Сперматогенез - Spermatogenesis
| Сперматогенез | |
|---|---|
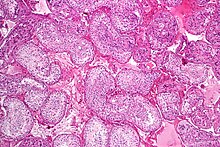
 Семенной канальец со созревающей спермой. H&E пятно. | |
 Зрелый человек Сперматозоид | |
| Идентификаторы | |
| MeSH | D013091 |
| Анатомическая терминология | |

Сперматогенез это процесс, посредством которого гаплоидный сперматозоиды развиваться из стволовые клетки в семенные канальцы из яички. Этот процесс начинается с митотическое деление из стволовые клетки расположены близко к базальной мембране канальцев.[1] Эти клетки называются сперматогониальные стволовые клетки. Их митотическое деление дает два типа клеток. Клетки типа A пополняют запас стволовых клеток, а клетки типа B дифференцируются в первичные сперматоциты. Первичный сперматоцит делится мейотически (Мейоз I) на два вторичных сперматоцита; каждый вторичный сперматоцит делится на два равных гаплоида сперматиды пользователя Meiosis II. Сперматиды превращаются в сперматозоиды (сперматозоиды) в процессе спермиогенез. Они развиваются в зрелые сперматозоиды, также известные как сперматозоиды.[2] Таким образом, первичный сперматоцит дает начало двум клеткам, вторичным сперматоцитам, а два вторичных сперматоцита путем их подразделения производят четыре сперматозоида и четыре гаплоидных клетки.[3]
Сперматозоиды - зрелый самец гаметы у многих организмов, размножающихся половым путем. Таким образом, сперматогенез - это мужской вариант гаметогенез, из которых женский эквивалент оогенез. В млекопитающие это происходит в семенные канальцы мужчины яички поэтапно. Сперматогенез во многом зависит от оптимальных условий для правильного протекания процесса и важен для половое размножение. Метилирование ДНК и гистоновая модификация были вовлечены в регулирование этого процесса.[4] Это начинается в половое созревание и обычно продолжается непрерывно до самой смерти, хотя с возрастом можно заметить небольшое уменьшение количества продуцируемой спермы (см. Мужское бесплодие ).
Сперматогенез начинается в нижней части семенных трубок, и постепенно клетки углубляются в трубочки и перемещаются по ним, пока зрелые сперматозоиды не достигнут просвета, где откладываются зрелые сперматозоиды. Деление происходит асинхронно; если трубку разрезать поперек, можно наблюдать различные состояния созревания. Группа клеток с разными состояниями созревания, которые генерируются одновременно, называется сперматогенной волной.[5]
Цель
Сперматогенез производит зрелые мужские гаметы, обычно называемые сперма но более конкретно известный как сперматозоиды, которые способны оплодотворять женскую гамету, ооцит, в течение зачатие для производства одноклеточного человека, известного как зигота. Это краеугольный камень половое размножение и включает две гаметы, которые вносят половину нормального набора хромосомы (гаплоидный ) приводить к хромосомно нормальному (диплоид ) зигота.
Чтобы сохранить количество хромосом в потомстве, которое различается между разновидность - одна из каждой гаметы должна иметь половину обычного количества хромосом, присутствующих в других клетках тела. В противном случае у потомства будет вдвое больше нормального количества хромосом, что может привести к серьезным отклонениям от нормы. У людей хромосомные аномалии, возникающие в результате неправильного сперматогенеза, приводят к врожденным дефектам и аномальным врожденным дефектам (Синдром Дауна, Синдром Клайнфельтера ) и в большинстве случаев самопроизвольный аборт развивающегося плода.
