Видообразование - Speciation

Видообразование это эволюционный процесс эволюции популяций, чтобы стать отличными разновидность. Биолог Оратор Ф. Кук ввел термин в 1906 году для кладогенез, разделение родословных, в отличие от анагенез, филетическая эволюция внутри линий.[1][2][3] Чарльз Дарвин был первым, кто описал роль естественный отбор в видообразовании в своей книге 1859 г. О происхождении видов.[4] Он также определил половой отбор как вероятный механизм, но сочли это проблематичным.
В природе существует четыре географических способа видообразования, в зависимости от степени видообразования. население изолированы друг от друга: аллопатрический, периферийный, парапатрический, и сочувствующий. Видообразование также может быть вызвано искусственно через животноводство, сельское хозяйство или лабораторные эксперименты. Ли генетический дрейф вносит незначительный или основной вклад в видообразование, является предметом продолжающихся дискуссий.
Быстрое симпатрическое видообразование может происходить через полиплоидия, например, путем удвоения числа хромосом; результат - потомство, которое немедленно репродуктивно изолированный от родительской популяции. Новые виды также могут быть созданы с помощью гибридизация затем, если естественный отбор благоприятствует гибриду, репродуктивная изоляция.
Историческое прошлое
При решении вопроса о происхождении видов возникают два ключевых вопроса: (1) каковы эволюционные механизмы видообразования и (2) что объясняет обособленность и индивидуальность видов в биоте? Со времен Чарльза Дарвина усилия по пониманию природы видов в первую очередь были сосредоточены на первом аспекте, и сейчас широко признано, что решающим фактором происхождения новых видов является репродуктивная изоляция.[5] Далее мы сосредоточимся на втором аспекте происхождения видов.
Дилемма Дарвина: почему существуют виды?
В О происхождении видов (1859) Дарвин интерпретировал биологическую эволюцию с точки зрения естественного отбора, но был озадачен группированием организмов по видам.[6] Глава 6 книги Дарвина озаглавлена «Трудности теории». Обсуждая эти «трудности», он отметил: «Во-первых, почему, если виды произошли от других видов незаметно тонкими градациями, не везде мы видим бесчисленные переходные формы? Почему не вся природа находится в замешательстве, а не существуют виды, как мы видим. их, четко определенные? " Эту дилемму можно назвать отсутствием или редкостью переходных разновидностей в пространстве обитания.[7]
Еще одна дилемма,[8] К первому относится отсутствие или редкость переходных разновидностей во времени. Дарвин указал, что согласно теории естественного отбора, «бесчисленные переходные формы должны были существовать», и задавался вопросом, «почему мы не находим их встроенными в бесчисленное множество в земную кору». Тот факт, что четко определенные виды действительно существуют в природе как в пространстве, так и во времени, подразумевает, что некоторые фундаментальные особенности естественного отбора действуют для создания и сохранения видов.[6]
Влияние полового размножения на видообразование
Утверждалось, что решение первой дилеммы Дарвина заключается в том, что пересечение половое размножение имеет внутреннюю стоимость редкости.[9][10][11][12][13] Цена редкости возникает следующим образом. Если при градиенте ресурсов эволюционирует большое количество отдельных видов, каждый из которых идеально приспособлен к очень узкой полосе этого градиента, каждый вид обязательно будет состоять из очень небольшого числа членов. Поиск партнера в этих обстоятельствах может вызвать трудности, если многие из соседних особей принадлежат к другим видам. В этих условиях, если размер популяции какого-либо вида случайно увеличится (за счет того или иного соседнего вида, если среда насыщена), это немедленно упростит для его членов поиск сексуальных партнеров. . Представители соседних видов, размеры популяции которых уменьшились, испытывают большие трудности с поиском партнеров и поэтому образуют пары реже, чем более крупные виды. Это имеет эффект снежного кома, когда крупные виды растут за счет более мелких, более редких видов, что в конечном итоге заставляет их вымирание. В конце концов, осталось всего несколько видов, каждый из которых сильно отличается от другого.[9][10][12] Цена редкости включает в себя не только затраты, связанные с невозможностью найти помощника, но и косвенные затраты, такие как стоимость общения при поиске партнера при низкой плотности населения.

