Гипоталамус - Hypothalamus
| Гипоталамус | |
|---|---|
 Расположение гипоталамуса человека | |
 Расположение гипоталамуса (синий) по отношению к гипофизу и остальному мозгу | |
| подробности | |
| Часть | Мозг |
| Идентификаторы | |
| латинский | гипоталамус |
| MeSH | D007031 |
| НейроЛекс МНЕ БЫ | birnlex_734 |
| TA98 | A14.1.08.401 A14.1.08.901 |
| TA2 | 5714 |
| FMA | 62008 |
| Анатомические термины нейроанатомии | |
В гипоталамус (от Древнегреческий ὑπό, "под" и θάλαμος, «камера») является частью мозг который содержит ряд мелких ядра с множеством функций. Одна из важнейших функций гипоталамуса - связывать нервная система к эндокринная система через гипофиз. Гипоталамус расположен ниже таламус и является частью лимбическая система.[1] В терминологии нейроанатомия, он формирует вентральный часть промежуточный мозг. Все позвоночное животное мозг содержит гипоталамус. У людей это размер миндаль.
Гипоталамус отвечает за регуляцию определенных метаболический процессы и другие мероприятия автономная нервная система. Это синтезирует и выделяет определенные нейрогормоны, называется высвобождающие гормоны или гипоталамические гормоны, которые, в свою очередь, стимулируют или ингибируют секрецию гормоны со стороны гипофиза. Гипоталамус контролирует температура тела, голод, важные аспекты воспитания детей и поведение привязанности, жажда,[2] усталость, спать, и циркадные ритмы.[3]
Структура

Гипоталамус разделен на 3 области (супраоптический, туберальный, маммиллярный) в парасагиттальной плоскости, что указывает на передне-заднее расположение; и 3 области (перивентрикулярная, медиальная, латеральная) в коронарной плоскости, что указывает на медиально-латеральное расположение. Ядра гипоталамуса расположены в этих специфических областях и областях.[4] Он обнаружен во всех нервных системах позвоночных. У млекопитающих магноцеллюлярные нейросекреторные клетки в паравентрикулярное ядро и супраоптическое ядро гипоталамуса производят нейрогипофизарные гормоны, окситоцин и вазопрессин. Эти гормоны попадают в кровь в задней доли гипофиза.[5] Значительно меньше парвоцеллюлярные нейросекреторные клетки, нейроны паравентрикулярного ядра, релиз кортикотропин-рилизинг гормон и другие гормоны в гипофизарная портальная система, где эти гормоны диффундируют в передний гипофиз.
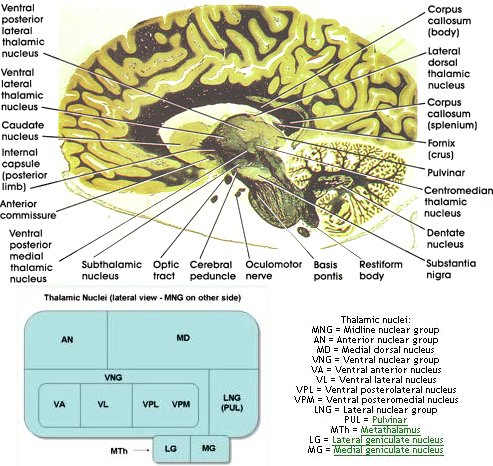
Ядра
Ядра гипоталамуса включают следующее:[6][7][8]
| Область, край | Площадь | Ядро | Функция[9] |
| Передний (супраоптический) | Преоптический | Преоптическое ядро | |
| Медиальный | Медиальное преоптическое ядро |
| |
| Супраоптическое ядро |
| ||
| Паравентрикулярное ядро |
| ||
| Переднее ядро гипоталамуса | |||
| Супрахиазматическое ядро | |||
| Боковой | |||
| Боковое ядро | Увидеть Боковой гипоталамус § Функция - первоисточник орексин нейроны, которые проецируются через головной и спинной мозг | ||
| Средний (туберальный) | Медиальный | Дорсомедиальное ядро гипоталамуса |
|
| Вентромедиальное ядро |
| ||
| Дугообразное ядро |
| ||
| Боковой | Боковое ядро | Увидеть Боковой гипоталамус § Функция - первоисточник орексин нейроны, которые проецируются через головной и спинной мозг | |
| Боковые туберальные ядра | |||
| Задний (маммиллярный) | Медиальный | Маммиллярные ядра (часть маммиллярные тела ) | |
| Заднее ядро |
| ||
| Боковой | Боковое ядро | Увидеть Боковой гипоталамус § Функция - первоисточник орексин нейроны, которые проецируются через головной и спинной мозг | |
| Туберомаммиллярное ядро[11] |
|
- Смотрите также

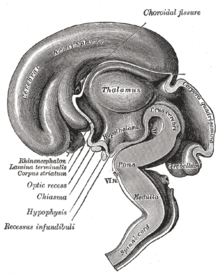
Поперечный разрез гипоталамуса обезьяны показывает два основных ядра гипоталамуса по обе стороны от заполненного жидкостью третьего желудочка.

