Индуцированные стволовые клетки - Induced stem cells
Индуцированные стволовые клетки (iSC) находятся стволовые клетки происходит от соматический, репродуктивный, плюрипотентный или другие типы клеток намеренно эпигенетический перепрограммирование. Они классифицируются как тотипотент (iTC), плюрипотентный (ИПСК) или прародитель (мультипотентная - iMSC, также называемая индуцированной мультипотентной клеткой-предшественником - iMPC) или всесильный - (iUSC) согласно их потенциал развития и степень дедифференцировки. Прародители получают так называемыми прямое перепрограммирование или направил дифференциация и называются также индуцированными соматические стволовые клетки.
Широко известны три метода:[1]
- Трансплантация ядер взяты из соматических клеток в ооцит (яйцеклетку) без собственного ядра (удаляется в лаборатории)[2][3][4][5]
- Слияние соматических клеток с плюрипотентными стволовыми клетками[6] и
- Трансформация соматических клеток в стволовые клетки с использованием генетического материала, кодирующего перепрограммирование белковых факторов,[7][8][9] рекомбинантные белки;[10] микроРНК,[11][12][13][14][15] синтетическая самовоспроизводящаяся полицистронная РНК[16] и низкомолекулярные биологически активные вещества.[17][18][19]
Естественные процессы
В 1895 г. Томас Морган удалил один из лягушка два бластомеры и обнаружил, что амфибии могут образовывать целое эмбрионы из оставшейся части. Это означало, что клетки могут изменять свой путь дифференцировки. В 1924 году Спеманн и Мангольд продемонстрировали ключевое значение клеточно-клеточной индукции во время развития животных.[20] Обратимое преобразование клеток одного дифференцированного типа клеток в другой называется метаплазия.[21] Этот переход может быть частью нормального процесса созревания или вызван побуждением.
Одним из примеров является преобразование Ирис клетки в линза клетки в процессе созревания и трансформации пигментный эпителий сетчатки клетки в нервную сетчатку во время регенерации у взрослых тритон глаза. Этот процесс позволяет организму заменять клетки, не подходящие для новых условий, более подходящими новыми клетками. В Дрозофила имагинальные диски, клетки должны выбирать из ограниченного числа стандартных состояний дискретной дифференцировки. Тот факт, что трансдетерминация (изменение пути дифференцировки) часто происходит для группы клеток, а не для отдельных клеток, показывает, что она индуцируется, а не является частью созревания.[22]
Исследователи смогли определить минимальные условия и факторы, которых было бы достаточно для запуска каскада молекулярных и клеточных процессов, чтобы дать команду плюрипотентным клеткам организовать эмбрион. Они показали, что противоположные градиенты из костный морфогенетический белок (BMP) и Узловой, два трансформирующий фактор роста члены семьи, которые действуют как морфогены, достаточны, чтобы вызвать молекулярные и клеточные механизмы, необходимые для организации, in vivo или же in vitro, незафиксированные ячейки из данио бластула животный полюс в хорошо развитую эмбрион.[23]
Некоторые типы зрелых специализированных взрослых клеток могут естественным образом превращаться в стволовые клетки. Например, «главные» клетки экспрессируют маркер стволовых клеток Troy. Хотя они обычно производят пищеварительную жидкость для желудка, они могут превращаться в стволовые клетки для временного ремонта повреждений желудка, таких как порез или повреждение от инфекции. Более того, они могут совершать этот переход даже при отсутствии заметных повреждений и способны восполнять целые желудочные единицы, по сути выступая в качестве неподвижных «резервных» стволовых клеток.[24] Дифференцированные эпителиальные клетки дыхательных путей могут превращаться в стабильные и функциональные стволовые клетки. in vivo.[25]После повреждения зрелые окончательно дифференцированные клетки почек дедифференцируются в более примордиальные версии самих себя, а затем дифференцируются в типы клеток, нуждающиеся в замене в поврежденной ткани.[26] Макрофаги могут самообновляться за счет локальной пролиферации зрелых дифференцированных клеток.[27][28] У тритонов мышечная ткань регенерируется из специализированных мышечных клеток, которые дедифференцируются и забывают, каким типом клеток они были. Эта способность к регенерации не снижается с возрастом и может быть связана с их способностью производить новые стволовые клетки из мышечных клеток по мере необходимости.[29]
Разнообразные неопухолевые стволовые клетки обладают способностью генерировать несколько типов клеток. Например, многолинейно-дифференцирующие стрессоустойчивые (Муза) клетки - это стрессоустойчивые стволовые клетки взрослого человека, которые могут самообновляться. Они образуют характерные кластеры клеток в суспензионной культуре, которые экспрессируют набор генов, связанных с плюрипотентностью, и могут дифференцироваться в энтодермальный, эктодермальный и мезодермальные клетки как in vitro, так и in vivo.[30][31][32][33][34]
Другие хорошо задокументированные примеры трансдифференциация подробно описаны их значение в развитии и возрождении.[35][36]

Индуцированные тотипотентные клетки
SCNT-опосредованный
Индуцированные тотипотентные клетки можно получить путем перепрограммирования соматических клеток с помощью перенос ядра соматической клетки (SCNT). Процесс включает в себя отсасывание ядра соматической (телесной) клетки и введение его в ооцит, ядро которого было удалено.[3][5][37][38][39][40]
Используя подход, основанный на протоколе, описанном Tachibana et al.,[3] hESCs могут быть созданы с помощью SCNT с использованием ядер дермальных фибробластов как у 35-летнего мужчины среднего возраста, так и у пожилого 75-летнего мужчины, что позволяет предположить, что возрастные изменения не обязательно являются препятствием для ядерного репрограммирования на основе SCNT клеток человека.[41] Такое перепрограммирование соматических клеток в плюрипотентное состояние имеет огромные возможности для регенеративная медицина. К сожалению, клетки, созданные с помощью этой технологии, потенциально не полностью защищены от иммунная система пациента (донора ядер), потому что у них одинаковые митохондриальный ДНК, как донор ооцитов, вместо митохондриальной ДНК пациентов. Это снижает их ценность как источника трансплантация аутологичных стволовых клеток терапия, а пока,[42] неясно, может ли он вызвать иммунный ответ пациента после лечения.
Индуцированные андрогенетические гаплоидные эмбриональные стволовые клетки можно использовать вместо сперматозоидов для клонирования. Эти клетки, синхронизированные в фазе М и введенные в ооцит, могут дать жизнеспособное потомство.[43]
Эти разработки вместе с данными о возможности получения неограниченного количества ооцитов из митотически активных репродуктивных стволовых клеток,[44] предлагают возможность промышленного производства трансгенных сельскохозяйственных животных. Повторное клонирование жизнеспособных мышей с помощью метода SCNT, который включает ингибитор гистондеацетилазы, трихостатин, добавленный в среду для культивирования клеток,[45] показать, что можно повторно клонировать животных на неопределенный срок без видимого накопления ошибок перепрограммирования или геномных ошибок[46] Однако исследования в области технологий получения сперматозоидов и яйцеклеток из стволовых клеток повышаются. биоэтический вопросы.[47]
Такие технологии также могут иметь далеко идущие клинические применения для преодоления цитоплазматических дефектов в ооцитах человека.[3][48] Например, технология может предотвратить унаследованные митохондриальная болезнь от передачи будущим поколениям. Генетический материал митохондрий передается от матери к ребенку. Мутации могут вызывать диабет, глухоту, нарушения зрения, желудочно-кишечные расстройства, болезни сердца, слабоумие и другие неврологические заболевания. Ядро одного человеческого яйца было перенесено в другое, включая его митохондрии, в результате чего образовалась клетка, которая может считаться имеющей двух матерей. Затем яйца были оплодотворены, и полученные эмбриональные стволовые клетки несли замененную митохондриальную ДНК.[49]В качестве доказательства того, что метод безопасен, автор этого метода указывает на существование здоровых обезьян, которым сейчас более четырех лет, и которые являются продуктом митохондриальных трансплантатов с разным генетическим происхождением.[50]
В позднем поколении теломераза -дефицитных (Terc - / -) мышей, SCNT-опосредованное репрограммирование смягчает дисфункцию теломер и митохондриальные дефекты в большей степени, чем репрограммирование на основе iPSC.[51]
Были описаны другие достижения в клонировании и тотипотентной трансформации.[52]
Получено без SCNT
Недавно некоторым исследователям удалось получить тотипотентные клетки без помощи SCNT. Тотипотентные клетки получали с использованием эпигенетических факторов, таких как зародышевая изоформа гистона ооцита.[53]Перепрограммирование in vivo путем временной индукции четырех факторов Oct4, Sox2, Klf4 и c-Myc у мышей придает свойства тотипотентности. Внутрибрюшинная инъекция таких iPS-клеток in vivo приводит к образованию структур, подобных эмбриону, которые экспрессируют эмбриональные и внезародышевые (трофэктодермальный ) маркеры.[54]Потенциал развития плюрипотентных стволовых клеток мышей для получения как эмбриональных, так и экстраэмбриональных клонов также может быть увеличен с помощью микроРНК. miR-34a дефицит, ведущий к сильной индукции эндогенного ретровирусы МУЭРВ-Л (MERVL).[55][56]
Омоложение до ИПСК

ИПСК впервые были получены в виде трансплантируемых тератокарцинома индуцируется трансплантатами, взятыми из эмбрионов мыши.[57] Тератокарцинома образовалась из соматических клеток.[58] Генетически мозаичные мыши были получены из клеток злокачественной тератокарциномы, что подтверждает их плюрипотентность.[59][60][61] Оказалось, что клетки тератокарциномы способны поддерживать культуру плюрипотентных эмбриональная стволовая клетка в недифференцированном состоянии, снабжая питательную среду различными факторами.[62] В 1980-х годах стало ясно, что трансплантация плюрипотентных / эмбриональных стволовых клеток в организм взрослых млекопитающих обычно приводит к образованию тератомы, которая затем может превратиться в злокачественную тератокарциному.[63] Однако введение клеток тератокарциномы в эмбрион на стадии бластоцисты привело к их включению в внутренняя клеточная масса и часто получали нормальное химерное (то есть состоящее из клеток разных организмов) животное.[64][65][66] Это указывало на то, что причиной тератомы является диссонанс - взаимное недопонимание между молодыми донорскими клетками и окружающими взрослыми клетками (так называемый "реципиент"ниша ").
В августе 2006 года японские исследователи отказались от ооцита, как в случае с SCNT. Перепрограммируя эмбриональную мышь фибробласты в плюрипотентные стволовые клетки посредством эктопической экспрессии четырех факторы транскрипции, а именно 4 октября, Sox2, Klf4 и c-Myc, они доказали, что сверхэкспрессия небольшого числа факторов может подтолкнуть клетку к переходу в новое стабильное состояние, которое связано с изменениями активности тысяч генов.[7]

Таким образом, механизмы репрограммирования связаны, а не независимы, и сосредоточены на небольшом количестве генов.[67]Свойства IPSC очень похожи на ESC.[68] Было показано, что ИПСК поддерживают разработку мышей, полностью состоящих из ИПСК, с использованием тетраплоид (4n) эмбрион,[69] самый строгий анализ потенциала развития. Тем не менее, некоторые генетически нормальные ИПСК не смогли продуцировать полностью ИПСК мышей из-за аберрантного эпигенетического молчания импринтированных Ген Dlk1-Dio3 кластер.[18] Команда, возглавляемая Хансом Шёлером (открывшим ген Oct4 еще в 1989 году), показала, что сверхэкспрессия Oct4 вызывает массовую активацию нецелевого гена во время репрограммирования, ухудшая качество ИПСК. По сравнению с OSKM (Oct4, Sox2, Klf4 и c-Myc), которые демонстрируют паттерны патологического импринтинга и дифференцировки, перепрограммирование SKM (Sox2, Klf4 и c-Myc) генерирует ИПСК с высоким потенциалом развития (почти в 20 раз выше, чем у OSKM). эквивалентно эмбриональная стволовая клетка, что определяется их способностью генерировать мышей, полностью инфицированных ИПСК, посредством комплементации тетраплоидных эмбрионов.[70][71]
Важным преимуществом ИПСК перед ЭСК является то, что они могут происходить из взрослых клеток, а не из эмбрионов. Таким образом, появилась возможность получать ИПСК от взрослых и даже пожилых пациентов.[9][72][73]
Перепрограммирование соматических клеток на ИПСК приводит к омоложению. Было обнаружено, что репрограммирование приводит к удлинению и последующему укорочению теломер после их дифференцировки обратно в фибробластоподобные производные.[74] Таким образом, репрограммирование приводит к восстановлению длины теломер эмбриона,[75] и, следовательно, увеличивает потенциальное число клеточных делений, иначе ограниченное Лимит Хейфлика.[76]
Однако из-за диссонанса между омоложенными клетками и окружающей нишей более старых клеток реципиента инъекция его собственного ИПСК обычно приводит к иммунная реакция,[77] которые можно использовать в медицинских целях,[78] или образование опухолей, таких как тератома.[79] Предполагается, что причина заключается в том, что некоторые клетки, дифференцированные от ESC и iPSC in vivo, продолжают синтезировать эмбриональные клетки. изоформы белка.[80] Таким образом, иммунная система может обнаруживать и атаковать клетки, которые не взаимодействуют должным образом.
Небольшая молекула под названием MitoBloCK-6 может заставить плюрипотентные стволовые клетки умирать, вызывая апоптоз (через цитохром с выпуск через митохондриальный внешняя мембрана) в плюрипотентных стволовых клетках человека, но не в дифференцированных клетках. Вскоре после дифференцировки дочерние клетки стали устойчивыми к смерти. Когда MitoBloCK-6 вводили в дифференцированные клеточные линии, клетки оставались здоровыми. Было высказано предположение, что ключ к их выживанию связан с изменениями, которым подвергаются митохондрии плюрипотентных стволовых клеток в процессе дифференцировки клеток. Эта способность MitoBloCK-6 разделять плюрипотентные и дифференцированные клеточные линии может снизить риск тератом и других проблем в регенеративной медицине.[81]
В 2012 году другие маленькие молекулы (селективные цитотоксические ингибиторы плюрипотентных стволовых клеток человека - hPSC), которые предотвращают образование тератом у мышей плюрипотентными стволовыми клетками человека. Самое сильное и селективное из них (ПлюриСин №1) подавляет стеароил-коА десатураза (ключевой фермент в олеиновая кислота биосинтез), что в конечном итоге приводит к апоптозу. С помощью этой молекулы недифференцированные клетки могут быть выборочно удалены из культуры.[82][83] Эффективная стратегия избирательного удаления плюрипотентных клеток с потенциалом тератомы направлена на специфические для плюрипотентных стволовых клеток антиапоптотический фактор (ы) (т.е. сурвивин или Bcl10). Однократное лечение химическими ингибиторами сурвивина (например, кверцетин или YM155) могут вызывать селективную и полную гибель клеток недифференцированных hPSC и, как утверждается, достаточны для предотвращения образования тератомы после трансплантации.[84] Однако маловероятно, что какое-либо предварительное разрешение может обеспечить повторную посадку iPSC или ESC. После избирательного удаления плюрипотентных клеток они быстро появляются снова, превращая дифференцированные клетки в стволовые, что приводит к опухолям.[85] Это может быть связано с расстройством let-7 регуляция его мишени Nr6a1 (также известного как Ядерный фактор зародышевой клетки - GCNF), эмбриональный репрессор транскрипции генов плюрипотентности, который регулирует экспрессию генов во взрослых фибробластах после микро-РНК потеря миРНК.[86]
Образование тератомы плюрипотентными стволовыми клетками может быть вызвано низкой активностью Фермент PTEN, как сообщается, способствуют выживанию небольшой популяции (0,1–5% от общей популяции) высоко канцерогенных, агрессивных, вызывающих тератому эмбрионоподобных клеток карциномы во время дифференцировки. Выживание этих клеток-инициаторов тератомы связано с неудачной репрессией Наног а также склонность к повышенному метаболизму глюкозы и холестерина.[87] Эти клетки-инициаторы тератомы также экспрессировали более низкое соотношение p53 / p21 по сравнению с неканцерогенными клетками.[88]В связи с указанными выше проблемами безопасности использование ИПСК для клеточной терапии все еще ограничено.[89] Однако их можно использовать для множества других целей, включая моделирование заболеваний,[90] скрининг (выборочный отбор) лекарственных препаратов, тестирование на токсичность различных препаратов.[91]

Ткани, выращенные из ИПСК, помещенных в «химерные» эмбрионы на ранних стадиях развития мыши, практически не вызывают иммунного ответа (после того, как эмбрионы вырастают во взрослых мышей) и подходят для аутотрансплантация[92]В то же время полное перепрограммирование взрослых клеток in vivo в тканях путем временной индукции четырех факторов Oct4, Sox2, Klf4 и c-Myc у мышей приводит к появлению тератом из множества органов.[54] Более того, частичное репрограммирование клеток в сторону плюрипотентности in vivo у мышей демонстрирует, что неполное репрограммирование влечет за собой эпигенетические изменения (неудачное подавление Поликомб цели и измененные Метилирование ДНК ) в клетках, которые вызывают развитие рака.[93]
Химическое побуждение
Используя исключительно маленькие молекулы, Deng Hongkui и его коллеги продемонстрировали, что эндогенных «мастер-генов» достаточно для репрограммирования клеточных судеб. Они индуцировали плюрипотентное состояние во взрослых клетках мышей, используя семь низкомолекулярных соединений.[17]Эффективность метода довольно высока: он смог преобразовать 0,02% клеток взрослой ткани в ИПСК, что сопоставимо со скоростью конверсии вставки генов. Авторы отмечают, что мыши, полученные из ЦПСК, были "на 100% жизнеспособными и, по-видимому, здоровы до 6 месяцев ». Таким образом, эта стратегия химического репрограммирования потенциально может использоваться для создания функциональных желаемых типов клеток для клинического применения.[95][96]
В 2015 году была создана надежная система химического перепрограммирования с выходом в 1000 раз больше, чем в ранее сообщенном протоколе. Таким образом, химическое перепрограммирование стало многообещающим методом управления судьбой клеток.[97]
Дифференциация от индуцированной тератомы
Тот факт, что ИПСК человека способны образовывать тератомы не только в организме человека, но и в организме некоторых животных, в частности у мышей или свиней, позволил разработать метод дифференциации ИПСК in vivo. С этой целью ИПСК с агентом для индукции дифференцировки в клетки-мишени вводят в генетически модифицированный свинья или мышь, которые подавили активацию иммунной системы на человеческих клетках. Образовавшаяся тератома вырезается и используется для выделения необходимых дифференцированных человеческих клеток.[98] посредством моноклональное антитело к тканеспецифическим маркерам на поверхности этих клеток. Этот метод успешно используется для производства функциональных миелоидных, эритроидных и лимфоидных клеток человека, пригодных для трансплантации (пока только мышам).[99]Мыши, которым прививали гематопоэтические клетки, происходящие из тератомы ИПСК человека, продуцировали В- и Т-клетки человека, способные к функциональным иммунным ответам. Эти результаты дают надежду на то, что создание индивидуальных клеток пациента in vivo возможно, обеспечивая материалы, которые могут быть полезны для трансплантации, создания человеческих антител и скрининга лекарств. Использование MitoBloCK-6[81] и / или PluriSIn # 1, дифференцированные клетки-предшественники могут быть дополнительно очищены от тератомы, образующей плюрипотентные клетки. Тот факт, что дифференцировка происходит даже в нише тератомы, дает надежду на то, что полученные клетки будут достаточно стабильными для стимулов, способных вызвать их переход обратно в дедифференцированное (плюрипотентное) состояние и, следовательно, безопасными. Сходная система дифференцировки in vivo, дающая приживляемые гемопоэтические стволовые клетки из ИПСК мыши и человека у животных с тератомой, в сочетании с маневром для облегчения гемопоэза, была описана Suzuki et al.[100] Они отметили, что ни лейкемия, ни опухоли не наблюдались у реципиентов после внутривенной инъекции полученных из ИПСК гемопоэтических стволовых клеток облученным реципиентам. Более того, эта инъекция привела к многолинейному и долгосрочному восстановлению гемолимфопоэтической системы при серийных переносах. Такая система представляет собой полезный инструмент для практического применения ИПСК при лечении гематологических и иммунологических заболеваний.[101]
Для дальнейшего развития этого метода животное, в котором выращивают трансплантат человеческой клетки, например мышь, должно иметь настолько модифицированный геном, чтобы все его клетки экспрессировали и имели на своей поверхности человеческий SIRPα.[102]Чтобы предотвратить отторжение после трансплантации пациенту аллогенного органа или ткани, выращенных из плюрипотентных стволовых клеток in vivo у животного, эти клетки должны экспрессировать две молекулы: CTLA4-Ig, который нарушает костимуляторные пути Т-клеток и PD-L1, который активирует путь ингибирования Т-клеток.[103]
Смотрите также: США 20130058900 патент.
Дифференцированные типы клеток
Клетки сетчатки
В ближайшем будущем начнутся клинические испытания, призванные продемонстрировать безопасность использования ИПСК для клеточной терапии людей с возрастной дегенерацией желтого пятна, заболеванием, вызывающим слепоту из-за повреждения сетчатки. Есть несколько статей, описывающих методы получения клеток сетчатки из ИПСК.[104][105]и как использовать их для клеточной терапии.[106][107] Сообщения о трансплантации пигментного эпителия сетчатки, полученного из ИПСК, показали улучшенное поведение под контролем зрения экспериментальных животных в течение 6 недель после трансплантации.[108] Однако клинические испытания были успешными: у десяти пациентов, страдающих пигментным ретинитом, было восстановлено зрение, в том числе у женщины, у которой осталось только 17 процентов зрения.[109]
Эпителиальные клетки легких и дыхательных путей
Хронические заболевания легких, такие как идиопатический фиброз легких и муковисцидоз или хроническая обструктивная болезнь легких и астма являются ведущими причинами заболеваемости и смертности во всем мире, вызывая значительное человеческое, социальное и финансовое бремя. Таким образом, существует острая необходимость в эффективной клеточной терапии и легкое тканевая инженерия.[110][111]Несколько протоколов были разработаны для генерации большинства типов клеток дыхательная система, что может быть полезно для получения терапевтических клеток, специфичных для пациента.[112][113][114][115][116]
Репродуктивные клетки
Некоторые линии ИПСК обладают потенциалом дифференцироваться в мужские половые клетки и ооцитоподобные клетки в соответствующей нише (путем культивирования в ретиноевой кислоте и среде для дифференцировки фолликулярной жидкости свиньи или трансплантации семенных канальцев). Более того, трансплантация ИПСК вносит вклад в восстановление семенников бесплодных мышей, демонстрируя потенциальную возможность образования гамет из ИПСК in vivo и in vitro.[117]
Индуцированные стволовые клетки-предшественники
Прямая трансдифференцировка
Риск рака и опухолей создает необходимость в разработке методов более безопасных клеточных линий, подходящих для клинического использования. Альтернативный подход - так называемое «прямое репрограммирование» - трансдифференцировка клеток без перехода через плюрипотентное состояние.[118][119][120][121][122][123][124] В основе этого подхода лежало то, что 5-азацитидин - реагент деметилирования ДНК - может вызывать образование миогенный, хондрогенные и адипогенные клоны в линии бессмертных клеток эмбриональных фибробластов мыши[125] и что активации одного гена, позже названного MyoD1, достаточно для такого перепрограммирования.[126] По сравнению с ИПСК, для репрограммирования которых требуется не менее двух недель, образование индуцированных клеток-предшественников иногда происходит в течение нескольких дней, а эффективность репрограммирования обычно во много раз выше. Это перепрограммирование не всегда требует деления клеток.[127] Клетки, полученные в результате такого перепрограммирования, более подходят для клеточной терапии, поскольку они не образуют тератом.[123]Например, Chandrakanthan et al., & Pimanda описывают создание ткане-регенеративных мультипотентных стволовых клеток (iMS-клеток) путем временной обработки зрелых костных и жировых клеток фактором роста (фактор роста тромбоцитов –AB (PDGF-AB)) и 5-азацитидин. Эти авторы утверждают, что: «В отличие от первичных мезенхимальных стволовых клеток, которые используются в клинической практике с небольшим количеством объективных данных для ускорения восстановления тканей, iMS-клетки вносят непосредственный вклад в регенерацию ткани in vivo контекстно-зависимым образом, не образуя опухоли», и поэтому значительный простор для применения в регенерации тканей ».[128][129][130]
Трансдифференцировка одного фактора транскрипции
Изначально только ранние эмбриональные клетки можно было уговорить изменить свою идентичность. Зрелые клетки устойчивы к изменению своей идентичности после того, как они перейдут к определенному виду. Однако кратковременная экспрессия одного фактора транскрипции, фактора ELT-7 GATA, может преобразовать идентичность полностью дифференцированных, специализированных неэнтодермальных клеток глотка в полностью дифференцированные клетки кишечника в неповрежденном личинки и взрослые аскариды Caenorhabditis elegans без необходимости в дедифференцированном промежуточном продукте.[131]
Трансдифференцировка с помощью активатора, опосредованного CRISPR
Судьбой клетки можно эффективно управлять с помощью редактирование эпигенома. В частности, путем прямой активации экспрессии специфических эндогенных генов с помощью CRISPR -опосредованный активатор. Когда dCas9 (который был модифицирован так, что он больше не разрезает ДНК, но все еще может быть направлен к конкретным последовательностям и связываться с ними) в сочетании с активаторами транскрипции, он может точно управлять экспрессией эндогенных генов. Используя этот метод, Wei et al. Увеличили экспрессию эндогенного Cdx2 и Gata6 гены с помощью активаторов, опосредованных CRISPR, таким образом, непосредственно превращая мышиные эмбриональные стволовые клетки в две внеэмбриональные линии, то есть в типичные стволовые клетки трофобласта и внэмбриональные клетки энтодермы.[132] Аналогичный подход был использован для индукции активации эндогенных генов Brn2, Ascl1 и Myt1l для преобразования эмбриональных фибробластов мыши в индуцированные нейрональные клетки.[133] Таким образом, активация транскрипции и эпигенетическое ремоделирование эндогенных основных факторов транскрипции достаточны для конверсии между типами клеток. Быстрая и устойчивая активация эндогенных генов в их природном хроматиновом контексте с помощью этого подхода может облегчить репрограммирование с помощью временных методов, которые избегают геномной интеграции и обеспечивают новую стратегию преодоления эпигенетических барьеров на пути спецификации клеточных судеб.