Редкость приносит с собой другие затраты. Редкие и необычные черты редко приносят пользу. В большинстве случаев они обозначают (безмолвный ) мутация, что почти наверняка будет вредным. Следовательно, сексуальным созданиям надлежит избегать партнеров, обладающих редкими или необычными чертами (койнофилия ).[15][16] Таким образом, сексуальные популяции быстро теряют редкие или периферические фенотипические особенности, тем самым канализируя весь внешний вид, как показано на прилагаемой иллюстрации Африканский карликовый зимородок, Ispidina picta. Это единообразие всех взрослых представителей одного полового вида стимулировало распространение полевые гиды на птиц, млекопитающих, рептилий, насекомых и многих других таксоны, в котором вид может быть описан одной иллюстрацией (или двумя, в случае половой диморфизм ). Как только популяция станет столь же однородной по внешнему виду, как это типично для большинства видов (что показано на фотографии африканского карликового зимородка), ее представители будут избегать спаривания с представителями других популяций, которые внешне отличаются от них самих.[17] Таким образом, избегание самок, демонстрирующих редкие и необычные фенотипические особенности, неизбежно приводит к репродуктивной изоляции, одному из признаков видообразования.[18][19][20][21]
В противоположном случае организмов, которые размножаться бесполым путем, нет стоимости редкости; следовательно, мелкомасштабная адаптация дает только преимущества. Таким образом, бесполые организмы очень часто демонстрируют непрерывное изменение формы (часто во многих различных направлениях), которое Дарвин ожидал от эволюции, превращая их в классификацию «видов» (вернее, морфовиды ) очень сложно.[9][15][16][22][23][24]
Режимы

Все формы естественного видообразования имели место в течение эволюция; Тем не менее, продолжаются дебаты относительно относительной важности каждого механизма в управлении автомобилем. биоразнообразие.[25]
Одним из примеров естественного видообразования является разнообразие трехиглая колюшка, а морской рыба, которая после последний ледниковый период, претерпел видообразование в новые пресная вода колонии в изолированных озерах и ручьях. По оценкам, на протяжении 10 000 поколений у колючек структурные различия больше, чем у разных роды рыб, включая изменения в плавниках, изменения количества или размера их костных пластинок, изменчивую структуру челюстей и различия в цвете.[26]
Аллопатрический
Во время аллопатрической (от древнегреческого аллос, "другое" + Патра, «отечество») видообразование, популяция разделяется на две географически изолированные популяции (например, путем фрагментация среды обитания из-за географических изменений, таких как горное образование ). Затем изолированные популяции подвергаются генотипической или фенотипический расхождения как: (а) они подвергаются разным селективный давления; (б) они независимо подвергаются генетическому дрейфу; (c) разные мутации возникают в двух популяциях. Когда популяции снова вступают в контакт, они развиваются так, что репродуктивно изолированы и больше не способны обмениваться гены. Островная генетика это термин, связанный с тенденцией небольших изолированных генетических пулов производить необычные черты. Примеры включают островная карликовость и радикальные изменения в некоторых известных цепочках островов, например на Комодо. В Галапагосские острова особенно известны своим влиянием на Чарльза Дарвина. За пять недель пребывания там он услышал, что Галапагосские черепахи можно было определить по острову, и заметил, что зяблики различались от одного острова к другому, но только девять месяцев спустя он подумал, что такие факты могут показать, что виды изменчивы. Когда он вернулся в Англия, его предположения об эволюции углубились после того, как эксперты сообщили ему, что это отдельные виды, а не просто разновидности, и, как известно, все другие разные птицы Галапагосских островов были видами зябликов. Хотя зяблики были менее важны для Дарвина, более недавние исследования показали, что птицы, ныне известные как Зяблики Дарвина быть классическим случаем адаптивного эволюционного излучения.[27]
Перипатрический
При перипатрическом видообразовании, подформе аллопатрического видообразования, новые виды образуются в изолированных, более мелких периферических популяциях, которым не дают обмениваться генами с основной популяцией. Это связано с концепцией эффект основателя, поскольку небольшие популяции часто подвергаются узкие места. Часто предполагается, что генетический дрейф играет важную роль в перипатрическом видообразовании.[28][29]
Тематические исследования включают исследование фауны птиц Майром;[30] австралийская птица Петройка многоцветная;[31] и репродуктивная изоляция в популяциях Дрозофила при условии ограничения населения.[нужна цитата ]
Парапатрический
В парапатрическом видообразовании существует лишь частичное разделение зон двух расходящихся популяций, обусловленное географией; особи каждого вида могут время от времени контактировать или пересекать места обитания, но при этом снижается приспособленность гетерозигота приводит к отбору поведения или механизмов, предотвращающих их скрещивание. Парапатрическое видообразование моделируется как непрерывная изменчивость в пределах «единой», связанной среды обитания, действующая как источник естественного отбора, а не на эффектах изоляции местообитаний, возникающих в результате перипатрического и аллопатрического видообразования.[32]
Парапатрическое видообразование может быть связано с дифференциально-зависимым отбор. Даже если есть поток генов между двумя популяциями сильный дифференциальный отбор может препятствовать ассимиляции, и в конечном итоге могут развиться разные виды.[33] Различия в среде обитания могут быть более важными в развитии репродуктивной изоляции, чем время изоляции. Кавказские скальные ящерицы Даревская рудис, D. valentini и Д. Порщинский все гибридизировать друг с другом в их гибридная зона; однако гибридизация сильнее между Д. Порщинский и Д. рудис, которые отделились раньше, но живут в аналогичных местообитаниях, чем между D. valentini и два других вида, которые разделились позже, но живут в разных климатических средах обитания.[34]
Экологи называют[требуется разъяснение ] парапатрическое и перипатрическое видообразование с точки зрения экологические ниши. Для успеха нового вида должна быть доступна ниша. Кольцевые виды Такие как Ларус Считается, что чайки иллюстрируют процесс видообразования, хотя ситуация может быть более сложной.[35] Трава Anthoxanthum odoratum может начинать парапатрическое видообразование в районах минного заражения.[36]
Симпатрический

Симпатрическое видообразование - это образование двух или более видов-потомков от одного предкового вида, занимающих одно и то же географическое положение.