Расположение у людей
Сперматогенез происходит в нескольких структурах мужская репродуктивная система. Начальные стадии происходят в семенниках и переходят к придаток яичка где развивающиеся гаметы созревают и хранятся до эякуляция. В семенные канальцы яичек являются отправной точкой для процесса, где сперматогониальные стволовые клетки прилегающие к внутренней стенке канальцев разделяют в центростремительном направлении, начиная со стенок и переходя во внутреннюю часть, или просвет- производить незрелую сперму.[2] Созревание происходит в придатке яичка. Местоположение [семенники / мошонка] особенно важно, так как процесс сперматогенеза требует более низкой температуры для производства жизнеспособных сперматозоидов, в частности, на 1-8 ° C ниже нормальной температуры тела 37 ° C (98,6 ° F).[6] Клинически небольшие колебания температуры, например, от спортивного ремня поддержки, не вызывают ухудшения жизнеспособности или количества сперматозоидов.[7]
Продолжительность
Для человека весь процесс сперматогенеза по разным оценкам занимает 74 дня.[8][9] (по данным биопсий, меченных тритием) и примерно 120 дней[10] (по измерениям ДНК-часов). Включая транспортировку по системе воздуховодов, это занимает 3 месяца. Семенники производят от 200 до 300 миллионов сперматозоидов ежедневно.[11] Однако только около половины или 100 миллионов из них становятся жизнеспособными сперматозоидами.[12]
Этапы
Весь процесс сперматогенеза можно разбить на несколько отдельных этапов, каждая из которых соответствует определенному типу клеток человека. В следующей таблице плоидность, число копий и количество хромосом / хроматид приведены для одной клетки, как правило, до синтеза и деления ДНК (в G1, если применимо). Первичный сперматоцит задерживается после синтеза ДНК и перед делением.
| Тип ячейки | плоидность / хромосомы у человека | Номер копии ДНК /хроматиды в человеческом | Процесс введен ячейкой |
| сперматогоний (типы Ad, Ap и B) | диплоид (2N) / 46 | 2C / 46 | сперматоцитогенез (митоз ) |
| начальный сперматоцит | диплоид (2N) / 46 | 4C / 2x46 | сперматидогенез (мейоз I) |
| два вторичных сперматоциты | гаплоид (N) / 23 | 2C / 2x23 | сперматидогенез (мейоз II) |
| четыре сперматиды | гаплоид (N) / 23 | С / 23 | спермиогенез |
| четыре функциональных сперматозоиды | гаплоид (N) / 23 | С / 23 | семяизвержение |
Сперматоцитогенез


Сперматоцитогенез - мужская форма гаметоцитогенез и приводит к образованию сперматоциты обладающий половиной нормального набора генетического материала. В сперматоцитогенезе диплоид сперматогоний, который находится в базальном отделе семенных канальцев, митотически делится, производя две диплоидные промежуточные клетки, называемые первичные сперматоциты. Затем каждый первичный сперматоцит перемещается в адлюминальный отсек семенных канальцев и дублирует его ДНК и впоследствии подвергается мейоз I произвести два гаплоида вторичные сперматоциты, который позже еще раз разделится на гаплоидный сперматиды. Это деление подразумевает источники генетической изменчивости, такие как случайное включение одной из родительских хромосом и хромосомный кроссовер что увеличивает генетическую изменчивость гамет. В Повреждение ДНК Ответная реакция (DDR) играет важную роль в сперматогенезе. Протеин FMRP связывается с мейотические хромосомы и регулирует динамику аппарата DDR во время сперматогенеза.[13] FMRP кажется необходимым для восстановление повреждений ДНК.
Каждое деление клетки от сперматогония до сперматиды неполное; клетки остаются связанными друг с другом мостиками цитоплазмы, что обеспечивает синхронное развитие. Не все сперматогонии делятся с образованием сперматоцитов; в противном случае запасы сперматогоний закончились бы. Вместо, сперматогониальные стволовые клетки делятся митотически, чтобы производить копии самих себя, обеспечивая постоянный приток сперматогониев для подпитки сперматогенеза.[14]
Сперматидогенез
Сперматидогенез - это создание сперматиды из вторичных сперматоцитов. Вторичные сперматоциты, полученные ранее, быстро входят в мейоз II и делятся с образованием гаплоидных сперматид. Краткость этой стадии означает, что вторичные сперматоциты редко встречаются в гистологический исследования.
