скелеты губка и / или спикулы карбонат кальция или же диоксид кремния. Все губки сидячий водные животные. Хотя есть пресноводные виды, подавляющее большинство из них - морские (соленые), от приливных зон до глубин, превышающих 8800 м (5,5 миль).
Хотя большинство из примерно 5000–10 000 известных видов питаются бактерии и другие частицы пищи в воде, некоторые фотосинтез микроорганизмы как эндосимбионты и эти союзы часто производят больше еды и кислорода, чем потребляют. Некоторые виды губок, обитающие в условиях скудной пищи, стали плотоядные животные что охотятся в основном на мелких ракообразные.[27]

Биоразнообразие губок. На этом фото четыре вида губок.

Цветочная корзина Венеры на глубине 2572 метра





Линней ошибочно идентифицировали губки как растения в порядке Водоросли.[28] На долгое время после этого губки были выделены в отдельное царство, Паразоа (смысл рядом с животными).[29] Теперь они классифицируются как парафилетический филюм из которого произошли высшие животные.[30]
Морские книдарии

Книдарианцы (По-гречески крапива) отличаются наличием жалящие клетки, специализированные клетки, которые они используют в основном для поимки добычи. Книдарианцы включают кораллы, морские анемоны, медуза и гидрозоаны. Они образуют филюм содержащий более 10 000[31] разновидность из животные встречается исключительно в водной (в основном морской) среде. Их тела состоят из мезоглея, неживое желеобразное вещество, зажатое между двумя слоями эпителий это в основном один толстая ячейка. У них две основные формы тела: плавание медузы и сидячий полипы, оба из которых радиально симметричный с ртом, окруженным щупальцами, несущими книдоциты. Обе формы имеют один отверстие и полости тела, которые используются для пищеварения и дыхание.
Ископаемые книдарии были обнаружены в породах, сформированных около 580 миллион лет назад. Окаменелости книдарийцев, которые не строят минерализованный структуры редки. В настоящее время ученые думают, что книдарианцы, гребневики и билатерии более тесно связаны с известковые губки чем они к другим губки, и это антозои являются эволюционными «тетками» или «сестрами» других книдарий и наиболее тесно связаны с билатерианами.
Книдарианцы являются простейшими животными, у которых клетки организованы в ткани.[32] В звездочка морской анемон используется как модельный организм в исследованиях.[33] За ним легко ухаживать в лаборатории и протокол был разработан, который может давать большое количество эмбрионов ежедневно.[34] Существует замечательная степень сходства в сохранности и сложности последовательности генов морского анемона и позвоночных.[34] В частности, гены, участвующие в формировании головы у позвоночных, также присутствуют в анемоне.[35][36]

Морские анемоны распространены в водоемы

Их щупальца ужалить и парализовать рыбку

Если остров опускается ниже уровня моря, рост кораллов может идти в ногу с подъемом воды и образовывать атолл





В биолюминесценция морской орех имеет преходящий анус[38] который образуется только тогда, когда ему нужно испражняться

Turritopsis dohrnii достигает биологическое бессмертие перенося свои клетки обратно в детство [39][40]




Морские черви
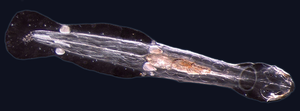
черви (Древнеанглийский для змея) обычно имеют длинные цилиндрические трубчатые тела и не имеют конечности. Морские черви различаются по размеру от микроскопических до более 1 метра (3,3 фута) в длину для некоторых морских многощетинковых червей (щетинистые черви )[41] и до 58 метров (190 футов) для морского немертинского червя (ботинок червь ).[42] Некоторые морские черви занимают небольшое количество паразитический ниши, обитающие внутри тел других животных, в то время как другие живут более свободно в морской среде или роясь под землей.
Различные группы морских червей связаны между собой лишь отдаленно, поэтому они встречаются в нескольких разных тип такой как Аннелида (сегментированные черви), Chaetognatha (стреловидные черви), Hemichordata, и Phoronida (подковообразные черви). Многие из этих червей имеют специальные щупальца, используемые для обмена кислорода и углекислого газа, а также могут использоваться для размножения. Некоторые морские черви трубчатые черви, такой как гигантский трубчатый червь который живет в водах рядом с водой вулканы и выдерживает температуру до 90 градусов Цельсия.
Платигельминты (плоские черви) образуют другой тип червей, который включает в себя класс Цестода паразитических ленточных червей. Морской цепень Полигонопор гигантский, найденный в кишечнике кашалоты, может вырасти до более 30 м (100 футов).[43][44]
Нематоды (круглые черви) составляют следующий тип червей с трубчатыми пищеварительная система и отверстие на обоих концах.[45][46] Описано более 25000 видов нематод,[47][48] из которых более половины являются паразитическими. Было подсчитано, что еще миллион остаются неописанными.[49] Они повсеместно распространены в морской, пресноводной и наземной среде, где часто превосходят по численности других животных как по индивидуальным, так и по видовым показателям. Они встречаются во всех уголках Земли. литосфера, от вершины гор до подножия океанические желоба.[50] По подсчетам они составляют 90% всех животных на дно океана.[51] Их численное преобладание, часто превышающее миллион особей на квадратный метр и составляющее около 80% всех индивидуальных животных на Земле, разнообразие их жизненных циклов и их присутствие на различных трофических уровнях указывают на важную роль во многих экосистемах.[52]