Часто цитируемые примеры симпатрического видообразования обнаруживаются у насекомых, которые становятся зависимыми от различных хозяин растения в той же местности.[37][38]

Самый известный пример симпатрического видообразования - это цихлиды из Восточная Африка населяющий Озера рифтовой долины, особенно Озеро Виктория, Озеро Малави и Озеро Танганьика. Существует более 800 описанных видов, и, по оценкам, в регионе может быть более 1600 видов. Их эволюция приводится как пример того, как естественный и половой отбор.[39][40] Исследование 2008 года предполагает, что симпатрическое видообразование произошло у Пещерные саламандры Теннесси.[41] Симпатрическое видообразование, обусловленное экологическими факторами, может также объяснить необычайное разнообразие ракообразных, обитающих в глубинах Сибири. озеро Байкал.[42]
Окулировочное видообразование было предложено как особая форма симпатрического видообразования, при которой небольшие группы особей становятся все более изолированными от предков, размножаясь преимущественно друг с другом. Этот тип видообразования может быть обусловлен сочетанием различных преимуществ инбридинга, таких как выражение благоприятных рецессивных фенотипов, снижение рекомбинационной нагрузки и снижение стоимости секса.[43]

Муха боярышник (Rhagoletis pomonella ), также известная как яблочная муха личинки, похоже, претерпевает симпатрическое видообразование.[44] Разные популяции боярышника питаются разными фруктами. Особая популяция появилась в Северной Америке в XIX веке спустя некоторое время после яблоки, неместный вид. Эта популяция, питающаяся яблоками, обычно питается только яблоками, а не исторически предпочитаемыми фруктами. боярышник. Нынешняя популяция боярышника обычно не питается яблоками. Некоторые доказательства, такие как шесть из тринадцати аллозим loci различны, мухи боярышника созревают позже и созревают дольше, чем яблони; и то, что существует мало свидетельств межпородного скрещивания (исследователи задокументировали частоту гибридизации 4–6%), позволяет предположить, что имеет место симпатрическое видообразование.[45]
Методы отбора
Армирование

Армирование, иногда называемое Эффект Уоллеса, это процесс, посредством которого естественный отбор увеличивает репродуктивную изоляцию.[18] Это может произойти после того, как две популяции одного и того же вида разделятся, а затем снова войдут в контакт. Если их репродуктивная изоляция была полной, то они уже превратились бы в два отдельных несовместимых вида. Если их репродуктивная изоляция будет неполной, то дальнейшее спаривание между популяциями приведет к гибридам, которые могут быть, а могут и не быть фертильными. Если гибриды бесплодны или плодовиты, но менее приспособлены, чем их предки, тогда произойдет дальнейшая репродуктивная изоляция и по существу произойдет видообразование (например, как в лошади и ослы ).[46]
Причина этого заключается в том, что если каждый из родителей гибридного потомства естественным образом выбрал черты для своих определенных условий, гибридное потомство будет иметь черты от обоих, поэтому не будет соответствовать ни экологической нише, а также любому из родителей. Низкая приспособленность гибридов заставит отбор отдать предпочтение ассортативная вязка, который будет контролировать гибридизацию. Иногда это называют эффектом Уоллеса по имени биолога-эволюциониста. Альфред Рассел Уоллес который предположил в конце 19 века, что это может быть важным фактором видообразования.[47]
И наоборот, если потомство гибридов более приспособлено, чем их предки, то популяции снова сольются в один и тот же вид в пределах области, с которой они контактируют.
Подкрепление, благоприятствующее репродуктивной изоляции, необходимо как для парапатрического, так и для симпатрического видообразования. Без подкрепления географическая область контакта между различными формами одного и того же вида, называемая их «гибридной зоной», не превратится в границу между разными видами. Гибридные зоны - это регионы, где встречаются и скрещиваются различные популяции. В этих регионах очень распространено гибридное потомство, которое обычно создается разошедшимися видами, попадающими в вторичный контакт. Без подкрепления эти два вида имели бы неконтролируемые инбридинг. Подкрепление может быть вызвано в экспериментах по искусственному отбору, как описано ниже.
