Панцирь брюхоногого моллюска - Gastropod shell


1 – пупок
2 – колумеллярная коса
3 – отверстие
4 – Колумелла
5 – шов
6 – оборот тела
7 – вершина

В панцирь брюхоногих моллюсков является частью тела брюхоногие моллюски или улитка, своего рода моллюск. Оболочка - это экзоскелет, который защищает от хищников, механических повреждений и обезвоживания, но также служит для прикрепления мышц и хранения кальция. Некоторые брюхоногие моллюски кажутся без панциря (слизни ), но может иметь остаток в мантии, или оболочка уменьшена так, что тело не может быть втянут внутрь (полуслаг ). У некоторых улиток также есть крышка, закрывающая отверстие в раковине, известная как отверстие, что обеспечивает дополнительную защиту. Изучение раковины моллюсков известен как конхология. Биологическое изучение брюхоногих моллюсков и других моллюсков в целом является малакология. Термины морфологии раковины варьируются в зависимости от видовой группы. Отличным источником терминологии, касающейся панцирей брюхоногих, является книга Джона Б. Берча «Как узнать восточных наземных улиток», которая сейчас находится в свободном доступе в цифровой библиотеке Hathi Trust.[1]
Слои оболочки
Раковина брюхоногих моллюсков состоит из трех основных слоев, секретируемых мантия. Известковый центральный слой, tracum, обычно состоит из карбонат кальция осажден в органическую матрицу, известную как конхиолин. Самый внешний слой - это периостракум, который устойчив к истиранию и обеспечивает большую часть окраски скорлупы. Тело улитки контактирует с самым внутренним гладким слоем, который может состоять из перламутра или перламутра, плотной горизонтально упакованной формы конхиолина, который накладывается на периостракум по мере роста улитки.
Морфология
 | | |
| Морфология типичной спиралевидной оболочки. Оболочка Zonitoides nitidus, наземная улитка, имеет правую намотку. Верхнее изображение: вид сверху, видны мутовки и вершина | Фото корпуса Zonitoides nitidus с апикальный вид, апертурный вид и базальный вид |
Морфология раковины брюхоногих моллюсков обычно довольно постоянна среди особей вида. Управляющими переменными являются:
- Скорость роста за оборот вокруг оси намотки. Высокие показатели дают формы с широким ртом, такие как морское ушко, низкие скорости дают сильно скрученные формы, такие как Turritella или некоторые из Planorbidae.
- Форма образующей кривой примерно эквивалентна форме апертуры. Он может быть круглым, например, в ракушка тюрбана, удлиненные, как в конус или иметь неправильную форму с расширением сифонального канала, как в Murex.
- Скорость перемещения образующей кривой вдоль оси наматывания, определяющая, насколько высокоразвитой становится полученная оболочка. Он может варьироваться от нуля (плоская планиспиральная оболочка) до почти диаметра отверстия.
- Неровности или «скульптуры», такие как ребра, шипы, выступы и варикозные узлы, сделанные улиткой, регулярно меняющие форму образующей кривой в процессе роста, например, у многих видов Murex.
- Онтологический рост меняется по мере того, как животное достигает взрослого возраста. Хорошие примеры - расширяющаяся губа взрослого человека. раковина и загнутый внутрь край каури.
Некоторые из этих факторов можно смоделировать математически, и существуют программы для создания чрезвычайно реалистичных изображений. Ранние работы Дэвид Рауп на аналоговом компьютере также было обнаружено множество возможных комбинаций, которые никогда не были приняты ни одним настоящим брюхоногим.
Некоторые формы раковин чаще встречаются в определенных средах, хотя есть много исключений. Омываемые волнами высокоэнергетические среды, например скалистые приливной Зона, обычно населяют улиток, раковины которых имеют широкое отверстие, относительно небольшую площадь поверхности и высокую скорость роста за оборот. Формы с высокими шпилями и высокой скульптурой становятся все более распространенными в тихой водной среде. Панцирь роющих форм, таких как оливковый и Теребра, гладкие, вытянутые и лишены сложной скульптуры, чтобы уменьшить сопротивление при движении по песку. На суше формы с высоким шпилем часто ассоциируются с вертикальными поверхностями, тогда как улитки с плоским панцирем, как правило, живут на земле.
Несколько брюхоногих моллюсков, например Vermetidae, цементируют оболочку и растут вдоль твердых поверхностей, таких как камни или другие оболочки.
Хиральность
Большинство раковин брюхоногих моллюсков спирально свернуто. Большинство (более 90%)[2] брюхоногих моллюсков правый (правые) раковины, но небольшое меньшинство видов и родов практически всегда левый (левша) и очень немногие виды (например, Amphidromus perversus[3]) показывают смесь правых и левосторонних лиц. Встречаются также аномально левосторонние формы правосторонних видов, и некоторые из них очень востребованы сборщиками раковин.[нужна цитата ]