Гигантские трубчатые черви группа вокруг гидротермальные источники

Lamellibrachia luymes, а холодная утечка трубочистка, живет более 250 лет.

Нематоды повсеместно псевдоэломаты который может паразитировать на морских растениях и животных.

Мотыля обычно встречаются на дне мелководных морских вод

Черви Bobbit находятся устраивать засады на хищников которые живут на морском дне





Иглокожие

Иглокожие (По-гречески колючая кожа) - это тип, который содержит только морских беспозвоночных. Взрослых можно узнать по их радиальная симметрия (обычно пятибалльный) и включают морская звезда, морские ежи, песочные доллары, и морские огурцы, так же хорошо как морские лилии.[53] Иглокожие встречаются на любой глубине океана, начиная с приливная зона к абиссальная зона. Тип насчитывает около 7000 живых разновидность,[54] что делает его второй по величине группой дейтеростомы (суперфильм), после хордовые (которые включают позвоночные, Такие как птицы, Рыбы, млекопитающие, и рептилии ).
Иглокожие уникальны среди животных тем, что имеют двустороннюю симметрию на личиночной стадии, но пятикратную симметрию (пентамеризм, особый тип радиальной симметрии) у взрослых.[55]
Иглокожие важны как биологически, так и геологически. С биологической точки зрения существует несколько других группировок, столь многочисленных в биотическая пустыня из глубокое море, а также более мелкие океаны. Большинство иглокожих способны возродить ткани, органы, конечности и воспроизводить бесполым путем; в некоторых случаях они могут пройти полную регенерацию из одной конечности. Геологически ценность иглокожих заключается в их окостеневший скелеты, которые вносят основной вклад во многие известняк формаций и может дать ценные ключи к разгадке геологической среды. Они были наиболее часто используемыми видами в регенеративных исследованиях в 19 и 20 веках. Кроме того, некоторые ученые считают, что радиация иглокожих была причиной Мезозойская морская революция.

Иглокожие буквально означает «колючая кожа», так как это арбуз морской еж иллюстрирует

Морские огурцы фильтровать питание планктона и взвешенных веществ

Бентопелагические морские огурцы может подняться с морского дна и подняться на высоту до 1000 м (3300 футов) по толщине воды

В охра морская звезда был первым краеугольный камень хищник быть изученным. Они ограничивают моллюски которые могут захлестнуть приливные сообщества.[56]

Красочный морские лилии на мелководье





Помимо трудно классифицируемых Аркаруа (а Докембрийский животное с пятичленной радиальной симметрией, напоминающей иглокожие), первые дефинитивные члены филума появились около начала Кембрийский.
Морские моллюски

Моллюски (Латинское для мягкий) образуют филюм около 85 000 сохранившийся признанный разновидность.[57] По количеству видов они самые крупные морской тип, составляющий около 23% всех названных морских организмы.[58] Моллюски имеют более разнообразные формы, чем другие типы беспозвоночных. Они очень разнообразны не только по размеру, но и по анатомический структура, но также в поведении и в среде обитания. Большинство видов до сих пор обитают в океанах, от побережья до абиссальная зона, но некоторые образуют значительная часть пресноводной фауны и земной экосистемы.
Тип моллюсков делится на 9 или 10 таксономический классы, два из которых вымерший. Эти классы включают брюхоногие моллюски, двустворчатые моллюски и головоногие моллюски, а также другие менее известные, но отличительные классы. Брюхоногие моллюски с защитными оболочками называются улитки (морские улитки ), тогда как брюхоногие моллюски без защитных панцирей называются слизни (морские слизни ). Брюхоногие моллюски являются самыми многочисленными моллюсками с точки зрения классифицированных видов, составляя 80% от общего числа.[25] Двустворчатые моллюски включают моллюски, устрицы, моллюски, моллюски, гребешки, и многие другие семьи. Насчитывается около 8000 видов морских двустворчатых моллюсков (в том числе солоноватая вода и эстуарий видов) и около 1200 пресноводных видов. Головоногие моллюски включают осьминог, Кальмар и каракатица. Они встречаются во всех океанах, и неврологически являются наиболее развитыми из беспозвоночных.[59] Выявлено около 800 ныне живущих видов морских головоногих моллюсков,[60] и примерно 11000 вымерших таксоны были описаны.[61] Полностью пресноводных головоногих моллюсков нет.[62]

Колоссальный кальмар, самый крупный из всех беспозвоночных [63]

В наутилус это живое ископаемое мало что изменилось с тех пор, как он появился 500 миллионов лет назад как один из первых головоногие моллюски.[64][65][66]

морской брюхоногие моллюски находятся морские улитки или же морские слизни. Этот голожаберный это морской слизень.