Ядра гипоталамуса

Ядра гипоталамуса на одной стороне гипоталамуса, показанные на трехмерной компьютерной реконструкции[12]



Подключения
Гипоталамус тесно связан с другими частями Центральная нервная система, в частности ствол мозга и его ретикулярная формация. В рамках лимбическая система, он связан с другими лимбическими структурами, включая миндалина и перегородка, а также связан с областями автономная нервная система.
Гипоталамус получает множество сигналов от мозговой ствол, наиболее заметный из ядро одиночного тракта, то голубое пятно, а вентролатеральный мозг.
Наиболее нервные волокна в гипоталамусе проходят двумя путями (двунаправленными).
- Проекции по площадям каудальный в гипоталамус проходят через медиальный пучок переднего мозга, то маммиллотегментарный тракт и дорсальный продольный пучок.
- Проекции в области, ростральные по отношению к гипоталамусу, выполняются маммиллоталамический тракт, то свод и конечная полоса.
- Проекции на районы симпатическая двигательная система (боковой рог сегменты позвоночника T1-L2 / L3) переносятся гипоталамоспинальный тракт и они активируют симпатический моторный путь.
Половой диморфизм
Несколько ядер гипоталамуса сексуально диморфный; то есть между самцами и самками существуют явные различия как в строении, так и в функциях.[13] Некоторые различия очевидны даже в грубой нейроанатомии: наиболее заметным является половое диморфное ядро в пределах преоптическая область,[13] в котором различия - это тонкие изменения в связности и химической чувствительности определенных наборов нейронов. Важность этих изменений можно понять по функциональным различиям между мужчинами и женщинами. Например, самцы большинства видов предпочитают запах и внешний вид самок самцам, что играет важную роль в стимулировании полового поведения самцов. При поражении полового диморфного ядра это предпочтение самок самцами уменьшается. Также характер секреции гормон роста половой диморфизм;[14] Вот почему у многих видов взрослые самцы заметно отличаются от самок.
Чувствительность к стероидам яичников
Другие поразительные функциональные диморфизмы проявляются в поведенческих реакциях на яичниковые стероиды взрослого. Мужчины и женщины реагируют на стероиды яичников по-разному, отчасти потому, что экспрессия чувствительных к эстрогену нейронов в гипоталамусе сексуально диморфна; т.е. рецепторы эстрогенов экспрессируются в разных наборах нейронов.
Эстроген и прогестерон может влиять на экспрессию генов в определенных нейронах или вызывать изменения в клеточная мембрана потенциал и киназа активация, приводящая к разнообразным негеномным клеточным функциям. Эстроген и прогестерон связываются с родственными им рецепторы ядерных гормонов, которые перемещаются в ядро клетки и взаимодействуют с участками ДНК, известными как элементы гормонального ответа (ОПЧ) или быть привязанным к другому фактор транскрипции сайт привязки. Рецептор эстрогена (ER) таким образом трансактивирует другие факторы транскрипции, несмотря на отсутствие элемент эстрогенового ответа (ERE) в проксимальной промоторной области гена. В целом, ER и рецепторы прогестерона (PR) являются активаторами генов с увеличенной мРНК и последующим синтезом белка после воздействия гормона.[нужна цитата ]
Мужской и женский мозг различаются по распределению рецепторов эстрогена, и это различие является необратимым следствием воздействия неонатальных стероидов. Рецепторы эстрогена (и рецепторы прогестерона) обнаруживаются в основном в нейронах переднего и медиобазального гипоталамуса, а именно:
- то преоптическая область (где LHRH расположены нейроны, регулирующие дофаминовые реакции и материнское поведение;[15]
- то перивентрикулярное ядро где соматостатин расположены нейроны, регулирующие уровень стресса;[16]
- то вентромедиальный гипоталамус который регулирует чувство голода и сексуальное возбуждение.
Развитие
В неонатальном периоде гонадные стероиды влияют на развитие нейроэндокринного гипоталамуса. Например, они определяют способность женщин демонстрировать нормальный репродуктивный цикл, а мужчин и женщин - проявлять соответствующее репродуктивное поведение во взрослой жизни.