Если удерживать свернутый в спираль панцирь брюхоногих моллюсков шпилем вверх и отверстием, более или менее обращенным к наблюдателю, в правом панцире будет отверстие с правой стороны, а в левом панцире - с левой стороны. . Эту хиральность брюхоногих моллюсков иногда упускают из виду, когда фотографии свернутых в спираль брюхоногих моллюсков «переворачиваются» неспециалистом перед использованием в публикации. Это «переворот» изображения приводит к тому, что нормальный правый брюхоногий моллюск оказывается редким или аномальным левым.
Синстральность возникала самостоятельно 19 раз среди морских брюхоногих моллюсков с начала их существования. Кайнозойский.[4] Эта леворукость чаще встречается у пресноводных и наземных легких. Но все же правосторонние виды брюхоногих моллюсков составляют 99% от общего числа.[5]
Хиральность брюхоногих моллюсков проявляется в раннем дроблении (спиральный разрез ) и ген Узловой впутан.[6] Более недавнее исследование (2013) коррелирует асимметричное скручивание оболочки с лево-правой асимметричной экспрессией гена декапентаплегии в мантии.[7]
Смешанные популяции намотки
В некоторых случаях левосторонняя и правосторонняя намотка обнаруживается в одной и той же популяции.[8] Синистральные мутанты обычно правых видов и правые мутанты обычно левосторонних видов редки, но хорошо задокументированы среди наземных улиток в целом.[8] Популяции или виды с нормально смешанной спиралевидностью встречаются гораздо реже и, насколько известно, ограничиваются, за одним исключением, несколькими родами древесных тропических улиток.[8] Помимо Амфидром, кубинский Liguus vittatus (Свейнсон), Гаитянский Liguus virgineus (Линней) (семья Орталициды ), немного Гавайский Партулина и многие гавайские Ахатинелла (семья Ахатинеллиды ), а также несколько видов островов Тихого океана Partula (семья Partulidae ), как известно, имеют смешанные правосторонние и левосторонние популяции.[8]
Возможное исключение может касаться некоторых европейских клаузилиды подсемейства Алопиины.[8] Они обязательны кальцифилы живут изолированными колониями на выходах известняков.[8] Несколько наборов видов различаются только направлением намотки, но данные неубедительны относительно того, живут ли вместе левосторонние и правосторонние раковины.[8] Соос (1928, стр. 372–385)[требуется полная цитата ] суммировал предыдущие обсуждения проблемы и пришел к выводу, что правые и левые популяции были разными видами.[8] Другие заявили, что эти популяции не были отдельными, и вопрос далеко не решен.[8] Перуанская клаузилида, Нения каллистоглипта Пилсбри (1949, стр. 216–217),[требуется полная цитата ] также был описан как вид амфидрома.[8]
Генетика обратной спирали у редкого правого мутанта другой клаузилиды, Алинда бипликата (Монтегю), был изучен Дегнером (1952).[8] Механизм такой же, как у Radix peregra (Мюллер), при этом направление намотки определяется простым Менделевский рецессивный.[8]
Стандартные способы просмотра оболочки

Апертурный вид раковины Valvata Sincera

Вид сбоку оболочки Valvata Sincera

Пупочный вид оболочки Valvata Sincera

Этот вид живого животного со спины Calliostoma bairdii также показан апикальный вид его раковины