Спермиогенез
Во время спермиогенеза сперматиды начинают образовывать хвост, разрастаясь. микротрубочки на одной из центриолей, переходящей в базальное тело. Эти микротрубочки образуют аксонема. Позже центриоль видоизменяется в процессе уменьшение центросомы.[15] Передняя часть хвоста (называемая средней частью) утолщается, потому что митохондрии расположены вокруг аксонемы, чтобы обеспечить снабжение энергией. Сперматида ДНК также проходит упаковку, становясь сильно уплотняемой. ДНК сначала упаковывается со специфическими основными ядерными белками, которые впоследствии заменяются на протамины во время удлинения сперматид. В результате плотно упакованный хроматин транскрипционно неактивен. В аппарат Гольджи окружает теперь конденсированное ядро, становясь акросома.
Затем созревание происходит под влиянием тестостерона, который удаляет оставшиеся ненужные цитоплазма и органеллы. Избыточная цитоплазма, известная как остаточные тела, является фагоцитированный окружая клетки Сертоли в яички. Полученные сперматозоиды теперь созрели, но не имеют подвижности. Зрелые сперматозоиды освобождаются от защитного Клетки Сертоли в просвет семенной канальец в процессе, называемом семяизвержение.
Неподвижные сперматозоиды транспортируются в придаток яичка в тестикулярная жидкость секретируется клетками Сертоли с помощью перистальтическое сокращение. Находясь в придатке яичка, сперматозоиды приобретают подвижность и становятся способными к оплодотворению. Однако транспорт зрелых сперматозоидов через оставшуюся часть мужская репродуктивная система достигается за счет сокращения мышц, а не недавно приобретенной подвижности сперматозоидов.
Роль клеток Сертоли
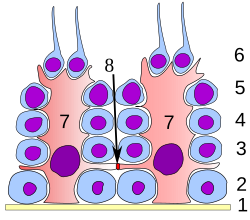
На всех стадиях дифференцировки сперматогенные клетки находятся в тесном контакте с клетками Сертоли, которые, как считается, обеспечивают структурную и метаболическую поддержку развивающихся сперматозоидов. Одиночная клетка Сертоли простирается от базальной мембраны до просвета семенного канальца, хотя цитоплазматические отростки трудно различить на уровне световой микроскопии.
Клетки Сертоли выполняют ряд функций во время сперматогенеза, они поддерживают развивающиеся гаметы следующим образом:
- Поддерживать среду, необходимую для развития и созревания, с помощью гемато-яичковый барьер
- Секретные вещества, инициирующие мейоз
- Секрет поддерживающей тестикулярной жидкости
- Секрет андрогенсвязывающий белок (ABP), который концентрирует тестостерон в непосредственной близости от развивающихся гамет
- Тестостерон необходим в очень больших количествах для поддержания репродуктивного тракта, а АД обеспечивает гораздо более высокий уровень фертильности.
- Секретные гормоны, влияющие на контроль гипофиза над сперматогенезом, особенно полипептидный гормон, ингибин
- Остаточная цитоплазма фагоцитоза, оставшаяся после спермиогенеза
- Секреция антимюллеров гормон вызывает ухудшение мюллерова протока[16]
- Защищайте сперматиды от иммунной системы мужчины через гемато-яичковый барьер
- Внести свой вклад в сперматогониальные стволовые клетки ниша
В молекулы межклеточной адгезии ICAM-1 и растворимый ICAM-1 иметь антагонистические эффекты на узкие стыки формируя гемато-яичковый барьер.[17] ICAM-2 молекулы регулируют адгезию сперматид на апикальной стороне барьера (по направлению к просвет ).[17]
Влияющие факторы
Процесс сперматогенеза очень чувствителен к колебаниям окружающей среды, особенно гормоны и температура. Тестостерон требуется в больших локальных концентрациях для поддержания процесса, что достигается за счет связывания тестостерона с помощью андрогенсвязывающий белок присутствует в семенных канальцах. Тестостерон вырабатывается интерстициальными клетками, также известными как Клетки Лейдига, которые расположены рядом с семенными канальцами.
Семенной эпителий чувствителен к повышенной температуре у людей и некоторых других видов, и на него неблагоприятно влияют температуры, превышающие нормальную температуру тела. Следовательно, яички расположены вне тела в кожаном мешочке, называемом мошонка. Оптимальная температура поддерживается на уровне 2 ° C (человек ) (8 ° С мышь ) ниже температуры тела. Это достигается за счет регулирования кровотока.[18] и позиционирование к теплу тела и от него кремастерная мышца и Дартош гладкая мускулатура мошонки.