Поэтапное моделирование процесса регенерации
Другой способ перепрограммирования - моделирование процессов, происходящих во время амфибия регенерация конечностей. В уроделе амфибий, ранним шагом в регенерации конечностей является дедифференцировка волокон скелетных мышц до клеточных элементов, которые пролиферируют в ткани конечностей. Однако последовательная обработка мышечного волокна миосеверином малыми молекулами, обратный (в киназа Aurora B ингибитор) и некоторые другие химические вещества: BIO (ингибитор киназы гликогенсинтазы-3), лизофосфатидная кислота (плейотропный активатор рецепторов, связанных с G-белком), SB203580 (p38 MAP киназа ингибитор), или SQ22536 (ингибитор аденилатциклазы) вызывает образование новых типов мышечных клеток, а также других типов клеток, таких как предшественники жировых клеток, клеток костей и нервной системы.[134]
Трансдифференцировка на основе антител
Исследователи обнаружили, что GCSF - подражание антитело может активировать рецептор, стимулирующий рост, на костный мозг клетки таким образом, что стволовые клетки костного мозга, которые обычно развиваются в лейкоциты, становятся нейральными клетками-предшественниками. Техника[135] позволяет исследователям искать в больших библиотеках антител и быстро выбирать те, которые обладают желаемым биологическим эффектом.[136][137][138]
Перепрограммирование бактериями
Желудочно-кишечный тракт человека заселен огромным сообществом симбионтов и комменсалов. Исследователи демонстрируют феномен репрограммирования соматических клеток бактериями и генерации мультипотенциальных клеток из клеток дермальных фибробластов взрослого человека путем включения молочнокислых бактерий. [139] Эта клеточная трансдифференцировка вызывается рибосомами и «может происходить через донорские бактерии, которые проглатываются и перевариваются клетками-хозяевами, что может вызывать рибосомный стресс и стимулировать пластичность клеточного развития».[140]
Условно перепрограммированные клетки
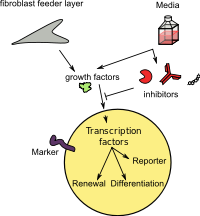
Шлегель и Лю[141] продемонстрировали, что комбинация фидерных ячеек[142][143][144] и Ро-киназа ингибитор (Y-27632) [145][146] побуждает нормальные и опухолевые эпителиальные клетки многих тканей бесконечно размножаться in vitro. Этот процесс происходит без необходимости трансдукции экзогенных вирусных или клеточных генов. Эти клетки были названы «условно перепрограммированными клетками (CRC)».[147] Индукция CRC происходит быстро и является результатом перепрограммирования всей популяции клеток. CRC не экспрессируют высокие уровни белков, характерных для ИПСК или эмбриональных стволовых клеток (ЭСК) (например, Sox2, Oct4, Nanog или Klf4). Эта индукция CRC обратима, и удаление Y-27632 и питателей позволяет клеткам нормально дифференцироваться.[141][148][149] Технология CRC может генерировать 2×106 клетки через 5-6 дней после биопсии иглой и могут генерировать культуры из криоконсервированной ткани и менее чем из четырех жизнеспособных клеток. CRC сохраняют нормальный кариотип и остаются неопухолевыми. Этот метод также позволяет эффективно создавать культуры клеток из опухолей человека и грызунов.[141][150][151]
Способность быстро генерировать множество опухолевых клеток из небольших образцов биопсии и замороженной ткани предоставляет значительные возможности для клеточной диагностики и лечения (включая тестирование химиочувствительности) и значительно увеличивает ценность биобанков.[141][150][151] Используя технологию CRC, исследователи смогли определить эффективную терапию для пациента с редким типом опухоли легкого.[152] Группа Энглемана[153] описывает фармакогеномную платформу, которая способствует быстрому открытию комбинаций лекарств, которые могут преодолеть устойчивость с помощью системы CRC. Кроме того, метод CRC позволяет проводить генетические манипуляции с эпителиальными клетками ex vivo и их последующую оценку in vivo у одного и того же хозяина. В то время как первоначальные исследования показали, что совместное культивирование эпителиальных клеток с клетками Swiss 3T3 J2 было необходимо для индукции CRC, при использовании планшетов для культивирования через лунки физический контакт между фидерами и эпителиальными клетками не требуется для индукции CRC и, что более важно, необходимо облучение питающих клеток для этой индукции. В соответствии с экспериментами трансвелл, кондиционированная среда индуцирует и поддерживает CRC, что сопровождается одновременным повышением активности клеточной теломеразы. Активность кондиционированной среды напрямую коррелирует с апоптозом питательных клеток, индуцированным радиацией. Таким образом, условное перепрограммирование эпителиальных клеток опосредуется комбинацией Y-27632 и растворимого фактора (факторов), высвобождаемого апоптотическими питающими клетками.[154]
Riegel et al.[155] демонстрируют, что мышиные ME клетки, выделенные из нормальных молочных желез или из опухолей молочной железы, индуцированных вирусом опухоли молочной железы мышей (MMTV) -Neu, можно культивировать неограниченное время как условно перепрограммированные клетки (CRC). Маркеры, связанные с предшественниками клеточной поверхности, быстро индуцируются в нормальных ME-CRC мыши по сравнению с ME-клетками. Однако экспрессия определенных субпопуляций предшественников молочной железы, таких как CD49f + ESA + CD44 +, значительно снижается на более поздних пассажах. Тем не менее, мышиные ME-CRCs, выращенные в трехмерном внеклеточном матриксе, дали начало ацинарным структурам молочных желез. ME-CRC, выделенные из опухолей молочной железы трансгенных мышей MMTV-Neu, экспрессируют высокие уровни HER2 / neu, а также маркеры опухолевых клеток, такие как CD44 +, CD49f + и ESA + (EpCam). Эти паттерны экспрессии поддерживаются в более поздних пассажах CRC. У ME-CRC раннего и позднего пассажа из опухолей MMTV-Neu, которые были имплантированы в жировые подушечки молочных желез сингенных или голых мышей, развивались сосудистые опухоли, которые метастазировали в течение 6 недель после трансплантации. Важно отметить, что гистопатология этих опухолей неотличима от гистопатологии родительских опухолей, которые развиваются у мышей MMTV-Neu. Application of the CRC system to mouse mammary epithelial cells provides an attractive model system to study the genetics and phenotype of normal and transformed mouse epithelium in a defined culture environment and in vivo transplant studies.
A different approach to CRC is to inhibit CD47 - а мембранный белок это тромбоспондин-1 рецептор. Loss of CD47 permits sustained proliferation of primary мышиный endothelial cells, increases asymmetric division and enables these cells to spontaneously reprogram to form multipotent эмбриоидное тело -like clusters. CD47 knockdown acutely increases мРНК levels of c-Myc and other stem cell transcription factors in cells in vitro and in vivo. Thrombospondin-1 is a key environmental signal that inhibits stem cell self-renewal via CD47. Thus, CD47 antagonists enable cell self-renewal and reprogramming by overcoming negative regulation of c-Myc and other stem cell transcription factors.[156] In vivo blockade of CD47 using an antisense морфолино increases survival of mice exposed to lethal total body irradiation due to increased proliferative capacity of bone marrow-derived cells and radioprotection of radiosensitive gastrointestinal tissues.[157]
Lineage-specific enhancers
Дифференцированный макрофаги can self-renew in tissues and expand long-term in culture.[27] Under certain conditions macrophages can divide without losing features they have acquired while specializing into иммунные клетки – which is usually not possible with дифференцированные клетки. The macrophages achieve this by activating a gene network similar to one found in embryonic stem cells. Single-cell analysis revealed that, in vivo, proliferating macrophages can derepress a macrophage-specific enhancer repertoire associated with a gene network controlling self-renewal. This happened when concentrations of two transcription factors named MafB и c-Maf were naturally low or were inhibited for a short time. Genetic manipulations that turned off MafB and c-Maf in the macrophages caused the cells to start a self-renewal program. The similar network also controls embryonic stem cell self-renewal but is associated with distinct embryonic stem cell-specific enhancers.[28]
Hence macrophages isolated from MafB- and c-Maf-double deficient mice divide indefinitely; the self-renewal depends on c-Myc и Klf4.[158]
Indirect lineage conversion
Indirect lineage conversion is a reprogramming methodology in which somatic cells transition through a plastic intermediate state of partially reprogrammed cells (pre-iPSC), induced by brief exposure to reprogramming factors, followed by differentiation in a specially developed chemical environment (artificial niche).[159]
This method could be both more efficient and safer, since it does not seem to produce tumors or other undesirable genetic changes and results in much greater yield than other methods. However, the safety of these cells remains questionable. Since lineage conversion from pre-iPSC relies on the use of iPSC reprogramming conditions, a fraction of the cells could acquire pluripotent properties if they do not stop the de-differentation process in vitro or due to further de-differentiation in vivo.[160]
Outer membrane glycoprotein
A common feature of pluripotent stem cells is the specific nature of protein гликозилирование of their outer membrane. That distinguishes them from most nonpluripotent cells, although not белые кровяные клетки.[161] В гликаны on the stem cell surface respond rapidly to alterations in cellular state and signaling and are therefore ideal for identifying even minor changes in cell populations. Много маркеры стволовых клеток are based on cell surface glycan epitopes including the widely used markers SSEA-3, SSEA-4, Tra 1-60 and Tra 1-81.[162] Suila Heli et al.[163] speculate that in human stem cells extracellular O-GlcNAc and extracellular O-LacNAc, play a crucial role in the fine tuning of Notch сигнальный путь - a highly conserved cell signaling system, that regulates cell fate specification, differentiation, left–right asymmetry, apoptosis, somitogenesis, angiogenesis and plays a key role in stem cell proliferation (reviewed by Perdigoto and Bardin[164] and Jafar-Nejad et al.[165])
Changes in outer membrane protein glycosylation are markers of cell states connected in some way with pluripotency and differentiation.[166] The glycosylation change is apparently not just the result of the initialization of gene expression, but perform as an important gene regulator involved in the acquisition and maintenance of the undifferentiated state.[167]
For example, activation of гликопротеин ACA,[168] linking glycosylphosphatidylinositol on the surface of the progenitor cells in human peripheral blood, induces increased expression of genes Wnt, Notch-1, BMI1 и HOXB4 through a signaling cascade PI3K /Акт /mTor /PTEN and promotes the formation of a self-renewing population of hematopoietic stem cells.[169]
Furthermore, dedifferentiation of progenitor cells induced by ACA-dependent signaling pathway leads to ACA-induced pluripotent stem cells, capable of differentiating in vitro into cells of all three ростковые отростки.[170]Изучение лектины ' ability to maintain a culture of pluripotent human stem cells has led to the discovery of lectin Erythrina crista-galli (ECA), which can serve as a simple and highly effective matrix for the cultivation of human pluripotent stem cells.[171]
Reprogramming with a proteoglycan
An alternative strategy to convert somatic cells to pluripotent states may be continuous stimulation of fibroblasts by a single ECM протеогликан, фибромодулин.[172] Such cells exhibit capability for skeletal muscle regeneration with markedly less tumorigenic risk when compared with iPSCs.[173] The decreased tumorigenicity of such cells is related to CDKN2B upregulation during the recombinant human fibromodulin reprogramming process[174]
Reprogramming through a physical approach
Белок клеточной адгезии E-кадгерин is indispensable for a robust pluripotent фенотип.[175] During reprogramming for iPS cell generation, N-кадгерин can replace function of E-cadherin.[176] These functions of cadherins are not directly related to adhesion because sphere morphology helps maintaining the "stemness" of stem cells.[177] Moreover, sphere formation, due to forced growth of cells on a low attachment surface, sometimes induces reprogramming. For example, neural progenitor cells can be generated from fibroblasts directly through a physical approach without introducing exogenous reprogramming factors.
Physical cues, in the form of parallel microgrooves on the surface of cell-adhesive substrates, can replace the effects of small-molecule epigenetic modifiers and significantly improve reprogramming efficiency. The mechanism relies on the mechanomodulation of the cells' epigenetic state. Specifically, "decreased histone deacetylase activity and upregulation of the expression of WD repeat domain 5 (WDR5) – a subunit of H3 methyltranferase – by microgrooved surfaces lead to increased histone H3 acetylation and methylation". Nanofibrous scaffolds with aligned fibre orientation produce effects similar to those produced by microgrooves, suggesting that changes in cell morphology may be responsible for modulation of the epigenetic state.[178]

Substrate rigidity is an important biophysical cue influencing neural induction and subtype specification. For example, soft substrates promote neuroepithelial conversion while inhibiting neural crest differentiation of hESCs in a BMP4 -зависимая манера. Mechanistic studies revealed a multi-targeted mechanotransductive process involving mechanosensitive Smad фосфорилирование and nucleocytoplasmic shuttling, regulated by rigidity-dependent Бегемот /YAP activities and actomyosin цитоскелет integrity and сократимость.[179]
Mouse embryonic stem cells (mESCs) undergo self-renewal in the presence of the цитокин фактор ингибирования лейкемии (LIF). Following LIF withdrawal, mESCs differentiate, accompanied by an increase in cell–substratum адгезия and cell spreading. Restricted cell spreading in the absence of LIF by either culturing mESCs on chemically defined, weakly adhesive biosubstrates, or by manipulating the цитоскелет allowed the cells to remain in an undifferentiated and pluripotent state. The effect of restricted cell spreading on mESC self-renewal is not mediated by increased intercellular adhesion, as inhibition of mESC adhesion using a function blocking anti E-cadherin antibody or миРНК does not promote differentiation.[180]Possible mechanisms of stem cell fate predetermination by physical interactions with the extracellular matrix have been described.[181][182]
A new method has been developed that turns cells into stem cells faster and more efficiently by 'squeezing' them using 3D microenvironment stiffness and density of the surrounding gel. The technique can be applied to a large number of cells to produce stem cells for medical purposes on an industrial scale.[183][184][185]
Cells involved in the reprogramming process change morphologically as the process proceeds. This results in physical difference in adhesive forces among cells. Substantial differences in 'adhesive signature' between pluripotent stem cells, partially reprogrammed cells, differentiated progeny and somatic cells allowed to develop separation process for isolation of pluripotent stem cells in микрофлюидные устройства,[186] который:
- fast (separation takes less than 10 minutes);
- efficient (separation results in a greater than 95 percent pure iPS cell culture);
- innocuous (cell survival rate is greater than 80 percent and the resulting cells retain normal transcriptional profiles, differentiation potential and karyotype).
Stem cells possess mechanical memory (they remember past physical signals) – with the Сигнальный путь бегемота factors:[187] Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding domain (TAZ) acting as an intracellular mechanical rheostat—that stores information from past physical environments and influences the cells' fate.[188][189]
Нервные стволовые клетки
Stroke and many neurodegenerative disorders such as Parkinson's disease, Alzheimer's disease, amyotrophic lateral sclerosis need cell replacement therapy. The successful use of converted neural cells (cNs) in transplantations open a new avenue to treat such diseases.[190] Nevertheless, induced neurons (iNs), directly converted from fibroblasts are terminally committed and exhibit very limited proliferative ability that may not provide enough autologous donor cells for transplantation.[191] Self-renewing induced neural stem cells (iNSCs) provide additional advantages over iNs for both basic research and clinical applications.[121][122][123][192][193]
For example, under specific growth conditions, mouse fibroblasts can be reprogrammed with a single factor, Sox2, to form iNSCs that self-renew in culture and after transplantation can survive and integrate without forming tumors in mouse brains.[194] INSCs can be derived from adult human fibroblasts by non-viral techniques, thus offering a safe method for autologous transplantation or for the development of cell-based disease models.[193]
Neural chemically induced progenitor cells (ciNPCs) can be generated from mouse tail-tip fibroblasts and human urinary somatic cells without introducing exogenous factors, but - by a chemical cocktail, namely VCR (V, VPA, inhibitor of HDACs; C, CHIR99021, an inhibitor of GSK-3 kinases and R, RepSox, ингибитор TGF beta signaling pathways ), under a physiological hypoxic condition.[195] Alternative cocktails with inhibitors of histone deacetylation, glycogen synthase kinase and TGF-β pathways (where: бутират натрия (NaB) or Трихостатин А (TSA) could replace VPA, Лития хлорид (LiCl) or lithium carbonate (Li2CO3) could substitute CHIR99021, or Repsox may be replaced with SB-431542 или же Траниласт ) show similar efficacies for ciNPC induction.[195]Zhang, et al.,[196] also report highly efficient reprogramming of mouse fibroblasts into induced neural stem cell-like cells (ciNSLCs) using a cocktail of nine components.
Multiple methods of direct transformation of somatic cells into induced neural stem cells have been described.[197]
Proof of principle experiments demonstrate that it is possible to convert transplanted human fibroblasts and human астроциты directly in the brain that are engineered to express inducible forms of neural reprogramming genes, into neurons, when reprogramming genes (Ascl1, Brn2a и Myt1l ) are activated after transplantation using a drug.[198]
Астроциты – the most common нейроглиальный brain cells, which contribute to шрам formation in response to injury – can be directly reprogrammed in vivo to become functional neurons that formed networks in mice without the need of cell transplantation.[199] The researchers followed the mice for nearly a year to look for signs of tumor formation and reported finding none. The same researchers have turned scar-forming astrocytes into progenitor cells called neuroblasts that regenerated into neurons in the injured adult spinal cord.[200]
Oligodendrocyte precursor cells
Без миелин to insulate neurons, nerve signals quickly lose power. Diseases that attack myelin, such as multiple sclerosis, result in nerve signals that cannot propagate to nerve endings and as a consequence lead to cognitive, motor and sensory problems. Transplantation of олигодендроцит precursor cells (OPCs), which can successfully create myelin sheaths around nerve cells, is a promising potential therapeutic response. Direct lineage conversion of mouse and rat fibroblasts into oligodendroglial cells provides a potential source of OPCs. Conversion by forced expression of both eight[201] or of the three[202] transcription factors Sox10, Olig2 and Zfp536, may provide such cells.
Cardiomyocytes
Cell-based in vivo therapies may provide a transformative approach to augment vascular and muscle growth and to prevent non-contractile scar formation by delivering transcription factors[118] or microRNAs[14] к сердцу.[203] Cardiac fibroblasts, which represent 50% of the cells in the mammalian heart, can be reprogrammed into кардиомиоцит -like cells in vivo by local delivery of cardiac core transcription factors ( GATA4, MEF2C, TBX5 and for improved reprogramming plus ESRRG, MESP1, Myocardin and ZFPM2) after coronary перевязка.[118][204] These results implicated therapies that can directly remuscularize the heart without cell transplantation. However, the efficiency of such reprogramming turned out to be very low and the phenotype of received cardiomyocyte-like cells does not resemble those of a mature normal cardiomyocyte. Furthermore, transplantation of cardiac transcription factors into injured murine hearts resulted in poor cell survival and minimal expression of cardiac genes.[205]
Meanwhile, advances in the methods of obtaining cardiac myocytes in vitro occurred.[206][207] Efficient cardiac differentiation of human iPS cells gave rise to progenitors that were retained within infarcted rat hearts and reduced remodeling of the heart after ischemic damage.[208]
The team of scientists, who were led by Sheng Ding, used a cocktail of nine chemicals (9C) for transdifferentiation of human skin cells into beating heart cells. With this method, more than 97% of the cells began beating, a characteristic of fully developed, healthy heart cells. The chemically induced cardiomyocyte-like cells (ciCMs) uniformly contracted and resembled human cardiomyocytes in their transcriptome, epigenetic, and electrophysiological properties. When transplanted into infarcted mouse hearts, 9C-treated fibroblasts were efficiently converted to ciCMs and developed into healthy-looking heart muscle cells within the organ.[209] This chemical reprogramming approach, after further optimization, may offer an easy way to provide the cues that induce heart muscle to regenerate locally.[210]
В другом исследовании ischemic cardiomyopathy in the murine infarction model was targeted by iPS cell transplantation. It synchronized failing ventricles, offering a regenerative strategy to achieve resynchronization and protection from декомпенсация by dint of improved left ventricular conduction and contractility, reduced scarring and reversal of structural remodelling.[211]One protocol generated populations of up to 98% cardiomyocytes from hPSCs simply by modulating the canonical Сигнальный путь Wnt at defined time points in during differentiation, using readily accessible small molecule compounds.[212]
Discovery of the mechanisms controlling the formation of cardiomyocytes led to the development of the drug ITD-1, which effectively clears the cell surface from TGF-β receptor type II and selectively inhibits intracellular TGF-β signaling. It thus selectively enhances the differentiation of uncommitted мезодерма to cardiomyocytes, but not to vascular smooth muscle and endothelial cells.[213]
One project seeded decellularized mouse hearts with human iPSC-derived multipotential cardiovascular progenitor cells. The introduced cells migrated, proliferated and differentiated in situ into cardiomyocytes, smooth muscle cells and endothelial cells to reconstruct the hearts. In addition, the heart's extracellular matrix (the substrate of heart scaffold) signalled the human cells into becoming the specialised cells needed for proper heart function. After 20 days of perfusion with growth factors, the engineered heart tissues started to beat again and were responsive to drugs.[214]
Reprogramming of cardiac fibroblasts into induced cardiomyocyte-like cells (iCMs) на месте represents a promising strategy for cardiac regeneration. Mice exposed in vivo, to three cardiac transcription factors GMT (Gata4, Mef2c, Tbx5) and the small-molecules: SB-431542 (the transforming growth factor (TGF)-β inhibitor), and XAV939 (the WNT inhibitor) for 2 weeks after myocardial infarction showed significantly improved reprogramming (reprogramming efficiency increased eight-fold) and cardiac function compared to those exposed to only GMT.[215]
See also: review[216]
Rejuvenation of the muscle stem cell
The elderly often suffer from progressive мышечная слабость and regenerative failure owing in part to elevated activity of the p38α and p38β mitogen-activated kinase pathway in senescent skeletal muscle stem cells. Subjecting such stem cells to transient inhibition of p38α and p38β in conjunction with culture on soft гидрогель substrates rapidly expands and rejuvenates them that result in the return of their strength.[217]
In geriatric mice, resting satellite cells lose reversible quiescence by switching to an irreversible pre-senescence state, caused by derepression of p16 INK4a (also called Cdkn2a). On injury, these cells fail to activate and expand, even in a youthful environment. p16INK4a silencing in geriatric satellite cells restores quiescence and muscle regenerative functions.[218]
Myogenic progenitors for potential use in disease modeling or cell-based therapies targeting skeletal muscle could also be generated directly from induced pluripotent stem cells using free-floating spherical culture (EZ spheres) in a culture medium supplemented with high concentrations (100 ng/ml) of fibroblast growth factor-2 (FGF-2 ) и фактор роста эпидермиса.[219]
Гепатоциты
Unlike current protocols for deriving гепатоциты from human fibroblasts, Saiyong Zhu et al., (2014)[220] did not generate iPSCs but, using small molecules, cut short reprogramming to pluripotency to generate an induced multipotent progenitor cell (iMPC) state from which endoderm progenitor cells and subsequently hepatocytes (iMPC-Heps) were efficiently differentiated. After transplantation into an immune-deficient mouse model of human liver failure, iMPC-Heps proliferated extensively and acquired levels of hepatocyte function similar to those of human primary adult hepatocytes. iMPC-Heps did not form tumours, most probably because they never entered a pluripotent state.

These results establish the feasibility of significant liver repopulation of mice with human hepatocytes generated in vitro, which removes a long-standing roadblock on the path to autologous liver cell therapy.
Cocktail of small molecules, Y-27632, A-83-01 (a TGFβ kinase/activin receptor like kinase (ALK5 ) inhibitor), and CHIR99021 (potent inhibitor of ГСК-3 ), can convert rat and mouse mature hepatocytes in vitro into proliferative bipotent cells – CLiPs (chemically induced liver progenitors). CLiPs can differentiate into both mature hepatocytes and biliary epithelial cells that can form functional ductal structures. In long-term culture CLiPs did not lose their proliferative capacity and their hepatic differentiation ability, and can repopulate chronically injured liver tissue.[221]
Insulin-producing cells
Осложнения Сахарный диабет Такие как сердечно-сосудистые заболевания, retinopathy, невропатия, нефропатия and peripheral circulatory diseases depend on sugar dysregulation due to lack of инсулин from pancreatic бета-клетки and can be lethal if they are not treated. One of the promising approaches to understand and cure diabetes is to use pluripotent stem cells (PSCs), including embryonic stem cells (ESCs) and induced PCSs (iPSCs).[222] Unfortunately, human PSC-derived insulin-expressing cells resemble human fetal β cells rather than adult β cells. In contrast to adult β cells, fetal β cells seem functionally immature, as indicated by increased базальный глюкоза secretion and lack of glucose stimulation and confirmed by РНК-последовательность чьи стенограммы.[223]
An alternative strategy is the conversion of fibroblasts towards distinct endodermal progenitor cell populations and, using cocktails of signalling factors, successful differentiation of these endodermal progenitor cells into functional beta-like cells both in vitro and in vivo.[224]
Overexpression of the three факторы транскрипции, PDX1 (required for pancreatic bud outgrowth and beta-cell maturation), NGN3 (required for endocrine precursor cell formation) and MAFA (for beta-cell maturation) combination (called PNM) can lead to the transformation of some cell types into a beta cell-like state.[225] An accessible and abundant source of functional insulin-producing cells is кишечник. PMN expression in human intestinal "органоиды " stimulates the conversion of intestinal epithelial cells into β-like cells possibly acceptable for трансплантация.[226]
Nephron Progenitors
Adult proximal tubule cells were directly transcriptionally reprogrammed to нефрон progenitors of the embryonic почка, using a pool of six genes of instructive transcription factors (SIX1, SIX2, OSR1, Eyes absent homolog 1(EYA1), Homeobox A11 (HOXA11) and Snail homolog 2 (SNAI2)) that activated genes consistent with a cap мезенхима /nephron progenitor phenotype in the adult proximal tubule cell line.[227]The generation of such cells may lead to cellular therapies for adult почечная болезнь. Embryonic kidney organoids placed into adult rat kidneys can undergo onward development and vascular development.[228]
Blood vessel cells
As blood vessels age, they often become abnormal in structure and function, thereby contributing to numerous age-associated diseases including myocardial infarction, ischemic stroke and atherosclerosis of arteries supplying the heart, brain and lower extremities. So, an important goal is to stimulate vascular growth for the collateral circulation to prevent the exacerbation of these diseases. Induced Vascular Progenitor Cells (iVPCs) are useful for cell-based therapy designed to stimulate coronary collateral growth. They were generated by partially reprogramming endothelial cells.[159] The vascular commitment of iVPCs is related to the epigenetic memory of endothelial cells, which engenders them as cellular components of growing blood vessels. That is why, when iVPCs were implanted into миокард, they engrafted in blood vessels and increased coronary collateral flow better than iPSCs, mesenchymal stem cells, or native endothelial cells.[229]
Ex vivo genetic modification can be an effective strategy to enhance stem cell function. For example, cellular therapy employing genetic modification with Pim-1 kinase (a downstream effector of Акт, which positively regulates neovasculogenesis) of Костный мозг –derived cells[230] or human cardiac progenitor cells, isolated from failing myocardium[231] results in durability of repair, together with the improvement of functional parameters of myocardial hemodynamic performance.