На фотографиях или иллюстрациях панцирь брюхоногого моллюска можно показать ориентированным рядом стандартных способов:
- апертурный вид: это наиболее распространенный угол обзора. Оболочка показана целиком, с отверстием, обращенным к зрителю, и вершиной вверху. Если при таком взгляде отверстие находится с правой стороны, то намотка оболочки является «правосторонней» или правосторонней; если при таком взгляде отверстие находится с левой стороны, то оболочка имеет "левостороннюю" или левостороннюю намотку.
- вид сверху (или же вид со спины): оболочка показана апертурой на 180 ° от наблюдателя и вершиной вверху.
- пупочный вид (или же базальный вид): оболочка показана прямо снизу. В большинстве случаев, когда есть пупок, он хорошо виден.
- апикальный вид: оболочка показана смотрящей вниз прямо на вершину.
Описание

Раковина начинается с личиночной раковины, (обычно) минутной эмбриональный завитки, известные как протоконх, который часто довольно сильно отличается от остальной оболочки и не имеет линий роста. Из протоконха, который образует вершину шпиль, катушки или завитки оболочки постепенно увеличиваются в размерах. Обычно мутовки круглые или эллиптические в сечении. Шпиль может быть высоким или низким, широким или тонким, в зависимости от того, как расположены витки оболочки, и соответственно изменяется апикальный угол оболочки. Мутовки иногда неплотно прилегают друг к другу (как в Скалярный эпитоний ). Они также могут перекрывать более ранние обороты, так что более ранние обороты могут быть в значительной степени или полностью покрыты более поздними. Когда возникает изгиб, пространство между ним и шов над ним находится область, известная как «плечо» раковины. Угол плеча может быть гладким или килевым, а иногда может иметь узлы или шипы.
Самая простая форма скульптура Панцирь брюхоногих моллюсков состоит из продольных и / или поперечных гребней. Первичные спирали могут появляться в регулярной последовательности по обе стороны от первой первичной, что обычно становится углом плеча, если возникает изгиб. Вторичные спирали могут возникать путем вставки между первичными и обычно отсутствуют в молодой оболочке, за исключением некоторых сильно ускоренных типов. У более специализированных видов между предшествующими группами вставлены третичные спирали. Ребра представляют собой регулярные поперечные складки оболочки, которые обычно проходят от шва к шовному материалу. Обычно они расположены равномерно и пересекаются спиралями. У специализированных типов, когда формируется плечевой угол, они концентрируются в виде узлов под этим углом, исчезают с плеча вверху и тела внизу. Шипы могут заменить узлы на более поздних стадиях. Они образуют выемки на краю скорлупы и впоследствии отбрасываются, часто оставаясь открытыми спереди. Неровные шипы также могут возникать на различных участках поверхности раковины (см. Platyceras ).
Когда в период покоя на краю или внешней кромке раковины образуется ряд шипов, эта особенность иногда остается позади в виде варикоз как в (Murex ) и многие из Ранеллиды. Варикозное расширение вен также может быть образовано простым расширением наружной губы и последующим возобновлением роста от основания расширения.
В отверстие или перистом оболочки может быть простым или различным образом модифицированным. Обычно распознаются внешняя и внутренняя (колумеллярная) губа. Они могут быть непрерывными друг с другом или могут быть разделены передней выемкой. Это в некоторых типах (Фузинус и т. д.) вытягивается в передний сифональный канал, большей или меньшей длины.
В некоторых таксонах присутствует верхняя или задняя выемка, и это может привести к образованию гребня или выступа рядом с швом (Clavilithes ). Наружная (боковая) выемка или выемка, иногда продолжающаяся в разрез, встречается у определенных типов (Pleurotomidae, Pleurotomaridae, Bellerophontidae и т. Д.), И постепенное закрытие этой щели может привести к появлению четко выраженной полосы разреза. В некоторых случаях прорезь оставляют в виде отверстия (Fissurellidae ), или периодическим обновлением в виде последовательности дырок (Haliotis ). На внешнюю выемку часто указывает только отраженный ход линий роста на раковине.
На внутренней стороне внешней губы различные гребни или складки, называемые лира иногда встречаются, а иногда они могут быть крепкими и зубчатыми (Неринея ). Подобные гребни или колумеллярные складки или складки чаще встречаются на внутренней губе, рядом с колумеллой или центральным витком спирали. Они могут быть наклонными или перпендикулярными оси наматывания (горизонтальными), немногочисленными или многочисленными, легко заметными или далеко внутри оболочки, чтобы быть невидимыми, за исключением сломанных оболочек. Когда ось навивки полая (перфорированный шпиль), отверстие в основании образует пупок. Пупок сильно различается по размеру и может быть полностью или частично покрыт расширением или мозолью внутренней губы (Натика ).
Много Недавний панцири, когда животное живое или панцирь только что пустой, имеют верхний слой оболочки из рогового, гладкого или волосатого эпидермиса или периостракум, белковый слой, который иногда бывает достаточно толстым, чтобы скрыть цветные отметины на поверхности скорлупы. Периостракум, как и окраска, редко сохраняется в ископаемых раковинах.
Апертурный конец панциря брюхоногих моллюсков - передний конец, ближайший к голове животного; вершина шпиля часто находится на заднем конце или, по крайней мере, на дорсальной стороне. Большинство авторов изображают раковины вершиной шпиля вверх. В жизни, когда мягкие части этих улиток втянуты, в некоторых группах отверстие раковины закрывается роговой или известковой крышка, дверная структура, которая выделяется и прикрепляется к верхней поверхности задней части стопы. Жаберная крышка имеет очень изменчивую форму у разных групп улиток, у которых она есть.[9]
Части оболочки