Одним из важных механизмов является теплообмен между потоками артериальной и венозной крови в яичках. Специализированные анатомические структуры состоят из двух зон спирали вдоль внутренней семенной артерии. Такое анатомическое расположение продлевает время контакта и теплообмена между потоками тестикулярной артериальной и венозной крови и может частично объяснять температурный градиент между аортальной и тестикулярной артериальной кровью, наблюдаемый у собак и баранов. Более того, снижение пульсового давления, происходящее в проксимальной трети длины спирали внутренней семенной артерии.[требуется разъяснение ][19][20] Кроме того, снижается активность сперматогенной рекомбиназы, что считается важным фактором дегенерации яичек.[требуется разъяснение ][21]
Диетический дефицит (например, витаминов B, E и A), анаболические стероиды, металлы (кадмий и свинец), рентгеновское излучение, диоксин, алкоголь и инфекционные заболевания также отрицательно влияют на скорость сперматогенеза.[нужна цитата ] Кроме того, мужская зародышевая линия чувствительна к повреждению ДНК, вызванному окислительным стрессом, и это повреждение, вероятно, оказывает значительное влияние на оплодотворение и беременность.[22] Воздействие пестицидов также влияет на сперматогенез.[23]
Гормональный контроль
Гормональный контроль сперматогенеза зависит от вида. У людей механизм до конца не изучен; однако известно, что начало сперматогенеза происходит в период полового созревания из-за взаимодействия гипоталамус, гипофиз и Клетки Лейдига. Если гипофиз удален, сперматогенез может быть инициирован фолликулостимулирующего гормона (ФСГ) и тестостерон.[24] В отличие от ФСГ, лютеинизирующий гормон (ЛГ), по-видимому, играет небольшую роль в сперматогенезе, кроме индукции выработки гонадного тестостерона.[24][25]
ФСГ стимулирует как производство андрогенсвязывающий белок (ABP) пользователя Клетки Сертоли, и формирование гемато-яичковый барьер. ABP необходимо для концентрации тестостерона на достаточно высоком уровне, чтобы инициировать и поддерживать сперматогенез. Уровни интратестикулярного тестостерона в 20–100 или 50–200 раз превышают его концентрацию в крови, хотя среди здоровых мужчин наблюдаются колебания в 5–10 раз.[26][27] ФСГ может инициировать секвестрацию тестостерона в яичках, но после его выработки для поддержания сперматогенеза требуется только тестостерон.[24] Однако повышение уровня ФСГ увеличивает производство сперматозоидов, предотвращая апоптоз из сперматогония типа А. Гормон ингибин действует на снижение уровня ФСГ. Исследования на моделях грызунов показывают, что гонадотропины (как ЛГ, так и ФСГ) поддерживают процесс сперматогенеза, подавляя проапоптотические сигналы, и, следовательно, способствуют выживанию сперматогенных клеток.[28]
Сами клетки Сертоли опосредуют части сперматогенеза посредством выработки гормонов. Они способны вырабатывать гормоны эстрадиол и ингибин. Клетки Лейдига также способны производить эстрадиол в дополнение к своему основному продукту - тестостерону. Было обнаружено, что эстроген необходим для сперматогенеза у животных.[29][30] Однако мужчина с синдром нечувствительности к эстрогенам (дефектный ERα ) была обнаружена выработка спермы с нормальным количество сперматозоидов, хоть и ненормально низкий жизнеспособность спермы; был ли он бесплоден, неясно.[31] Слишком высокий уровень эстрогена может быть вредным для сперматогенеза из-за подавления секреции гонадотропина и увеличения выработки тестостерона в организме.[32] Пролактин также, по-видимому, важен для сперматогенеза.[25]
Смотрите также
Рекомендации
- ^ де Крецер, Д. М .; Loveland, K. L .; Meinhardt, A .; Simorangkir, D .; Рэфорд, Н. (1998-04-01). «Сперматогенез». Репродукция человека. 13 (Suppl_1): 1–8. Дои:10.1093 / humrep / 13.suppl_1.1. ISSN 0268-1161. PMID 9663765.