Экологический
Экологический отбор - это «взаимодействие людей с окружающей их средой во время приобретения ресурсов».[48] Естественный отбор неотъемлемо вовлечен в процесс видообразования, посредством чего «в условиях экологического видообразования популяции в различных средах или популяции, эксплуатирующие разные ресурсы, испытывают противоположное давление естественного отбора на черты, которые прямо или косвенно вызывают эволюцию репродуктивной изоляции».[49] Доказательства роли, которую играет экология в процессе видообразования, существуют. Исследования популяций колюшки подтверждают наличие экологически связанного видообразования, возникающего как побочный продукт.[50] наряду с многочисленными исследованиями параллельного видообразования, где изоляция развивается между независимыми популяциями видов, адаптирующихся к контрастирующим средам, чем между независимыми популяциями, адаптирующимися к аналогичным условиям.[51] Экологическое видообразование происходит с большим количеством доказательств, «... накопленных в результате нисходящих исследований адаптации и репродуктивной изоляции».[51]
Половой отбор
Широко признано, что половой отбор мог вызвать видообразование во многих кладах, независимо от естественный отбор.[52] Однако термин «видообразование» в этом контексте обычно используется в двух различных, но не исключающих друг друга смыслах. Первое и наиболее часто используемое значение относится к «рождению» новых видов. То есть разделение существующего вида на два отдельных вида или отпочкование нового вида от родительского вида, оба движимые биологической «модной причудой» (предпочтение какой-либо особенности или особенностей одного или обоих полов, не обязательно обладающих адаптивными качествами).[52][53][54][55] Во втором смысле «видообразование» относится к широко распространенной тенденции половых существ группироваться в четко определенные виды,[56][19] вместо того, чтобы формировать континуум фенотипы как во времени, так и в пространстве - что было бы более очевидным или логическим следствием естественного отбора. Это действительно было признано Дарвин как проблемные, и включены в его О происхождении видов (1859 г.) под заголовком «Трудности с теорией».[6] Есть несколько предложений, как выбор партнера может сыграть значительную роль в решении Дилемма Дарвина.[19][9][15][16][17][57] Если видообразование происходит в отсутствие естественного отбора, его можно назвать неэкологическое видообразование[58][59].
Искусственное видообразование


Новые виды созданы животноводство, но даты и методы зарождения таких видов не ясны. Часто домашний аналог дикого предка все еще может скрещиваться и производить плодовитое потомство, как в случае домашних животных. крупный рогатый скот, которые можно рассматривать как один и тот же вид, что и несколько разновидностей диких бык, гаур, як и т. д. или отечественные овца которые могут скрещиваться с муфлон.[60][61]
Наиболее документированные создания новых видов в лаборатории были выполнены в конце 1980-х годов. Уильям Р. Райс и Джордж У. Солт разводили Drosophila melanogaster плодовые мошки используя лабиринт с тремя различными вариантами среды обитания, такими как свет / темнота и влажность / сухость. Каждое поколение было помещено в лабиринт, и группы мух, вышедших из двух из восьми выходов, были разделены для размножения друг с другом в своих соответствующих группах. После тридцати пяти поколений две группы и их потомство были репродуктивно изолированы из-за их сильных предпочтений в среде обитания: они спаривались только в тех областях, которые они предпочитали, и поэтому не спаривались с мухами, которые предпочитали другие области.[62] История таких попыток описана Райс и Элен Э. Хостерт (1993).[63][64]Дайан Додд использовала лабораторный эксперимент, чтобы показать, как репродуктивная изоляция может развиваться в Drosophila pseudoobscura плодовых мушек через несколько поколений, помещая их в разные среды, на основе крахмала и мальтозы.[65]

Многим другим людям было легко воспроизвести эксперимент Додда, в том числе с другими видами плодовых мух и другими продуктами питания.[66] Исследования 2005 года показали, что эта быстрая эволюция репродуктивной изоляции на самом деле может быть следствием инфекции, вызванной Вольбахия бактерии.[67]
С другой стороны, эти наблюдения согласуются с представлением о том, что сексуальные существа по своей природе не хотят спариваться с людьми, внешний вид или поведение которых отличается от нормы. Риск того, что такие отклонения вызваны наследственная дезадаптация очень высокий. Таким образом, если сексуальное существо, неспособное предсказать будущее направление естественного отбора, настроено производить наиболее приспособленное потомство, оно будет избегать самок с необычными привычками или особенностями.[68][69][15][16][17] Тогда сексуальные существа неизбежно будут группироваться в репродуктивно изолированные виды.[16]
Генетика
Было обнаружено несколько генов видообразования. Обычно они связаны с процессом усиления поздних стадий видообразования. В 2008 году сообщалось о гене видообразования, вызывающем репродуктивную изоляцию.[70] Это вызывает гибридное бесплодие между родственными подвидами. Порядок видообразования трех групп от общего предка может быть неясным или неизвестным; собрание из трех таких видов называется «трихотомией».