Stem cells extracted from fat tissue after липосакция may be coaxed into becoming progenitor гладкая мышца cells (iPVSMCs) found in arteries and veins.[232]
The 2D culture system of human iPS cells[233] in conjunction with triple marker selection (CD34 (a surface glycophosphoprotein expressed on developmentally early embryonic fibroblasts), NP1 (receptor – neuropilin 1) and KDR (kinase insert domain-containing receptor)) for the isolation of vasculogenic precursor cells from human iPSC, generated endothelial cells that, after transplantation, formed stable, functional mouse blood vessels in vivo, lasting for 280 days.[234]
To treat infarction, it is important to prevent the formation of fibrotic scar tissue. This can be achieved in vivo by transient application of паракринный factors that redirect native heart progenitor stem cell contributions from scar tissue to cardiovascular tissue. For example, in a mouse myocardial infarction model, a single intramyocardial injection of human фактор роста эндотелия сосудов А mRNA (VEGF-A modRNA), modified to escape the body's normal defense system, results in long-term improvement of heart function due to mobilization and redirection of epicardial progenitor cells toward cardiovascular cell types.[235]
Blood stem cells
красные кровяные тельца
РБК переливание is necessary for many patients. However, to date the supply of RBCs remains labile. In addition, transfusion risks infectious disease transmission. A large supply of safe RBCs generated in vitro would help to address this issue. Ex vivo erythroid cell generation may provide alternative transfusion products to meet present and future clinical requirements.[236][237] Red blood cells (RBC)s generated in vitro from mobilized CD34 positive cells have normal survival when transfused into an autologous recipient.[238] RBC produced in vitro contained exclusively гемоглобин плода (HbF), which rescues the functionality of these RBCs. In vivo the switch of fetal to adult hemoglobin was observed after infusion of nucleated erythroid precursors derived from iPSCs.[239] Although RBCs do not have nuclei and therefore can not form a tumor, their immediate erythroblasts precursors have nuclei. The terminal maturation of erythroblasts into functional RBCs requires a complex remodeling process that ends with extrusion of the nucleus and the formation of an enucleated RBC.[240] Cell reprogramming often disrupts enucleation. Transfusion of in vitro-generated RBCs or erythroblasts does not sufficiently protect against tumor formation.
В арил hydrocarbon receptor (AhR) pathway (which has been shown to be involved in the promotion of cancer cell development) plays an important role in normal blood cell development. AhR activation in human hematopoietic progenitor cells (HPs) drives an unprecedented expansion of HPs, megakaryocyte- and erythroid-lineage cells.[241] See also Concise Review:[242][243]В SH2B3 gene encodes a negative regulator of cytokine signaling and naturally occurring loss-of-function variants in this gene increase RBC counts in vivo. Targeted suppression of SH2B3 in primary human hematopoietic stem and progenitor cells enhanced the maturation and overall yield of in-vitro-derived RBCs. Moreover, inactivation of SH2B3 by CRISPR /Cas9 genome editing in human pluripotent stem cells allowed enhanced erythroid cell expansion with preserved differentiation.[244](See also overview.[243][245])

Тромбоциты
Тромбоциты help prevent hemorrhage in thrombocytopenic patients and patients with thrombocythemia. A significant problem for multitransfused patients is refractoriness to platelet transfusions. Thus, the ability to generate platelet products ex vivo and platelet products lacking HLA antigens in serum-free media would have clinical value.An РНК-интерференция -based mechanism used a лентивирусный вектор to express short-hairpin RNAi targeting β2-microglobulin transcripts in CD34-positive cells. Generated platelets demonstrated an 85% reduction in class I HLA antigens. These platelets appeared to have normal function in vitro[246][247]
One clinically-applicable strategy for the derivation of functional platelets from human iPSC involves the establishment of stable immortalized megakaryocyte progenitor cell lines (imMKCLs) through доксициклин -dependent overexpression of BMI1 и BCL-XL. The resulting imMKCLs can be expanded in culture over extended periods (4–5 months), even after криоконсервация. Halting the overexpression (by the removal of doxycycline from the medium) of c-MYC, BMI1 и BCL-XL in growing imMKCLs led to the production of CD42b + platelets with functionality comparable to that of native platelets on the basis of a range of assays in vitro and in vivo.[248]Thomas et al., describe a forward programming strategy relying on the concurrent exogenous expression of 3 transcription factors: GATA1, FLI1 и TAL1. The forward programmed мегакариоциты proliferate and differentiate in culture for several months with megakaryocyte purity over 90% reaching up to 2x105 mature megakaryocytes per input hPSC. Functional platelets are generated throughout the culture allowing the prospective collection of several transfusion units from as few as one million starting hPSCs.[249]See also overview[250]
Immune cells
A specialised type of лейкоцит, известный как cytotoxic T лимфоциты (CTLs), are produced by the иммунная система and are able to recognise specific markers on the surface of various infectious or tumour cells, causing them to launch an attack to kill the harmful cells. Thence, immunotherapy with functional antigen-specific T cells has potential as a therapeutic strategy for combating many cancers and viral infections.[251] However, cell sources are limited, because they are produced in small numbers naturally and have a short lifespan.
A potentially efficient approach for generating antigen-specific CTLs is to revert mature immune T cells into iPSCs, which possess indefinite proliferative capacity in vitro and after their multiplication to coax them to redifferentiate back into T cells.[252][253][254]
Another method combines iPSC and рецептор химерного антигена (МАШИНА)[255] technologies to generate human T cells targeted to CD19, an antigen expressed by malignant В-клетки, in tissue culture.[256] This approach of generating therapeutic human T cells may be useful for cancer immunotherapy and other medical applications.
Invariant natural killer T (iNKT) cells have great clinical potential as адъюванты for cancer immunotherapy. iNKT cells act as innate T lymphocytes and serve as a bridge between the врожденный и acquired immune systems. They augment anti-tumor responses by producing интерферон-гамма (IFN-γ).[257] The approach of collection, reprogramming/dedifferentiation, re-differentiation and injection has been proposed for related tumor treatment.[258]
Дендритные клетки (DC) are specialized to control T-cell responses. DC with appropriate genetic modifications may survive long enough to stimulate antigen-specific CTL and after that be completely eliminated. DC-like antigen-presenting cells obtained from human induced pluripotent stem cells can serve as a source for вакцинация терапия.[259]
CCAAT/enhancer binding protein-α (C/EBPα) induces transdifferentiation of В-клетки в макрофаги at high efficiencies[260] and enhances reprogramming into iPS cells when co-expressed with transcription factors Oct4, Sox2, Klf4 and Myc.[261] with a 100-fold increase in iPS cell reprogramming efficiency, involving 95% of the population.[262]Furthermore, C/EBPa can convert selected human B cell lymphoma and leukemia cell lines into macrophage-like cells at high efficiencies, impairing the cells' tumor-forming capacity.[263]
Thymic epithelial cells rejuvenation
В вилочковая железа is the first organ to deteriorate as people age. This shrinking is one of the main reasons the immune system becomes less effective with age. Diminished expression of the thymic epithelial cell фактор транскрипции FOXN1 has been implicated as a component of the mechanism regulating age-related involution.[264][265]
Клэр Блэкберн and colleagues show that established age-related thymic involution can be reversed by forced upregulation of just one transcription factor – FOXN1 in the thymic epithelial cells in order to promote омоложение, proliferation and differentiation of these cells into fully functional thymic epithelium.[266]This rejuvenation and increased proliferation was accompanied by upregulation of genes that promote клеточный цикл прогрессия (циклин D1, ΔNстр. 63, FgfR2IIIb ) and that are required in the thymic epithelial cells to promote specific aspects of Т-клетка development (Dll4, Kitl, Ccl25, Cxcl12, Cd40, Cd80, Ctsl, Pax1 ). In the future, this method may be widely used to enhance immune function and combat Inflammaging in patients by rejuvenating the thymus на месте.[267]
Мезенхимальные стволовые клетки
Индукция
Mesenchymal stem/stromal cells (MSCs) are under investigation for applications in cardiac, renal, neural, joint and bone repair, as well as in inflammatory conditions and hemopoietic cotransplantation.[268] This is because of their immunosuppressive properties and ability to differentiate into a wide range of mesenchymal-lineage tissues. MSCs are typically harvested from adult bone marrow or fat, but these require painful invasive procedures and are low-frequency sources, making up only 0.001–0.01% of bone marrow cells and 0.05% in liposuction aspirates.[269] Of concern for autologous use, in particular in the elderly most in need of tissue repair, MSCs decline in quantity and quality with age.[268][270][271]
IPSCs could be obtained by the cells rejuvenation of even centenarians.[9][41] Because iPSCs can be harvested free of ethical constraints and culture can be expanded indefinitely, they are an advantageous source of MSCs.[272] IPSC treatment with SB-431542 leads to rapid and uniform MSC generation from human iPSCs. (SB-431542 is an inhibitor of activin/TGF- pathways by blocking фосфорилирование из ALK4, ALK5 и ALK7 receptors.) These iPS-MSCs may lack teratoma-forming ability, display a normal stable karyotype in culture and exhibit growth and differentiation characteristics that closely resemble those of primary MSCs. It has potential for in vitro scale-up, enabling MSC-based therapies.[273] MSC derived from iPSC have the capacity to aid periodontal regeneration and are a promising source of readily accessible stem cells for use in the clinical treatment of periodontitis.[274][275]
Lai et al., & Lu report the chemical method to generate MSC-like cells (iMSCs), from human primary dermal fibroblasts using six chemical inhibitors (SP600125, SB202190, Go6983, Y-27632, PD0325901, and CHIR99021) with or without 3 growth factors (transforming growth factor-β (TGF-β), basic fibroblast growth factor (bFGF), and leukemia inhibitory factor (LIF)). The chemical cocktail directly converts human fibroblasts to iMSCs with a monolayer culture in 6 days, and the conversion rate was approximately 38%.[276]
Besides cell therapy in vivo, the culture of human mesenchymal stem cells can be used in vitro for mass-production of экзосомы, which are ideal vehicles for drug delivery.[277]
Dedifferentiated adipocytes
Жировой tissue, because of its abundance and relatively less invasive harvest methods, represents a source of mesenchymal stem cells (MSCs). Unfortunately, liposuction aspirates are only 0.05% MSCs.[269] However, a large amount of mature adipocytes, which in general have lost their proliferative abilities and therefore are typically discarded, can be easily isolated from the adipose cell suspension and dedifferentiated into липид -free fibroblast-like cells, named dedifferentiated fat (DFAT) cells. DFAT cells re-establish active proliferation ability and express multipotent capacities.[278] Compared with adult stem cells, DFAT cells show unique advantages in abundance, isolation and homogeneity. Under proper induction culture in vitro or proper environment in vivo, DFAT cells could demonstrate adipogenic, osteogenic, chondrogenic and myogenic potentials. They could also exhibit perivascular characteristics and elicit neovascularization.[279][280][281]
Chondrogenic cells
Хрящ is the connective tissue responsible for frictionless joint movement. Its degeneration ultimately results in complete loss of joint function in the late stages of остеоартроз. As an avascular and hypocellular tissue, cartilage has a limited capacity for self-repair. Хондроциты are the only cell type in cartilage, in which they are surrounded by the extracellular matrix that they secrete and assemble.
One method of producing cartilage is to induce it from iPS cells.[282] Alternatively, it is possible to convert fibroblasts directly into induced chondrogenic cells (iChon) without an intermediate iPS cell stage, by inserting three reprogramming factors (c-MYC, KLF4 and SOX9).[283] Human iChon cells expressed marker genes for chondrocytes (type II collagen) but not fibroblasts.
Имплантированные в дефекты суставного хряща крыс, человеческие клетки iChon выживали, образуя хрящевую ткань, по крайней мере, в течение четырех недель без опухолей. В этом методе используется c-MYC, который, как считается, играет важную роль в онкогенезе и включает ретровирус ввести факторы репрограммирования, исключив его из неизменного использования в терапии человека.[252][254][284]
Источники клеток для перепрограммирования
Наиболее часто используемые источники для перепрограммирования - клетки крови.[285][286][287][288][289] и фибробласты, полученные при биопсии кожи,[290] но взяв клетки из моча менее инвазивен.[291][292][293][294] Последний метод не требует биопсии или забора крови. По состоянию на 2013 год стволовые клетки, полученные из мочи, были дифференцированы на эндотелиальные, остеогенные, хондрогенные, адипогенные, скелетные миогенные и нейрогенные линии без образования тератом.[295] Следовательно, их эпигенетическая память приспособлена для репрограммирования в iPS-клетки. Однако в моче появляется небольшое количество клеток, была достигнута только низкая эффективность преобразования и риск бактериального заражения относительно высок.
Еще одним многообещающим источником клеток для репрограммирования являются мезенхимальные стволовые клетки, полученные из волосяных фолликулов человека.[296]
Происхождение соматических клеток, используемых для репрограммирования, может влиять на эффективность репрограммирования.[297][298] функциональные свойства полученных индуцированных стволовых клеток[299] и способность образовывать опухоли.[300][301]
IPSC сохраняют эпигенетическую память о своей ткани происхождения, что влияет на их способность к дифференцировке.[284][299][302][303][304][305]Эта эпигенетическая память не обязательно проявляется на стадии плюрипотентности - ИПСК, полученные из разных тканей, обладают правильной морфологией, экспрессируют маркеры плюрипотентности и способны дифференцироваться на три эмбриональных слоя in vitro и in vivo. Однако эта эпигенетическая память может проявляться во время повторной дифференцировки в определенные типы клеток, которые требуют определенных локусов, несущих остаточные эпигенетические метки.
Смотрите также
- Трансдифференцировка
- Примеры трансдифференцировки in vitro с помощью подхода, учитывающего происхождение
- Примеры трансдифференцировки in vitro с использованием подхода начальной фазы эпигенетической активации
- Примеры трансдифференцировки in vivo с помощью подхода, учитывающего происхождение
- Ниши стволовых клеток, вызванные травмами
- Факторы транскрипции
- Факторы роста
- Факторы первопроходца
- Клеточная дифференциация
- CAF-1
Рекомендации
- ^ Yamanaka, S .; Блау, Х. М. (2010). «Ядерное перепрограммирование до плюрипотентного состояния тремя подходами». Природа. 465 (7299): 704–712. Bibcode:2010Натура.465..704л. Дои:10.1038 / природа09229. ЧВК 2901154. PMID 20535199.
- ^ Гурдон Дж. Б. и Ян Уилмут (2011) Перенос ядер в яйца и ооциты Cold Spring Harb Perspect Biol; 3: a002659
- ^ а б c d Tachibana, M .; Amato, P .; Спарман, М .; Gutierrez, N.M .; Типпнер-Хеджес, Р.; Ma, H .; Kang, E .; Fulati, A .; Lee, H. S .; Sritanaudomchai, H .; Мастерсон, К .; Larson, J .; Eaton, D .; Sadler-Fredd, K .; Battaglia, D .; Ли, Д .; Wu, D .; Jensen, J .; Patton, P .; Gokhale, S .; Stouffer, R.L .; Wolf, D .; Миталипов, С. (2013). «Стволовые эмбриональные клетки человека, полученные путем переноса ядра соматической клетки». Клетка. 153 (6): 1228–1238. Дои:10.1016 / j.cell.2013.05.006. ЧВК 3772789. PMID 23683578.
- ^ Noggle, S .; Fung, H.L .; Гор, А .; Martinez, H .; Satriani, K. C .; Проссер, Р .; Oum, K .; Paull, D .; Druckenmiller, S .; Freeby, M .; Greenberg, E .; Zhang, K .; Goland, R .; Sauer, M. V .; Leibel, R.L .; Эгли, Д. (2011). «Человеческие ооциты перепрограммируют соматические клетки до плюрипотентного состояния». Природа. 478 (7367): 70–75. Bibcode:2011Натура 478 ... 70N. Дои:10.1038 / природа10397. PMID 21979046.
- ^ а б Pan, G .; Wang, T .; Yao, H .; Пей, Д. (2012). «Репрограммирование соматических клеток для регенеративной медицины: SCNT против IPS клеток». BioEssays. 34 (6): 472–476. Дои:10.1002 / bies.201100174. PMID 22419173.
- ^ Do, J. T .; Han, D. W .; Gentile, L; Собек-Клоке, я; Stehling, M; Lee, H.T .; Шелер, Х. Р. (2007). «Стирание клеточной памяти путем слияния с плюрипотентными клетками». Стволовые клетки. 25 (4): 1013–1020. Дои:10.1634 / стволовые клетки.2006-0691. PMID 17218392.
- ^ а б Takahashi, K .; Tanabe, K .; Охнуки, М .; Нарита, М .; Ichisaka, T .; Tomoda, K .; Яманака, С. (2007). «Индукция плюрипотентных стволовых клеток из фибробластов взрослого человека с помощью определенных факторов». Клетка. 131 (5): 861–872. Дои:10.1016 / j.cell.2007.11.019. HDL:2433/49782. PMID 18035408.
- ^ Wang, W .; Yang, J .; Liu, H .; Лу, Д .; Чен, X .; Зенонос, З .; Campos, L. S .; Rad, R .; Guo, G .; Zhang, S .; Брэдли, А .; Лю П. (2011). «Быстрое и эффективное перепрограммирование соматических клеток в индуцированные плюрипотентные стволовые клетки рецептором ретиноевой кислоты гамма и гомологом рецептора печени 1». Труды Национальной академии наук. 108 (45): 18283–18288. Bibcode:2011PNAS..10818283W. Дои:10.1073 / pnas.1100893108. ЧВК 3215025. PMID 21990348.
- ^ а б c Лапассет, Л .; Milhavet, O .; Prieur, A .; Besnard, E .; Babled, A .; Ait-Hamou, N .; Leschik, J .; Pellestor, F .; Ramirez, J. -M .; De Vos, J .; Lehmann, S .; Леметр, Ж. -М. (2011). «Омоложение стареющих и столетних человеческих клеток путем перепрограммирования через плюрипотентное состояние». Гены и развитие. 25 (21): 2248–2253. Дои:10.1101 / gad.173922.111. ЧВК 3219229. PMID 22056670.
- ^ Чжоу, H .; Wu, S .; Joo, J. Y .; Zhu, S .; Han, D. W .; Lin, T .; Траугер, С .; Bien, G .; Yao, S .; Zhu, Y .; Сюздак, Г.; Schöler, H.R .; Duan, L .; Дин, С. (2009). «Создание индуцированных плюрипотентных стволовых клеток с использованием рекомбинантных белков». Стволовая клетка. 4 (5): 381–384. Дои:10.1016 / j.stem.2009.04.005. PMID 19398399.
- ^ Ли, З .; Рана, Т. М. (2012). Использование микроРНК для увеличения образования индуцированных плюрипотентных стволовых клеток. Текущие протоколы в биологии стволовых клеток. 20. С. 4D.4.1–14. Дои:10.1002 / 9780470151808.sc04a04s20. ISBN 978-0470151808. PMID 22415842.
- ^ Anokye-Danso, F .; Триведи, К. М .; Juhr, D .; Gupta, M .; Cui, Z .; Tian, Y .; Zhang, Y .; Yang, W .; Gruber, P.J .; Эпштейн, Дж. А .; Морриси, Э. Э. (2011). «Высокоэффективное miRNA-опосредованное репрограммирование соматических клеток мыши и человека до плюрипотентности». Стволовая клетка. 8 (4): 376–388. Дои:10.1016 / j.stem.2011.03.001. ЧВК 3090650. PMID 21474102.
- ^ Miyoshi, N .; Ishii, H .; Nagano, H .; Haraguchi, N .; Dewi, D. L .; Kano, Y .; Nishikawa, S .; Tanemura, M .; Mimori, K .; Tanaka, F .; Сайто, Т .; Nishimura, J .; Takemasa, I .; Mizushima, T .; Ikeda, M .; Yamamoto, H .; Секимото, М .; Доки, Й .; Мори, М. (2011). «Перепрограммирование клеток мыши и человека до плюрипотентности с использованием зрелых микроРНК». Стволовая клетка. 8 (6): 633–638. Дои:10.1016 / j.stem.2011.05.001. PMID 21620789.
- ^ а б Jayawardena, T. M .; Егемназаров, Б .; Финч, Э. А .; Zhang, L .; Payne, J. A .; Пандья, К .; Zhang, Z .; Rosenberg, P .; Мироцу, М .; Дзау, В. Дж. (2012). «МикроРНК-опосредованное прямое перепрограммирование in vitro и in vivo сердечных фибробластов в кардиомиоциты». Циркуляционные исследования. 110 (11): 1465–1473. Дои:10.1161 / CIRCRESAHA.112.269035. ЧВК 3380624. PMID 22539765.
- ^ Бао, X .; Чжу, X .; Liao, B .; Бенда, С .; Zhuang, Q .; Pei, D .; Цинь, Б .; Эстебан, М.А. (2013). «МикроРНК в репрограммировании соматических клеток». Текущее мнение в области клеточной биологии. 25 (2): 208–214. Дои:10.1016 / j.ceb.2012.12.004. PMID 23332905.
- ^ Yoshioka, N .; Gros, E .; Li, H.R .; Kumar, S .; Дьякон, Д. С .; Maron, C .; Муотри, А. Р .; Chi, N.C .; Fu, X. D .; Ю. Б. Д .; Дауди, С. Ф. (2013). «Эффективное создание ИПСК человека с помощью синтетической саморепликативной РНК». Стволовая клетка. 13 (2): 246–254. Дои:10.1016 / j.stem.2013.06.001. ЧВК 3845961. PMID 23910086.
- ^ а б Hou, P .; Li, Y .; Чжан, X .; Liu, C .; Guan, J .; Li, H .; Чжао, Т .; Ye, J .; Yang, W .; Лю, К .; Ge, J .; Xu, J .; Zhang, Q .; Zhao, Y .; Дэн, Х. (2013). «Плюрипотентные стволовые клетки, индуцированные из соматических клеток мышей с помощью низкомолекулярных соединений». Наука. 341 (6146): 651–654. Bibcode:2013Наука ... 341..651H. Дои:10.1126 / science.1239278. PMID 23868920.
Efe, J. A .; Дин, С. (2011). «Развитие биологии малых молекул: управление судьбой и идентичностью клеток». Философские труды Королевского общества B: биологические науки. 366 (1575): 2208–2221. Дои:10.1098 / rstb.2011.0006. ЧВК 3130415. PMID 21727126. - ^ а б Stadtfeld, M .; Apostolou, E .; Ferrari, F .; Choi, J .; Walsh, R.M .; Chen, T .; Ooi, S. S. K .; Kim, S. Y .; Bestor, T. H .; Shioda, T .; Парк, П. Дж .; Хохедлингер, К. (2012). «Аскорбиновая кислота предотвращает потерю импринтинга Dlk1-Dio3 и способствует образованию мышей, полностью дифференцированных по iPS, из терминально дифференцированных В-клеток». Природа Генетика. 44 (4): 398–405, S1–2. Дои:10.1038 / ng.1110. ЧВК 3538378. PMID 22387999.
- ^ Пандиан, Г. Н .; Сугияма, Х. (2012). «Программируемые генетические переключатели для управления транскрипционной машиной плюрипотентности». Биотехнологический журнал. 7 (6): 798–809. Дои:10.1002 / biot.201100361. PMID 22588775.
Пандиан, Г. Н .; Nakano, Y .; Sato, S .; Morinaga, H .; Bando, T .; Nagase, H .; Сугияма, Х. (2012). «Синтетическая малая молекула для быстрой индукции множественных генов плюрипотентности в эмбриональных фибробластах мыши». Научные отчеты. 2: 544. Bibcode:2012НатСР ... 2Е.544П. Дои:10.1038 / srep00544. ЧВК 3408130. PMID 22848790. - ^ Де Робертис, Эдвард М. (2006). "Spemannорганизатор и саморегуляция у эмбрионов амфибий ». Обзоры природы Молекулярная клеточная биология. 7 (4): 296–302. Дои:10.1038 / nrm1855. ЧВК 2464568. PMID 16482093.
- ^ Слэк, Дж. М. У. (2009). «Метаплазия и репрограммирование соматических клеток». Журнал патологии. 217 (2): 161–8. Дои:10.1002 / путь.2442. PMID 18855879.