Морская улитка Сиринкс аруанский имеет самый большой панцирь среди всех живых брюхоногих моллюсков

У моллюсков обычно есть глаза. Граничит с краем мантии гребешок, а двустворчатый моллюск, может быть более 100 простые глаза.

Мидия обыкновенная, другой двустворчатый моллюск





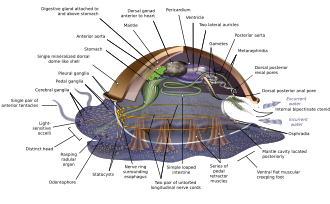
Моллюски имеют настолько разнообразные формы, что многие учебники основывают свои описания анатомии моллюсков на обобщенных или гипотетический предковый моллюск. Этот генерализованный моллюск не сегментирован и двусторонне симметричный с нижней стороной, состоящей из одного мускульного оплачивать.[67][68]:484–628 Помимо этого, у него есть еще три ключевые особенности. Во-первых, у него мускулистый плащ, называемый мантия покрывающий его внутренности и содержащий значительную полость, используемую для дыхания и выделение. А ракушка секретируется мантией, покрывает верхнюю поверхность.[68] Во-вторых (кроме двустворчатых) у него есть грубый язык, называемый радула используется для кормления. В-третьих, у него есть нервная система включая сложную пищеварительную систему с микроскопическими мышечными волосками, называемыми реснички источать слизь. У генерализованного моллюска два парных нервные шнуры (три у двустворчатых моллюсков). В мозг, у видов, которые имеют один, обводит пищевод. Большинство моллюсков имеют глаза и все они оснащены датчиками, определяющими химические вещества, вибрацию и прикосновение. Самый простой вид моллюсков репродуктивная система полагается на внешнее оплодотворение, но встречаются и более сложные варианты. Все производят яйца, из которых может возникнуть трохофор личинки, более сложный велигер личинки или миниатюрные взрослые особи. Изображение очень похоже на современное. моноплакофораны, и некоторые предполагают, что он может напоминать очень ранних моллюсков.[67]:284–291[67]:298–300[69][70]
Существуют веские доказательства появления морских брюхоногих моллюсков, головоногие моллюски и двустворчатые моллюски в Кембрийский период 541 до 485,4 миллион лет назад. Однако эволюционная история появления обоих моллюсков из предковых Лофотрохозоа и об их разнообразии в хорошо известные жизни и ископаемое формы до сих пор являются предметом жарких споров среди ученых.
Морские членистоногие

Членистоногие (По-гречески шарнирные ноги) есть экзоскелет (внешний скелет ), а сегментированный корпус и сочлененный придатки (парные придатки). Они образуют филюм который включает насекомые, паукообразные, многоножки, и ракообразные. Для членистоногих характерны суставные конечности и кутикула сделано из хитин, часто минерализованный карбонат кальция. Членистоногие план тела состоит из сегменты, каждый с парой придатки. Жесткая кутикула тормозит рост, поэтому членистоногие периодически заменяют ее линька. Их универсальность позволила им стать самыми богатыми видами среди всех экологические гильдии в большинстве сред.
Размеры морских членистоногих варьируются от микроскопических. ракообразный Стиготантул к Японский краб-паук. Первичная внутренняя полость членистоногих - это гемоцель, который вмещает их внутренние органы, и через которые их гемолимфа - аналог кровь - циркулирует; у них есть открытые системы кровообращения. Как и их внешность, внутренние органы членистоногих обычно состоят из повторяющихся сегментов. Их нервная система «лестничный», с парными вентральный нервные шнуры прохождение всех сегментов и формирование парных ганглии в каждом сегменте. Их головы образованы слиянием различного количества сегментов, а их мозги образуются путем слияния ганглиев этих сегментов и охватывают пищевод. В респираторный и выделительный системы членистоногих различаются в зависимости от среды обитания и подтип к которому они принадлежат.
Их видение основано на различных комбинациях сложные глаза и пигментная яма глазки: у большинства видов глазки могут определять только направление, с которого исходит свет, и сложные глаза являются основным источником информации, но главные глаза пауки это глазки, которые могут формировать изображения и, в некоторых случаях, могут поворачиваться, чтобы отслеживать добычу. У членистоногих также есть широкий спектр химических и механических датчиков, в основном основанных на модификациях многих щетинки (щетина), выступающие из кутикулы. Способы размножения и развития членистоногих разнообразны; все наземные виды используют внутреннее оплодотворение, но это часто происходит путем косвенной передачи спермы через придаток или землю, а не путем прямой инъекции. Все морские виды откладывают яйца и используют либо внутренние, либо внешнее оплодотворение. Птенцы членистоногих варьируются от миниатюрных взрослых особей до личинок, у которых нет суставных конечностей и которые в конечном итоге полностью исчезают. метаморфоза произвести взрослую форму.