Видообразование через полиплоидию

Полиплоидия это механизм, который вызвал множество событий быстрого видообразования в сочувствие потому что потомство, например, от тетраплоидных х диплоидных скрещиваний часто приводит к триплоидному стерильному потомству.[71] Однако не все полиплоиды репродуктивно изолированы от родительских растений, и поток генов все еще может происходить, например, через триплоидные гибридные х диплоидные спаривания, которые производят тетраплоиды, или спаривания между мейотически нередуцированный гаметы из диплоидов и гаметы из тетраплоидов (см. также гибридное видообразование ).
Было высказано предположение, что многие из существующих видов растений и животных подверглись полиплоидизации в своей эволюционной истории.[72][73] Размножение успешных полиплоидных видов иногда бывает бесполым путем партеногенез или же апомиксис, так как по неизвестным причинам многие бесполые организмы полиплоидны. Известны редкие случаи полиплоидных млекопитающих, но чаще всего они приводят к внутриутробной смерти.
Гибридное видообразование
Гибридизация двух разных видов иногда приводит к различию фенотип. Этот фенотип также может быть более подходящим, чем родительская линия, и поэтому естественный отбор может затем благоприятствовать этим особям. В конце концов, если репродуктивная изоляция будет достигнута, это может привести к появлению отдельного вида. Однако добиться репродуктивной изоляции между гибридами и их родителями особенно сложно, и поэтому видообразование гибридов считается чрезвычайно редким явлением. В Марианская кряква считается, что он возник в результате гибридного видообразования.
Гибридизация - важный способ видообразования растений, поскольку полиплоидия (имея более двух экземпляров каждого хромосома ) переносится растениями легче, чем животными.[74][75] Полиплоидия важна для гибридов, поскольку она позволяет воспроизводить два разных набора хромосом, каждый из которых может спариваться с идентичным партнером во время мейоза.[73] Полиплоиды также обладают большим генетическим разнообразием, что позволяет им избегать инбридинговая депрессия в небольших популяциях.[76]
Гибридизация без изменения числа хромосом называется видообразованием гомоплоидного гибрида. Считается очень редким, но был показан в Геликоний бабочки[77] и подсолнухи. Полиплоидное видообразование, которое включает изменение числа хромосом, является более распространенным явлением, особенно у видов растений.
Транспозиция генов
Феодосий Добжанский, который изучал плодовых мушек на заре генетических исследований в 1930-х годах, предположил, что части хромосом, которые переключаются из одного места в другое, могут вызвать разделение вида на два разных вида. Он наметил, каким образом участки хромосом могут перемещаться в геноме. Эти подвижные секции могут вызывать бесплодие у межвидовых гибридов, что может оказывать давление на видообразование. Теоретически его идея была здравой, но ученые долго спорили, действительно ли это произошло в природе. В конце концов было показано, что конкурирующая теория, предполагающая постепенное накопление мутаций, встречается в природе так часто, что генетики в значительной степени отклонили гипотезу движущегося гена.[78] Однако исследования 2006 года показывают, что переход гена с одной хромосомы на другую может способствовать рождению новых видов.[79] Это подтверждает механизм репродуктивной изоляции, ключевой компонент видообразования.[80]
Тарифы
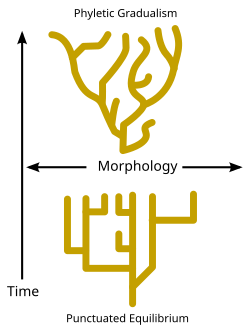
Есть дебаты относительно скорости, с которой происходят события видообразования в течение геологического времени. В то время как некоторые эволюционные биологи утверждают, что события видообразования оставались относительно постоянными и постепенными с течением времени (известные как «филетический градуализм» - см. Диаграмму), некоторые палеонтологи Такие как Найлз Элдридж и Стивен Джей Гулд[81] утверждали, что виды обычно остаются неизменными в течение длительных периодов времени, и что видообразование происходит только в течение относительно коротких интервалов; эта точка зрения известна как прерывистое равновесие. (См. Диаграмму и Дилемма Дарвина.)