Терминология, используемая для описания панцирей брюхоногих моллюсков, включает:
- Диафрагма: открытие оболочки
- Губа: перистом: край апертуры
- Апекс: несколько самых маленьких оборотов раковины
- Оборот тела (или последний оборот): самый большой оборот, в котором находится основная часть висцеральной массы моллюска.
- Колумелла: «столбик» на оси вращения оболочки
- Operculum: "люк" оболочки
- Теменная мозоль: гребень на внутренней губе отверстия у некоторых брюхоногих моллюсков
- Периостракум: тонкий слой органической «кожицы», которая образует внешний слой панциря многих видов.
- Перистом: часть оболочки, которая находится прямо вокруг отверстия, также известная как губа
- Коса: складки на колумелле.
- Протоконх: ядерный или эмбриональный оборот; раковина личинки, часто остается на месте даже на взрослой раковине
- Скульптура: орнамент на внешней поверхности раковины
- Лира: выпуклые линии или выступы на поверхности скорлупы
- Сифональный канал: расширение апертуры у некоторых брюхоногих моллюсков
- Шпиль: часть раковины над оборотом тела.
- Шовный: Переход между завитки большинства брюхоногих моллюсков
- Телеоконх: вся оболочка без протоконха; постъядерные обороты.
- Пупок: в раковинах, у которых мутовки раздвигаются по мере роста, на нижней стороне раковины имеется глубокое углубление, доходящее до шпиля; это пупок
- Варикоз: на раковинах некоторых моллюсков выступающие и утолщенные вертикальные ребра обозначают окончание периода быстрого роста; это варикоз
- Whorl: каждое из полных оборотов спирали оболочки
Форма корпуса
Общая форма раковины варьируется. Например, по соотношению высоты и ширины можно выделить три группы:[10]
- продолговатый - высота намного больше ширины
- шаровидная или коническая раковина - высота и ширина раковины примерно одинаковы
- депрессивный - ширина намного больше высоты