- ^ Wei, G .; Schubiger, G .; Сложнее, F .; Мюллер, А. М. (2000). «Пластичность стволовых клеток у млекопитающих и трансдетерминация у Дрозофила: Общие темы? ". Стволовые клетки. 18 (6): 409–14. Дои:10.1634 / стволовые клетки.18-6-409. PMID 11072028.
Worley, M. I .; Setiawan, L .; Харихаран, И. К. (2012). «Регенерация и трансдетерминация в Дрозофила Воображаемый Диски ». Ежегодный обзор генетики. 46: 289–310. Дои:10.1146 / annurev-genet-110711-155637. PMID 22934642. - ^ Сюй, Пэн-Фэй; Хуссен, Натали; Ферри-Лагно, Карин Ф .; Thisse, Бернар; Thisse, Кристина (2014). «Конструирование эмбриона позвоночного из двух противоположных градиентов морфогенов». Наука. 344 (6179): 87–89. Bibcode:2014Наука ... 344 ... 87X. Дои:10.1126 / science.1248252. PMID 24700857.
- ^ Штанге, Д. Э .; Koo, B.K .; Huch, M .; Сиббел, Г .; Basak, O .; Любимова А .; Kujala, P .; Bartfeld, S .; Koster, J .; Geahlen, J. H .; Peters, P.J .; Van Es, J. H .; Ван Де Ветеринг, М .; Mills, J.C .; Клеверс, Х. (2013). «Дифференцированные клетки Troy + Chief действуют как резервные стволовые клетки для создания всех линий эпителия желудка». Клетка. 155 (2): 357–68. Дои:10.1016 / j.cell.2013.09.008. ЧВК 4094146. PMID 24120136.
- ^ Tata, P. R .; Mou, H .; Пардо-Саганта, А .; Zhao, R .; Прабху, М .; Закон, Б. М .; Винарский, В .; Cho, J. L .; Бретон, С .; Sahay, A .; Medoff, B.D .; Раджагопал, Дж. (2013). «Дедифференцировка коммитированных эпителиальных клеток в стволовые клетки in vivo». Природа. 503 (7475): 218–23. Bibcode:2013Натура.503..218Т. Дои:10.1038 / природа12777. ЧВК 4035230. PMID 24196716.
- ^ Кусаба, Т .; Lalli, M .; Kramann, R .; Кобаяши, А .; Хамфрис, Б. Д. (2013). «Дифференцированные эпителиальные клетки почек восстанавливают поврежденный проксимальный каналец». Труды Национальной академии наук. 111 (4): 1527–1532. Дои:10.1073 / pnas.1310653110. ЧВК 3910580. PMID 24127583.
- ^ а б Сивеке, Майкл Х .; Аллен, Джудит Э. (2013). «За пределами стволовых клеток: самообновление дифференцированных макрофагов». Наука. 342 (6161): 1242974. Дои:10.1126 / science.1242974. PMID 24264994.
- ^ а б Soucie, E.L .; Weng, Z .; Geirsdóttir, L .; и другие. (2016). «Клоноспецифические энхансеры активируют гены самообновления в макрофагах и эмбриональных стволовых клетках». Наука. 351 (6274): aad5510. Дои:10.1126 / science.aad5510. ЧВК 4811353. PMID 26797145.
- ^ Sandoval-Guzmán, T .; Wang, H .; Хаттак, С .; Schuez, M .; Roensch, K .; Nacu, E .; Tazaki, A .; Joven, A .; Tanaka, E.M .; Саймон, А. С. (2014). «Фундаментальные различия в дифференцировке и рекрутировании стволовых клеток во время регенерации скелетных мышц у двух видов саламандр». Стволовая клетка. 14 (2): 174–87. Дои:10.1016 / j.stem.2013.11.007. PMID 24268695.
- ^ Курода, Y; Wakao, S; Китада, М; Мураками, Т; Нодзима, М. Дезава, М (2013). «Выделение, культивирование и оценка многолинейных дифференцирующихся стрессоустойчивых (Muse) клеток». Нат Проток. 8 (7): 1391–415. Дои:10.1038 / nprot.2013.076. PMID 23787896.
- ^ Kuroda, Y .; Китада, М .; Wakao, S .; и другие. (2010). «Уникальные мультипотентные клетки в популяциях мезенхимальных клеток взрослого человека». PNAS. 107 (19): 8639–8643. Bibcode:2010PNAS..107.8639K. Дои:10.1073 / pnas.0911647107. ЧВК 2889306. PMID 20421459.
- ^ Огура, Ф; Wakao, S; Курода, Y; Цучияма, К. Багери, М; Heneidi, S; Чазенбалк, G; Айба, S; Дезава, М (2014). «Человеческая жировая ткань обладает уникальной популяцией плюрипотентных стволовых клеток с неопухолевой и низкой теломеразной активностью: потенциальные последствия для регенеративной медицины». Стволовые клетки Dev. 23 (7): 717–28. Дои:10.1089 / scd.2013.0473. PMID 24256547.
- ^ Heneidi, S; Симерман, АА; Келлер, Э; Singh, P; Ли, Х; и другие. (2013). «Пробужденный клеточным стрессом: выделение и характеристика новой популяции плюрипотентных стволовых клеток, полученных из жировой ткани человека». PLOS ONE. 8 (6): e64752. Bibcode:2013PLoSO ... 864752H. Дои:10.1371 / journal.pone.0064752. ЧВК 3673968. PMID 23755141.
- ^ Шигемото, Т; Курода, Y; Wakao, S; Дезава, М (2013). «Новый подход к сбору сателлитных клеток из скелетных мышц взрослых на основе их устойчивости к стрессу». Стволовые клетки Trans Med. 2 (7): 488–498. Дои:10.5966 / sctm.2012-0130. ЧВК 3697816. PMID 23748608.
- ^ Сисахтнежад, С .; Матин, М. М. (2012). «Трансдифференцировка: процесс перепрограммирования клеток и молекул». Исследования клеток и тканей. 348 (3): 379–96. Дои:10.1007 / s00441-012-1403-у. PMID 22526624.
- ^ Меррелл, Эллисон Дж. (2016). «Пластичность взрослых клеток in vivo: де-дифференцировка и трансдифференцировка снова в моде». Обзоры природы Молекулярная клеточная биология. 17 (7): 413–425. Дои:10.1038 / nrm.2016.24. ЧВК 5818993. PMID 26979497.
- ^ Диннес, Андраш; Тиан, Сючунь Синди; Обак, Бьорн (17 апреля 2013 г.). Роберт А. Мейерс (ред.). Перенос ядер для клонирования животных. Джон Вили и сыновья. С. 299–344. ISBN 978-3-527-66854-0.
- ^ Жюльен, Джером; Паске, Винсент; Halley-Stott, Ричард П .; Миямото, Кей; Гурдон, Дж. Б. (23 июня 2011 г.). «Механизмы ядерного репрограммирования яйцами и ооцитами: детерминированный процесс?». Обзоры природы Молекулярная клеточная биология. 12 (7): 453–459. Дои:10.1038 / nrm3140. ЧВК 3657683. PMID 21697902.
- ^ Кэмпбелл, Кейт Х. С. (март 2002 г.). «Предыстория переноса ядер и их применения в сельском хозяйстве и терапевтической медицине *». Журнал анатомии. 200 (3): 267–275. Дои:10.1046 / j.1469-7580.2002.00035.x. ЧВК 1570687. PMID 12033731.
- ^ 8 647 872 долларов США патент
- ^ а б Chung, Y. G .; Eum, J. H .; Lee, J. E .; Shim, S. H .; Sepilian, V .; Hong, S.W .; Lee, Y .; Treff, N.R .; Choi, Y.H .; Kimbrel, E.A .; Dittman, R.E .; Lanza, R .; Ли, Д. Р. (2014). «Перенос ядра соматической клетки человека с использованием взрослых клеток». Стволовая клетка. 14 (6): 777–780. Дои:10.1016 / j.stem.2014.03.015. PMID 24746675.
- ^ Махла RS (2016). «Применение стволовых клеток в регенеративной медицине и трепевтике болезней». Международный журнал клеточной биологии. 2016 (7): 1–24. Дои:10.1155/2016/6940283. ЧВК 4969512. PMID 27516776.
- ^ Ян, H .; Shi, L .; Wang, B.A .; Liang, D .; Чжун, С .; Liu, W .; Nie, Y .; Liu, J .; Zhao, J .; Gao, X .; Li, D .; Xu, G.L .; Ли, Дж. (2012). «Получение генетически модифицированных мышей путем инъекции в ооциты андрогенетических гаплоидных эмбриональных стволовых клеток». Клетка. 149 (3): 605–617. Дои:10.1016 / j.cell.2012.04.002. PMID 22541431.
- ^ Hayashi, K .; Огуши, С .; Куримото, К .; Shimamoto, S .; Ohta, H .; Сайтоу М. (2012). «Потомство из ооцитов, полученных in vitro из первичных зародышевых клеток, подобных клеткам мышей». Наука. 338 (6109): 971–975. Bibcode:2012Наука ... 338..971H. Дои:10.1126 / science.1226889. PMID 23042295.
- ^ Кишигами, S .; Mizutani, E .; Ohta, H .; Hikichi, T .; Thuan, N.V .; Wakayama, S .; Bui, H.T .; Вакаяма, Т. (2006). «Значительное улучшение техники клонирования мышей при лечении трихостатином А после переноса соматического ядра». Сообщения о биохимических и биофизических исследованиях. 340 (1): 183–189. Дои:10.1016 / j.bbrc.2005.11.164. PMID 16356478.
- ^ Wakayama, S .; Кохда, Т .; Обоката, H .; Токоро, М .; Li, C .; Terashita, Y .; Mizutani, E .; Nguyen, V.T .; Кишигами, S .; Ишино, Ф .; Вакаяма, Т. (2013). «Успешное последовательное повторное клонирование при помощи мыши в нескольких поколениях». Стволовая клетка. 12 (3): 293–297. Дои:10.1016 / j.stem.2013.01.005. PMID 23472871.
- ^ "Официальный сайт Президентской комиссии по изучению вопросов биоэтики". Архивировано из оригинал на 2008-09-16. Получено 2014-02-25.
- ^ Paull, D .; Emmanuele, V .; Weiss, K. A .; Treff, N .; Стюарт, Л .; Hua, H .; Циммер, М .; Kahler, D. J .; Goland, R. S .; Noggle, S.A .; Проссер, Р .; Hirano, M .; Sauer, M. V .; Эгли, Д. (2012). «Перенос ядерного генома в ооциты человека устраняет варианты митохондриальной ДНК». Природа. 493 (7434): 632–637. Дои:10.1038 / природа11800. PMID 23254936.
- ^ Tachibana, M .; Amato, P .; Спарман, М .; Woodward, J .; Санчис, Д. М .; Ma, H .; Gutierrez, N.M .; Типпнер-Хеджес, Р.; Kang, E .; Lee, H. S .; Ramsey, C .; Мастерсон, К .; Battaglia, D .; Ли, Д .; Wu, D .; Jensen, J .; Patton, P .; Gokhale, S .; Stouffer, R .; Миталипов, С. (2012). «На пути к генной терапии наследственных митохондриальных заболеваний зародышевой линии». Природа. 493 (7434): 627–631. Дои:10.1038 / природа11647. ЧВК 3561483. PMID 23103867.
- ^ Отметьте Hayden, E. (2013). «Регулирующие органы взвешивают преимущества« трех родительского »оплодотворения». Природа. 502 (7471): 284–285. Bibcode:2013Натура.502..284C. Дои:10.1038 / 502284a. PMID 24132269.
- ^ Le, R .; Kou, Z .; Jiang, Y .; Li, M .; Хуанг, Б .; Liu, W .; Li, H .; Kou, X .; Он, W .; Rudolph, K. L .; Ju, Z .; Гао, С. (2014). «Усиленное омоложение теломер в плюрипотентных клетках, перепрограммированных посредством переноса ядра по сравнению с индуцированными плюрипотентными стволовыми клетками». Стволовая клетка. 14 (1): 27–39. Дои:10.1016 / j.stem.2013.11.005. PMID 24268696.
- ^ Чибелли, Хосе; Ланца, Роберт; Кэмпбелл, Кейт Х.С.; Уэст, Майкл Д. (14 сентября 2002 г.). Принципы клонирования. Академическая пресса. ISBN 978-0-08-049215-5.
- ^ Shinagawa, T .; Takagi, T .; Tsukamoto, D .; Tomaru, C .; Huynh, L.M .; Sivaraman, P .; Кумаревель, Т .; Inoue, K .; Nakato, R .; Katou, Y .; Садо, Т .; Takahashi, S .; Огура, А .; Shirahige, K .; Исии, С. (2014). «Варианты гистонов, обогащенные ооцитами, усиливают репрограммирование индуцированных плюрипотентных стволовых клеток». Стволовая клетка. 14 (2): 217–227. Дои:10.1016 / j.stem.2013.12.015. PMID 24506885.
- ^ а б Abad, M. A .; Mosteiro, L .; Pantoja, C .; Cañamero, M .; Район, Т .; Ors, I .; Graña, O .; Megías, D .; Domínguez, O .; Мартинес, Д .; Manzanares, M .; Ортега, С .; Серрано, М. (2013). «Перепрограммирование in vivo дает тератомы и iPS-клетки с характеристиками тотипотентности». Природа. 502 (7471): 340–345. Bibcode:2013Натура.502..340A. Дои:10.1038 / природа12586. PMID 24025773.
Наик, Гаутам (11 сентября 2013 г.). «Новое обещание для стволовых клеток - WSJ.com». Online.wsj.com. Получено 2014-01-30. - ^ Вандербург Б.Б. (2017). Дефицит микроРНК miR-34a вызывает тотипотентные стволовые клетки. ReliaWire
- ^ Джин Чой, Йонг; Линь, Чао-По; Риссо, Давиде; Чен, Шон; Аквинский Ким, Фома; Как Тан, Мэн; Билли Ли, Джин; Ву, Ялей; Чен, Кайфу; Сюань, Чжэньюй; Макфарлан, Тодд; Пэн, Вэйцзюнь; Ллойд, К. К. Кент; Ён Ким, Санг; Скорость, Теренс П .; Он, Лин (2017). «Дефицит microRNAmiR-34a увеличивает потенциал клеточной судьбы в плюрипотентных стволовых клетках». Наука. 355 (6325): eaag1927. Дои:10.1126 / science.aag1927. ЧВК 6138252. PMID 28082412.
- ^ Стивенс, Л. С. (1970). «Развитие трансплантируемых тератокарцином из интратестикулярных трансплантатов до- и постимплантационных эмбрионов мыши». Биология развития. 21 (3): 364–382. Дои:10.1016/0012-1606(70)90130-2. PMID 5436899.
- ^ Минц, B; Кронмиллер, C; Кастер, Р. П. (1978). «Тератокарциномы соматического происхождения». Труды Национальной академии наук Соединенных Штатов Америки. 75 (6): 2834–2838. Bibcode:1978ПНАС ... 75.2834М. Дои:10.1073 / pnas.75.6.2834. ЧВК 392659. PMID 275854.
- ^ Минц, B; Illmensee, K (1975). «Нормальные генетически мозаичные мыши, полученные из клеток злокачественной тератокарциномы». Proc Natl Acad Sci USA. 72 (9): 3585–3589. Bibcode:1975ПНАС ... 72.3585М. Дои:10.1073 / пнас.72.9.3585. ЧВК 433040. PMID 1059147.
- ^ Martin, G.R .; Эванс, М. Дж. (1975). «Дифференциация клональных линий клеток тератокарциномы: формирование эмбриоидных телец in vitro». Proc Natl Acad Sci USA. 72 (4): 1441–1445. Bibcode:1975ПНАС ... 72.1441М. Дои:10.1073 / pnas.72.4.1441. ЧВК 432551. PMID 1055416.
- ^ Illmensee, K; Минц, Б. (1976). «Totipotency и нормальная дифференцировка единичных клеток тератокарциномы, клонированных путем инъекции в бластоцисты». Труды Национальной академии наук Соединенных Штатов Америки. 73 (2): 549–553. Bibcode:1976ПНАС ... 73..549И. Дои:10.1073 / пнас.73.2.549. ЧВК 335947. PMID 1061157.
- ^ Мартин, Г. Р. (1981). «Выделение плюрипотентной клеточной линии из ранних эмбрионов мыши, культивированных в среде, кондиционированной стволовыми клетками тератокарциномы». Proc Natl Acad Sci USA. 78 (12): 7634–7638. Bibcode:1981PNAS ... 78.7634M. Дои:10.1073 / pnas.78.12.7634. ЧВК 349323. PMID 6950406.
- ^ Мартин, Г. Р. (1980). «Тератокарциномы и эмбриогенез млекопитающих». Наука. 209 (4458): 768–776. Bibcode:1980Sci ... 209..768M. Дои:10.1126 / science.6250214. PMID 6250214.
- ^ Papaioannou, V.E .; Gardner, R.L .; McBurney, M.W .; Бабине, С; Эванс, М. Дж. (1978). «Участие культивируемых клеток тератокарциномы в эмбриогенезе мышей». Журнал эмбриологии и экспериментальной морфологии. 44: 93–104. PMID 650144.
- ^ Грэм, К. Ф. (январь 1977 г.). Клетки тератокарциномы и нормальный эмбриогенез мыши мыши = Майкл И. Шерман. MIT Press. ISBN 978-0-262-19158-6.
- ^ Ильмензее, К. (14 июня 2012 г.). Л. Б. Рассел (ред.). Обратное развитие злокачественных новообразований и нормализация дифференцировки клеток тератокарциномы у химерных мышей. Springer London, Limited. С. 3–24. ISBN 978-1-4684-3392-0. PMID 378217.
- ^ Стюарт, Х. Т .; Van Oosten, A. L .; Радзишевская, А .; Мартелло, G .; Миллер, А .; Dietmann, S .; Nichols, J .; Сильва, Дж. С. Р. (2014). «NANOG усиливает активацию STAT3 и синергетически стимулирует наивную плюрипотентную программу». Текущая биология. 24 (3): 340–346. Дои:10.1016 / j.cub.2013.12.040. ЧВК 3982126. PMID 24462001.
- ^ Чой, Джихо; Ли, Сухён; Кряква, Уильям; и другие. (2015). «Сравнение генетически подобранных клеточных линий показывает эквивалентность ИПСК человека и ЭСК». Природа Биотехнологии. 33 (11): 1173–1181. Дои:10.1038 / nbt.3388. ЧВК 4847940. PMID 26501951.
- ^ Boland, M. J .; Hazen, J. L .; Назор, К. Л .; Родригес, А. Р .; Гиффорд, Вт; Мартин, G; Куприянов, С; Болдуин, К. К. (2009). «Взрослые мыши, полученные из индуцированных плюрипотентных стволовых клеток». Природа. 461 (7260): 91–94. Bibcode:2009Натура 461 ... 91Б. Дои:10.1038 / природа08310. PMID 19672243.
Канг, Л .; Wang, J .; Zhang, Y .; Kou, Z .; Гао, С. (2009). «Клетки IPS могут поддерживать полноценное развитие тетраплоидных эмбрионов с бластоцистами». Стволовая клетка. 5 (2): 135–138. Дои:10.1016 / j.stem.2009.07.001. PMID 19631602. - ^ Качество индуцированных плюрипотентных стволовых клеток значительно повышается за счет исключения того, что считалось наиболее важным фактором репрограммирования.
- ^ Исключение Oct4 из коктейля Яманака раскрывает потенциал развития ИПСК
- ^ Яги, Т .; Kosakai, A .; Ито, Д .; Okada, Y .; Akamatsu, W .; Nihei, Y .; Nabetani, A .; Ishikawa, F .; Arai, Y .; Hirose, N .; Окано, H .; Сузуки, Н. (2012). «Создание индуцированных плюрипотентных стволовых клеток от долгожителей для исследования нейродегенеративных заболеваний». PLOS ONE. 7 (7): e41572. Bibcode:2012PLoSO ... 741572Y. Дои:10.1371 / journal.pone.0041572. ЧВК 3405135. PMID 22848530.
- ^ Rohani, L .; Johnson, A. A .; Арнольд, А .; Штольцинг, А. (2014). «Признак старения: признак индуцированных плюрипотентных стволовых клеток?». Ячейка старения. 13 (1): 2–7. Дои:10.1111 / acel.12182. ЧВК 4326871. PMID 24256351.
- ^ Yehezkel, S; Ребибо-Саббах, А; Сегев, У; Цукерман, М; Shaked, R; Хубер, я; Гепштейн, Л; Скорецкий, К; Селиг, S (2011). «Перепрограммирование теломерных областей во время создания индуцированных человеком плюрипотентных стволовых клеток и последующей дифференциации в фибробластоподобные производные». Эпигенетика. 6 (1): 63–75. Дои:10.4161 / epi.6.1.13390. ЧВК 3052915. PMID 20861676.
- ^ West, M.D .; Вазири, Х (2010). «Назад к бессмертию: восстановление длины теломер эмбриона во время индуцированной плюрипотентности». Регенеративная медицина. 5 (4): 485–488. Дои:10.2217 / rme.10.51. PMID 20632849.
- ^ Marión, R.M .; Бласко, М.А. (2010). «Омоложение теломер при ядерном репрограммировании». Текущее мнение в области генетики и развития. 20 (2): 190–196. Дои:10.1016 / j.gde.2010.01.005. PMID 20176474.
Gourronc, F.A .; Клингельхутц, А. Дж. (2012). «Терапевтические возможности: поддержание теломер в индуцибельных плюрипотентных стволовых клетках». Мутационные исследования / Фундаментальные и молекулярные механизмы мутагенеза. 730 (1–2): 98–105. Дои:10.1016 / j.mrfmmm.2011.05.008. ЧВК 3179558. PMID 21605571. - ^ Чжао, Т .; Zhang, Z. N .; Rong, Z .; Сюй, Ю. (2011). «Иммуногенность индуцированных плюрипотентных стволовых клеток». Природа. 474 (7350): 212–215. Дои:10.1038 / природа10135. PMID 21572395.
- ^ Dhodapkar, M. V .; Дходапкар, К. М. (2011). «Спонтанный и вызванный терапией иммунитет к генам плюрипотентности у людей: клинические последствия, возможности и проблемы». Иммунология рака, Иммунотерапия. 60 (3): 413–418. Дои:10.1007 / s00262-010-0944-8. ЧВК 3574640. PMID 21104412.
- ^ Gutierrez-Aranda, I .; Ramos-Mejia, V .; Bueno, C .; Munoz-Lopez, M .; Реал, П. Дж .; Mácia, A .; Sanchez, L .; Ligero, G .; Garcia-Parez, J. L .; Менендес, П. (2010). «Плюрипотентные стволовые клетки человека развивают тератому более эффективно и быстрее, чем эмбриональные стволовые клетки человека, независимо от места инъекции». Стволовые клетки. 28 (9): 1568–1570. Дои:10.1002 / шток.471. ЧВК 2996086. PMID 20641038.
- ^ Chang, C.J .; Mitra, K .; Koya, M .; Velho, M .; Desprat, R .; Lenz, J .; Бухасира, Э. Э. (2011). «Производство эмбриональных и эмбриональных эритроцитов из индуцированных человеком плюрипотентных стволовых клеток». PLOS ONE. 6 (10): e25761. Bibcode:2011PLoSO ... 625761C. Дои:10.1371 / journal.pone.0025761. ЧВК 3192723. PMID 22022444.
- ^ а б Дабир, Д. В .; Hasson, S.A .; Setoguchi, K .; Johnson, M.E .; Wongkongkathep, P .; Douglas, C.J .; Zimmerman, J .; Damoiseaux, R .; Teitell, M. A .; Келер, К. М. (2013). "Малые молекулы ингибитора редокс-регулируемой транслокации белков в митохондрии". Клетка развития. 25 (1): 81–92. Дои:10.1016 / j.devcel.2013.03.006. ЧВК 3726224. PMID 23597483.
- ^ Бен-Давид, Ури; Ган, Цин-Фэнь; Голан-Лев, Тамар; и другие. (2013). «Селективное удаление плюрипотентных стволовых клеток человека с помощью ингибитора синтеза олеата, обнаруженного на высокопроизводительном экране». Стволовая клетка. 12 (2): 167–179. Дои:10.1016 / j.stem.2012.11.015. PMID 23318055.
- ^ Лу, К. Дж. (2013). «Маленькие молекулы против тератом». Биржа науки и бизнеса. 6 (7): 158. Дои:10.1038 / scibx.2013.158.
- ^ Ли, М. -О .; Moon, S. H .; Jeong, H. -C .; Yi, J. -Y .; Lee, T. -H .; Shim, S. H .; Rhee, Y. -H .; Lee, S. -H .; О, С. -Дж .; Lee, M. -Y .; Han, M. -J .; Чо, Ю. С .; Chung, H. -M .; Kim, K. -S .; Ча, Х. -Дж. (2013). «Ингибирование образования тератомы, полученной из плюрипотентных стволовых клеток, небольшими молекулами». Труды Национальной академии наук. 110 (35): E3281–90. Bibcode:2013PNAS..110E3281L. Дои:10.1073 / pnas.1303669110. ЧВК 3761568. PMID 23918355.
- ^ Chaffer, C.L .; Brueckmann, I .; Scheel, C .; Kaestli, A.J .; Wiggins, P.A .; Rodrigues, L.O .; Brooks, M .; Reinhardt, F .; Вс, Ы .; Поляк, К .; Arendt, L.M .; Kuperwasser, C .; Bierie, B .; Вайнберг, Р. А. (2011). «Нормальные и неопластические не стволовые клетки могут спонтанно превращаться в стеблевое состояние». Труды Национальной академии наук. 108 (19): 7950–7955. Bibcode:2011PNAS..108.7950C. Дои:10.1073 / pnas.1102454108. ЧВК 3093533. PMID 21498687.