- Если самка крысы вводят однократно тестостерон в первые дни послеродовой жизни (в «критический период» влияния половых стероидов), гипоталамус необратимо маскулинизируется; взрослая крыса будет неспособна генерировать всплеск ЛГ в ответ на эстроген (характеристика самок), но будет способна проявлять мужской сексуальное поведение (восхождение на сексуально восприимчивую женщину).[17]
- Напротив, самец крысы кастрированные сразу после рождения будут феминизированный, и взрослый покажет женский пол сексуальное поведение в ответ на эстроген (сексуальная восприимчивость, лордоз поведение ).[17]
У приматов влияние на развитие андрогены менее ясен, и последствия менее понятны. В мозгу тестостерон ароматизируется (чтобы эстрадиол ), который является основным активным гормоном, влияющим на развитие. Человек яичко секретирует высокий уровень тестостерона примерно с 8 недели жизни плода до 5-6 месяцев после рождения (аналогичный перинатальный всплеск тестостерона наблюдается у многих видов), процесс, который, по-видимому, лежит в основе мужского фенотипа. Эстроген из материнского кровотока относительно неэффективен, отчасти из-за высокого уровня циркулирующих стероидсвязывающих белков во время беременности.[17]
Половые стероиды не только важные факторы, влияющие на развитие гипоталамуса; особенно, предпубертатный стресс в раннем возрасте (у крыс) определяет способность взрослого гипоталамуса реагировать на острый стрессор.[18] В отличие от рецепторов гонадных стероидов, глюкокортикоид рецепторы очень широко распространены по всему мозгу; в паравентрикулярное ядро, они обеспечивают контроль отрицательной обратной связи CRF синтез и секреция, но в других местах их роль недостаточно изучена.
Функция
Гормональный выброс
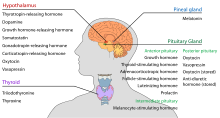
Гипоталамус имеет центральную нейроэндокринный функции, в первую очередь за счет контроля над передний гипофиз, который, в свою очередь, регулирует различные эндокринные железы и органы. Высвобождающие гормоны (также называемые рилизинг-факторами) производятся в ядрах гипоталамуса, а затем транспортируются вдоль аксоны либо к срединное возвышение или задней доли гипофиза, где они хранятся и выпускаются по мере необходимости.[19]
- Передний гипофиз
В гипоталамо-аденогипофизарной оси рилизинг-гормоны, также известные как гипофизиотропные или гипоталамические гормоны, высвобождаются из срединного возвышения, удлинения гипоталамуса, в гипофизарная портальная система, который переносит их в переднюю долю гипофиза, где они выполняют свои регулирующие функции в отношении секреции аденогипофизарных гормонов.[20] Эти гипофизиотропные гормоны стимулируются парвоцеллюлярными нейросекреторными клетками, расположенными в перивентрикулярной области гипоталамуса. После попадания в капилляры третьего желудочка гипофизиотропные гормоны проходят через так называемый гипоталамо-гипофизарный портальный кровоток. Достигнув места назначения в передней доле гипофиза, эти гормоны связываются со специфическими рецепторами, расположенными на поверхности клеток гипофиза. В зависимости от того, какие клетки активируются посредством этого связывания, гипофиз либо начнет секретировать, либо прекратит выделять гормоны в остальную часть кровотока.[21]
Другие гормоны, выделяемые срединным возвышением, включают: вазопрессин, окситоцин, и нейротензин.[23][24][25][26]
- Задней доли гипофиза
В гипоталамо-нейрогипофизарной оси нейрогипофизарные гормоны выделяются в кровоток из задней доли гипофиза, который на самом деле является продолжением гипоталамуса.
| Секретируемый гормон | Сокращение | Произведено | Эффект |
|---|---|---|---|
| Окситоцин | OXY или OXT | Магноцеллюлярные нейросекреторные клетки паравентрикулярного ядра и супраоптическое ядро | Сокращение матки Лактация (рефлекс подавления) |
| Вазопрессин (антидиуретический гормон) | ADH или AVP | Магноцеллюлярные и парвоцеллюлярные нейросекреторные клетки паравентрикулярного ядра, магноцеллюлярные клетки супраоптического ядра | Повышение проницаемости для воды клеток дистальный каналец и сборный канал в почках и, таким образом, обеспечивает реабсорбцию воды и выведение концентрированной мочи |
Также известно, что гипоталамо-гипофизарно-надпочечниковая система (HPA) гормоны связаны с некоторыми кожными заболеваниями и гомеостазом кожи. Имеются данные о связи гиперактивности гормонов HPA со стрессовыми кожными заболеваниями и кожными опухолями.[27]
Стимуляция
Гипоталамус координирует многие гормональные и поведенческие циркадные ритмы, сложные паттерны нейроэндокринный выходы, комплекс гомеостатический механизмы и важные модели поведения. Следовательно, гипоталамус должен реагировать на множество различных сигналов, некоторые из которых генерируются извне, а некоторые - внутренне. Дельта волна передача сигналов, возникающая либо в таламусе, либо в коре головного мозга, влияет на секрецию рилизинг-гормонов; GHRH и пролактин стимулируются пока TRH заблокирован.
Гипоталамус реагирует на:
- Свет: продолжительность светового дня и фотопериод для регулирования циркадный и сезонные ритмы
- Обонятельный стимулы, в том числе феромоны
- Стероиды, в том числе гонадные стероиды и кортикостероиды
- Нейронно передаваемая информация, исходящая, в частности, от сердца, кишечная нервная система (из желудочно-кишечный тракт ),[28] и репродуктивный тракт.[нужна цитата ]
- Автономный входы
- Стимулы, передающиеся через кровь, в том числе лептин, грелин, ангиотензин, инсулин, гормоны гипофиза, цитокины, плазменные концентрации глюкозы и осмолярность и т. д.