продолговатая оболочка Булгарика зубчатая

шаровидная оболочка Sphincterochila Candidissima

депрессивная оболочка Escargot de Quimper


Ниже приведены основные изменения формы раковины брюхоногих моллюсков.[11]
- Регулярно по спирали:
- Буллоид: пузырчатый Булла
- Целоконоид представляет собой слегка вогнутую коническую оболочку, в которой угол приращения постоянно увеличивается во время роста (см .: Каллиостома )
- Конусообразная, обратноконическая. Конус
- Контабулированный, короткий, с лопаточными мутовками
- Вогнутый: отверстие, равное длине раковины, почти или полностью скрывает шпиль. Cypraea
- Цилиндрические, куколочные. Lioplax, Куколка
- Циртоконоид: приближается к конической форме, но с выпуклыми сторонами (см .: Гиббула )
- Вдавленный, двояковыпуклый. Ethalia carneolata
- Дискоидальный. Элахорбис
- В форме уха. Haliotis
- Удлиненный, шиловидный, приподнятый. Теребра
- Мало-мутовчатый. Helix pomatia.
- Веретеновидная, веретеновидная. Фузинус
- Гиббус. Мутовки вздуты за пределы нормального контура увеличения (обычно со стороны апертуры). Streptaxis.
- Шаровидный. Натика
- Многомутовый. Миллереликс перегрина.
- Короткие, щечкообразные. Buccinum
- Трохиформные, пирамидальные, конические с плоским основанием. Trochus
- Турбинированный: конический, с закругленным основанием. Турбо
- Башеночный, туррикулярный, вавилонный; продолговатый панцирь с загнутыми углами или выступами в верхней части мутовками. Turritella
- Скалярно, мутовки не заходят внутрь. Скалярный эпитоний
- Неправильно спиралевидный, эволютный. Силикария, Верметус
- Трубчатый.
- В форме щита. Умбракулюм
- В форме лодочки, в форме тапочки. Crepidula
- Коническая или блюдцеобразная форма. Надколенник
- Биконический: имеет форму двух конических форм, которые соприкасаются со своими основаниями и сужаются на обоих концах: Fasciolaria tulipa
- Грушевидная: сочетание двух форм: яйцевидно-конической и конической. Фикус
Детальное различение формы может быть:[12][13]

форма крышки

форма уха

неритиформный

планиспиральный

вдавленный трохиформный или вальватиформный

трохиформ

яйцевидно-конический

конический

удлиненно-конической или турриформной или петлевидной формы

верхняя форма

форма веретена - морская улитка Сиринкс аруанский имеет самый большой панцирь среди всех ныне живущих брюхоногих моллюсков.

форма клуба - Расческа Venus Murex

форма яйца

неправильная форма












Размеры
Наиболее часто используемые размеры панциря брюхоногих моллюсков: высота оболочки, ширина оболочки, высоту проема и ширину проема. Также часто используется количество оборотов.
В этом контексте высота (или длина) оболочки - это ее максимальное измерение вдоль центральной оси. Ширина (или ширина, или диаметр) - это максимальное измерение оболочки под прямым углом к центральной оси. Оба термина относятся только к описанию панциря, а не к ориентации панциря на живом животном.
Самая большая высота любой раковины у морских видов улиток. Сиринкс аруанский, который может достигать 91 см.[14]
В центральная ось представляет собой воображаемую ось по длине оболочки, вокруг которой в свернутой оболочке спиралью закручиваются обороты. Центральная ось проходит через колумеллу, центральную опору раковины.
Эволюционные изменения
Среди предлагаемых ролей, связанных с изменчивостью снарядов во время эволюция включают механическую стабильность,[15] защита от хищников[16] и климатический отбор.[17][18]
Раковины некоторых брюхоногих моллюсков уменьшились или частично уменьшились во время их эволюция. Это сокращение можно увидеть во всех слизни, у полуслизней и у различных других морских и неморских брюхоногих моллюсков. Иногда сокращение панциря связывают с хищным способом кормления.
Некоторые таксоны в процессе эволюции утратили спиральность своей раковины.[19] В соответствии с Закон Долло, восстановить свертывание оболочки после ее утраты невозможно. Несмотря на это, родов в семье немного. Calyptraeidae которые изменили время их развития (гетерохрония ) и вернул (реэволюция ) спиральная оболочка из предыдущего состояния размотанной бездомный -подобная оболочка.[19]
Тафономический подразумеваемое
В достаточно больших количествах панцири брюхоногих моллюсков могут оказывать достаточное влияние на условия окружающей среды, чтобы повлиять на способность органических остатков в окружающей среде окаменевать.[20] Например, в Формирование парка динозавров Ископаемая яичная скорлупа гадрозавра встречается редко.[20] Это потому, что поломка дубильные вещества из местных хвойный из-за растительности древние воды стали кислыми.[20] Фрагменты яичной скорлупы присутствуют только в двух микрофоссилий места, где преобладают сохранившиеся раковины беспозвоночных, в том числе брюхоногих.[20] Это было медленное растворение этих снарядов, высвобождающее карбонат кальция в воду, которая подняла воду pH достаточно высокий, чтобы предотвратить растворение фрагментов яичной скорлупы до того, как они смогут окаменеть.[20]
Разнообразие форм