Подковообразный краб, а живое ископаемое членистоногие 450 миллионов лет назад

- Ракообразные

Многие рачки очень маленькие, как этот крошечный амфипода, и составляют значительную часть океана зоопланктон

В полосатая креветка-очиститель рачок, распространенный в тропиках.

В Тасманский гигантский краб является долгожителем и медленно растет, что делает его уязвимым для перелова.[72]

В Японский краб-паук имеет самый длинный размах ног среди всех членистоногих.




Эволюционное происхождение членистоногих восходит к Кембрийский период. Группа обычно рассматривается как монофилетический, и многие анализы подтверждают размещение членистоногих с циклоневры (или составляющие их клады) в суперфилуме Экдизозоа. Однако в целом базальный отношения Metazoa еще не решены. Точно так же до сих пор активно обсуждаются отношения между различными группами членистоногих.
Другой тип
- Тихоходка, Лобоподия, (Онихофора )
- Нечерепные (беспозвоночные) хордовые: Цефалохордовые, Туниката и Haikouella. Эти беспозвоночные - близкие родственники позвоночных.
- Нечерепные хордовые - близкие родственники позвоночных.

В ланцетник, небольшая полупрозрачная рыба, похожая на Цефалохордовые, является ближайшим живым беспозвоночным родственником позвоночных.[73][74]

Флуоресцентные морские брызги, Rhopalaea crassa. Оболочки могут дать ключ к разгадке происхождения позвоночных (и, следовательно, человека).[75]

Салп цепь
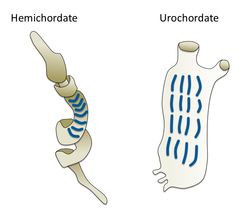
Жаберные щели в желудевом черве (слева) и оболочке (справа)