Фу, Вэй (2012). «Остаточные недифференцированные клетки во время дифференцировки индуцированных плюрипотентных стволовых клеток in vitro и in vivo». Стволовые клетки и развитие. 21 (4): 521–529. Дои:10.1089 / scd.2011.0131. PMID 21631153. - ^ Гуртан, А. М .; Рави, А .; Rahl, P. B .; Bosson, A.D .; Jnbaptiste, C.K .; Bhutkar, A .; Whittaker, C.A .; Янг, Р. А .; Шарп, П. А. (2013). «Let-7 подавляет Nr6a1 и программу развития в середине беременности во взрослых фибробластах». Гены и развитие. 27 (8): 941–954. Дои:10.1101 / gad.215376.113. ЧВК 3650230. PMID 23630078.
Wang, H .; Ван, X .; Сюй, X .; Zwaka, T. P .; Куни, А. Дж. (2013). «Эпигенетическое репрограммирование гена ядерного фактора зародышевой клетки необходимо для правильной дифференциации индуцированных плюрипотентных клеток». Стволовые клетки. 31 (12): 2659–2666. Дои:10.1002 / шток.1367. ЧВК 3731425. PMID 23495137. - ^ Lindgren, A. G .; Нацухара, К .; Tian, E .; Винсент, Дж. Дж .; Li, X .; Jiao, J .; Wu, H .; Banerjee, U .; Кларк, А. Т. (2011). «Потеря Pten вызывает начало опухоли после дифференциации мышиных плюрипотентных стволовых клеток из-за неудачной репрессии Nanog». PLOS ONE. 6 (1): e16478. Bibcode:2011PLoSO ... 616478L. Дои:10.1371 / journal.pone.0016478. ЧВК 3029365. PMID 21304588.
- ^ Град, я; Hibaoui, Y; Jaconi, M; Чича, Л; Бергстрём-Тенгзелиус, Р. Sailani, M. R .; Pelte, M. F .; Дахун, S; Mitsiadis, T. A .; Töhönen, V; Bouillaguet, S; Antonarakis, S.E .; Кере, Дж; Zucchelli, M; Ховатта, О; Феки, А (2011). «Праймирование NANOG перед полным перепрограммированием может вызвать опухоли зародышевых клеток». Европейские клетки и материалы. 22: 258–274, обсуждение 274. Дои:10.22203 / ecm.v022a20. PMID 22071697.
- ^ Окано, H .; Накамура, М .; Yoshida, K .; Okada, Y .; Tsuji, O .; Nori, S .; Ikeda, E .; Yamanaka, S .; Миура, К. (2013). «Шаги к безопасной клеточной терапии с использованием индуцированных плюрипотентных стволовых клеток». Циркуляционные исследования. 112 (3): 523–533. Дои:10.1161 / CIRCRESAHA.111.256149. PMID 23371901.
Cunningham, J. J .; Ulbright, T. M .; Pera, M. F .; Лойенга, Л. Х. Дж. (2012). «Уроки тератом человека для разработки безопасных методов лечения стволовыми клетками». Природа Биотехнологии. 30 (9): 849–857. Дои:10.1038 / nbt.2329. PMID 22965062. - ^ Беллин, М .; Marchetto, M.C .; Gage, F. H .; Маммери, К. Л. (2012). «Индуцированные плюрипотентные стволовые клетки: новый пациент?». Обзоры природы Молекулярная клеточная биология. 13 (11): 713–726. Дои:10.1038 / nrm3448. PMID 23034453.
Sandoe, J .; Эгган, К. (2013). «Возможности и проблемы моделей нейродегенеративных заболеваний плюрипотентных стволовых клеток». Природа Неврология. 16 (7): 780–789. Дои:10.1038 / № 3425. PMID 23799470. - ^ Takahashi, K .; Яманака, С. (2013). «Индуцированные плюрипотентные стволовые клетки в медицине и биологии». Разработка. 140 (12): 2457–2461. Дои:10.1242 / dev.092551. PMID 23715538.
Fu, X; Сюй, Y (2012). «Проблемы клинического применения плюрипотентных стволовых клеток: на пути к геномной и функциональной стабильности». Геномная медицина. 4 (6): 55. Дои:10,1186 / gm354. ЧВК 3698533. PMID 22741526. - ^ Araki, R .; Уда, М .; Hoki, Y .; Sunayama, M .; Накамура, М .; Ando, S .; Sugiura, M .; Ideno, H .; Shimada, A .; Nifuji, A .; Абэ, М. (2013). «Незначительная иммуногенность терминально дифференцированных клеток, полученных из индуцированных плюрипотентных или эмбриональных стволовых клеток». Природа. 494 (7435): 100–104. Bibcode:2013Натура.494..100А. Дои:10.1038 / природа11807. PMID 23302801.
Wahlestedt, M .; Norddahl, G.L .; Sten, G .; Угале, А .; Фриск, М. -А. М .; Mattsson, R .; Deierborg, T .; Sigvardsson, M .; Брайдер, Д. (2013). «Эпигенетический компонент старения гемопоэтических стволовых клеток, поддающийся перепрограммированию в молодое состояние» (PDF). Кровь. 121 (21): 4257–4264. Дои:10.1182 / кровь-2012-11-469080. PMID 23476050. - ^ Охниши, К .; Семи, К .; Ямамото, Т .; Shimizu, M .; Tanaka, A .; Mitsunaga, K .; Ямада, Ю. (2014). «Преждевременное прекращение репрограммирования in vivo приводит к развитию рака через изменение эпигенетической регуляции». Клетка. 156 (4): 663–677. Дои:10.1016 / j.cell.2014.01.005. PMID 24529372.
- ^ Mfopou JK, De Groote V, Xu X, Heimberg H, Bouwens L (май 2007 г.). «Звуковой еж и другие растворимые факторы дифференциации эмбриоидных телец тормозят развитие поджелудочной железы». Стволовые клетки. 25 (5): 1156–1165. Дои:10.1634 / стволовые клетки.2006-0720. PMID 17272496.
- ^ Де Лос, Анхелес; Дейли, Г.К. (2013). «Химическая логика для перепрограммирования плюрипотентности». Клеточные исследования. 23 (12): 1337–1338. Дои:10.1038 / cr.2013.119. ЧВК 3847567. PMID 23979017.
- ^ Federation, A.J .; Bradner, J. E .; Мейснер, А. (2013). «Использование малых молекул в репрограммировании соматических клеток». Тенденции в клеточной биологии. 24 (3): 179–187. Дои:10.1016 / j.tcb.2013.09.011. ЧВК 3943685. PMID 24183602.
- ^ Zhao, Y .; Чжао, Т .; Гуань, Дж. (2015). «XEN-подобное государство связывает соматические клетки с плюрипотентностью во время химического репрограммирования». Клетка. 163 (7): 1678–1691. Дои:10.1016 / j.cell.2015.11.017. PMID 26686652.
- ^ Накаучи Хиромицу; Камия Акихидэ; Сузуки Нао; Ито Кейичи; Ямазаки Сатоши (2011) Способ получения клеток, индуцированных для дифференциации от плюрипотентных стволовых клеток. Заявка на договор о патентной кооперации, патент: WO2011071085 (A1). 2011-06-16 (C12N5 / 07)
- ^ Amabile, G .; Welner, R. S .; Nombela-Arrieta, C .; d'Alise, A.M .; Di Ruscio, A .; Ебралидзе, А.К .; Крайцберг, Ю .; Е, М .; Кочер, О .; Neuberg, D. S .; Храпко, К .; Зильберштейн, Л. Э .; Тенен, Д. Г. (2012). «Получение in vivo трансплантируемых гемопоэтических клеток человека из индуцированных плюрипотентных стволовых клеток». Кровь. 121 (8): 1255–1264. Дои:10.1182 / кровь-2012-06-434407. ЧВК 3701251. PMID 23212524.
- ^ Suzuki, N .; Yamazaki, S .; Yamaguchi, T .; Окабе, М .; Masaki, H .; Takaki, S .; Оцу, М .; Накаучи, Х. (2013). «Получение приживляемых гемопоэтических стволовых клеток из индуцированных плюрипотентных стволовых клеток путем образования тератомы». Молекулярная терапия. 21 (7): 1424–1431. Дои:10,1038 / мт.2013,71. ЧВК 3705943. PMID 23670574.
- ^ Chou, B.K .; Ye, Z .; Ченг, Л. (2013). «Генерация и насаждение гематопоэтических клеток, полученных из ИПСК, in vivo». Молекулярная терапия. 21 (7): 1292–1293. Дои:10.1038 / мт.2013.129. ЧВК 3702102. PMID 23812546.
- ^ Ямаути, Т; Такенака, К; Урата, S; Шима, Т; Кикушигэ, Y; Токуяма, Т; Ивамото, К; Нишихара, М; Ивасаки, H; Миямото, Т; Honma, N; Накао, М; Матодзаки, Т; Акаши, К. (2013). «Полиморфная Sirpa является генетической детерминантой для линий мышей на основе NOD для достижения эффективного приживления клеток человека». Кровь. 121 (8): 1316–1325. Дои:10.1182 / кровь-2012-06-440354. PMID 23293079.
- ^ Rong, Z .; Wang, M .; Hu, Z .; Страднер, М .; Zhu, S .; Kong, H .; Yi, H .; Goldrath, A .; Yang, Y.G .; Xu, Y .; Фу, X. (2014). «Эффективный подход к предотвращению иммунного отторжения аллотрансплантатов, полученных из ESC человека». Стволовая клетка. 14 (1): 121–130. Дои:10.1016 / j.stem.2013.11.014. ЧВК 4023958. PMID 24388175.
- ^ Hirami, Y .; Osakada, F .; Takahashi, K .; Окита, К .; Yamanaka, S .; Ikeda, H .; Yoshimura, N .; Такахаши, М. (2009). «Генерация клеток сетчатки из плюрипотентных стволовых клеток мыши и человека». Письма о неврологии. 458 (3): 126–131. Дои:10.1016 / j.neulet.2009.04.035. HDL:2433/97948. PMID 19379795.
- ^ Buchholz, D. E .; Hikita, S.T .; Rowland, T. J .; Фридрих, А. М .; Hinman, C.R .; Johnson, L.V .; Клегг, Д. О. (2009). «Получение функционального пигментного эпителия сетчатки из индуцированных плюрипотентных стволовых клеток». Стволовые клетки. 27 (10): 2427–2434. Дои:10.1002 / стержень.189. PMID 19658190.
- ^ Ян, Джин; Нонг, Ева; Цанг, Стивен Х (2013). «Индуцированные плюрипотентные стволовые клетки и лечение дегенерации сетчатки». Эксперт преподобный офтальмол. 8 (1): 5–8. Дои:10.1586 / EOP.12.75.
- ^ Поля, Марка А .; Хван, Джон; Гонг, Цзе; Цай, Хуэй; Дель Приоре, Лучиан (9 декабря 2012 г.). Стивен Цанг (ред.). Глаз как орган-мишень для терапии стволовыми клетками. Springer. С. 1–30. ISBN 978-1-4614-5493-9.
- ^ Ли, У; Цай, Ю.Т .; Хсу, CW; и другие. (2012). «Долгосрочная безопасность и эффективность трансплантатов индуцированных человеком плюрипотентных стволовых клеток (iPS) на доклинической модели пигментного ретинита». Мол. Med. 18 (1): 1312–1319. Дои:10.2119 / молмед.2012.00242. ЧВК 3521789. PMID 22895806.
- ^ «Могут ли стволовые клетки снова заставить слепых видеть?. Образ жизни исследователя. 15 июля 2013 г.
- ^ Tzouvelekis, A .; Ntolios, P .; Бурос, Д. (2013). «Лечение стволовыми клетками хронических заболеваний легких». Дыхание. 85 (3): 179–192. Дои:10.1159/000346525. PMID 23364286.
- ^ Вагнер, Дарси Э .; Бонвиллен, Райан В .; Дженсен, Тодд; Жирар, Эрик Д .; Bunnell, Bruce A .; Финк, Кристин М .; Hoffman5, Andrew M .; Weiss2, Daniel J. (июль 2013 г.). «Можно ли использовать стволовые клетки для создания новых легких? Биоинженерия легких ex vivo с децеллюляризованными каркасами цельного легкого». Респирология. 18 (6): 895–911. Дои:10.1111 / соотв. 12102. ЧВК 3729745. PMID 23614471.
- ^ Wong, A. P .; Россант, Дж. (2013). «Создание эпителия легких из плюрипотентных стволовых клеток». Текущие отчеты патобиологии. 1 (2): 137–145. Дои:10.1007 / s40139-013-0016-9. ЧВК 3646155. PMID 23662247.
- ^ Mou, H .; Zhao, R .; Sherwood, R .; Ahfeldt, T .; Lapey, A .; Wain, J .; Сицилийский, L .; Извольский, К .; Lau, F. H .; Musunuru, K .; Cowan, C .; Раджагопал, Дж. (2012). «Создание мультипотентных предшественников легких и дыхательных путей из мышиных ESC и ИПСК, специфичных для пациентов с кистозным фиброзом». Стволовая клетка. 10 (4): 385–397. Дои:10.1016 / j.stem.2012.01.018. ЧВК 3474327. PMID 22482504.
- ^ Ghaedi, M .; Calle, E. A .; Mendez, J. J .; Gard, A. L .; Balestrini, J .; Бут, А .; Бове, П. Ф .; Gui, L .; White, E. S .; Никласон, Л. Э. (2013). «Альвеолярный эпителий, полученный из iPS-клеток человека, повторно заселяет внеклеточный матрикс легких». Журнал клинических исследований. 123 (11): 4950–4962. Дои:10.1172 / JCI68793. ЧВК 3809786. PMID 24135142.
- ^ Ghaedi, M .; Mendez, J. J .; Бове, П. Ф .; Сиварапатна, А .; Raredon, M.S.B .; Никласон, Л. Э. (2014). «Дифференциация альвеолярного эпителия индуцированных человеком плюрипотентных стволовых клеток во вращающемся биореакторе». Биоматериалы. 35 (2): 699–710. Дои:10.1016 / j.biomaterials.2013.10.018. ЧВК 3897000. PMID 24144903.
- ^ Huang, S. X. L .; Islam, M. N .; О'Нил, Дж .; Hu, Z .; Yang, Y.G .; Чен, Ю. В .; Mumau, M .; Грин, M.D .; Vunjak-Novakovic, G .; Bhattacharya, J .; Снок, Х. В. (2013). «Эффективное создание эпителиальных клеток легких и дыхательных путей из плюрипотентных стволовых клеток человека». Природа Биотехнологии. 32 (1): 84–91. Дои:10.1038 / nbt.2754. ЧВК 4101921. PMID 24291815.
- ^ Niu, Z .; Hu, Y .; Чу, З .; Ю, М .; Bai, Y .; Wang, L .; Хуа, Дж. (2013). «Дифференциация зародышевых клеток из индуцированных плюрипотентных стволовых клеток (ИПСК)». Биохимия и функции клетки. 31 (1): 12–19. Дои:10.1002 / cbf.2924. PMID 23086862.
Ян, С .; Бо, Дж .; Хм.; Guo, X .; Tian, R .; Sun, C .; Zhu, Y .; Li, P .; Liu, P .; Zou, S .; Huang, Y .; Ли, З. (2012). «Получение мужских половых клеток из индуцированных плюрипотентных стволовых клеток in vitro и в восстановленных семенных канальцах». Распространение клеток. 45 (2): 91–100. Дои:10.1111 / j.1365-2184.2012.00811.x. ЧВК 6496715. PMID 22324506.
Panula, S .; Medrano, J. V .; Kee, K .; Bergstrom, R .; Nguyen, H.N .; Байерс, В .; Wilson, K. D .; Wu, J. C .; Саймон, С .; Hovatta, O .; Рейо Пера, Р.А. (2010). «Дифференциация половых клеток человека из плюрипотентных стволовых клеток, индуцированных плодом и взрослым». Молекулярная генетика человека. 20 (4): 752–762. Дои:10,1093 / hmg / ddq520. ЧВК 3024045. PMID 21131292. - ^ а б c Qian, L .; Huang, Y .; Spencer, C.I .; Фоли, А .; Vedantham, V .; Liu, L .; Конвей, С. Дж .; Fu, J.D .; Шривастава, Д. (2012). «In vivo перепрограммирование сердечных фибробластов мыши в индуцированные кардиомиоциты». Природа. 485 (7400): 593–598. Bibcode:2012Натура.485..593Q. Дои:10.1038 / природа11044. ЧВК 3369107. PMID 22522929.
- ^ Szabo, E .; Rampalli, S .; Risueño, R.M .; Schnerch, A .; Mitchell, R .; Fiebig-Comyn, A .; Levadoux-Martin, M .; Бхатия, М. (2010). «Прямое преобразование человеческих фибробластов в предшественников многократной крови». Природа. 468 (7323): 521–526. Bibcode:2010Натура.468..521С. Дои:10.1038 / природа09591. PMID 21057492.
- ^ Efe, J. A .; Hilcove, S .; Kim, J .; Чжоу, H .; Ouyang, K .; Wang, G .; Chen, J .; Дин, С. (2011). «Превращение мышиных фибробластов в кардиомиоциты с использованием стратегии прямого репрограммирования». Природа клеточной биологии. 13 (3): 215–222. Дои:10.1038 / ncb2164. PMID 21278734.
- ^ а б Lujan, E .; Chanda, S .; Ahlenius, H .; Sudhof, T. C .; Верниг, М. (2012). «Прямое превращение фибробластов мыши в самообновляющиеся трипотентные клетки-предшественники нейронов». Труды Национальной академии наук. 109 (7): 2527–2532. Bibcode:2012PNAS..109.2527L. Дои:10.1073 / pnas.1121003109. ЧВК 3289376. PMID 22308465.
- ^ а б Thier, M .; Wörsdörfer, P .; Озера, Ю. Б .; Gorris, R .; Herms, S .; Opitz, T .; Seiferling, D .; Quandel, T .; Hoffmann, P .; Nöthen, M. M .; Brüstle, O .; Эденхофер, Ф. (2012). «Прямое преобразование фибробластов в стабильно расширяемые нервные стволовые клетки». Стволовая клетка. 10 (4): 473–479. Дои:10.1016 / j.stem.2012.03.003. PMID 22445518.
- ^ а б c Han, D. W .; Tapia, N .; Германн, А .; Hemmer, K .; Höing, S .; Araúzo-Bravo, M.J .; Zaehres, H .; Wu, G .; Франк, S .; Moritz, S. R .; Гребер, Б .; Yang, J. H .; Lee, H.T .; Schwamborn, J.C .; Сторч, А .; Шелер, Х. Р. (2012). «Прямое перепрограммирование фибробластов в нервные стволовые клетки с помощью определенных факторов». Стволовая клетка. 10 (4): 465–472. Дои:10.1016 / j.stem.2012.02.021. PMID 22445517.
- ^ Величко, Сергей; Канг, Кюри; Ким, Сон Мин; Квак, Тэ Хван; Ким, Ки-Пё; Пак, Чанхёк; Хонг, Квонхо; Чанг, Чихе; Хён, Чон Гын (апрель 2019). «Слияние факторов репрограммирования меняет траекторию преобразования соматической линии передачи». Отчеты по ячейкам. 27 (1): 30–39.e4. Дои:10.1016 / j.celrep.2019.03.023. PMID 30943410.
- ^ Taylor, S.M .; Джонс, П. А. (1979). «Множество новых фенотипов, индуцированных в клетках 10T1 / 2 и 3T3, обработанных 5-азацитидином». Клетка. 17 (4): 771–779. Дои:10.1016/0092-8674(79)90317-9. PMID 90553.
- ^ Lassar, A. B .; Paterson, B.M .; Weintraub, H (1986). «Трансфекция локуса ДНК, который опосредует превращение фибробластов 10T1 / 2 в миобласты». Клетка. 47 (5): 649–656. Дои:10.1016/0092-8674(86)90507-6. PMID 2430720.
Davis, R. L .; Weintraub, H .; Лассар, А. Б. (1987). «Экспрессия одной трансфицированной кДНК превращает фибробласты в миобласты». Клетка. 51 (6): 987–1000. Дои:10.1016 / 0092-8674 (87) 90585-х. PMID 3690668.
Weintraub, H; Tapscott, S.J .; Davis, R. L .; Thayer, M. J .; Adam, M. A .; Lassar, A. B .; Миллер, А. Д. (1989). «Активация мышечно-специфических генов в клеточных линиях пигмента, нервов, жира, печени и фибробластов путем принудительной экспрессии MyoD». Труды Национальной академии наук Соединенных Штатов Америки. 86 (14): 5434–5438. Bibcode:1989PNAS ... 86,5434 Вт. Дои:10.1073 / pnas.86.14.5434. ЧВК 297637. PMID 2748593. - ^ Vierbuchen, T .; Верниг, М. (2011). «Прямое преобразование родословной: неестественно, но полезно?». Природа Биотехнологии. 29 (10): 892–907. Дои:10.1038 / nbt.1946. ЧВК 3222779. PMID 21997635.
- ^ Чандракантан, Ваше; и другие. (2016). «PDGF-AB и 5-азацитидин индуцируют преобразование соматических клеток в ткане-регенеративные мультипотентные стволовые клетки». Труды Национальной академии наук. 113 (16): E2306 – E2315. Bibcode:2016PNAS..113E2306C. Дои:10.1073 / pnas.1518244113. ЧВК 4843430. PMID 27044077.
- ^ Ученые разработали новую систему восстановления стволовых клеток. Портал стволовых клеток
- ^ Может ли эта новая стволовая клетка изменить правила игры для регенеративного лечения?. Ирландский экзаменатор
- ^ Риддл, М. Р .; Weintraub, A .; Nguyen, K. C. Q .; Холл, Д. Н .; Ротман, Дж. Х. (2013). «Трансдифференцировка и ремоделирование постэмбриональных клеток C. elegans с помощью одного фактора транскрипции». Разработка. 140 (24): 4844–4849. Дои:10.1242 / dev.103010. ЧВК 3848185. PMID 24257624.
- ^ Wei, S; Zou, Q; Lai, S; и другие. (2016). «Превращение эмбриональных стволовых клеток во внеэмбриональные клоны активаторами, опосредованными CRISPR». Sci. Представитель. 6: 19648. Bibcode:2016НатСР ... 619648Вт. Дои:10.1038 / srep19648. ЧВК 4726097. PMID 26782778.
- ^ Блэк, Джошуа Б.; Адлер, Андрей Ф .; Ван, Хун-Ганг; Д'Ипполито, Энтони М .; Hutchinson, Hunter A .; Редди, Тимоти Э .; Питт, Джеффри С .; Leong, Kam W .; Герсбах, Чарльз А. (2016). «Целевое эпигенетическое ремоделирование эндогенных локусов с помощью транскрипционных активаторов на основе CRISPR / Cas9 напрямую превращает фибробласты в нейрональные клетки». Стволовая клетка. 19 (3): 406–414. Дои:10.1016 / j.stem.2016.07.001. ЧВК 5010447. PMID 27524438.
- ^ Jung, D. W .; Уильямс, Д. Р. (2011). «Новый химически определенный подход к производству мультипотентных клеток из терминально дифференцированных тканевых синцитий». ACS Химическая биология. 6 (6): 553–562. Дои:10.1021 / cb2000154. PMID 21322636.
"В этом выпуске". ACS Химическая биология. 7 (4): 619. 2012. Дои:10.1021 / cb300127f. - ^ Чжан, Хункай; Уилсон, Ян А .; Лернер, Ричард А. (2012). «Выбор антител, регулирующих фенотип, из библиотек внутриклеточных комбинаторных антител». PNAS. 109 (39): 15728–15733. Bibcode:2012PNAS..10915728Z. Дои:10.1073 / pnas.1214275109. ЧВК 3465454. PMID 23019357.
- ^ Антитело, которое трансформирует стволовые клетки костного мозга непосредственно в клетки мозга
- ^ Xie, J .; Zhang, H .; Да, К .; Лернер, Р. А. (23 апреля 2013 г.). «Отбор комбинаторных антител на основе аутокринной передачи сигналов, которые трансдифференцируют стволовые клетки человека». Труды Национальной академии наук. 110 (20): 8099–8104. Bibcode:2013PNAS..110.8099X. Дои:10.1073 / pnas.1306263110. ЧВК 3657831. PMID 23613575.
- ^ Ринг, Карен (12 сентября 2017 г.). «Ученые, финансируемые CIRM, открывают новый способ получения стволовых клеток с использованием антител».
- ^ Это на.; Охта, К. (2015). «Перепрограммирование соматических клеток человека бактериями». Развитие, рост и дифференциация. 57 (4): 305–312. Дои:10.1111 / dgd.12209. PMID 25866152.
- ^ Установлено, что рибосомы индуцируют плюрипотентность соматических клеток. Технологические сети. НОВОСТИ 07 фев 2018
- ^ а б c d Лю, X .; Ори, В .; Chapman, S .; Юань, H .; Albanese, C .; Каллакури, Б .; Тимофеева, О. А .; Nealon, C .; Дакич, А .; Simic, V .; Haddad, B.R .; Rhim, J. S .; Дритчило, А .; Riegel, A .; McBride, A .; Шлегель, Р. (2012). «Клетки-ингибиторы ROCK и питающие клетки вызывают условное перепрограммирование эпителиальных клеток». Американский журнал патологии. 180 (2): 599–607. Дои:10.1016 / j.ajpath.2011.10.036. ЧВК 3349876. PMID 22189618.
- ^ Rheinwald, J. G .; Грин, H (1975). «Серийное культивирование штаммов эпидермальных кератиноцитов человека: образование ороговевших колоний из единичных клеток». Клетка. 6 (3): 331–343. Дои:10.1016 / S0092-8674 (75) 80001-8. PMID 1052771.
- ^ Хью, Ю.-Л. (2011). Изучение биологических последствий повреждения ДНК, вызванного облученными фидерными клетками фибробластов J2-3T3 и HPV16: характеристика биологических функций Mll (Кандидатская диссертация). Университетский колледж Лондона.
- ^ Шумиэль, И. (2012). «Радиационный гормезис: аутофагия и другие клеточные механизмы». Международный журнал радиационной биологии. 88 (9): 619–628. Дои:10.3109/09553002.2012.699698. PMID 22702489.