Прерывистая эволюция
Эволюция может быть чрезвычайно быстрой, как показано в создании одомашненных животных и растений за очень короткий геологический промежуток времени, охватывающий всего несколько десятков тысяч лет. Кукуруза (Zea Mays), например, был созданный в Мексика всего за несколько тысяч лет, примерно от 7000 до 12000 лет назад.[82] Это поднимает вопрос, почему долгосрочная скорость эволюции намного медленнее, чем это теоретически возможно.[83][84][85][86]







Эволюция навязывается видам или группам. В некоторых странах это не планируется и не предпринимается. Ламаркист путь.[87] Мутации, от которых зависит процесс, являются случайными событиями, и, за исключением "тихие мутации "которые не влияют на функциональность или внешний вид носителя, поэтому обычно являются невыгодными, и их шансы оказаться полезными в будущем исчезающе малы. Таким образом, хотя вид или группа могут выиграть от способности адаптироваться к новому окружающей среде за счет накопления широкого диапазона генетических вариаций, это в ущерб отдельные лица которые должны нести эти мутации до тех пор, пока небольшое, непредсказуемое меньшинство из них в конечном итоге не внесет вклад в такую адаптацию. Таким образом способность для развития потребуется групповой выбор, концепция, дискредитированная (например) Джордж К. Уильямс,[88] Джон Мейнард Смит[89] и Ричард Докинз[90][91][92][93] как выборочно невыгодный для человека.
Таким образом, решение второй дилеммы Дарвина может быть следующим:
Если сексуальные особи оказываются в невыгодном положении из-за передачи мутаций своему потомству, они будут избегать мутантных партнеров со странными или необычными характеристиками.[69][15][16][57] Мутации, влияющие на внешний вид их носителей, редко передаются следующему и последующим поколениям. Поэтому они редко будут проверяться естественным отбором. Таким образом, эволюция фактически останавливается или значительно замедляется. Единственные мутации, которые могут накапливаться в популяции, с точки зрения прерывистого равновесия, - это мутации, которые не оказывают заметного влияния на внешний вид и функциональность своих носителей (т. Е. Они «молчаливы» илинейтральные мутации, "которые могут использоваться и используются для отслеживания родство и возраст популяций и видов.[15][94]) Этот аргумент подразумевает, что эволюция может происходить только в том случае, если мутантных спариваний нельзя избежать в результате острой нехватки потенциальных партнеров. Скорее всего, это произойдет в небольшие изолированные сообщества. Чаще всего они встречаются на небольших островах, в отдаленных долинах, озерах, речных системах или пещерах.[95] или во время последствий массовое вымирание.[94] В этих обстоятельствах не только сильно ограничен выбор партнеров, но и узкие места популяции, эффекты основателя, генетический дрейф и инбридинг вызывают быстрые, случайные изменения в генетическом составе изолированной популяции.[95] Более того, гибридизация с родственными видами, заключенными в один и тот же изолят, может внести дополнительные генетические изменения. Если изолированное население, такое как эта переживает свои генетические потрясения, и впоследствии расширится в незанятую нишу или в нишу, в которой он имеет преимущество перед своими конкурентами, возникнет новый вид или подвид. С геологической точки зрения это будет резкое событие. Возобновление избегания мутантных партнеров после этого снова приведет к эволюционной стагнации.[81][84]
В очевидном подтверждении этого прерываемого равновесия взгляда на эволюцию Окаменелости эволюционный прогресс обычно состоит из видов, которые внезапно появляются и, в конечном итоге, исчезают через сотни тысяч или миллионы лет без каких-либо изменений внешнего вида.[81][94][96] Графически эти ископаемые виды представлены линиями, параллельными оси времени, длина которых показывает, как долго каждый из них существовал. Тот факт, что линии остаются параллельными оси времени, иллюстрирует неизменный внешний вид каждого из ископаемых видов, изображенных на графике. Во время существования каждого вида новые виды появляются через случайные промежутки времени, каждый также существует много сотен тысяч лет, прежде чем исчезнуть без изменения внешнего вида. Точное родство этих параллельных видов, как правило, невозможно определить. Это показано на диаграмме, изображающей распространение видов гомининов во времени так как гоминины отделены от линии, которая привела к эволюции наших ближайших живых родственников приматов, шимпанзе.[96]
Подобные эволюционные временные рамки см., Например, в палеонтологическом списке Африканские динозавры, Азиатские динозавры, то Lampriformes и Amiiformes.
Смотрите также
Рекомендации
- ^ Берлоше 1998, п.3
- ^ Кук, Оратор Ф. (30 марта 1906 г.). «Факторы видообразования». Наука. 23 (587): 506–507. Bibcode:1906Sci .... 23..506C. Дои:10.1126 / science.23.587.506. PMID 17789700.
- ^ Кук, Оратор Ф. (ноябрь 1908 г.). «Эволюция без изоляции». Американский натуралист. 42 (503): 727–731. Дои:10.1086/279001.
- ^ Виа, Сара (16 июня 2009 г.). «Естественный отбор в действии во время видообразования». PNAS. 106 (Приложение 1): 9939–9946. Bibcode:2009PNAS..106.9939V. Дои:10.1073 / pnas.0901397106. ЧВК 2702801. PMID 19528641.