- Стресс
- Проникновение микроорганизмов путем повышения температуры тела и установки термостата тела вверх.
Обонятельные раздражители
Обонятельные раздражители важны для половое размножение и нейроэндокринный функционируют у многих видов. Например, если беременная мышь подвергается воздействию мочи «чужого» самца в критический период после полового акта, то беременность не наступает ( Эффект Брюса ). Таким образом, во время коитуса самка мыши формирует точную «обонятельную память» о своем партнере, которая сохраняется в течение нескольких дней. Феромональные сигналы помогают синхронизации эструс у многих видов; у женщин синхронно менструация также могут возникать из-за феромонных сигналов, хотя роль феромонов у людей оспаривается.
Стимулы, передающиеся через кровь
Пептид гормоны имеют важное влияние на гипоталамус, и для этого они должны пройти через гематоэнцефалический барьер. Гипоталамус частично ограничен специализированными областями мозга, в которых отсутствует эффективный гематоэнцефалический барьер; то капилляр эндотелий на этих участках фенестрирован, чтобы обеспечить свободный проход даже больших белков и других молекул. Некоторые из этих участков являются участками нейросекреции - нейрогипофиз и срединное возвышение. Однако в других местах мозг анализирует состав крови. Два из этих сайтов, SFO (подформенный орган ) и OVLT (Organum vasculosum терминальной пластинки ) так называемые окружные желудочковые органы, где нейроны находятся в тесном контакте с кровью и CSF. Эти структуры имеют плотную васкуляризацию и содержат осморецептивные и натрий-рецептивные нейроны, которые контролируют питьевой, вазопрессин релиз, экскреция натрия и аппетит натрия. Они также содержат нейроны с рецепторами для ангиотензин, предсердный натрийуретический фактор, эндотелин и релаксин, каждый из которых важен для регулирования баланса жидкости и электролитов. Нейроны в проекте OVLT и SFO к супраоптическое ядро и паравентрикулярное ядро, а также в преоптические области гипоталамуса. Окружные желудочковые органы также могут быть местом действия интерлейкины вызвать лихорадку и секрецию АКТГ посредством воздействия на паравентрикулярные нейроны.[нужна цитата ]
Неясно, как все пептиды, влияющие на гипоталамическую активность, получают необходимый доступ. На случай, если пролактин и лептин, есть свидетельства активного поглощения в сосудистое сплетение из крови в спинномозговая жидкость (CSF). Некоторые гормоны гипофиза оказывают отрицательное влияние на секрецию гипоталамуса; Например, гормон роста отвечает гипоталамусу, но как он попадает в мозг, не ясно. Есть также свидетельства того, что центральные действия пролактин.[нужна цитата ]
Результаты показали, что гормон щитовидной железы (T4) поглощается гипоталамической глиальные клетки в инфундибулярное ядро / срединное возвышение, и что здесь он преобразован в Т3 дейодиназой 2 типа (D2). После этого Т3 транспортируется в тиреотропин-рилизинг-гормон (TRH ) -производящие нейроны в паравентрикулярное ядро. Рецепторы гормонов щитовидной железы были найдены в этих нейроны, что указывает на то, что они действительно чувствительны к стимулам Т3. Кроме того, эти нейроны экспрессировали MCT8, а гормон щитовидной железы транспортер, подтверждая теорию о том, что Т3 транспортируется в них. Затем Т3 может связываться с рецептором тироидного гормона в этих нейронах и влиять на выработку тиреотропин-рилизинг-гормона, тем самым регулируя выработку гормона щитовидной железы.[29]
Гипоталамус функционирует как тип термостат для тела.[30] Он устанавливает желаемую температуру тела и стимулирует выработку и удержание тепла для повышения температуры крови до более высокого уровня или потоотделение и расширение сосудов для охлаждения крови до более низкой температуры. Все лихорадка результат повышенного настроения в гипоталамусе; повышенная температура тела по любой другой причине классифицируется как гипертермия.[30] Редко прямое повреждение гипоталамуса, например, от Инсульт, вызовет жар; это иногда называют гипоталамическая лихорадка. Однако чаще такое повреждение вызывает аномально низкую температуру тела.[30]
Стероиды
Гипоталамус содержит нейроны, которые сильно реагируют на стероиды и глюкокортикоиды - (стероидные гормоны надпочечник, выпущен в ответ на АКТГ ). Он также содержит специализированные нейроны, чувствительные к глюкозе (в дугообразное ядро и вентромедиальный гипоталамус ), которые важны для аппетит. В преоптической области располагаются термочувствительные нейроны; это важно для TRH секреция.
Нейронный
Окситоцин секреция в ответ на сосание или вагино-цервикальную стимуляцию опосредуется некоторыми из этих путей; вазопрессин секреция в ответ на сердечно-сосудистые раздражители, исходящие от хеморецепторов в сонная артерия и дуга аорты, и от низкого давления рецепторы предсердного объема, опосредуется другими. У крысы стимуляция влагалище также вызывает пролактин секреция, и это приводит к псевдобеременность после бесплодного спаривания. У кролика коитус вызывает рефлекс овуляция. В овце, шейный стимуляция в присутствии высокого уровня эстрогена может вызвать материнское поведение в девственной овце. Все эти эффекты опосредуются гипоталамусом, и информация передается в основном по спинным путям, которые передаются в ствол мозга. Стимуляция сосков стимулирует высвобождение окситоцина и пролактина и подавляет высвобождение LH и ФСГ.