- ^ Куросава, Х. (2012). "Application of Rho-associated protein kinase (ROCK) inhibitor to human pluripotent stem cells". Журнал биологии и биоинженерии. 114 (6): 577–581. Дои:10.1016/j.jbiosc.2012.07.013. PMID 22898436.
- ^ Terunuma, A.; Limgala, R. P.; Park, C. J.; Choudhary, I.; Vogel, J. C. (2010). "Efficient Procurement of Epithelial Stem Cells from Human Tissue Specimens Using a Rho-Associated Protein Kinase Inhibitor Y-27632". Тканевая инженерия, часть А. 16 (4): 1363–1368. Дои:10.1089/ten.tea.2009.0339. ЧВК 2862604. PMID 19912046.
- ^ Wu, X., Wang, S., Li, M., Li, J., Shen, J., Zhao, Y., ... & Kaboli, P. J. (2020). Conditional reprogramming: next generation cell culture. Acta Pharmaceutica Sinica B. 10(8), 1360-1381 Дои:10.1016/j.apsb.2020.01.011
- ^ Chapman, Sandra; Liu, Xuefeng; Meyers, Craig; Schlegel, Richard; McBride, Alison A. (1 July 2010). "Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor". Журнал клинических исследований. 120 (7): 2619–2626. Дои:10.1172/JCI42297. ЧВК 2898606. PMID 20516646.
- ^ Suprynowicz, F. A.; Upadhyay, G.; Krawczyk, E.; Kramer, S. C.; Hebert, J. D.; Лю, X .; Юань, H .; Cheluvaraju, C.; Clapp, P. W.; Boucher, R. C.; Kamonjoh, C. M.; Randell, S. H.; Schlegel, R. (2012). "Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells". Труды Национальной академии наук. 109 (49): 20035–20040. Bibcode:2012PNAS..10920035S. Дои:10.1073/pnas.1213241109. ЧВК 3523865. PMID 23169653.
- ^ а б Agarwal, S .; Rimm, D. L. (2012). "Making Every Cell Like He Ла". Американский журнал патологии. 180 (2): 443–445. Дои:10.1016/j.ajpath.2011.12.001. PMID 22192626.
- ^ а б Lisanti, MP; Tanowitz, HB (Apr 2012). "Translational discoveries, personalized medicine, and living biobanks of the future". Американский журнал патологии. 180 (4): 1334–1336. Дои:10.1016/j.ajpath.2012.02.003. PMID 22453029.
- ^ Юань, H .; Myers, S.; Wang, J .; Чжоу, Д .; Woo, J. A.; Kallakury, B.; Ju, A.; Bazylewicz, M.; Carter, Y. M.; Albanese, C.; Grant, N.; Shad, A.; Dritschilo, A.; Лю, X .; Schlegel, R. (2012). "Use of Reprogrammed Cells to Identify Therapy for Respiratory Papillomatosis". Медицинский журнал Новой Англии. 367 (13): 1220–1227. Дои:10.1056/NEJMoa1203055. ЧВК 4030597. PMID 23013073.
- ^ Crystal, AS; Shaw, AT; Sequist, LV; Friboulet, L; Niederst, MJ; Lockerman, EL; Frias, RL; Gainor, JF; Amzallag, A; Greninger, P; Ли, Д; Kalsy, A; Gomez-Caraballo, M; Elamine, L; Howe, E; Hur, W; Lifshits, E; Robinson, HE; Katayama, R; Faber, AC; Awad, MM; Ramaswamy, S; Mino-Kenudson, M; Iafrate, AJ; Benes, CH; Engelman, JA (2014). "Patient-derived models of acquired resistance can identify effective drug combinations for cancer". Наука. 346 (6216): 1480–1486. Bibcode:2014Sci...346.1480C. Дои:10.1126/science.1254721. ЧВК 4388482. PMID 25394791.
- ^ Palechor-Ceron, N.; Suprynowicz, F. A.; Upadhyay, G.; Dakic, A.; Minas, T.; Simic, V.; Johnson, M .; Albanese, C.; Schlegel, R.; Лю, X. (2013). "Radiation Induces Diffusible Feeder Cell Factor(s) That Cooperate with ROCK Inhibitor to Conditionally Reprogram and Immortalize Epithelial Cells". Американский журнал патологии. 183 (6): 1862–1870. Дои:10.1016/j.ajpath.2013.08.009. ЧВК 5745544. PMID 24096078.
- ^ Saenz, F. R.; Ory, V.; AlOtaiby, M.; Rosenfield, S.; Furlong, M.; Cavalli, L. R.; Johnson, M. D.; Лю, X .; Schlegel, R.; Wellstein, A.; Riegel, A. T. (2014). "Conditionally Reprogrammed Normal and Transformed Mouse Mammary Epithelial Cells Display a Progenitor-Cell–Like Phenotype". PLOS ONE. 9 (5): e97666. Bibcode:2014PLoSO...997666S. Дои:10.1371/journal.pone.0097666. ЧВК 4022745. PMID 24831228.
- ^ Kaur, Sukhbir; Soto-Pantoja, David R.; Stein, Erica V.; и другие. (2013). "Thrombospondin-1 Signaling through CD47 Inhibits Self-renewal by Regulating c-Myc and Other Stem Cell Transcription Factors". Научные отчеты. 3: 1673. Bibcode:2013NatSR...3E1673K. Дои:10.1038/srep01673. ЧВК 3628113. PMID 23591719.
- ^ Soto-Pantoja, D. R.; Ridnour, L. A.; Wink, D. A.; Roberts, D. D. (2013). "Blockade of CD47 increases survival of mice exposed to lethal total body irradiation". Научные отчеты. 3: 1038. Bibcode:2013NatSR...3E1038S. Дои:10.1038/srep01038. ЧВК 3539147. PMID 23301159.
- ^ Gentek, R.; Molawi, K.; Sieweke, M. H. (2014). "Tissue macrophage identity and self-renewal". Иммунологические обзоры. 262 (1): 56–73. Дои:10.1111/imr.12224. PMID 25319327.
- ^ а б Kurian, Leo; Sancho-Martinez, Ignacio; Nivet, Emmanuel; Juan; Izpisua Belmonte, Carlos (2012). "Conversion of human fibroblasts to angioblast-like progenitor cells". Природные методы. 10 (1): 77–83. Дои:10.1038/nmeth.2255. ЧВК 3531579. PMID 23202434.
- ^ Morris, S. A.; Daley, G. Q. (2013). "A blueprint for engineering cell fate: current technologies to reprogram cell identity". Клеточные исследования. 23 (1): 33–48. Дои:10.1038/cr.2013.1. ЧВК 3541663. PMID 23277278.
- ^ Wang, Y. C.; Nakagawa, M.; Garitaonandia, I.; Slavin, I.; Altun, G.; Lacharite, R. M.; Nazor, K. L.; Tran, H.T .; Lynch, C. L.; Leonardo, T. R.; Liu, Y .; Peterson, S. E.; Laurent, L. C.; Yamanaka, S.; Loring, J. F. (2011). "Specific lectin biomarkers for isolation of human pluripotent stem cells identified through array-based glycomic analysis". Клеточные исследования. 21 (11): 1551–1563. Дои:10.1038/cr.2011.148. ЧВК 3364725. PMID 21894191.
- ^ Thomson, J. A .; Itskovitz-Eldor, J; Шапиро, С. С .; Waknitz, M. A.; Swiergiel, J. J.; Marshall, V. S.; Jones, J. M. (1998). «Линии эмбриональных стволовых клеток, полученные из бластоцист человека». Наука. 282 (5391): 1145–1147. Bibcode:1998Научный ... 282.1145Т. Дои:10.1126 / science.282.5391.1145. PMID 9804556.
- ^ Suila, H.; Hirvonen, T .; Ritamo, I.; Natunen, S.; Tuimala, J.; Laitinen, S.; Anderson, H.; Nystedt, J.; Räbinä, J.; Valmu, L. (2014). "Extracellular O-Linked N-Acetylglucosamine is Enriched in Stem Cells Derived from Human Umbilical Cord Blood". BioResearch Open Access. 3 (2): 39–44. Дои:10.1089/biores.2013.0050. ЧВК 3995142. PMID 24804163.
- ^ Perdigoto, C. N.; Bardin, A. J. (2013). "Sending the right signal: Notch and stem cells". Biochimica et Biophysica Acta (BBA) - Общие предметы. 1830 (2): 2307–2322. Дои:10.1016/j.bbagen.2012.08.009. PMID 22917651.
- ^ Jafar-Nejad, H.; Leonardi, J.; Fernandez-Valdivia, R. (2010). "Role of glycans and glycosyltransferases in the regulation of Notch signaling". Гликобиология. 20 (8): 931–949. Дои:10.1093/glycob/cwq053. ЧВК 2912550. PMID 20368670.
- ^ Alisson-Silva, Frederico; Deivid; Rodrigues, Carvalho; Vairo, Leandro; и другие. (2014). "and Adriane R Todeschini (2014). Evidences for the involvement of cell surface glycans in stem cell pluripotency and differentiation". Гликобиология. 24 (5): 458–468. Дои:10.1093/glycob/cwu012. PMID 24578376.
- ^ Hasehira, K.; Tateno, H.; Onuma, Y.; Ито, Й .; Asashima, M.; Hirabayashi, J. (2012). "Structural and Quantitative Evidence for Dynamic Glycome Shift on Production of Induced Pluripotent Stem Cells". Молекулярная и клеточная протеомика. 11 (12): 1913–1923. Дои:10.1074/mcp.M112.020586. ЧВК 3518133. PMID 23023295.
- ^ Becker Kojic', Z. A. (2002). "A Novel Human Erythrocyte Glycosylphosphatidylinositol (GPI)-anchored Glycoprotein ACA. Isolation, Purification, Primary Structure Determination, and Molecular Parameters of its Lipid Structure". Журнал биологической химии. 277 (43): 40472–40478. Дои:10.1074/jbc.M202416200. PMID 12167612.
- ^ Becker-Kojić, Z. A.; Ureña-Peralta, J. R.; Saffrich, R; Rodriguez-Jiménez, F. J.; Rubio, M. P.; Rios, P; Romero, A; Ho, A. D.; Stojković, M (2013). "A novel human glycoprotein ACA is an upstream regulator of human hematopoiesis". Вестник экспериментальной биологии и медицины. 155 (4): 536–551. Дои:10.1007/s10517-013-2195-0. PMID 24143385.
- ^ Becker-Kojič, ZA; Ureña-Peralta, JR; Zipančić, I.; Stojkovič, M. (2013). "Activation of surface glycoprotein ACA induced pluripotent hematopoietic progenitor cells" (PDF). Cell Technologies in Biology and Medicine. 9 (2): 85–101.
- ^ Mikkola, M. (2013). Human pluripotent stem cells: glycomic approaches for culturing and characterization. HDL:10138/37669. ISBN 978-952-10-8444-7.
- ^ Zheng, Z., Jian, J., Zhang, X., Zara, J. N., Yin, W., Chiang, M., ... & Soo, C. (2012). Reprogramming of human fibroblasts into multipotent cells with a single ECM proteoglycan, fibromodulin. Biomaterials, 33(24), 5821-5831. PMID 22622142 Дои:10.1016/j.biomaterials.2012.04.049
- ^ Yang, P., Li, C., Lee, M., Marzvanyan, A., Zhao, Z. H., Ting, K., ... & Zheng, Z. (2020). Photopolymerizable hydrogel encapsulated fibromodulin-reprogrammed cells for muscle regeneration. Tissue engineering. Part A. https://doi.org/10.1089/ten.tea.2020.0026
- ^ Zheng, Z., Li, C., Ha, P., Chang, G. X., Yang, P., Zhang, X., ... & Mills, Z. (2019). CDKN2B upregulation prevents teratoma formation in multipotent fibromodulin-reprogrammed cells. Journal of Clinical Investigation, 129(8), 3236-3251. Дои:10.1172/JCI125015 ЧВК 6668700 PMID 31305260
- ^ Redmer, T.; Diecke, S.; Grigoryan, T.; Quiroga-Negreira, A.; Birchmeier, W.; Besser, D. (2011). "E-cadherin is crucial for embryonic stem cell pluripotency and can replace OCT4 during somatic cell reprogramming". Отчеты EMBO. 12 (7): 720–726. Дои:10.1038/embor.2011.88. ЧВК 3128971. PMID 21617704.
- ^ Bedzhov, I.; Alotaibi, H.; Basilicata, M. F.; Ahlborn, K.; Liszewska, E.; Brabletz, T.; Stemmler, M. P. (2013). "Adhesion, but not a specific cadherin code, is indispensable for ES cell and induced pluripotency". Stem Cell Research. 11 (3): 1250–1263. Дои:10.1016/j.scr.2013.08.009. PMID 24036274.
- ^ Su, G.; Zhao, Y .; Wei, J .; Xiao, Z.; Chen, B .; Han, J .; Chen, L .; Guan, J.; Wang, R .; Dong, Q .; Dai, J. (2013). "Direct conversion of fibroblasts into neural progenitor-like cells by forced growth into 3D spheres on low attachment surfaces". Биоматериалы. 34 (24): 5897–5906. Дои:10.1016/j.biomaterials.2013.04.040. PMID 23680365.
- ^ Downing, T. L.; Soto, J.; Morez, C.; Houssin, T.; Fritz, A.; Yuan, F.; Chu, J.; Patel, S.; Schaffer, D. V.; Li, S. (2013). "Biophysical regulation of epigenetic state and cell reprogramming". Материалы Природы. 12 (12): 1154–1162. Bibcode:2013NatMa..12.1154D. Дои:10.1038/nmat3777. PMID 24141451.
- ^ Sun, Yubing; Koh; Aw Yong, Meng; Villa-Diaz, Luis G.; Fu, Jianping (2014). "Hippo/YAP-mediated rigidity-dependent motor neuron differentiation of human pluripotent stem cells". Материалы Природы. 13 (6): 599–604. Bibcode:2014NatMa..13..599S. Дои:10.1038/nmat3945. ЧВК 4051885. PMID 24728461.
- ^ Murray, P .; Prewitz, M.; Hopp, I.; Wells, N.; Zhang, H .; Купер, А .; Parry, K. L.; Short, R.; Antoine, D. J.; Edgar, D. (2013). "The self-renewal of mouse embryonic stem cells is regulated by cell–substratum adhesion and cell spreading". Международный журнал биохимии и клеточной биологии. 45 (11): 2698–2705. Дои:10.1016/j.biocel.2013.07.001. ЧВК 3898852. PMID 23871934.
- ^ Guilak, F.; Коэн, Д. М .; Estes, B. T.; и другие. (2009). "Control of stem cell fate by physical interactions with the extracellular matrix". Стволовая клетка. 5 (1): 17–26. Дои:10.1016/j.stem.2009.06.016. ЧВК 2768283. PMID 19570510.
- ^ Worley, K .; Certo, A.; Wan, L. Q. (2012). "Geometry–Force Control of Stem Cell Fate". BioNanoScience. 3: 43–51. Дои:10.1007/s12668-012-0067-0.
- ^ Caiazzo, Massimiliano; Okawa, Yuya; Ranga, Adrian; Piersigilli, Alessandra; Tabata, Yoji; Lutolf, Matthias P. (2016). "Defined three-dimensional microenvironments boost induction of pluripotency". Материалы Природы. 15 (3): 344–352. Bibcode:2016NatMa..15..344C. Дои:10.1038/nmat4536. PMID 26752655.
- ^ Squeezing cells into stem cells. ScienceDaily, 11 January 2016
- ^ Roy, B., Yuan, L., Lee, Y., Bharti, A., Mitra, A., & Shivashankar, G. V. (2020). Fibroblast rejuvenation by mechanical reprogramming and redifferentiation. Proceedings of the National Academy of Sciences, 117(19), 10131-10141 Дои:10.1073/pnas.1911497117 ЧВК 7229653 PMID 32350144
- ^ Сингх, А .; Suri, S.; Ли, Т .; Chilton, J. M.; Cooke, M. T.; Chen, W.; Fu, J.; Stice, S. L.; Lu, H.; McDevitt, T. C.; García, A. S. J. (2013). "Adhesion strength–based, label-free isolation of human pluripotent stem cells". Природные методы. 10 (5): 438–444. Дои:10.1038/nmeth.2437. ЧВК 3641175. PMID 23563795.
- ^ Wang, Kainan; Degerny, Cindy; Xu, Minghong; Yang, Xiang-Jiao (2009). "YAP, TAZ, and Yorkie: A conserved family of signal-responsive transcriptional coregulators in animal development and human disease". Biochemistry and Cell Biology. 87 (1): 77–91. Дои:10.1139/O08-114. PMID 19234525.
- ^ Ян, С .; Tibbitt, M.W.; Basta, L.; Anseth, K.S. (2014). "Mechanical memory and dosing influence stem cell fate". Материалы Природы. 13 (6): 645–652. Bibcode:2014NatMa..13..645Y. Дои:10.1038/nmat3889. ЧВК 4031270. PMID 24633344.
- ^ Nampe, D.; Tsutsui, H. (2013). "Engineered Micromechanical Cues Affecting Human Pluripotent Stem Cell Regulations and Fate". Журнал автоматизации лабораторий. 18 (6): 482–493. Дои:10.1177/2211068213503156. PMID 24062363.
- ^ Zhang, W.; Duan, S .; Li, Y .; Сюй, X .; Qu, J .; Zhang, W.; Liu, G. H. (2012). "Converted neural cells: Induced to a cure?". Белки и клетки. 3 (2): 91–97. Дои:10.1007/s13238-012-2029-2. ЧВК 4104580. PMID 22410787.
- ^ Ян, Н .; Ng, Y. H.; Pang, Z. P.; Südhof, T. C.; Wernig, M. (2011). "Induced Neuronal Cells: How to Make and Define a Neuron". Стволовая клетка. 9 (6): 517–525. Дои:10.1016/j.stem.2011.11.015. ЧВК 4377331. PMID 22136927.
- ^ Sheng, C.; Zheng, Q .; Wu, J .; Xu, Z .; Sang, L.; Wang, L .; Guo, C .; Zhu, W .; Tong, M.; Liu, L .; Li, W .; Liu, Z. H.; Zhao, X. Y.; Wang, L .; Chen, Z .; Zhou, Q. (2012). "Generation of dopaminergic neurons directly from mouse fibroblasts and fibroblast-derived neural progenitors". Клеточные исследования. 22 (4): 769–772. Дои:10.1038/cr.2012.32. ЧВК 3317566. PMID 22370632.
- ^ а б Maucksch, C.; Firmin, E.; и другие. (2012). "Non-viral generation of neural precursor-like cells from adult human fibroblasts". J Stem Cells Regen Med. 8 (3): 1–9.
- ^ Ring, K. L.; Tong, L. M.; Balestra, M. E.; Javier, R.; Andrews-Zwilling, Y.; Li, G .; Walker, D.; Zhang, W. R.; Kreitzer, A. C.; Huang, Y. (2012). "Direct Reprogramming of Mouse and Human Fibroblasts into Multipotent Neural Stem Cells with a Single Factor". Стволовая клетка. 11 (1): 100–109. Дои:10.1016/j.stem.2012.05.018. ЧВК 3399516. PMID 22683203.
- ^ а б Ченг, Линь; Hu, Wenxiang; Qiu, Binlong; Чжао, Цзянь; Yu, Yongchun; Guan, Wuqiang; Wang, Min; Yang, Wuzhou; Pei, Gang (18 March 2014). "Generation of neural progenitor cells by chemical cocktails and hypoxia". Клеточные исследования. 24 (6): 665–679. Дои:10.1038/cr.2014.32. ЧВК 4042166. PMID 24638034.
- ^ Zhang, Mingliang; Lin, Yuan-Hung; Sun, Yujiao Jennifer; Zhu, Saiyong; Zheng, Jiashun; Liu, Kai; Cao, Nan; Подобно; Хуанг, Ядун; Ding, Sheng (May 2016). "Pharmacological Reprogramming of Fibroblasts into Neural Stem Cells by Signaling-Directed Transcriptional Activation". Стволовая клетка. 18 (5): 653–667. Дои:10.1016/j.stem.2016.03.020. ЧВК 4864020. PMID 27133794.
- ^ Liu, G. H.; Yi, F.; Сузуки, К .; Qu, J .; Belmonte, J. C. I. (2012). "Induced neural stem cells: A new tool for studying neural development and neurological disorders". Клеточные исследования. 22 (7): 1087–1091. Дои:10.1038/cr.2012.73. ЧВК 3391017. PMID 22547025.
- ^ Torper, O.; Pfisterer, U.; Wolf, D. A.; Pereira, M.; Lau, S.; Jakobsson, J.; Bjorklund, A.; Grealish, S.; Parmar, M. (2013). "Generation of induced neurons via direct conversion in vivo". Труды Национальной академии наук. 110 (17): 7038–7043. Bibcode:2013PNAS..110.7038T. Дои:10.1073/pnas.1303829110. ЧВК 3637783. PMID 23530235.
- ^ Niu, W.; Zang, T.; Zou, Y .; Fang, S .; Smith, D.K .; Bachoo, R.; Zhang, C. L. (2013). "In vivo reprogramming of astrocytes to neuroblasts in the adult brain". Природа клеточной биологии. 15 (10): 1164–1175. Дои:10.1038/ncb2843. ЧВК 3867822. PMID 24056302.
- ^ Su, Z.; Niu, W.; Liu, M. L.; Zou, Y .; Zhang, C. L. (2014). "In vivo conversion of astrocytes to neurons in the injured adult spinal cord". Nature Communications. 5: 3338. Bibcode:2014NatCo...5.3338S. Дои:10.1038/ncomms4338. ЧВК 3966078. PMID 24569435.
- ^ Najm, F. J.; Lager, A. M.; Zaremba, A.; Wyatt, K.; Caprariello, A. V.; Factor, D. C.; Karl, R. T.; Maeda, T .; Miller, R. H.; Tesar, P. J. (2013). "Transcription factor–mediated reprogramming of fibroblasts to expandable, myelinogenic oligodendrocyte progenitor cells". Природа Биотехнологии. 31 (5): 426–433. Дои:10.1038/nbt.2561. ЧВК 3678540. PMID 23584611.
- ^ Ян, Н .; Zuchero, J. B.; Ahlenius, H.; Marro, S.; Ng, Y. H.; Vierbuchen, T.; Hawkins, J. S.; Geissler, R.; Barres, B.A .; Wernig, M. (2013). "Generation of oligodendroglial cells by direct lineage conversion". Природа Биотехнологии. 31 (5): 434–439. Дои:10.1038/nbt.2564. ЧВК 3677690. PMID 23584610.
- ^ Xu, C. (2012). "Turning cardiac fibroblasts into cardiomyocytes in vivo". Тенденции в молекулярной медицине. 18 (10): 575–576. Дои:10.1016/j.molmed.2012.06.009. PMID 22770847.
- ^ Fu, J. D.; Stone, N. R.; Liu, L .; Spencer, C. I.; Qian, L.; Hayashi, Y.; Delgado-Olguin, P.; Ding, S .; Bruneau, B. G.; Srivastava, D. (2013). "Direct Reprogramming of Human Fibroblasts toward a Cardiomyocyte-like State". Отчеты о стволовых клетках. 1 (3): 235–247. Дои:10.1016/j.stemcr.2013.07.005. ЧВК 3849259. PMID 24319660.
- ^ Chen, J. X.; Krane, M.; Deutsch, M. -A.; Wang, L .; Rav-Acha, M.; Gregoire, S.; Engels, M. C.; Rajarajan, K.; Karra, R.; Abel, E. D.; Wu, J. C.; Milan, D.; Wu, S. M. (2012). "Inefficient Reprogramming of Fibroblasts into Cardiomyocytes Using Gata4, Mef2c, and Tbx5". Циркуляционные исследования. 111 (1): 50–55. Дои:10.1161/CIRCRESAHA.112.270264. ЧВК 3390172. PMID 22581928.
- ^ Burridge, P. W.; Keller, G.; Gold, J. D.; Wu, J. C. (2012). "Production of De Novo Cardiomyocytes: Human Pluripotent Stem Cell Differentiation and Direct Reprogramming". Стволовая клетка. 10 (1): 16–28. Дои:10.1016/j.stem.2011.12.013. ЧВК 3255078. PMID 22226352.
- ^ Wang, H .; Cao, N.; Spencer, C. I.; Nie, B.; Ma, T .; Xu, T .; Zhang, Y .; Ван, X .; Srivastava, D.; Ding, S. (2014). "Small Molecules Enable Cardiac Reprogramming of Mouse Fibroblasts with a Single Factor, Oct4". Отчеты по ячейкам. 6 (5): 951–960. Дои:10.1016/j.celrep.2014.01.038. ЧВК 4004339. PMID 24561253.
- ^ Карпентер, Л .; Carr, C .; Yang, C. T.; Stuckey, D. J.; Clarke, K.; Watt, S. M. (2012). "Efficient Differentiation of Human Induced Pluripotent Stem Cells Generates Cardiac Cells That Provide Protection Following Myocardial Infarction in the Rat". Стволовые клетки и развитие. 21 (6): 977–986. Дои:10.1089/scd.2011.0075. ЧВК 3315757. PMID 22182484.
- ^ Cao, Nan; Хуанг, Ю; Zheng, Jiashun; Srivastava, Deepak; Ding, Sheng (2016). "Conversion of human fibroblasts into functional cardiomyocytes by small molecules". Наука. 352 (6290): 1216–1220. Bibcode:2016Sci...352.1216C. Дои:10.1126/science.aaf1502. PMID 27127239.
- ^ Scientists Turn Skin Cells into Heart Cells and Brain Cells Using Drugs. Институты Гладстона. News Center
- ^ Yamada, S.; Nelson, T. J.; Kane, G. C.; Martinez-Fernandez, A.; Crespo-Diaz, R. J.; Ikeda, Y .; Perez-Terzic, C.; Terzic, A. (2013). "IPS Cell Intervention Rescues Wall Motion Disparity Achieving Biological Cardiac Resynchronization Post-Infarction". Журнал физиологии. 591 (17): 4335–4349. Дои:10.1113/jphysiol.2013.252288. ЧВК 3779120. PMID 23568891.