- ^ а б Шарма С., Ханукоглу А., Ханукоглу И. (2018). «Локализация эпителиального натриевого канала (ENaC) и CFTR в зародышевом эпителии яичка, клетках Сертоли и сперматозоидах». Журнал молекулярной гистологии. 49 (2): 195–208. Дои:10.1007 / s10735-018-9759-2. PMID 29453757. S2CID 3761720.
- ^ "Сперматозоиды в анатомии Грея". Получено 2010-10-07.
- ^ Песня, Нин; Лю, Цзе; Ан, Шукай; Нишино, Томоя; Хисикава, Ёситака; Кодзи, Такехико (2011). «Иммуногистохимический анализ модификаций гистона H3 в зародышевых клетках во время сперматогенеза мышей». Acta Histochemica et Cytochemica. 44 (4): 183–90. Дои:10.1267 / ahc.11027. ЧВК 3168764. PMID 21927517.
- ^ Шульце, В. (24 апреля 2009 г.). «Свидетельства волны сперматогенеза в семенниках человека». Андрология. 14 (2): 200–207. Дои:10.1111 / j.1439-0272.1982.tb03124.x. PMID 7103139.
- ^ «мошонка». Encyclopdia Britannica. Энциклопедия Britannica Online. Encyclopdia Britannica Inc., 2015. Интернет. 14 января 2015 <http://www.britannica.com/EBchecked/topic/530078/scrotum >.
- ^ Ван С., Макдональд В., Люнг А., Суперлано Л., Берман Н., Халл Л., Свердлов Р.С. (1997). «Влияние повышенной температуры мошонки на выработку спермы у нормальных мужчин». Fertil. Стерил. 68 (2): 334–9. Дои:10.1016 / s0015-0282 (97) 81525-7. PMID 9240266.
- ^ Хеллер CG, Клермон Y (1964). «Кинетика зародышевого эпителия человека». Недавнее Prog Horm Res. 20: 545–571. PMID 14285045.
- ^ Аманн Р.П. (2008). «Цикл семенного эпителия у человека: необходимость пересмотра?». Дж. Андрол. 29 (5): 469–487. Дои:10.2164 / jandrol.107.004655. PMID 18497337.
- ^ Форстер П., Хохофф С., Дункельманн Б., Шюренкамп М., Пфайффер Н., Нойхубер Ф., Бринкманн Б. (2015). «Повышенная частота мутаций зародышевой линии у отцов-подростков». Proc R Soc B. 282 (1803): 20142898. Дои:10.1098 / rspb.2014.2898. ЧВК 4345458. PMID 25694621.
- ^ Padubidri, VG; Дафтари, С. Н., ред. (2011). Учебник гинекологии Шоу (15-е изд.). п. 201. ISBN 978-81-312-2548-6.
- ^ Джонсон Л., Петти С.С., Нивз В.Б. (1983). «Дальнейшая количественная оценка сперматогенеза человека: потеря зародышевых клеток во время постпрофазы мейоза и ее связь с ежедневным производством спермы». Биол. Репрод. 29 (1): 207–15. Дои:10.1095 / биолрепрод29.1.207. PMID 6615966.
- ^ Алпатов Р., Леш Б.Дж., Накамото-Киношита М., Бланко А., Чен С., Штютцер А., Армаш К.Дж., Саймон М.Д., Сюй К., Али М., Мурн Дж., Присич С., Кутателадзе Т.Г., Вакоч С.Р., Мин Дж., Кингстон Р.Э., Фишл В., Уоррен С.Т., Пейдж, округ Колумбия, Ши И (май 2014 г.). «Хроматин-зависимая роль хрупкого белка X-умственной отсталости FMRP в ответе на повреждение ДНК». Клетка. 157 (4): 869–81. Дои:10.1016 / j.cell.2014.03.040. ЧВК 4038154. PMID 24813610.
- ^ Фишельсон, Лев; Гон, Офер; Холденгребер, Веред; Делареа, Якоб (2007). «Сравнительный сперматогенез, сперматоцитогенез и образование сперматозоидов у самцов живородящих видов рыб клинид (Teleostei: Clinidae, Blennioidei)». Анатомический рекорд. 290 (3): 311–23. Дои:10.1002 / ar.20412. PMID 17525946.