Turritella communis, многовитковая раковина башенной улитки

Рентгеновское изображение Turritella

Раковина морской улитки каури - Cypraea nebrites

Скалярный эпитоний спиральная оболочка





Рентгеновское изображение Cypraea

рентгеновский снимок изображение оболочки Tonna Galea
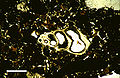
Тонкий срез в плоскополяризованном свете микроскопической раковины брюхоногих моллюсков, от Голоцен лагунные отложения Райс-Бэй, Остров Сан-Сальвадор, Багамы. Масштабная линейка 500 мкм.





Рекомендации
Эта статья включает в себя текст из ссылок, являющийся общественным достоянием,[1][8][9] и текст CC-BY-2.0 из ссылки.[18]
- ^ а б Джон. Б. Берч (1962). «Как узнать восточных наземных улиток; ключ к изображению для определения наземных улиток Соединенных Штатов, встречающихся к востоку от водораздела Скалистых гор» WM. C. Brown Company Publishers, Дубьюк, Айова. 214 страниц.
- ^ Шильтхуйзен М. и Дэвисон А. (2005). «Запутанная эволюция хиральности улитки». Naturwissenschaften 92(11): 504–515. PMID 16217668. Дои:10.1007 / s00114-005-0045-2.
- ^ «Amphidromus perversus (Linnaeus, 1758)». jaxshells.org.
- ^ Гират Вермей (2002). "География эволюционных возможностей: гипотеза и два случая брюхоногих моллюсков". Интегративная и сравнительная биология 42(5): 9359–40. Дои:10.1093 / icb / 42.5.935.
- ^ Асами Т. (1993) "Генетические вариации спиральной хиральности улиток". Forma, 8, 263–276
- ^ Майерс П. З. (13 апреля, 2009 г.) "Улитки есть узловые!". Большой палец панды, по состоянию на 3 мая 2009 г.
- ^ Кейсуке Симидзу, Минору Иидзима, Давин Его Превосходительство Сетиамарга Исао Сарасина, Тетсухиро Кудо, Такахиро Асами, Эди Гиттенбергер, Казуёси Эндо; "Лево-правое асимметричное выражение dpp в мантии брюхоногих моллюсков коррелирует с асимметричным свертыванием раковины ". EvoDevo 2013, 4:15. Дои:10.1186/2041-9139-4-15.
- ^ а б c d е ж грамм час я j k л м
 Лэйдлоу Ф. Ф. & Солем А. (1961). "Род наземных улиток Амфидром: синоптический каталог ». Филдианская зоология 41(4): 505–720.
Лэйдлоу Ф. Ф. & Солем А. (1961). "Род наземных улиток Амфидром: синоптический каталог ». Филдианская зоология 41(4): 505–720. - ^ а б Грабау А.В. & Шимер Х. В. (1909) Индекс окаменелостей Северной Америки Беспозвоночные. Том I.. A. G. Seiler & Company, Нью-Йорк. страницы стр. 582 –584.
- ^ Фолкнер Г., Обрдлик П., Кастелла Э. и Спейт М. К. Д. (2001). Панцирные гастроподы Западной Европы. München: Friedrich-Held-Gesellschaft, 267 стр.
- ^ Джордж Вашингтон Трайон, Структурная и систематическая конхология, 1882, стр. 43–44
- ^ Хершлер Р. и Пондер В. Ф. (1998). «Обзор морфологических признаков гидробиоидных улиток». Вклад Смитсоновского института в зоологию 600: 1–55. HDL:10088/5530.
- ^ Танец П.С. Снаряды.[страница нужна ]
- ^ Уэллс Ф. Э., Уокер Д. И. и Джонс Д. С. (ред.) (2003) "Пища великанов - полевые наблюдения за диетой Сиринкс аруанский (Linnaeus, 1758) (Turbinellidae) самый крупный из ныне живущих брюхоногих моллюсков. ". Морская флора и фауна Дампира, Западная Австралия. Музей Западной Австралии, Перт.