- ^ Lian, X.; Hsiao, C.; Wilson, G .; Zhu, K .; Hazeltine, L. B.; Azarin, S. M.; Raval, K. K.; Zhang, J .; Kamp, T. J.; Palecek, S. P. (2012). "Cozzarelli Prize Winner: Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling". Труды Национальной академии наук. 109 (27): E1848–57. Дои:10.1073/pnas.1200250109. ЧВК 3390875. PMID 22645348.
- ^ Willems, E.; Cabral-Teixeira, J.; Schade, D.; Cai, W.; Reeves, P.; Bushway, P. J.; Lanier, M.; Walsh, C .; Kirchhausen, T.; Izpisua Belmonte, J. C.; Cashman, J.; Mercola, M. (2012). "Small Molecule-Mediated TGF-β Type II Receptor Degradation Promotes Cardiomyogenesis in Embryonic Stem Cells". Стволовая клетка. 11 (2): 242–252. Дои:10.1016/j.stem.2012.04.025. ЧВК 3419596. PMID 22862949.
- ^ Lu, T. Y.; Lin, B .; Kim, J .; Салливан, М .; Tobita, K.; Salama, G.; Yang, L. (2013). "Repopulation of decellularized mouse heart with human induced pluripotent stem cell-derived cardiovascular progenitor cells". Nature Communications. 4: 2307. Bibcode:2013NatCo...4.2307L. Дои:10.1038/ncomms3307. PMID 23942048.
- ^ Mohamed, Tamer M. A.; Stone, Nicole R.; Berry, Emily C.; и другие. (2016). "Chemical Enhancement of In Vitro and In Vivo Direct Cardiac Reprogramming". Тираж. 135 (10): 978–995. Дои:10.1161/CIRCULATIONAHA.116.024692. ЧВК 5340593. PMID 27834668.
- ^ Budniatzky, I.; Gepstein, L. (2014). "Concise Review: Reprogramming Strategies for Cardiovascular Regenerative Medicine: From Induced Pluripotent Stem Cells to Direct Reprogramming". Трансляционная медицина стволовых клеток. 3 (4): 448–457. Дои:10.5966/sctm.2013-0163. ЧВК 3973716. PMID 24591731. Архивировано из оригинал на 2014-04-07.
- ^ Cosgrove, B. D.; Gilbert, P. M.; Porpiglia, E.; Mourkioti, F.; Lee, S.P .; Corbel, S. Y.; Blau, H. M. (2014). «Омоложение популяции мышечных стволовых клеток восстанавливает силу травмированных старых мышц». Природа Медицина. 20 (3): 255–264. Дои:10,1038 / нм 3464. ЧВК 3949152. PMID 24531378.
- ^ Sousa-Victor, P.; Gutarra, S.; García-Prat, L.; Rodriguez-Ubreva, J.; Ortet, L.; Ruiz-Bonilla, V.; Jardí, M.; Ballestar, E.; González, S .; Serrano, A. L.; Perdiguero, E.; Muñoz-Cánoves, P. (2014). "Geriatric muscle stem cells switch reversible quiescence into senescence". Природа. 506 (7488): 316–321. Bibcode:2014Natur.506..316S. Дои:10.1038/nature13013. PMID 24522534.
- ^ Hosoyama; и другие. (2014). "and Masatoshi Suzuki (March, 2014). Derivation of Myogenic Progenitors Directly From Human Pluripotent Stem Cells Using a Sphere-Based Culture". Stem Cells Trans Med. 3 (5): 564–574. Дои:10.5966/sctm.2013-0143. ЧВК 4006483. PMID 24657962.
- ^ Zhu, S .; Rezvani, M.; Harbell, J.; Mattis, A. N.; Wolfe, A. R.; Benet, L. Z.; Willenbring, H.; Ding, S. (2014). "Mouse liver repopulation with hepatocytes generated from human fibroblasts". Природа. 508 (7494): 93–97. Bibcode:2014Natur.508...93Z. Дои:10.1038/nature13020. ЧВК 4161230. PMID 24572354.
- ^ Katsuda, Takeshi; Kawamata, Masaki; Hagiwara, Keitaro; Takahashi, Ryou-u; Yamamoto, Yusuke; Camargo, Fernando D.; Ochiya, Takahiro (January 2017). "Conversion of Terminally Committed Hepatocytes to Culturable Bipotent Progenitor Cells with Regenerative Capacity". Стволовая клетка. 20 (1): 41–55. Дои:10.1016/j.stem.2016.10.007. PMID 27840021.
- ^ Abdelalim, E. M.; Bonnefond, A.; Bennaceur-Griscelli, A.; Froguel, P. (2014). "Pluripotent Stem Cells as a Potential Tool for Disease Modelling and Cell Therapy in Diabetes". Обзоры и отчеты о стволовых клетках. 10 (3): 327–337. Дои:10.1007/s12015-014-9503-6. PMID 24577791.
- ^ Hrvatin, S.; O'Donnell, C. W.; Deng, F .; и другие. (2014). "Differentiated human stem cells resemble fetal, not adult, β cells". Труды Национальной академии наук. 111 (8): 3038–3043. Bibcode:2014PNAS..111.3038H. Дои:10.1073/pnas.1400709111. ЧВК 3939927. PMID 24516164.
- ^ Zhu, Saiyong; Russ, Holger A.; Ван, Сяоцзин; Zhang, Mingliang; Ma, Tianhua; Сюй, Дао; Tang, Shibing; Hebrok, Matthias; Ding, Sheng (2016). "Human pancreatic beta-like cells converted from fibroblasts". Nature Communications. 7: 10080. Bibcode:2016NatCo...710080Z. Дои:10.1038/ncomms10080. ЧВК 4729817. PMID 26733021.
- ^ Akinci, E; Banga, A; Tungatt, K; и другие. (2013). "Reprogramming of Various Cell Types to a Beta-Like State by Pdx1, Ngn3 and MafA". PLOS ONE. 8 (11): e82424. Bibcode:2013PLoSO...882424A. Дои:10.1371/journal.pone.0082424. ЧВК 3843737. PMID 24312421.
- ^ Chen, Y. J.; Finkbeiner, S. R.; Weinblatt, D.; Stanger, B. Z. (2014). "De Novo Formation of Insulin-Producing "Neo-β Cell Islets" from Intestinal Crypts". Отчеты по ячейкам. 6 (6): 1046–1058. Дои:10.1016/j.celrep.2014.02.013. ЧВК 4245054. PMID 24613355.
- ^ Hendry, C. E.; Vanslambrouck, J. M.; Ineson, J.; Suhaimi, N.; Takasato, M.; Rae, F.; Little, M. H. (2013). "Direct Transcriptional Reprogramming of Adult Cells to Embryonic Nephron Progenitors". Журнал Американского общества нефрологов. 24 (9): 1424–1434. Дои:10.1681/ASN.2012121143. ЧВК 3752949. PMID 23766537.
- ^ Ксинарис, Христодулос; Бенедетти, Валентина; Риццо, Паола; Аббат, Мауро; Корна, Даниэла; Аззоллини, Надя; Конти, Сара; Unbekandt, Матье; Дэвис, Джейми А .; Мориджи, Марина; Бениньи, Ариэла; Ремуцци, Джузеппе (ноябрь 2012 г.). «Созревание функциональных почечных органоидов, образованных из суспензий эмбриональных клеток». Журнал Американского общества нефрологов. 23 (11): 1857–1868. Дои:10.1681 / ASN.2012050505. ЧВК 3482737. PMID 23085631.
- ^ Инь, Л .; Оганян, В .; Fen Pung, Y .; Delucia, A .; Bailey, E .; Энрик, М .; Стеванов, К .; Kolz, C.L .; Guarini, G .; Чилиан, В. М. (2011). «Индукция сосудистых клеток-предшественников из эндотелиальных клеток стимулирует рост коронарных коллатералей». Циркуляционные исследования. 110 (2): 241–252. Дои:10.1161 / CIRCRESAHA.111.250126. ЧВК 3974272. PMID 22095729.
- ^ Quijada, P .; Toko, H .; Фишер, К. М .; Bailey, B .; Reilly, P .; Хант, К. Д .; Gude, N.A .; Avitabile, D .; Сассман, М.А. (2012). «Сохранение структуры миокарда улучшено за счет разработки Pim-1 клеток костного мозга». Циркуляционные исследования. 111 (1): 77–86. Дои:10.1161 / CIRCRESAHA.112.265207. ЧВК 3398618. PMID 22619278.
- ^ Mohsin, S .; Хан, М .; Toko, H .; Bailey, B .; Коттедж, кл. Т .; Wallach, K .; Nag, D .; Ли, А .; Siddiqi, S .; Lan, F .; Фишер, К. М .; Gude, N .; Quijada, P .; Avitabile, D .; Truffa, S .; Collins, B .; Дембицкий, З .; Wu, J. C .; Сассман, М.А. (2012). «Клетки-предшественники сердца человека, созданные с использованием киназы Pim-I, улучшают восстановление миокарда». Журнал Американского колледжа кардиологии. 60 (14): 1278–1287. Дои:10.1016 / j.jacc.2012.04.047. ЧВК 3461098. PMID 22841153.
- ^ Американская кардиологическая ассоциация (2012, 25 июля). Взрослые стволовые клетки от липосакции используются для создания кровеносных сосудов в лаборатории. ScienceDaily.
- ^ Wang, Z. Z .; Au, P .; Chen, T .; Shao, Y .; Daheron, L.M .; Bai, H .; Арцигиан, М .; Fukumura, D .; Jain, R.K .; Скадден, Д. Т. (2007). «Эндотелиальные клетки, полученные из эмбриональных стволовых клеток человека, образуют прочные кровеносные сосуды in vivo». Природа Биотехнологии. 25 (3): 317–318. Дои:10.1038 / nbt1287. PMID 17322871.
- ^ Samuel, R .; Daheron, L .; Liao, S .; Вардам, Т .; Kamoun, W. S .; Батиста, А .; Buecker, C .; Schafer, R .; Хан, X .; Au, P .; Scadden, D.T .; Duda, D.G .; Fukumura, D .; Джайн, Р. К. (2013). «Создание функционально компетентных и прочных инженерных кровеносных сосудов из индуцированных человеком плюрипотентных стволовых клеток». Труды Национальной академии наук. 110 (31): 12774–12779. Bibcode:2013ПНАС..11012774С. Дои:10.1073 / pnas.1310675110. ЧВК 3732948. PMID 23861493.
- ^ Zangi, L .; Луи, К. О .; von Gise, A .; Ma, Q .; Ebina, W .; Пташек, Л. М .; Später, D .; Xu, H .; Табебордбар, М .; Горбатов, Р .; Sena, B .; Nahrendorf, M .; Briscoe, D.M .; Li, R.A .; Пари, А. Дж .; Росси, Д. Дж .; Pu, W. T .; Чиен, К. Р. (2013). «Модифицированная мРНК управляет судьбой клеток-предшественников сердца и индуцирует регенерацию сосудов после инфаркта миокарда». Природа Биотехнологии. 31 (10): 898–907. Дои:10.1038 / nbt.2682. ЧВК 4058317. PMID 24013197.
- ^ Zeuner, A .; Мартелли, Ф .; Vaglio, S .; Federici, G .; Whitsett, C .; Мильаччо, А. Р. (2012). «Краткий обзор: эритроциты, полученные из стволовых клеток, как новые игроки в переливании крови». Стволовые клетки. 30 (8): 1587–1596. Дои:10.1002 / стержень.1136. ЧВК 3697769. PMID 22644674.
- ^ Hirose, S. I .; Takayama, N .; Накамура, S .; Nagasawa, K .; Очи, К .; Hirata, S .; Yamazaki, S .; Yamaguchi, T .; Оцу, М .; Sano, S .; Takahashi, N .; Sawaguchi, A .; Ито, М .; Като, Т .; Nakauchi, H .; Это, К. (2013). «Иммортализация эритробластов c-MYC и BCL-XL делает возможным производство крупномасштабных эритроцитов из плюрипотентных стволовых клеток человека». Отчеты о стволовых клетках. 1 (6): 499–508. Дои:10.1016 / j.stemcr.2013.10.010. ЧВК 3871399. PMID 24371805.
- ^ Giarratana, M. -C .; Rouard, H .; Dumont, A .; Кигер, Л .; Safeukui, I .; Ле Пеннек, П. -Ю .; Francois, S .; Trugnan, G .; Peyrard, T .; Мари, Т .; Jolly, S .; Hebert, N .; Mazurier, C .; Mario, N .; Harmand, L .; Lapillonne, H .; Devaux, J. -Y .; Дуэ, Л. (2011). «Доказательство принципа переливания эритроцитов, созданных in vitro». Кровь. 118 (19): 5071–5079. Дои:10.1182 / кровь-2011-06-362038. ЧВК 3217398. PMID 21885599.
- ^ Кобари, Л .; Yates, F .; Oudrhiri, N .; Francina, A .; Кигер, Л .; Mazurier, C .; Rouzbeh, S .; El-Nemer, W .; Hebert, N .; Giarratana, M. -C .; Francois, S .; Часовня, А .; Lapillonne, H .; Luton, D .; Bennaceur-Griscelli, A .; Дуэ, Л. (2012). «Плюрипотентные стволовые клетки, индуцированные человеком, могут достигать полного терминального созревания: доказательства in vivo и in vitro на модели эритропоэтической дифференцировки». Haematologica. 97 (12): 1795–1803. Дои:10.3324 / haematol.2011.055566. ЧВК 3590085. PMID 22733021.
- ^ Keerthivasan, G .; Wickrema, A .; Криспино, Дж. Д. (2011). «Энуклеация эритробластов». Stem Cells International. 2011: 1–9. Дои:10.4061/2011/139851. ЧВК 3189604. PMID 22007239.
- ^ Smith, B.W .; Rozelle, S. S .; Leung, A .; Ubellacker, J .; Парки, А .; Nah, S.K .; Французский, D .; Gadue, P .; Monti, S .; Чуй, Д. Х. К .; Steinberg, M. H .; Frelinger, A. L .; Michelson, A.D .; Theberge, R .; McComb, M.E .; Costello, C.E .; Коттон, Д. Н .; Мостославский, Г .; Sherr, D. H .; Мерфи, Дж. Дж. (2013). «Арилуглеводородный рецептор управляет размножением и дифференцировкой гемопоэтических клеток-предшественников». Кровь. 122 (3): 376–385. Дои:10.1182 / blood-2012-11-466722. ЧВК 3716202. PMID 23723449.
- ^ Шах, Сиддхартх; Хуанг, Сяосун; Чэн, Линьчжао (2014). «Основанные на стволовых клетках подходы к производству эритроцитов для переливания». Стволовые клетки Trans Med. 3 (3): 346–355. Дои:10.5966 / sctm.2013-0054. ЧВК 3952923. PMID 24361925.
- ^ а б Руссо, Г. Ф .; Mazurier, C .; Дуэ, Л. (2016). «Культивирование эритроцитов из стволовых клеток: решение настоящих и будущих проблем трансфузионной медицины?». Научная серия ISBT. 11: 111–117. Дои:10.1111 / voxs.12235.
- ^ Giani, F.C .; Fiorini, C .; Wakabayashi, A .; Людвиг, Л. С .; Салем, Р. М .; Jobaliya, C.D .; Го, М. Х. (2016). «Целевое применение генетических вариаций человека может улучшить производство красных кровяных телец из стволовых клеток». Стволовая клетка. 18 (1): 73–78. Дои:10.1016 / j.stem.2015.09.015. ЧВК 4707983. PMID 26607381.
- ^ Fujita, A .; Uchida, N .; Haro-Mora, J. J .; Винклер, Т .; Тисдейл, Дж. (2016). «Экспрессирующие β-глобин дефинитивные клетки-предшественники эритроида, полученные из эмбриональных и индуцированных мешочков, полученных из плюрипотентных стволовых клеток». Стволовые клетки. 34 (6): 1541–1552. Дои:10.1002 / шток.2335. ЧВК 4892957. PMID 26866725.
- ^ Фигейредо, Констанца; Гудева, Лилия; Хорн, Питер А .; Эйз-Веспер, Бритта; Блашик, Райнер; Сельцам, Аксель (23 апреля 2010 г.). «Получение HLA-дефицитных тромбоцитов из гемопоэтических клеток-предшественников». Переливание. 50 (8): 1690–1701. Дои:10.1111 / j.1537-2995.2010.02644.x. PMID 20412529.
- ^ Судзуки Д., Флахоу К., Йошикава Н., Стирблайт И., Хаяси Ю., Савагути А. ... и Мацумото Т. (2020). «Тромбоциты, производные ИПСК, лишенные HLA класса I, инертны по отношению к анти-HLA класса I и иммунитету естественных киллерных клеток». Отчеты о стволовых клетках, 14 (1), 49-59. Дои:10.1016 / j.stemcr.2019.11.011 ЧВК 6962657 PMID 31883921
- ^ Накамура, S .; Takayama, N .; Hirata, S .; Seo, H .; Endo, H .; Очи, К .; Fujita, K. I .; Koike, T .; Harimoto, K. I .; Dohda, T .; Watanabe, A .; Окита, К .; Takahashi, N .; Sawaguchi, A .; Yamanaka, S .; Nakauchi, H .; Nishimura, S .; Это, К. (2014). «Расширяемые клеточные линии мегакариоцитов позволяют получить клинически применимое образование тромбоцитов из индуцированных человеком плюрипотентных стволовых клеток». Стволовая клетка. 14 (4): 535–548. Дои:10.1016 / j.stem.2014.01.011. PMID 24529595.
- ^ Геварт, Седрик; Педерсен, Роджер; Оувеханд, Виллем; Соранцо, Николь; Брилл, Александр; Пономарев, Татьяна; Пэйн, Холли; Pask, Dean C .; Хоббс, Кэтрин М .; Буэ, Гвенаэль; Лампела, Риина; Долби, Аманда; Ву, Вин Хан; Арумугам, Мира; Кользани, Мария; Ховард, Дэниел; Троттер, Мэтью У .; Ян, Инь; Тейссен, Марлоуз; Васкес, Луэлла; Эванс, Аманда; Моро, Томас (7 апреля 2016 г.). «Крупномасштабное производство мегакариоцитов из плюрипотентных стволовых клеток человека с помощью химически определенного подхода прямого программирования». HDL:1810/253670. Цитировать журнал требует
| журнал =(помощь) - ^ Thon, J. N .; Медвец, Д. А .; Карлссон, С. М .; Итальяно, Дж. Э. (2015). «Дорожные препятствия в производстве тромбоцитов для переливания крови». Журнал тромбоза и гемостаза. 13: S55 – S62. Дои:10.1111 / jth.12942. ЧВК 5565795. PMID 26149051.
- ^ Riddell, S. R .; Гринберг, П. Д. (1995). «Принципы адоптивной Т-клеточной терапии вирусных заболеваний человека». Ежегодный обзор иммунологии. 13: 545–586. Дои:10.1146 / annurev.iy.13.040195.002553. PMID 7612234.
- ^ а б Nishimura, T .; Канеко, С .; Кавана-Тачикава, А .; Tajima, Y .; Goto, H .; Zhu, D .; Накаяма-Хосоя, К .; Iriguchi, S .; Uemura, Y .; Shimizu, T .; Takayama, N .; Yamada, D .; Nishimura, K .; Ohtaka, M .; Watanabe, N .; Takahashi, S .; Ивамото, А .; Koseki, H .; Наканиши, М .; Это, К .; Накаучи, Х. (2013). «Создание омоложенных антигенспецифических Т-клеток путем перепрограммирования до плюрипотентности и повторной дифференцировки». Стволовая клетка. 12 (1): 114–126. Дои:10.1016 / j.stem.2012.11.002. PMID 23290140.
- ^ Vizcardo, R .; Masuda, K .; Yamada, D .; Икава, Т .; Shimizu, K .; Fujii, S. I .; Koseki, H .; Кавамото, Х. (2013). «Регенерация человеческих опухолевых антиген-специфичных Т-клеток из ИПСК, полученных из зрелых CD8 + Т-клеток». Стволовая клетка. 12 (1): 31–36. Дои:10.1016 / j.stem.2012.12.006. PMID 23290135.
- ^ а б Lei, F .; Haque, R .; Xiong, X .; Сонг, Дж. (2012). «Направленная дифференцировка индуцированных плюрипотентных стволовых клеток по отношению к Т-лимфоцитам». Журнал визуализированных экспериментов (63): e3986. Дои:10.3791/3986. ЧВК 3389997. PMID 22617911.
- ^ Саделайн, М; Brentjens, R; Ривьер, I (2013). «Основные принципы конструирования рецепторов химерного антигена». Открытие рака. 3 (4): 388–398. Дои:10.1158 / 2159-8290.CD-12-0548. ЧВК 3667586. PMID 23550147.
- ^ Темели, М .; Kloss, C.C .; Ciriello, G .; Федоров, В. Д .; Perna, F .; Gonen, M .; Саделайн, М. (2013). «Генерация опухолевых Т-лимфоцитов человека из индуцированных плюрипотентных стволовых клеток для лечения рака». Природа Биотехнологии. 31 (10): 928–933. Дои:10.1038 / nbt.2678. ЧВК 5722218. PMID 23934177.
- ^ Pilones, K. A .; Aryankalayil, J .; Демария, С. (2012). «Инвариантные NKT-клетки как новые мишени для иммунотерапии солидных опухолей». Клиническая иммунология и иммунология развития. 2012: 1–11. Дои:10.1155/2012/720803. ЧВК 3483734. PMID 23118781.
- ^ Watarai, H .; Yamada, D .; Fujii, S. I .; Taniguchi, M .; Косеки, Х. (2012). «Индуцированная плюрипотентность как потенциальный путь к иммунотерапии рака, опосредованной клетками iNKT». Международный журнал гематологии. 95 (6): 624–631. Дои:10.1007 / s12185-012-1091-0. PMID 22592322.
- ^ Haruta, M .; Tomita, Y .; Юно, А .; Matsumura, K .; Икеда, Т .; Takamatsu, K .; Haga, E .; Коба, Ц .; Nishimura, Y .; Сенджу, С. (2012). «ТАР-дефицитные линии миелоидных клеток человека, полученные из iPS-клеток, как неограниченный источник клеток для антигенпрезентирующих клеток, подобных дендритным клеткам». Генная терапия. 20 (5): 504–513. Дои:10.1038 / gt.2012.59. PMID 22875043.
- ^ Xie, H .; Е, М .; Feng, R .; Граф Т. (2004). «Пошаговое перепрограммирование В-клеток в макрофаги». Клетка. 117 (5): 663–676. Дои:10.1016 / S0092-8674 (04) 00419-2. PMID 15163413.
Bussmann, L.H .; Schubert, A .; Vu Manh, T. P .; Де Андрес, Л .; Desbordes, S.C .; Parra, M .; Циммерманн, Т .; Рапино, Ф .; Rodriguez-Ubreva, J .; Ballestar, E .; Граф Т. (2009). «Надежная и высокоэффективная система репрограммирования иммунных клеток». Стволовая клетка. 5 (5): 554–566. Дои:10.1016 / j.stem.2009.10.004. PMID 19896445. - ^ Hanna, J .; Маркулаки, S .; Schorderet, P .; Кэри, Б. У .; Beard, C .; Wernig, M .; Creyghton, M.P .; Steine, E. J .; Cassady, J. P .; Форман, Р .; Ленгнер, С. Дж .; Dausman, J. A .; Jaenisch, R. (2008). «Прямое перепрограммирование терминально дифференцированных зрелых лимфоцитов В до плюрипотентности». Клетка. 133 (2): 250–264. Дои:10.1016 / j.cell.2008.03.028. ЧВК 2615249. PMID 18423197.
- ^ Ди Стефано, Б .; Sardina, J. L .; Van Oevelen, C .; Collombet, S .; Каллин, Э. М .; Vicent, G.P .; Lu, J .; Thieffry, D .; Беато, М .; Граф Т. (2013). «C / EBPα настраивает В-клетки для быстрого перепрограммирования в индуцированные плюрипотентные стволовые клетки» (PDF). Природа. 506 (7487): 235–239. Дои:10.1038 / природа12885. HDL:10803/283484. PMID 24336202.
- ^ Рапино, Ф .; Роблес, Э. Ф .; Richter-Larrea, J. A .; Каллин, Э. М .; Martinez-Climent, J. A .; Граф Т. (2013). «C / EBPα индуцирует высокоэффективную макрофагальную трансдифференцировку клеточных линий лимфомы и лейкемии и снижает их онкогенность». Отчеты по ячейкам. 3 (4): 1153–1163. Дои:10.1016 / j.celrep.2013.03.003. PMID 23545498.
- ^ Guo, J .; Feng, Y .; Barnes, P .; Huang, F. F .; Idell, S .; Су, Д. М .; Шамс, Х. (2012). «Делеция FoxN1 в медуллярном эпителии тимуса снижает реакцию периферических Т-клеток на инфекцию и имитирует старение». PLOS ONE. 7 (4): e34681. Bibcode:2012PLoSO ... 734681G. Дои:10.1371 / journal.pone.0034681. ЧВК 3326029. PMID 22514652.
- ^ Вс, л .; Guo, J .; Brown, R .; Amagai, T .; Zhao, Y .; Су, Д.-М. (2010). «Снижение экспрессии одного автономного гена эпителиальных клеток ускоряет возрастную инволюцию тимуса». Ячейка старения. 9 (3): 347–357. Дои:10.1111 / j.1474-9726.2010.00559.x. ЧВК 2894280. PMID 20156205.
- ^ Бреденкамп, Николай; Nowell, Craig S .; Блэкберн, К. Клэр (2014). «Регенерация старого тимуса одним фактором транскрипции». Разработка. 141 (8): 1627–1637. Дои:10.1242 / dev.103614. ЧВК 3978836. PMID 24715454.