Сердечно-сосудистые раздражители переносятся блуждающий нерв. Блуждающий нерв также передает разнообразную висцеральную информацию, в том числе, например, сигналы, возникающие при растяжении или опорожнении желудка, для подавления или стимулирования кормления, сигнализируя о высвобождении лептин или гастрин соответственно. И снова эта информация достигает гипоталамуса через реле в стволе мозга.
Кроме того, функция гипоталамуса реагирует на все три классических уровня и регулируется ими. моноаминовые нейротрансмиттеры, норадреналин, дофамин, и серотонин (5-гидрокситриптамин), в тех трактах, из которых он получает иннервацию. Например, норадренергические входы, возникающие из голубого пятна, имеют важные регулирующие эффекты на кортикотропин-рилизинг гормон (CRH) уровни.
Контроль приема пищи
| Пептиды, повышающие пищевое поведение | Пептиды, уменьшающие пищевое поведение |
|---|---|
| Грелин | Лептин |
| Нейропептид Y | (α, β, γ) -Гормоны, стимулирующие меланоциты |
| Агути-родственный пептид | Пептиды транскрипции, регулируемые кокаином и амфетамином |
| Орексины (А, Б) | Кортикотропин-рилизинг-гормон |
| Меланин-концентрирующий гормон | Холецистокинин |
| Галанин | Инсулин |
| Глюкагоноподобный пептид 1 |
Крайний боковой часть вентромедиальное ядро гипоталамуса отвечает за контроль еда потребление. Стимуляция этой области вызывает повышенное потребление пищи. Двусторонний поражение этой области вызывает полное прекращение приема пищи. Медиальные части ядра оказывают контролирующее влияние на латеральную часть. Двустороннее поражение медиальной части вентромедиального ядра вызывает гиперфагия и ожирение животного. Дальнейшее поражение боковой части вентромедиального ядра у того же животного приводит к полному прекращению приема пищи.
Существуют разные гипотезы, связанные с этим регулированием:[32]
- Липостатическая гипотеза: эта гипотеза утверждает, что жировой ткань производит гуморальный сигнал, который пропорционален количеству жира и действует на гипоталамус, уменьшая потребление пищи и увеличивая выработку энергии. Было очевидно, что гормон лептин действует на гипоталамус, уменьшая потребление пищи и увеличивая выработку энергии.
- Гипотеза гутпептида: желудочно-кишечный гормоны, такие как Grp, глюкагоны, CCK и другие утверждали, что препятствуют приему пищи. Пища, попадающая в желудочно-кишечный тракт, вызывает выброс этих гормонов, которые воздействуют на мозг, вызывая чувство сытости. Мозг содержит рецепторы CCK-A и CCK-B.
- Глюкостатическая гипотеза: активность центра насыщения в вентромедиальных ядрах, вероятно, регулируется глюкоза утилизация в нейронах. Было высказано предположение, что, когда их использование глюкозы низкое и, следовательно, когда разница в уровне глюкозы в артериовенозной крови у них низкая, активность нейронов снижается. В этих условиях деятельность центра питания не контролируется, и человек чувствует голод. Прием пищи быстро увеличивается за счет внутрижелудочкового введения 2-дезоксиглюкоза следовательно, снижение использования глюкозы в клетках.
- Термостатическая гипотеза: согласно этой гипотезе снижение температуры тела ниже заданного значения стимулирует аппетит, тогда как повышение температуры выше заданного значения подавляет аппетит.
Обработка страха
Медиальная зона гипоталамуса является частью цепи, которая контролирует мотивированное поведение, например, защитное поведение.[33] Анализ Fos -маркировка показала, что ряд ядер в «столбце поведенческого контроля» важен для регуляции выражения врожденного и условного защитного поведения.[34]
- Антихищное защитное поведение
Контакт с хищником (например, с кошкой) вызывает у лабораторных грызунов защитное поведение, даже если животное никогда не контактировало с кошкой.[35] В гипоталамусе это воздействие вызывает увеличение С маркировкой Fos клетки в переднем ядре гипоталамуса, дорсомедиальной части вентромедиального ядра и в вентролатеральной части премамиллярного ядра (PMDvl).[36] Премамиллярное ядро играет важную роль в выражении защитного поведения по отношению к хищнику, поскольку повреждения в этом ядре отменяют защитное поведение, такое как замирание и бегство.[36][37] PMD не модулирует защитное поведение в других ситуациях, так как поражения этого ядра оказали минимальное влияние на баллы замораживания после шока.[37] PMD имеет важные связи с спинным периакведуктальный серый, важная структура в выражении страха.[38][39] Кроме того, животные демонстрируют поведение оценки риска по отношению к окружающей среде, ранее связанной с кошкой. Анализ меченных Fos клеток показал, что PMDv1 является наиболее активированной структурой в гипоталамусе, и инактивация с помощью мусцимол до воздействия контекста отменяет защитное поведение.[36] Следовательно, гипоталамус, в основном PMDv1, играет важную роль в выражении врожденного и условного защитного поведения хищника.