Минералы из морской воды
Есть ряд морских беспозвоночных, которые используют минералы, присутствующие в море в таких ничтожных количествах, что их нельзя было обнаружить до появления спектроскопия. Ванадий сосредоточено некоторыми оболочки для использования в их кровяных тельцах до уровня, в десять миллионов раз превышающего уровень окружающей морской воды. Другие оболочники аналогичным образом концентрируют ниобий и тантал.[67]:947 Использование омаров медь в их дыхательном пигменте гемоцианин несмотря на то, что доля этого металла в морской воде мала.[67]:638 Хотя эти элементы присутствуют в океане в огромных количествах, их добыча человеком нерентабельна.[76]
Смотрите также
- Стеклянные морские существа
- морская жизнь
- Морская биология
- Морское позвоночное животное
- Список видов морских аквариумных беспозвоночных
- Wiwaxia
Примечания
- ^ Myxozoa считались исключением, но теперь считаются сильно модифицированными членами Книдария. Хименес-Гури, Ева; Филипп, Эрве; Окамура, Бет; Голландия, Питер В. Х. (6 июля 2007 г.). "Будденброкия Книдарианский червь ". Наука. 317 (5834): 116–118. Bibcode:2007Sci ... 317..116J. Дои:10.1126 / science.1142024. ISSN 0036-8075. PMID 17615357. S2CID 5170702.CS1 maint: ref = harv (связь)
Рекомендации
- ^ Дэвидсон, Майкл В. (26 мая 2005 г.). «Структура клетки животных». Молекулярные выражения. Таллахасси, Флорида: Университет штата Флорида. Получено 2008-09-03.
- ^ Фогель, Гретхен (20.09.2018). «Эта окаменелость - одно из самых ранних животных в мире, если судить по молекулам жира, сохранившимся полмиллиарда лет». Наука. AAAS. Получено 21 сентября 2018.
- ^ Бобровский, Илья (2018). «Древние стероиды сделали эдиакарскую ископаемую Дикинсонию одним из самых ранних животных». Наука. 361 (6408): 1246–1249. Bibcode:2018Научный ... 361.1246B. Дои:10.1126 / science.aat7228. PMID 30237355.
- ^ Retallack, G.J. (2007). «Рост, распад и уплотнение погребения Дикинсонии, культовой ископаемой эдиакарской останки» (PDF). Алчеринга: Австралазийский журнал палеонтологии. 31 (3): 215–240. Дои:10.1080/03115510701484705. S2CID 17181699.
- ^ Сперлинг, Эрик; Винтер, Якоб; Пизани, Давиде; Петерсон, Кевин (2008). "Плакозойское сродство к Дикинсония и эволюция способов питания позднедокембрийских многоклеточных животных " (PDF). In Cusack, M; Оуэн, А; Кларк, Н. (ред.). Программа с тезисами. Ежегодное собрание палеонтологической ассоциации. 52. Глазго, Великобритания. п. 81.
- ^ Голд, Д. А .; Runnegar, B .; Gehling, J. G .; Джейкобс, Д. К. (2015). «Реконструкция онтогенетического состояния предков поддерживает двуногую близость к Dickinsonia». Эволюция и развитие. 17 (6): 315–397. Дои:10.1111 / ede.12168. PMID 26492825.
- ^ Цзюнь-Юань Чен; Оливери, Паола; Фэн Гао; и другие. (1 августа 2002 г.). «Жизнь докембрийских животных: вероятные формы развития и взрослых книдарий из Юго-Западного Китая» (PDF). Биология развития. 248 (1): 182–196. Дои:10.1006 / dbio.2002.0714. ISSN 0012-1606. PMID 12142030. Архивировано из оригинал (PDF) 26 мая 2013 г.. Получено 2015-02-04.CS1 maint: ref = harv (связь)
- ^ Гражданкин, Дима (июнь 2004 г.). «Модели распространения в эдиакарских биотах: фации против биогеографии и эволюции». Палеобиология. 30 (2): 203–221. Дои:10.1666 / 0094-8373 (2004) 030 <0203: PODITE> 2.0.CO; 2. ISSN 0094-8373.CS1 maint: ref = harv (связь)
- ^ Сейлачер, Адольф (Август 1992 г.). "Вендобионта и Псаммокораллия: утраченные конструкции докембрийской эволюции". Журнал геологического общества. 149 (4): 607–613. Bibcode:1992JGSoc.149..607S. Дои:10.1144 / gsjgs.149.4.0607. ISSN 0016-7649. S2CID 128681462. Получено 2015-02-04.CS1 maint: ref = harv (связь)
- ^ Мартин, Марк В .; Гражданкин, Дмитрий В .; Bowring, Samuel A .; и другие. (5 мая 2000 г.). «Возраст неопротерозойского билатерианского тела и ископаемых остатков, Белое море, Россия: последствия для эволюции многоклеточных животных». Наука. 288 (5467): 841–845. Bibcode:2000Sci ... 288..841M. Дои:10.1126 / science.288.5467.841. ISSN 0036-8075. PMID 10797002. S2CID 1019572.CS1 maint: ref = harv (связь)