- ^ Атипичные центриоли во время полового размножения Tomer Avidor-Reiss *, Atul Khire, Emily L. Fishman и Kyoung H. Jo Curr Biol. 2015 16 ноября; 25 (22): 2956-63. DOI: 10.1016 / j.cub.2015.09.045. Epub 2015 17 октября. http://journal.frontiersin.org/article/10.3389/fcell.2015.00021/full
- ^ Hadley, Mac E .; Левин, Джон Э. (2007). Эндокринология (6-е изд.). Река Аппер Сэдл, Нью-Джерси: Prentice Hall. п. 369. ISBN 978-0-13-187606-4.
- ^ а б Сяо, X .; Мрук, Д. Д .; Ченг, К. Ю. (2013). «Молекулы межклеточной адгезии (ICAM) и сперматогенез». Обновление репродукции человека. 19 (2): 167–86. Дои:10.1093 / humupd / dms049. ЧВК 3576004. PMID 23287428.
- ^ Харрисон, Р.Г.; Вайнер, Дж. С. (1949). «Сосудистые паттерны семенников млекопитающих и их функциональное значение». Журнал экспериментальной биологии. 26 (3): 304–16, 2 пл. PMID 15407652.
- ^ Wallach, Edward E .; Kandeel, Fouad R .; Свердлов, Рональд С. (1 января 1988 г.). «Роль температуры в регуляции сперматогенеза и использование нагрева как метода контрацепции». Фертильность и бесплодие. 49 (1): 1–23. Дои:10.1016 / S0015-0282 (16) 59640-X. PMID 3275550.
- ^ Cameron, R.DA .; Блэкшоу, А. У. (1 мая 1980 г.). «Влияние повышенной температуры окружающей среды на сперматогенез у хряка». Размножение. 59 (1): 173–179. Дои:10.1530 / jrf.0.0590173. PMID 7401033.
- ^ Хотта, Ясуо; Фудзисава, Масато; Табата, Сатоши; Стерн, Герберт; Ёсида, Сёнэн (1 сентября 1988 г.). «Влияние температуры на рекомбинационную активность в семенниках грызунов». Экспериментальные исследования клеток. 178 (1): 163–168. Дои:10.1016/0014-4827(88)90387-4. PMID 2900772.
- ^ Lewis, S.E.M .; Айткен, Р. Дж. (24 мая 2005 г.). «Повреждение ДНК сперматозоидов влияет на оплодотворение и беременность». Исследования клеток и тканей. 322 (1): 33–41. Дои:10.1007 / s00441-005-1097-5. PMID 15912407. S2CID 27592293.
- ^ Мехрпур, Омид; Каррари, Париж; Замани, Насим; Tsatsakis, Aristides M .; Абдоллахи, Мохаммад (октябрь 2014 г.). «Профессиональное воздействие пестицидов и их последствия для мужской спермы и фертильности: обзор». Письма токсикологии. 230 (2): 146–156. Дои:10.1016 / j.toxlet.2014.01.029. PMID 24487096.
- ^ а б c Уильям Дж. Кремер; Роголь А.Д. (15 апреля 2008 г.). Энциклопедия спортивной медицины: публикация Медицинской комиссии МОК, Эндокринная система в спорте и физических упражнениях. Джон Вили и сыновья. С. 286–. ISBN 978-0-470-75780-2.
- ^ а б Fody EP, Walker EM (1985). «Влияние лекарств на мужскую и женскую репродуктивную систему». Анна. Clin. Лаборатория. Наука. 15 (6): 451–8. PMID 4062226.
- ^ Вольф-Бернхард Шилль; Фрэнк Х. Комхэр; Тимоти Б. Харгрив (26 августа 2006 г.). Андрология для клинициста. Springer Science & Business Media. С. 76–. ISBN 978-3-540-33713-3.
- ^ Эберхард Нишлаг; Герман М. Бере; Сьюзан Нишлаг (26 июля 2012 г.). Тестостерон: действие, дефицит, замещение. Издательство Кембриджского университета. С. 130–. ISBN 978-1-107-01290-5.