- Социальное поражение
Точно так же гипоталамус играет роль в социальное поражение: Ядра в медиальной зоне также мобилизуются во время встречи с агрессивным сородичем. У побежденного животного наблюдается повышение уровней Fos в половых диморфных структурах, таких как медиальное предоптическое ядро, вентролатеральная часть вентромедиального ядра и вентральное премаммилярное ядро.[40] Такие структуры важны для других форм социального поведения, таких как сексуальное и агрессивное поведение. Более того, мобилизируется и премамиллярное ядро, дорсомедиальная часть, но не вентролатеральная часть.[40] Поражения в этом ядре отменяют пассивное защитное поведение, такое как замораживание и поза «на спине».[40]
Дополнительные изображения


Человеческий мозг в разрезе слева в среднесагиттальном разрезе

Расположение гипоталамуса



Смотрите также
- Копептин
- Гипоталамо-гипофизарно-надпочечниковая ось (Ось HPA)
- Гипоталамо-гипофизарно-гонадная ось (Ось HPG)
- Гипоталамо-гипофизарно-тироидная ось (Ось HPT)
- Инцертогипоталамический путь
- Нейроэндокринология
- Неврология сна
использованная литература
- ^ Доктор Бори, К. Джордж. «Эмоциональная нервная система». Лимбическая система. Получено 18 апреля 2016.
- ^ "Словарь терминов по раку NCI". Национальный институт рака.
- ^ Сапер, Клиффорд Б.; Scammell, Thomas E .; Лу, июнь (2005). «Гипоталамическая регуляция сна и циркадных ритмов». Природа. 437 (7063): 1257–1263. Bibcode:2005Натура.437.1257S. Дои:10.1038 / природа04284. ISSN 1476-4687. PMID 16251950. S2CID 1793658.
- ^ Индербир Сингх (сентябрь 2011 г.). Учебник анатомии: Том 3: Голова и шея, центральная нервная система.. JP Medical Ltd., стр. 1101–. ISBN 978-93-5025-383-0.
- ^ Мелмед, S; Полонский, К.С.; Ларсен, PR; Кроненберг, HM (2011). Учебник эндокринологии Уильямса (12-е изд.). Сондерс. п. 107. ISBN 978-1437703245.
- ^ «Увеличенный вид гипоталамуса». psycheducation.org. Джим Фелпс. Архивировано из оригинал 15 декабря 2005 г.. Получено 7 февраля 2020.
- ^ «Схема ядер». Universe-review.ca. Получено 7 февраля 2020.
- ^ «Эмоции и лимбическая система». utdallas.edu. Люсьен Т. "Трес" Томпсон, Техасский университет в Далласе. Получено 7 февраля 2020.
- ^ Холл, Джон Э .; Гайтон, Артур С. (2011). Учебник медицинской физиологии Гайтона и Холла (12-е изд.). Saunders / Elsevier. ISBN 978-1416045748.
- ^ Йошида К., Ли Х, Кано Дж., Лазарус М., Сапер С.Б. (сентябрь 2009 г.). «Параллельные преоптические пути для терморегуляции». Журнал неврологии. 29 (38): 11954–64. Дои:10.1523 / JNEUROSCI.2643-09.2009. ЧВК 2782675. PMID 19776281.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 6: Широко распространяющиеся системы: моноамины, ацетилхолин и орексин». В Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. С. 175–176. ISBN 9780071481274.
В головном мозге гистамин синтезируется исключительно нейронами с их клеточными телами в туберомаммиллярном ядре (TMN), которое находится в задней части гипоталамуса. У человека примерно 64000 гистаминергических нейронов на каждой стороне. Эти клетки проецируются по всему головному и спинному мозгу. Области, которые получают особенно плотные проекции, включают кору головного мозга, гиппокамп, неостриатум, прилежащее ядро, миндалевидное тело и гипоталамус. ... В то время как наиболее охарактеризованная функция гистаминовой системы в головном мозге - регуляция сна и возбуждения, гистамин также участвует в обучении и памяти ... Также оказывается, что гистамин участвует в регуляции питания и энергетического баланса.
- ^ Бюллетень исследований мозга 35: 323–327, 1994
- ^ а б Хофман М.А., Свааб Д.Ф. (июнь 1989 г.). «Половое диморфное ядро преоптической области человеческого мозга: сравнительное морфометрическое исследование». Журнал анатомии. 164: 55–72. ЧВК 1256598. PMID 2606795.