- ^ Федонкин Михаил А .; Ваггонер, Бенджамин М. (28 августа 1997 г.). "Окаменелости позднего докембрия Кимберелла представляет собой моллюскоподобный двуногий организм ". Природа. 388 (6645): 868–871. Bibcode:1997Натура.388..868F. Дои:10.1038/42242. ISSN 0028-0836. S2CID 4395089.CS1 maint: ref = harv (связь)
- ^ Муи, Рич; Дэвид, Бруно (декабрь 1998 г.). "Эволюция в причудливом типе: гомологии первых иглокожих" (PDF). Американский зоолог. 38 (6): 965–974. Дои:10.1093 / icb / 38.6.965. ISSN 1540-7063. Получено 2015-02-05.CS1 maint: ref = harv (связь)
- ^ Макменамин, Марк А.С. (Сентябрь 2003 г.). Spriggina трилобитоидный экдизозойный. Geoscience Horizons Сиэтл 2003. Рефераты с программами. 35. Боулдер, Колорадо: Геологическое общество Америки. п. 105. OCLC 249088612. Получено 2007-11-24.CS1 maint: ref = harv (связь) Доклад № 40-2, представленный на ежегодном собрании Геологического общества Америки в Сиэтле в 2003 г. (2–5 ноября 2003 г.) 2 ноября 2003 г. Конгресс-центр штата Вашингтон.
- ^ Джих-Пай Линь; Гон, Самуэль М., III; Гелинг, Джеймс Дж .; и другие. (2006). "А Парванкорина-подобное членистоногое из кембрия Южного Китая ». Историческая биология: международный журнал палеобиологии. 18 (1): 33–45. Дои:10.1080/08912960500508689. ISSN 1029-2381. S2CID 85821717.
- ^ Баттерфилд, Николас Дж. (Декабрь 2006 г.). «Зацепление некоторых« червей »стеблевой группы: ископаемых лофотрохозоидов в сланцах Берджесс». BioEssays. 28 (12): 1161–1166. Дои:10.1002 / bies.20507. ISSN 0265-9247. PMID 17120226. S2CID 29130876.CS1 maint: ref = harv (связь)
- ^ а б Бенгтсон 2004, стр. 67–78
- ^ а б Гулд, Стивен Джей (1990) Прекрасная жизнь: сланцы Берджесс и природа истории W. W. Norton. ISBN 9780393307009.
- ^ Бадд, Грэм Э. (Февраль 2003 г.). «Кембрийская летопись окаменелостей и происхождение филы» (PDF). Интегративная и сравнительная биология. 43 (1): 157–165. Дои:10.1093 / icb / 43.1.157. ISSN 1557-7023. PMID 21680420. Получено 2015-02-06.CS1 maint: ref = harv (связь)
- ^ Бадд, Грэм Э. (март 1996 г.). «Морфология Opabinia regalis и реконструкция стволовой группы членистоногих». Lethaia. 29 (1): 1–14. Дои:10.1111 / j.1502-3931.1996.tb01831.x. ISSN 0024-1164.CS1 maint: ref = harv (связь)
- ^ Маршалл, Чарльз Р. (май 2006 г.). «Объяснение кембрийского« взрыва »животных». Ежегодный обзор наук о Земле и планетах. 34: 355–384. Bibcode:2006AREPS..34..355M. Дои:10.1146 / annurev.earth.33.031504.103001. ISSN 1545-4495. S2CID 85623607.CS1 maint: ref = harv (связь)
- ^ Валентин, Джеймс У. (2004). О происхождении Фила. Чикаго: Издательство Чикагского университета. п. 7. ISBN 978-0-226-84548-7.
Классификации организмов в иерархических системах использовались в семнадцатом и восемнадцатом веках. Обычно организмы группировались в соответствии с их морфологическим сходством, как их воспринимали эти первые исследователи, а затем эти группы группировались в соответствии с их сходством и так далее, чтобы сформировать иерархию.
CS1 maint: ref = harv (связь) - ^ а б Валентин, Джеймс В. (18.06.2004). О происхождении Фила. ISBN 9780226845487.
- ^ Эрвин, Дуглас; Валентин, Джеймс; Яблонски, Дэвид (1997). «Недавние находки окаменелостей и новые взгляды на развитие животных позволяют по-новому взглянуть на загадку взрыва животных в раннем кембрии». Американский ученый (Март Апрель).
- ^ а б Budd, G.E .; Дженсен, С. (май 2000 г.). "Критическая переоценка летописи окаменелостей билатерального типа". Биологические обзоры. 75 (2): 253–295. Дои:10.1111 / j.1469-185X.1999.tb00046.x. PMID 10881389. S2CID 39772232.
- ^ а б Ponder, W.F .; Линдберг, Д. Р., ред. (2008). Филогения и эволюция Mollusca. Беркли: Калифорнийский университет Press. п. 481. ISBN 978-0-520-25092-5.
- ^ Porifera (сущ.) Интернет-словарь этимологии. Дата обращения 18 августа 2016.
- ^ Vacelet & Duport 2004 С. 179–190.
- ^ "Spongia Linnaeus, 1759 г.". Всемирный регистр морских видов. Получено 18 июля, 2012.
- ^ Роуленд, С. М. и Стивенс, Т. (2001). «Археоциата: история филогенетической интерпретации». Журнал палеонтологии. 75 (6): 1065–1078. Дои:10.1666 / 0022-3360 (2001) 075 <1065: AAHOPI> 2.0.CO; 2. JSTOR 1307076.