- ^ О, Дж., Ван, В., Томас, Р., и Су, Д. М. (2020). Омоложение тимуса с помощью индуцированных эпителиальных клеток тимуса (iTEC) из фибробластов со сверхэкспрессией FOXN1 для противодействия воспалению. bioRxiv. https://doi.org/10.1101/2020.03.17.995357
- ^ а б Peng, Y .; Huang, S .; Cheng, B .; Не, X .; Enhe, J .; Feng, C .; Фу, X. (2013). «Мезенхимальные стволовые клетки: революция в терапевтических стратегиях возрастных заболеваний». Обзоры исследований старения. 12 (1): 103–115. Дои:10.1016 / j.arr.2012.04.005. PMID 22569401.
- ^ а б Бибак, К; Kern, S; Kocaömer, A; Ферлик, К; Бугерт, П. (2008). «Сравнение мезенхимальных стромальных клеток из различных тканей человека: костного мозга, жировой ткани и пуповинной крови». Биомедицинские материалы и инженерия. 18 (1 приложение): S71–76. PMID 18334717.
- ^ Ефименко, А .; Джояшвили, Н .; Калинина, Н .; Кочегура, Т .; Акчурин, Р .; Ткачук, В .; Парфёнова Ю. (2013). «Полученные из жировой ткани мезенхимальные стромальные клетки от пожилых пациентов с заболеванием коронарной артерии сохраняют свойства мезенхимальных стромальных клеток, но демонстрируют характеристики старения и имеют пониженный ангиогенный потенциал». Трансляционная медицина стволовых клеток. 3 (1): 32–41. Дои:10.5966 / sctm.2013-0014. ЧВК 3902283. PMID 24353175.
- ^ Штольцинг, А; Джонс, Э; McGonagle, D; Скатт, А (2008). «Возрастные изменения мезенхимальных стволовых клеток костного мозга человека: последствия для клеточной терапии». Механизмы старения и развития. 129 (3): 163–173. Дои:10.1016 / j.mad.2007.12.002. PMID 18241911.
- ^ Эберле, Ирина; Мусульманин, мохсен; Хеншлер, Рейнхард; Канц, Тобиас (2012). «Разработанные MSCS из индивидуальных iPS-клеток». Мезенхимальные стволовые клетки - основы и клиническое применение II. Достижения в области биохимической инженерии / биотехнологии. 130. С. 1–17. Дои:10.1007/10_2012_156. ISBN 978-3-642-37943-7. PMID 22915200.
- ^ Chen, Y. S .; Pelekanos, R.A .; Ellis, R.L .; Horne, R .; Wolvetang, E.J .; Фиск, Н. М. (2012). «Мезенгенная индукция малых молекул индуцированных человеком плюрипотентных стволовых клеток для создания мезенхимальных стволовых / стромальных клеток». Трансляционная медицина стволовых клеток. 1 (2): 83–95. Дои:10.5966 / sctm.2011-0022. ЧВК 3659681. PMID 23197756.
- ^ Хайнс, К; Мениканин, D; Хан, Дж; Марино, В; Мрозик, К; Gronthos, S; Бартольд, П. М. (2013). «Мезенхимальные стволовые клетки из iPS-клеток способствуют регенерации пародонта». Журнал стоматологических исследований. 92 (9): 833–839. Дои:10.1177/0022034513498258. PMID 23884555.
- ^ Хайнс, Ким; Гронтос, Стэн; Бартольд, П. Марк (1 марта 2014 г.). «ИПСК для регенерации тканей зубов». Текущие отчеты о здоровье полости рта. 1 (1): 9–15. Дои:10.1007 / s40496-013-0001-8.
- ^ Лай, Пей-Лунь; Линь, Сюань; Чен, Шан-Фу; Ян, Шан-Чжи; Хун, Го-Сюань; Чанг, Чинг-Фанг; Чанг, Сян-И; Ли Лу, Фрэнк; Ли, И-Сюань; Лю Юй-Чуань; Хуанг, Сяо-Чун; Лу, Жан (2017). «Эффективное создание химически индуцированных мезенхимальных стволовых клеток из дермальных фибробластов человека». Научные отчеты. 7: 44534. Bibcode:2017НатСР ... 744534Л. Дои:10.1038 / srep44534. ЧВК 5356011. PMID 28303927.
- ^ Лай, Руэнн Чай; Йео, Ронн Ви Йе; Тан, Сун Сим; Чжан, Бинь; Инь, Ицзюнь; Сзе, Ньюман Сиу Кван; Чу, Андре; Лим, Сай Кианг (2013). «Экзосомы мезенхимальных стволовых клеток: Будущая терапия на основе МСК?». Терапия мезенхимальными стволовыми клетками. С. 39–61. Дои:10.1007/978-1-62703-200-1_3. ISBN 978-1-62703-199-8.
Lai, R.C .; Yeo, R. W. Y .; Tan, K. H .; Лим, С. К. (2013). «Экзосомы для доставки лекарств - новое приложение для мезенхимальных стволовых клеток». Достижения биотехнологии. 31 (5): 543–551. Дои:10.1016 / j.biotechadv.2012.08.008. PMID 22959595.
Kosaka, N .; Такешита, Ф .; Yoshioka, Y .; Hagiwara, K .; Кацуда, Т .; Ono, M .; Очия, Т. (2013). «Экзосомальные опухолевые супрессивные микроРНК как новая терапия рака». Расширенные обзоры доставки лекарств. 65 (3): 376–382. Дои:10.1016 / j.addr.2012.07.011. PMID 22841506. - ^ Джумабай, М .; Abdmaulen, R .; Ly, A .; Cubberly, M. R .; Шахмирян, Л. Дж .; Гейдархан-Хагвалл, С .; Dumesic, D. A .; Yao, Y .; Бостром, К. И. (2014). «Плюрипотентные стволовые клетки, полученные из зрелых адипоцитов мышей и человека». Трансляционная медицина стволовых клеток. 3 (2): 161–171. Дои:10.5966 / sctm.2013-0107. ЧВК 3925054. PMID 24396033.
- ^ Poloni, A .; Maurizi, G .; Леони, П .; Серрани, Ф .; Mancini, S .; Frontini, A .; Zingaretti, M.C .; Siquini, W .; Sarzani, R .; Чинти, С. (2012). «Дедифференцированные адипоциты человека проявляют свойства, аналогичные свойствам мезенхимальных стволовых клеток, полученных из костного мозга». Стволовые клетки. 30 (5): 965–974. Дои:10.1002 / стержень.1067. PMID 22367678.
- ^ Shen, J. F .; Сугавара, А; Ямасита, Дж; Огура, H; Сато, S (2011). «Дедифференцированные жировые клетки: альтернативный источник взрослых мультипотентных клеток из жировой ткани». Международный журнал оральной науки. 3 (3): 117–124. Дои:10.4248 / IJOS11044. ЧВК 3470092. PMID 21789960.
- ^ Melief, S.M .; Zwaginga, J. J .; Fibbe, W. E .; Рулофс, Х. (2013). «Мультипотентные стромальные клетки, полученные из жировой ткани, обладают более высокой иммуномодулирующей способностью, чем их аналоги, полученные из костного мозга». Трансляционная медицина стволовых клеток. 2 (6): 455–463. Дои:10.5966 / sctm.2012-0184. ЧВК 3673757. PMID 23694810.
- ^ Cheng, A .; Hardingham, T. E .; Кимбер, С. Дж. (2013). «Восстановление хряща из плюрипотентных стволовых клеток». Тканевая инженерия, часть B: обзоры. 20 (4): 257–66. Дои:10.1089 / ten.teb.2012.0757. ЧВК 4123466. PMID 23957872.
- ^ Outani, H .; Окада, М .; Ямасита, А .; Nakagawa, K .; Yoshikawa, H .; Цумаки, Н. (2013). «Прямая индукция хондрогенных клеток из культуры дермальных фибробластов человека с помощью определенных факторов». PLOS ONE. 8 (10): e77365. Bibcode:2013 ПРИЛОЖЕНИЕ ... 877365O. Дои:10.1371 / journal.pone.0077365. ЧВК 3797820. PMID 24146984.
- ^ а б Crompton, J.G .; Rao, M .; Рестифо, Н. П. (2013). «Воспоминания о реинкарнированной Т-клетке». Стволовая клетка. 12 (1): 6–8. Дои:10.1016 / j.stem.2012.12.009. ЧВК 6352969. PMID 23290132.
- ^ Tan, H. -K .; Toh, C. -X. D .; Сумасшедший.; Ян, Б .; Лю, Т. М .; Lu, J .; Wong, C. -W .; Тан, Т. -К .; Li, H .; Syn, C .; Tan, E. -L .; Lim, B .; Lim, Y. -P .; Cook, S.A .; Ло, Ю. -Х. (2014). «Плюрипотентные стволовые клетки, индуцированные уколом пальца человека, способствуют развитию банка стволовых клеток». Трансляционная медицина стволовых клеток. 3 (5): 586–598. Дои:10.5966 / sctm.2013-0195. ЧВК 4006490. PMID 24646489.
- ^ Окита, К .; Yamakawa, T .; Matsumura, Y .; Sato, Y .; Amano, N .; Watanabe, A .; Goshima, N .; Яманака, С. (2013). «Эффективный невирусный метод для создания свободных от интеграции индуцированных человеком плюрипотентных стволовых клеток из пуповинной крови и клеток периферической крови». Стволовые клетки. 31 (3): 458–466. Дои:10.1002 / стержень.1293. PMID 23193063.
- ^ Geti, I .; Ормистон, М. Л .; Rouhani, F .; Тошнер, М .; Movassagh, M .; Nichols, J .; Mansfield, W .; Southwood, M .; Брэдли, А .; Rana, A. A .; Vallier, L .; Моррелл, Н. В. (2012). «Практичный и эффективный клеточный субстрат для создания индуцированных плюрипотентных стволовых клеток у взрослых: эндотелиальные клетки-предшественники, полученные из крови». Трансляционная медицина стволовых клеток. 1 (12): 855–865. Дои:10.5966 / sctm.2012-0093. ЧВК 3659672. PMID 23283547.
- ^ Staerk, J .; Dawlaty, M. M .; Gao, Q .; Maetzel, D .; Hanna, J .; Sommer, C.A .; Мостославский, Г .; Яениш Р. (2010). «Перепрограммирование клеток периферической крови человека в индуцированные плюрипотентные стволовые клетки». Стволовая клетка. 7 (1): 20–24. Дои:10.1016 / j.stem.2010.06.002. ЧВК 2917234. PMID 20621045.
- ^ Парк, Т. С .; Huo, J. S .; Peters, A .; Talbot, C.C .; Verma, K .; Циммерлин, Л .; Каплан, И. М .; Замбидис, Э. Т. (2012). «Активированные фактором роста цепи стволовых клеток и стромальные сигналы совместно ускоряют неинтегрированное перепрограммирование ИПСК миелоидных предшественников человека». PLOS ONE. 7 (8): e42838. Bibcode:2012PLoSO ... 742838P. Дои:10.1371 / journal.pone.0042838. ЧВК 3414503. PMID 22905176.
- ^ Yoshikawa, K .; Naitoh, M .; Kubota, H .; Ishiko, T .; Aya, R .; Yamawaki, S .; Сузуки, С. (2013). «Мультипотентные стволовые клетки эффективно собираются из кожи щек взрослого человека». Сообщения о биохимических и биофизических исследованиях. 431 (1): 104–110. Дои:10.1016 / j.bbrc.2012.12.069. HDL:2433/174817. PMID 23268344.
- ^ Чжоу, Т .; Бенда, С .; Duzinger, S .; Huang, Y .; Li, X .; Li, Y .; Guo, X .; Cao, G .; Chen, S .; Hao, L .; Chan, Y. -C .; Ng, K. -M .; Сай Хо, Дж .; Wieser, M .; Wu, J .; Redl, H .; Це, Х. -Ф .; Grillari, J .; Grillari-Voglauer, R .; Pei, D .; Эстебан, М.А. (2011). «Создание индуцированных плюрипотентных стволовых клеток из мочи». Журнал Американского общества нефрологов. 22 (7): 1221–1228. Дои:10.1681 / ASN.2011010106. ЧВК 3137570. PMID 21636641.
- ^ Чжоу, Т .; Бенда, С .; Dunzinger, S .; Huang, Y .; Ho, J.C .; Yang, J .; Wang, Y .; Zhang, Y .; Zhuang, Q .; Li, Y .; Бао, X .; Tse, H. F .; Grillari, J .; Grillari-Voglauer, R .; Pei, D .; Эстебан, М.А. (2012). «Создание индуцированных человеком плюрипотентных стволовых клеток из образцов мочи». Протоколы природы. 7 (12): 2080–2089. Дои:10.1038 / nprot.2012.115. PMID 23138349.
- ^ Wang, L .; Wang, L .; Huang, W .; Вс, ч .; Xue, Y .; Вс, З .; Liao, B .; Wang, H .; Бао, X .; Qin, D .; He, J .; Wu, W .; Итак, К. Ф .; Pan, G .; Пей, Д. (2012). «Генерация невральных клеток-предшественников без интеграции из клеток в моче человека». Природные методы. 10 (1): 84–89. Дои:10.1038 / nmeth.2283. HDL:10397/32665. PMID 23223155.
- ^ Цай, Джинглей; Чжан, Яньмэй; Лю, Пэнфэй; Чен, Шубин; Ву, Сюань; Солнце, Юхуа; Ли, Анг; Хуанг, Кэ; Ло, Жунпин; Ван, Лихуэй; Лю, Инь; Чжоу, Тин; Вэй, Шичэн; Пан, Гуанцзинь; Пей, Дуаньцин (2013). «Создание зубоподобных структур из свободных от интеграции плюрипотентных стволовых клеток, индуцированных мочой человека». Регенерация клеток. 2 (1): 2:6. Дои:10.1186/2045-9769-2-6. ЧВК 4230506. PMID 25408878.
- ^ Bharadwaj, S .; Лю, G .; Shi, Y .; Wu, R .; Ян, Б .; Он, Т .; Fan, Y .; Лу, X .; Чжоу, X .; Liu, H .; Атала, А .; Rohozinski, J .; Чжан, Ю. (2013). «Мультипотенциальная дифференцировка стволовых клеток, полученных из мочи человека: потенциал для терапевтического применения в урологии». Стволовые клетки. 31 (9): 1840–1856. Дои:10.1002 / шток.1424. PMID 23666768.
- ^ Wang, Y .; Liu, J .; Tan, X .; Li, G .; Gao, Y .; Лю, X .; Zhang, L .; Ли, Ю. (2012). «Индуцированные плюрипотентные стволовые клетки из мезенхимальных стволовых клеток волосяного фолликула человека». Обзоры и отчеты о стволовых клетках. 9 (4): 451–460. Дои:10.1007 / s12015-012-9420-5. ЧВК 3742959. PMID 23242965.
- ^ Schnabel, L.V .; Abratte, C.M .; Schimenti, J.C .; Southard, T. L .; Фортье, Л. А. (2012). «Генетический фон влияет на индуцированную генерацию плюрипотентных стволовых клеток». Исследования стволовых клеток и терапия. 3 (4): 30. Дои:10.1186 / scrt121. ЧВК 3580468. PMID 22862934.
- ^ Panopoulos, A.D .; Ruiz, S .; Yi, F .; Herrerías, A.D .; Батчелдер, Э. М .; Бельмонте, Дж. К. И. (2011). «Быстрое и высокоэффективное создание индуцированных плюрипотентных стволовых клеток из эндотелиальных клеток пупочной вены человека». PLOS ONE. 6 (5): e19743. Bibcode:2011PLoSO ... 619743P. Дои:10.1371 / journal.pone.0019743. ЧВК 3095638. PMID 21603572.
- ^ а б Polo, J.M .; Liu, S .; Фигероа, М. Э .; Kulalert, W .; Eminli, S .; Tan, K. Y .; Apostolou, E .; Stadtfeld, M .; Li, Y .; Shioda, T .; Natesan, S .; Пари, А. Дж .; Melnick, A .; Evans, T .; Хохедлингер, К. (2010). «Тип клеток влияет на молекулярные и функциональные свойства плюрипотентных стволовых клеток, индуцированных мышами». Природа Биотехнологии. 28 (8): 848–855. Дои:10.1038 / nbt.1667. ЧВК 3148605. PMID 20644536.
- ^ Миура, К .; Okada, Y .; Aoi, T .; Окада, А .; Takahashi, K .; Окита, К .; Nakagawa, M .; Koyanagi, M .; Tanabe, K .; Охнуки, М .; Ogawa, D .; Ikeda, E .; Окано, H .; Яманака, С. (2009). «Изменение безопасности индуцированных линий плюрипотентных стволовых клеток». Природа Биотехнологии. 27 (8): 743–745. Дои:10.1038 / nbt.1554. HDL:2433/120546. PMID 19590502.
- ^ Liang, Y .; Zhang, H .; Feng, Q. S .; Cai, M. B .; Deng, W .; Qin, D .; Yun, J. P .; Tsao, G. S. W .; Канг, Т .; Эстебан, М. А .; Pei, D .; Цзэн, Ю. X. (2013). «Склонность к онкогенезу в индуцированных человеком плюрипотентных стволовых клетках связана с геномной нестабильностью». Китайский журнал рака. 32 (4): 205–212. Дои:10.5732 / cjc.012.10065. ЧВК 3845575. PMID 22704487.
- ^ Kim, K .; Doi, A .; Wen, B .; Ng, K .; Zhao, R .; Cahan, P .; Kim, J .; Aryee, M. J .; Ji, H .; Эрлих, Л. И. Р .; Ябуучи, А .; Takeuchi, A .; Cunniff, K. C .; Hongguang, H .; McKinney-Freeman, S .; Naveiras, O .; Юн, Т. Дж .; Irizarry, R.A .; Jung, N .; Seita, J .; Hanna, J .; Murakami, P .; Jaenisch, R .; Weissleder, R .; Оркин, С. Х .; Weissman, I. L .; Файнберг, А. П .; Дейли, Г.К. (2010). «Эпигенетическая память в индуцированных плюрипотентных стволовых клетках». Природа. 467 (7313): 285–290. Bibcode:2010Натура.467..285K. Дои:10.1038 / природа09342. ЧВК 3150836. PMID 20644535.
- ^ Kim, K .; Zhao, R .; Doi, A .; Ng, K .; Unternaehrer, J .; Cahan, P .; Hongguang, H .; Loh, Y. H .; Aryee, M. J .; Lensch, M. W .; Li, H .; Коллинз, Дж. Дж .; Файнберг, А. П .; Дейли, Г.К. (2011). «Тип донорских клеток может влиять на эпигеном и потенциал дифференцировки индуцированных человеком плюрипотентных стволовых клеток». Природа Биотехнологии. 29 (12): 1117–1119. Дои:10.1038 / nbt.2052. ЧВК 3357310. PMID 22119740.
- ^ Бар-Нур, О .; Russ, H.A .; Efrat, S .; Бенвенисти, Н. (2011). «Эпигенетическая память и предпочтительная дифференцировка клонов в индуцированных плюрипотентных стволовых клетках, полученных из бета-клеток панкреатических островков». Стволовая клетка. 9 (1): 17–23. Дои:10.1016 / j.stem.2011.06.007. PMID 21726830.
- ^ Денкер, Х. В. (2012). «Пора пересмотреть стратегии индукции стволовых клеток». Клетки. 1 (4): 1293–1312. Дои:10.3390 / ячейки1041293. ЧВК 3901125. PMID 24710555.
дальнейшее чтение
- Скуделлари, Меган (2016). «Как iPS-клетки изменили мир». Природа. 534 (7607): 310–312. Bibcode:2016Натура.534..310S. Дои:10.1038 / 534310a. PMID 27306170.
- Турксен, Курсад; Надь, Андраш (2016). Индуцированные плюрипотентные стволовые (iPS) клетки. Методы молекулярной биологии. 1357. Дои:10.1007/978-1-4939-3055-5. ISBN 978-1-4939-3054-8.
- Шривастава, Дипак; ДеВитт, Натали (сентябрь 2016 г.). «Перепрограммирование клеток in vivo: новое поколение». Клетка. 166 (6): 1386–1396. Дои:10.1016 / j.cell.2016.08.055. PMID 27490631.
- Шривастава, Дипак; ДеВитт, Натали (сентябрь 2016 г.). «Перепрограммирование клеток in vivo: новое поколение». Клетка. 166 (6): 1386–1396. Дои:10.1016 / j.cell.2016.08.055. ЧВК 6234007. PMID 27610565.
- Табар, В .; Студер, Л. (2014). «Плюрипотентные стволовые клетки в регенеративной медицине: проблемы и недавний прогресс». Природа Обзоры Генетика. 15 (2): 82–92. Дои:10.1038 / nrg3563. ЧВК 4539940. PMID 24434846.
- Тан, юань; Оои, Сара; Ван, Лишэн (31 декабря 2013 г.). «Иммуногенность и онкогенность плюрипотентных стволовых клеток и их производных: генетические и эпигенетические перспективы». Текущие исследования стволовых клеток и терапия. 9 (1): 63–72. Дои:10.2174 / 1574888x113086660068. ЧВК 3873036. PMID 24160683.
- Яманака, Шинья (июнь 2012 г.). «Индуцированные плюрипотентные стволовые клетки: прошлое, настоящее и будущее». Стволовая клетка. 10 (6): 678–684. Дои:10.1016 / j.stem.2012.05.005. PMID 22704507.
- Takahashi, K .; Яманака, С. (28 мая 2013 г.). «Индуцированные плюрипотентные стволовые клетки в медицине и биологии». Разработка. 140 (12): 2457–2461. Дои:10.1242 / dev.092551. PMID 23715538.
- Asuelime, Grace E .; Ши, Яньхун (август 2012 г.). «Случай клеточной алхимии: перепрограммирование клонов и его потенциал в регенеративной медицине». Журнал молекулярной клеточной биологии. 4 (4): 190–196. Дои:10.1093 / jmcb / mjs005. ЧВК 3408064. PMID 22371436.
- Ленш, М. Уильям; Mummery, Кристин Л. (июнь 2013 г.). «От кражи огня до клеточного перепрограммирования: научная история, ведущая к Нобелевской премии 2012 года». Отчеты о стволовых клетках. 1 (1): 5–17. Дои:10.1016 / j.stemcr.2013.05.001. ЧВК 3757737. PMID 24052937.
- Чжоу, Ци (октябрь 2013 г.). «Индуцированные плюрипотентные стволовые клетки: текущий прогресс и перспективы на будущее». Геномика, протеомика и биоинформатика. 11 (5): 257–258. Дои:10.1016 / j.gpb.2013.10.001. ЧВК 4357784. PMID 24121127.
- Линь Цзи; Мэй-ронг Ли, Донг-дон Ти; Вэй-дон, Хан (2013). «Преобразование клеточного клона, вызванное микросредой: смещение акцента с внутреннего репрограммирования на внешнее воздействие». Обзоры исследований старения. 12 (1): 29–38. Дои:10.1016 / j.arr.2012.04.002. PMID 22561469.
- Такахаши, К. (2014). «Перепрограммирование клетки». Холодная весна Харб Perspect Biol. 6 (2): a018606. Дои:10.1101 / cshperspect.a018606. ЧВК 3941237. PMID 24492711.
- Нобелевская премия по физиологии и медицине 2012 года присуждена за открытие того, что зрелые клетки могут быть перепрограммированы, чтобы стать плюрипотентными
- Хусейн, Самер М.И.; Надь, Андраш А (2012). «Прогресс в области перепрограммирования: новые факторы, новые стратегии и новые взгляды». Текущее мнение в области генетики и развития. 22 (5): 435–443. Дои:10.1016 / j.gde.2012.08.007. PMID 22959308.
- Чжан, Йемин; Яо, Линь; Ю, Сия; Оу, июнь; Хуэй, Нин; Лю, Шаньжун (31 октября 2014 г.). «Плохая имитация естественного процесса». Клеточный цикл. 11 (24): 4536–4544. Дои:10.4161 / cc.22575. ЧВК 3562298. PMID 23114619.
- Луни, Камилла; Джулитти, Стефано; Серена, Елена; Феррари, Лука; Замбон, Алессандро; Гальяно, Онелия; Giobbe, Giovanni G; Михиелин, Федерика; Кнебель, Себастьян; Босио, Андреас; Эльвассор, Никола (2016). «Высокоэффективное перепрограммирование клеток с помощью микрофлюидики». Nat. Методы. 13 (5): 446–452. Дои:10.1038 / nmeth.3832. PMID 27088312. повышение КПД в 50 раз; небольшие объемы; дифференциация в желаемые клетки на той же платформе
- Mochiduki, Y .; Окита, К. (2012). «Методы генерации iPS-клеток для фундаментальных исследований и клинических приложений». Биотехнологический журнал. 7 (6): 789–797. Дои:10.1002 / biot.201100356. PMID 22378737.
- Мадонна, Розалинда (2012). "Индуцированные человеком плюрипотентные стволовые клетки: в поисках клинического применения". Молекулярная биотехнология. 52 (2): 193–203. Дои:10.1007 / s12033-012-9504-0. PMID 22302314.
- Лоренцо, М .; Fleischer, A .; Бачиллер, Д. (2012). «Создание индуцированных мышами и человеком плюрипотентных стволовых клеток (ИПСК) из первичных соматических клеток». Обзоры и отчеты о стволовых клетках. 9 (4): 435–450. Дои:10.1007 / s12015-012-9412-5. HDL:10261/98876. PMID 23104133.
- Подробные протоколы для перепрограммирования и анализа ИПСК
- Buganim, Y .; Faddah, D.A .; Яениш, Р. (2013). «Механизмы и модели репрограммирования соматических клеток». Природа Обзоры Генетика. 14 (6): 427–439. Дои:10.1038 / nrg3473. ЧВК 4060150. PMID 23681063.
| Источники / типы | |
|---|---|
| Клеточная активность | |
| Статьи по Теме | |
| |