- ^ Куинни К.М., Бонтуис П.Дж., Харрис Е.П., Шетти С.Р., Риссман Э.Ф. (2015). «Нервный гормон роста: региональная регуляция эстрадиолом и / или дополнением половых хромосом у самцов и самок мышей». Биология половых различий. 6: 8. Дои:10.1186 / s13293-015-0026-х. ЧВК 4434521. PMID 25987976.
- ^ Castañeyra-Ruiz L, González-Marrero I, Castañeyra-Ruiz A, González-Toledo JM, Castañeyra-Ruiz M, de Paz-Carmona H, Castañeyra-Perdomo A, Carmona-Calero EM (2013). «Распределение лютеинизирующего гормона-рилизинг-гормона в переднем гипоталамусе самок крыс». Анатомия ISRN. 2013: 1–6. Дои:10.5402/2013/870721. ЧВК 4392965. PMID 25938107.
- ^ Исгор С., Чекки М., Каббадж М., Акил Х., Уотсон С.Дж. (2003). «Бета-рецептор эстрогена в паравентрикулярном ядре гипоталамуса регулирует нейроэндокринную реакцию на стресс и регулируется кортикостероном». Неврология. 121 (4): 837–45. Дои:10.1016 / S0306-4522 (03) 00561-X. PMID 14580933. S2CID 31026141.
- ^ а б c Маккарти М.М., Арнольд А.П., Болл Г.Ф., Блауштайн Д.Д., Де Врис Г.Дж. (февраль 2012 г.). «Половые различия в мозге: не такая уж и неудобная правда». Журнал неврологии. 32 (7): 2241–7. Дои:10.1523 / JNEUROSCI.5372-11.2012. ЧВК 3295598. PMID 22396398.
- ^ Ромео Р.Д., Беллани Р., Каратсореос И.Н., Чхуа Н., Вернов М., Конрад С.Д., МакИвен Б.С. (апрель 2006 г.). «История стресса и пубертатное развитие взаимодействуют, чтобы сформировать пластичность оси гипоталамус-гипофиз-надпочечники». Эндокринология. 147 (4): 1664–74. Дои:10.1210 / en.2005-1432. PMID 16410296.
- ^ Боуэн, Р. «Обзор гормонов гипоталамуса и гипофиза». Получено 5 октября 2014.
- ^ Мелмед С., Джеймсон Дж. Л. (2005). «Заболевания передней доли гипофиза и гипоталамуса». В Kasper DL, Braunwald E, Fauci AS, et al. (ред.). Принципы внутренней медицины Харрисона (16-е изд.). Нью-Йорк, штат Нью-Йорк: Макгроу-Хилл. стр.2076 –97. ISBN 978-0-07-139140-5.
- ^ Медведь MF, Коннорс BW, Paradiso MA (2016). «Гипоталамический контроль передней доли гипофиза». Нейробиология: исследование мозга (4-е изд.). Филадельфия: Вольтерс Клувер. п. 528. ISBN 978-0-7817-7817-6.
- ^ Бен-Шломо А., Мелмед С. (март 2010 г.). «Передача сигналов рецептора соматостатина гипофиза». Тенденции в эндокринологии и метаболизме. 21 (3): 123–33. Дои:10.1016 / j.tem.2009.12.003. ЧВК 2834886. PMID 20149677.
- ^ Хорн AM, Робинсон IC, Финк G (февраль 1985 г.). «Окситоцин и вазопрессин в портальной крови гипофиза крыс: экспериментальные исследования на нормальных крысах и крысах Brattleboro». Журнал эндокринологии. 104 (2): 211–24. Дои:10.1677 / joe.0.1040211. PMID 3968510.
- ^ Дата Y, Мондал М.С., Мацукура С., Уэта Ю., Ямасита Х., Кайя Х., Кангава К., Наказато М. (март 2000 г.). «Распределение орексина / гипокретина в среднем возвышении крыс и гипофизе». Исследование мозга. Молекулярные исследования мозга. 76 (1): 1–6. Дои:10.1016 / s0169-328x (99) 00317-4. PMID 10719209.
- ^ Ватанобе Х., Такебе К. (апрель 1993 г.). «Высвобождение нейротензина in vivo из среднего уровня у крыс, подвергнутых овариэктомии, примированных эстрогеном, по оценке пушпульной перфузии: корреляция с выбросами лютеинизирующего гормона и пролактина». Нейроэндокринология. 57 (4): 760–4. Дои:10.1159/000126434. PMID 8367038.
- ^ Spinazzi R, Андрейс П.Г., Росси Г.П., Нуссдорфер Г.Г. (март 2006 г.). «Орексины в регуляции гипоталамо-гипофизарно-надпочечниковой оси». Фармакологические обзоры. 58 (1): 46–57. Дои:10.1124 / пр.58.1.4. PMID 16507882. S2CID 17941978.
- ^ Чон Ын Ким; Байк Ки Чо; Дэ Хо Чо; Хён Чжон Пак (2013). «Выражение оси гипоталамус-гипофиз-надпочечники при распространенных кожных заболеваниях: доказательства его связи с активностью заболеваний, связанных со стрессом». Национальный исследовательский фонд Кореи. Получено 4 марта 2014.