- ^ Sperling, E. A .; Pisani, D .; Петерсон, К. Дж. (1 января 2007 г.). «Пориферская парафилия и ее значение для докембрийской палеобиологии» (PDF). Геологическое общество, Лондон, Специальные публикации. 286 (1): 355–368. Bibcode:2007GSLSP.286..355S. Дои:10.1144 / SP286.25. S2CID 34175521. Архивировано из оригинал (PDF) 20 декабря 2009 г.. Получено 22 августа, 2012.
- ^ Чжан, З.-К. (2011). «Биоразнообразие животных: введение в классификацию более высокого уровня и таксономическое богатство» (PDF). Zootaxa. 3148: 7–12. Дои:10.11646 / zootaxa.3148.1.3.
- ^ "Nematostella vectensis v1.0 ". Портал генома. Калифорнийский университет. Получено 2014-01-19.
- ^ "Нематостелла". Nematostella.org. Получено 2014-01-18.
- ^ а б Генихович, Г. и У. Технау (2009). "Звездочка морского анемона Nematostella vectensis: Антозойный модельный организм для исследований в области сравнительной геномики и функциональной эволюционной биологии развития ». Протоколы Колд-Спринг-Харбор. 2009 (9): pdb.emo129. Дои:10.1101 / pdb.emo129. PMID 20147257.
- ^ «Откуда наша голова? Морская безмозглая ветреница проливает новый свет на эволюционное происхождение головы». Science Daily. 2013-02-12. Получено 2014-01-18.
- ^ Sinigaglia, C .; и другие. (2013). «Ген шесть3 / 6 формирования паттерна головы билатеральной головы контролирует развитие аборального домена у книдарии». PLOS Биология. 11 (2): e1001488. Дои:10.1371 / journal.pbio.1001488. ЧВК 3586664. PMID 23483856.
- ^ Карлескинт Дж., Ричард Тернер Р и, Джеймс Смолл Дж. (2012) Введение в морскую биологию Cengage Learning, издание 4, стр. 445. ISBN 9781133364467.
- ^ Майкл Ле Пейдж (март 2019 г.). «Животное с анусом, который приходит и уходит, может показать, как эволюционировали наши». Новый ученый.
- ^ Бавестрелло, Джорджио; Кристиан Зоммер; Мишель Сара (1992). «Двунаправленное преобразование Turritopsis nutricula (Hydrozoa)». Scientia Marina. 56 (2–3): 137–140.
- ^ Пираино, Стефано; Ф. Боеро; Б. Эшбах; В. Шмид (1996). «Изменение жизненного цикла вспять: превращение медуз в полипы и трансдифференцировка клеток Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический бюллетень. 190 (3): 302–312. Дои:10.2307/1543022. JSTOR 1543022. PMID 29227703. S2CID 3956265.
- ^ "Корнуолл - Природа - Суперзвезда Червь". BBC.
- ^ Марк Карвардин (1995) Книга рекордов Гиннеса. Издательство Guinness. п. 232.
- ^ «Стойкие паразиты». Журнал Тайм. 1957-04-08.
- ^ Харгис, Уильям Дж. (1985). «Паразитология и патология морских организмов Мирового океана». NOAA Tech. Представитель.
- ^ «Классификация паразитов животных».
- ^ Гарсия, Линн. «Классификация человеческих паразитов, переносчиков и подобных организмов» (PDF). Медицинский центр UCLA Лос-Анджелес, Калифорния.
- ^ Ходда, М. (2011). "Phylum Nematoda Cobb, 1932. В: Чжан, З.-К. (Ред.) Биоразнообразие животных: план классификации более высокого уровня и обзор таксономического богатства". Zootaxa. 3148: 63–95. Дои:10.11646 / zootaxa.3148.1.11.
- ^ Чжан, З. (2013). «Биоразнообразие животных: обновление классификации и разнообразия в 2013 году. В: Чжан, З.-К. (ред.) Биоразнообразие животных: краткое изложение классификации высокого уровня и исследования таксономического богатства (приложения 2013 г.)». Zootaxa. 3703 (1): 5–11. Дои:10.11646 / zootaxa.3703.1.3. S2CID 85252974.
- ^ Lambshead PJD (1993). «Последние достижения в исследованиях морского бентического биоразнообразия». Oceanis. 19 (6): 5–24.
- ^ Боргони Г., Гарсия-Мояно А., Литтхауэр Д., Берт В., Бестер А., ван Херден Е., Мёллер С., Эразмус М., Onstott TC (июнь 2011 г.). «Нематоды из глубоких недр Южной Африки». Природа. 474 (7349): 79–82. Bibcode:2011Натура 474 ... 79Б. Дои:10.1038 / природа09974. HDL:1854 / LU-1269676. PMID 21637257. S2CID 4399763.
- ^ Дановаро Р., Гамби С., Делль'Анно А., Коринальдези С., Фрашетти С., Ванройзель А., Винкс М., Гудей А.Дж. (январь 2008 г.). «Экспоненциальное ухудшение функционирования глубоководных экосистем, связанное с утратой бентического биоразнообразия». Curr. Биол. 18 (1): 1–8. Дои:10.1016 / j.cub.2007.11.056. PMID 18164201. S2CID 15272791. Сложить резюме – EurekAlert!.