- ^ Майр 1982, п. 273
- ^ а б c Дарвин 1859
- ^ Сепкоски, Дэвид (2012). «1. Дилемма Дарвина: палеонтология, летопись окаменелостей и эволюционная теория». Перечитывание летописи окаменелостей: рост палеобиологии как эволюционной дисциплины. Издательство Чикагского университета. С. 9–50. ISBN 978-0-226-74858-0.
Одно из его самых больших опасений заключалось в том, что «неполнота» летописи окаменелостей может быть использована для критики его теории: что очевидные «пробелы» в последовательности окаменелостей могут быть названы отрицательными доказательствами, по крайней мере, для его предположения, что все организмы произошли в результате мелких и постепенных изменений от общего предка.
- ^ Стоуэр, Ханна (2013). «Разрешение дилеммы Дарвина». Природа Обзоры Генетика. 14 (747): 747. Дои:10.1038 / nrg3614. S2CID 45302603.
Практически одновременное появление большинства современных форм телосложения животных во время кембрийского взрыва предполагает короткий интервал быстрой фенотипической и генетической эволюции, которую Дарвин считал слишком быстрой, чтобы ее можно было объяснить естественным отбором.
- ^ а б c d Бернштейн, Харрис; Байерли, Генри К.; Hopf, Frederic A .; и другие. (21 декабря 1985 г.). «Секс и появление видов». Журнал теоретической биологии. 117 (4): 665–690. Дои:10.1016 / S0022-5193 (85) 80246-0. PMID 4094459.
- ^ а б Hopf, Frederic A .; Хопф, Ф. В. (февраль 1985 г.). «Роль эффекта Аллее в упаковке видов». Теоретическая популяционная биология. 27 (1): 27–50. Дои:10.1016/0040-5809(85)90014-0.
- ^ Бернштейн и Бернштейн 1991
- ^ а б Michod 1995
- ^ Michod 1999
- ^ Хоккей, Дин и Райан, 2005 г., стр.176, 193
- ^ а б c d е ж Коэслаг, Йохан Х. (10 мая 1990 г.). «Койнофилия группирует сексуальных существ по видам, способствует застою и стабилизирует социальное поведение». Журнал теоретической биологии. 144 (1): 15–35. Дои:10.1016 / s0022-5193 (05) 80297-8. ISSN 0022-5193. PMID 2200930.
- ^ а б c d е ж Коеслаг, Йохан Х. (21 декабря 1995 г.). «На двигателе видообразования». Журнал теоретической биологии. 177 (4): 401–409. Дои:10.1006 / jtbi.1995.0256. ISSN 0022-5193.
- ^ а б c Poelstra, Jelmer W .; Виджай, Нагарджун; Bossu, Christen M .; и другие. (20 июня 2014 г.). «Геномный ландшафт, лежащий в основе фенотипической целостности перед лицом потока генов у ворон». Наука. 344 (6190): 1410–1414. Bibcode:2014Научный ... 344.1410P. Дои:10.1126 / science.1253226. PMID 24948738. S2CID 14431499.
Фенотипические различия между падалью и серой вороной в зоне гибридизации в Европе вряд ли связаны с ассортативным спариванием.
- Комментарий Мажуванчерри К. Унникришнана и Х. С. Ахилы. - ^ а б Ридли, Марк. «Видообразование - какова роль подкрепления в видообразовании?». Получено 2015-09-07. Адаптирован из Эволюция (2004), 3-е издание (Мальден, Массачусетс: Blackwell Publishing ), ISBN 978-1-4051-0345-9.
- ^ а б c Мэйнард Смит 1989, стр. 275–280
- ^ Майр 1988
- ^ Уильямс 1992, п. 118
- ^ Мейнард Смит, Джон (Декабрь 1983 г.). «Генетика застоя и пунктуации». Ежегодный обзор генетики. 17: 11–25. Дои:10.1146 / annurev.ge.17.120183.000303. PMID 6364957.
- ^ Клэпхэм, Тутин и Варбург 1952 г.
- ^ Грант 1971
- ^ Бейкер, Джейсон М. (июнь 2005 г.). «Адаптивное видообразование: роль естественного отбора в механизмах географического и негеографического видообразования» (PDF). Исследования по истории и философии науки Часть C: Исследования по истории и философии биологических и биомедицинских наук. 36 (2): 303–326. Дои:10.1016 / j.shpsc.2005.03.005. PMID 19260194.
- ^ Кингсли, Дэвид М. (январь 2009 г.). «Разнообразие выявлено: от атомов к чертам». Scientific American. 300 (1): 52–59. Дои:10.1038 / scientificamerican0109-52. PMID 19186749.
- ^ Саллоуэй, Фрэнк Дж. (30 сентября 1982 г.). "The Бигль коллекции зябликов Дарвина (Geospizinae) ». Вестник Британского музея (естественная история), зоология. 43 (2): 49–58.