- ^ Майер Э.А. (июль 2011 г.). «Внутренние чувства: зарождающаяся биология коммуникации между кишечником и мозгом». Обзоры природы. Неврология. 12 (8): 453–66. Дои:10.1038 / номер 3071. ЧВК 3845678. PMID 21750565.
- ^ Fliers E, Unmehopa UA, Alkemade A (июнь 2006 г.). «Функциональная нейроанатомия обратной связи гормонов щитовидной железы в гипоталамусе и гипофизе человека». Молекулярная и клеточная эндокринология. 251 (1–2): 1–8. Дои:10.1016 / j.mce.2006.03.042. PMID 16707210. S2CID 33268046.
- ^ а б c Фаучи, Энтони; и другие. (2008). Принципы внутренней медицины Харрисона (17-е изд.). McGraw-Hill Professional. стр.117 –121. ISBN 978-0-07-146633-2.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 10: Нейронный и нейроэндокринный контроль внутренней среды - Таблица 10: 3». В Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 263. ISBN 9780071481274.
- ^ Theologides A (май 1976 г.). «Промежуточные метаболиты, вызывающие анорексию». Американский журнал клинического питания. 29 (5): 552–8. Дои:10.1093 / ajcn / 29.5.552. PMID 178168.
- ^ Swanson LW (декабрь 2000 г.). «Церебральная регуляция мотивированного поведения полушарием мозга». Исследование мозга. 886 (1–2): 113–164. Дои:10.1016 / S0006-8993 (00) 02905-X. PMID 11119693. S2CID 10167219.
- ^ Кантерас, Н. (2002). «Медиальная гипоталамическая защитная система: годологическая организация и функциональные последствия». Фармакология, биохимия и поведение. 71 (3): 481–491. Дои:10.1016 / S0091-3057 (01) 00685-2. PMID 11830182. S2CID 12303256.
- ^ Ribeiro-Barbosa ER, Canteras NS, Cezário AF, Blanchard RJ, Blanchard DC (2005). «Альтернативная экспериментальная процедура для изучения защитных реакций хищников». Неврология и биоповеденческие обзоры. 29 (8): 1255–63. Дои:10.1016 / j.neubiorev.2005.04.006. PMID 16120464. S2CID 8063630.
- ^ а б c Сезарио А.Ф., Рибейро-Барбоса ER, Бальдо М.В., Кантерас Н.С. (сентябрь 2008 г.). «Гипоталамические участки, отвечающие на угрозы хищников - роль дорсального премамиллярного ядра в безусловном и условном антихищническом защитном поведении». Европейский журнал нейробиологии. 28 (5): 1003–15. Дои:10.1111 / j.1460-9568.2008.06392.x. PMID 18691328. S2CID 10073236.
- ^ а б Бланшар, округ Колумбия (2003). «Дорсальное предкаммиллярное ядро по-разному модулирует защитное поведение, вызванное различными опасными стимулами у крыс». Письма о неврологии. 345 (3): 145–148. Дои:10.1016 / S0304-3940 (03) 00415-4. PMID 12842277. S2CID 16406187.
- ^ Кантерас Н.С., Суонсон Л.В. (ноябрь 1992 г.). «Дорсальное премаммиллярное ядро: необычный компонент маммиллярного тела». Труды Национальной академии наук Соединенных Штатов Америки. 89 (21): 10089–93. Bibcode:1992PNAS ... 8910089C. Дои:10.1073 / pnas.89.21.10089. ЧВК 50283. PMID 1279669.
- ^ Бехбехани М.М. (август 1995 г.). «Функциональные характеристики периакведуктального серого среднего мозга». Прогресс в нейробиологии. 46 (6): 575–605. Дои:10.1016 / 0301-0082 (95) 00009-К. PMID 8545545. S2CID 24690642.
- ^ а б c Motta SC, Goto M, Gouveia FV, Baldo MV, Canteras NS, Swanson LW (март 2009 г.). «Анализ системы страха мозга показывает, что гипоталамус имеет решающее значение для реагирования на вторжение второстепенных представителей определенного вида». Труды Национальной академии наук Соединенных Штатов Америки. 106 (12): 4870–5. Bibcode:2009ПНАС..106.4870М. Дои:10.1073 / pnas.0900939106. ЧВК 2660765. PMID 19273843.
{kind=link}
дальнейшее чтение
- de Vries GJ, Södersten P (май 2009 г.). «Половые различия в мозге: взаимосвязь между структурой и функцией». Гормоны и поведение. 55 (5): 589–96. Дои:10.1016 / j.yhbeh.2009.03.012. ЧВК 3932614. PMID 19446075.
внешние ссылки
- Окрашенные изображения срезов головного мозга, в том числе «гипоталамус» на Проект BrainMaps
- Гипоталамус и гипофиз на endotexts.org
- Поиск NIF - Гипоталамус через Информационная структура по неврологии
- Диаграммы заполнения пространства и поперечные сечения ядер гипоталамуса: правый гипоталамус, передний, трубчатый, задний.
| Авторитетный контроль |
|---|