- ^ Платт Х.М. (1994). «предисловие». В Lorenzen S, Lorenzen SA (ред.). Филогенетическая систематика вольных нематод. Лондон: Общество лучей. ISBN 978-0-903874-22-9.
- ^ "Морская лилия". Научная энциклопедия. Получено 5 сен, 2014.
- ^ "Сеть разнообразия животных - иглокожие". Музей зоологии Мичиганского университета. Получено 26 августа 2012.
- ^ Фокс, Ричард. "Asterias forbesi". Анатомия беспозвоночных онлайн. Lander University. Получено 14 июн 2014.
- ^ Холзингер, К. (2005). Краеугольные камни. Получено 10 мая 2010 г. из «Архивная копия». Архивировано из оригинал на 2010-06-30. Получено 2010-05-12.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Чепмен, А.Д. (2009). Численность живых видов в Австралии и мире, 2-е издание. Австралийское исследование биологических ресурсов, Канберра. Проверено 12 января 2010 года. ISBN 978-0-642-56860-1 (напечатано); ISBN 978-0-642-56861-8 (онлайн).
- ^ Хэнкок, Ребекка (2008). «Признание исследования моллюсков». Австралийский музей. Архивировано из оригинал на 2009-05-30. Получено 2009-03-09.
- ^ Барнс, Р.С.К., Калоу П., Олив, П.Дж.У., Голдинг Д.В., Спайсер Дж. И. (2001). Беспозвоночные, синтез (3-е изд.). Великобритания: Blackwell Science.
- ^ "Добро пожаловать в CephBase". CephBase. Получено 29 января 2016.
- ^ Wilbur, Karl M .; Clarke, M.R .; Труман, Э. Р., ред. (1985), Моллюска, 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN 0-12-728702-7
- ^ "Есть ли пресноводные головоногие моллюски?". 16 января 2013 г.
- ^ Блэк, Ричард (26 апреля 2008 г.). «Колоссальный кальмар из морозилки». Новости BBC.
- ^ Юэн Каллэуэй (2 июня 2008 г.). «Простодушный наутилус показывает вспышку памяти». Новый ученый. Получено 7 марта 2012.
- ^ Кэтрин Филлипс (15 июня 2008 г.). «Живые воспоминания об ископаемых». Журнал экспериментальной биологии. 211 (12): iii. Дои:10.1242 / jeb.020370. S2CID 84279320.
- ^ Робин Крук и Дженнифер Бэзил (2008). "Двухфазная кривая памяти в наутилусе в камере, Наутилус помпилиус L. (Cephalopoda: Nautiloidea) ". Журнал экспериментальной биологии. 211 (12): 1992–1998. Дои:10.1242 / jeb.018531. PMID 18515730. S2CID 6305526.
- ^ а б c d е ж Рупперт, Эдвард Э .; Фокс, Ричард, С .; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание. Cengage Learning. ISBN 978-81-315-0104-7.
- ^ а б Хейворд, П.Дж. (1996). Справочник морской фауны Северо-Западной Европы. Издательство Оксфордского университета. ISBN 978-0-19-854055-7.
- ^ Giribet, G .; Окусу, А, А .; Lindgren, A.R., A.R .; Huff, S.W., S.W .; Schrödl, M, M .; Нисигучи, М.К., М.К. (май 2006 г.). «Доказательства клады, состоящей из моллюсков с последовательно повторяющимися структурами: моноплакофораны родственны хитонам». Труды Национальной академии наук Соединенных Штатов Америки. 103 (20): 7723–7728. Bibcode:2006PNAS..103.7723G. Дои:10.1073 / pnas.0602578103. ЧВК 1472512. PMID 16675549.
- ^ Хили, Дж. М. (2001). «Моллюска». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. С. 120–171. ISBN 978-0-19-551368-4.
- ^ Работаем, да мы. «Узнайте больше о First Life - Первая жизнь Дэвида Аттенборо». Архивировано из оригинал 26 января 2011 г.
- ^ Д. Р. Карри; Т. М. Уорд (2009). Южно-Австралийский гигантский краб (Pseudocarcinus gigas) Рыболовство (PDF). Южно-Австралийский научно-исследовательский институт. Отчет об оценке рыболовства для PIRSA. Архивировано из оригинал (PDF) 28 марта 2012 г.. Получено 9 декабря 2013.
- ^ Гевин, В. (2005). «Функциональная геномика сгущает биологический сюжет». PLOS Биология. 3 (6): e219. Дои:10.1371 / journal.pbio.0030219. ЧВК 1149496. PMID 15941356.
- ^ Геном ланцетника (амфиоксуса) и происхождение позвоночных Ars Technica, 19 июня 2008 г.
- ^ Лемэр, П. (2011). «Эволюционный перекресток в биологии развития: оболочники». Разработка. 138 (11): 2143–2152. Дои:10.1242 / dev.048975. PMID 21558365. S2CID 40452112.
- ^ Карсон, Рэйчел (1997). Море вокруг нас. Оксфордские книги в мягкой обложке. С. 190–191. ISBN 978-0195069976.