- ^ Бриттон Дж. К. (1995) "Взаимосвязь между положением на берегу и орнаментом ракушек у двух зависящих от размера морфотипов. Littorina striata, с оценкой потерь воды за счет испарения у этих морфотипов и у Melarhaphe neritoides". Гидробиология 309: 129–142. Дои:10.1007 / BF00014480.
- ^ Уилсон А. Б., Глаубрехт М. и Мейер А. (март 2004 г.) «Древние озера как эволюционные резервуары: свидетельства талассоидных брюхоногих моллюсков озера Танганьика». Труды Лондонского королевского общества, серия B - Биологические науки 271: 529–536. Дои:10.1098 / rspb.2003.2624.
- ^ Гудфренд Г. А. (1986) "Изменение формы и размера раковины наземных улиток и их причины - Обзор". Систематическая зоология 35: 204–223.
- ^ а б Пфеннингер М., Грабакова М., Стейнке Д. и Депраз А. (4 ноября 2005 г.) «Почему у улиток есть волосы? Байесовский вывод эволюции характера». BMC Эволюционная биология 5: 59. Дои:10.1186/1471-2148-5-59
- ^ а б Коллин Р. и Чиприани Р. (22 декабря 2003 г.) "Закон Долло и повторная эволюция свертывания оболочки". Труды Королевского общества B 270(1533): 2551–2555. Дои:10.1098 / rspb.2003.2517 PMID 14728776.
- ^ а б c d е Танке, Д.Х., Бретт-Сурман, М.К. 2001. Свидетельства появления птенцов и гадрозавров размером с птенцов (Reptilia: Ornithischia) из провинциального парка динозавров (формация парка динозавров: кампания), Альберта, Канада. С. 206–218. В: Мезозойская жизнь позвоночных - новые исследования, вдохновленные палеонтологией Филиппа Дж. Карри. Под редакцией Д.Танке и К. Карпентер. Издательство Индианского университета: Блумингтон. xviii + 577 с.
дальнейшее чтение
| Часть серии по |
| Ракушки |
|---|
 |
| Раковины моллюсков |
| О раковинах моллюсков |
| Другие ракушки |
- О хиральности
- ван Батенбург1 Ф. Х. Д. и Гиттенбергер Э. (1996). «Легкость фиксации смены намотки: компьютерные эксперименты по хиральности улиток». Наследственность 76: 278–286. Дои:10.1038 / хди.1996.41.
- Ванделт Дж. И Надь Л. М. (24 августа 2004 г.) «Лево-правая асимметрия: более чем один способ свернуть оболочку». Текущая биология 14(16): R654 – R656. Дои:10.1016 / j.cub.2004.08.010
внешняя ссылка
![]() СМИ, связанные с Панцири брюхоногих моллюсков в Wikimedia Commons
СМИ, связанные с Панцири брюхоногих моллюсков в Wikimedia Commons
- Брюхоногие моллюски Дж. Х. Лила - Информация о некоторых брюхоногих моллюсках тропической Западной Атлантики, особенно Карибского моря, имеющая отношение к рыболовству в этом регионе.
- Радиоуглеродное датирование раковин брюхоногих моллюсков
- Наир К. К. и Муте П. Т. (18 ноября 1961 г.) «Влияние рибонуклеазы на регенерацию оболочки в Ариофанта sp. ". Природа 192: 674–675. Дои:10.1038 / 192674b0.
- (на испанском) Антонио Руис Руис, Анхель Каркаба Посо, Ана И. Поррас Кревиллен и Хосе Р. Арребола Бургос Caracoles Terrestres de Andalúcia. Guía y manual de Identificación. 303 стр., ISBN 84-935194-2-1. (с сайта )