- ^ Pareek, Tej K .; Joshi, Ayesha R .; Саньял, Амартья; Дигхе, Раджан Р. (2007). «Понимание апоптоза мужских половых клеток из-за истощения гонадотропинов, вызванного антагонистами GnRH». Апоптоз. 12 (6): 1085–100. Дои:10.1007 / s10495-006-0039-3. PMID 17268770. S2CID 25378624.
- ^ О'Доннелл Л., Робертсон К.М., Джонс М.Э., Симпсон Э.Р. (2001). «Эстроген и сперматогенез». Endocr. Rev. 22 (3): 289–318. Дои:10.1210 / edrv.22.3.0431. PMID 11399746.
- ^ Карро С., Бураима-Лелонг Х, Делаланд С. (2012). «Роль эстрогенов в сперматогенезе». Передние биоски. 4: 1–11. Дои:10.2741 / e356. PMID 22201851.
- ^ Смит, Эрик П .; Бойд, Джефф; Франк, Грэм Р .; Такахаши, Хироюки; Коэн, Роберт М .; Спекер, Бонни; Уильямс, Тимоти С .; Lubahn, Dennis B .; Корах, Кеннет С. (1994). «Устойчивость к эстрогену, вызванная мутацией гена эстрогеновых рецепторов у человека». Медицинский журнал Новой Англии. 331 (16): 1056–1061. Дои:10.1056 / NEJM199410203311604. ISSN 0028-4793. PMID 8090165.
- ^ Эдмунд С. Сабанег-младший (20 октября 2010 г.). Мужское бесплодие: проблемы и решения. Springer Science & Business Media. С. 83–. ISBN 978-1-60761-193-6.
дальнейшее чтение
- Окано, Цукаса; Ишинива, Хироко; Онума, Манабу; Синдо, Джунджи; Йокохата, Ясуси; Тамаоки, Масанори (23 марта 2016 г.). «Влияние радиации окружающей среды на семенники и сперматогенез у диких крупных японских полевых мышей (Apodemus speciosus) из Фукусимы». Научные отчеты. 6 (1): 23601. Bibcode:2016НатСР ... 623601O. Дои:10.1038 / srep23601. ЧВК 4804236. PMID 27005329.
- Johnson, L .; Blanchard, T.L .; Varner, D.D .; Скратчфилд, W.L. (Ноябрь 1997 г.). «Факторы, влияющие на сперматогенез жеребца». Териогенология. 48 (7): 1199–1216. Дои:10.1016 / s0093-691x (97) 00353-1. PMID 16728209.
- Бардин, C.W. (1991). «Гипофизарно-яичковая ось». В йенах, S.S.C .; Джеффи, Р. Б. (ред.). Репродуктивная эндокринология (3-е изд.). Филадельфия: У. Б. Сондерс. ISBN 0721632068.
- Чемберс, Кристофер В .; Шафер, Мэри-Энн; Адгер, Гувер; Ом-Смит, Мэрилин; Millstein, Susan G .; Ирвин, Чарльз Э .; Шахтер, Юлиус; Милый, Ричард (февраль 1987 г.). «Микрофлора уретры у мальчиков-подростков: связь с сексуальной активностью и негонококковым уретритом». Журнал педиатрии. 110 (2): 314–321. Дои:10.1016 / с0022-3476 (87) 80180-4. PMID 3100755.
- Czyba, J.C .; Жирод, К. (1980). «Развитие нормального яичка». В Хафезе E.S.E. (ред.). Опустившиеся и крипторхидные яички. Гаага: Мартинус Нийхофф. ISBN 9024723337.
- Whitmore, Willet F .; Карш, Лоуренс; Гиттес, Рубен Ф. (октябрь 1985 г.). "Роль зародышевого эпителия и сперматогенеза в привилегированном выживании интратестикулярных трансплантатов". Журнал урологии. 134 (4): 782–786. Дои:10.1016 / s0022-5347 (17) 47438-6. PMID 2863395.
внешняя ссылка
| Биологические термины | |
|---|---|
| Сексуальный воспроизведение | |
| Сексуальность | |
| |