- ^ Джерри А. Койн; Х. Аллен Орр (2004). Видообразование. Sinauer Associates. п. 105. ISBN 978-0-87893-091-3.
- ^ Лоусон, Люсинда П .; Бейтс, Джон М .; Менегон, Микеле; Загрузчик, Саймон П. (2015). «Расхождение по краям: перипатрическая изоляция в горном шиповатом комплексе тростниковой лягушки». BMC Эволюционная биология. 15 (128): 128. Дои:10.1186 / s12862-015-0384-3. ЧВК 4487588. PMID 26126573.
- ^ Майр 1992, стр. 21–53
- ^ Токеши, М. (1999). Сосуществование видов: экологические и эволюционные перспективы. Оксфорд: Blackwell Science. п. 27. ISBN 0632061464. OCLC 47011551.
- ^ "Видообразование: происхождение новых видов | Изучение науки в Scitable". www.nature.com. Получено 2020-02-16.
- ^ Эндлер 1977
- ^ Тархнишвили, Давид; Мурцхваладзе, Марин; Гавашелишвили, Александр (август 2013). «Видообразование кавказских ящериц: климатическая неоднородность местообитаний важнее времени изоляции». Биологический журнал Линнеевского общества. 109 (4): 876–892. Дои:10.1111 / bij.12092.
- ^ Либерс, Дорит; Книжфф, Питер де; Хельбиг, Андреас Дж. (2004). «Комплекс сельдевых чаек не является кольцевым видом». Proc Biol Sci. 271 (1542): 893–901. Дои:10.1098 / rspb.2004.2679. ЧВК 1691675. PMID 15255043.
- ^ «Парапатрическое видообразование». Калифорнийский университет в Беркли. Получено 3 апреля 2017.
- ^ Федер, Джеффри Л .; Xianfa Xie; Рулл, Хуан; и другие. (3 мая 2005 г.). "Майр, Добжанский и Буш и сложности симпатрического видообразования в Раголетис". PNAS. 102 (Приложение 1): 6573–6580. Bibcode:2005PNAS..102.6573F. Дои:10.1073 / pnas.0502099102. ЧВК 1131876. PMID 15851672.
- ^ Берлоше, Стюарт Х .; Федер, Джеффри Л. (январь 2002 г.). "Симпатрическое видообразование у насекомых-фитофагов: выход за рамки споров?". Ежегодный обзор энтомологии. 47: 773–815. Дои:10.1146 / annurev.ento.47.091201.145312. PMID 11729091. S2CID 9677456.
- ^ Machado, Heather E .; Пыльца, Александр А .; Hofmann, Hans A .; и другие. (Декабрь 2009 г.). «Межвидовое профилирование экспрессии генов на основе сравнительной геномной гибридизации: обзор и новый подход к африканским рыбам-цихлидам». Интегративная и сравнительная биология. 49 (6): 644–659. Дои:10.1093 / icb / icp080. PMID 21665847.
- ^ Фан, Шаохуа; Элмер, Кэтрин Р .; Мейер, Аксель (5 февраля 2012 г.). «Геномика адаптации и видообразования у цихлид: последние достижения и анализ африканских и неотропических линий». Философские труды Королевского общества B. 367 (1587): 385–394. Дои:10.1098 / rstb.2011.0247. ЧВК 3233715. PMID 22201168.
- ^ Niemiller, Matthew L .; Фитцпатрик, Бенджамин М .; Миллер, Брайан Т. (май 2008 г.). "Recent divergence with gene flow in Tennessee cave salamanders (Plethodontidae: Gyrinophilus) inferred from gene genealogies". Молекулярная экология. 17 (9): 2258–2275. Дои:10.1111/j.1365-294X.2008.03750.x. PMID 18410292. S2CID 20761880.
- ^ Martens, Koen (May 1997). "Speciation in ancient lakes". Тенденции в экологии и эволюции. 12 (5): 177–182. Дои:10.1016/S0169-5347(97)01039-2. PMID 21238028.
- ^ Joly, E. (9 December 2011). "The existence of species rests on a metastable equilibrium between inbreeding and outbreeding. An essay on the close relationship between speciation, inbreeding and recessive mutations". Биология Директ. 6: 62. Дои:10.1186/1745-6150-6-62. ЧВК 3275546. PMID 22152499.
- ^ Feder, Jeffrey L.; Roethele, Joseph B.; Filchak, Kenneth; и другие. (Март 2003 г.). "Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella". Генетика. 163 (3): 939–953. ЧВК 1462491. PMID 12663534. Получено 2015-09-07.
- ^ Berlocher, Stewart H.; Bush, Guy L. (June 1982). "An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny". Систематическая зоология. 31 (2): 136–155. Дои:10.2307/2413033. JSTOR 2413033.
- ^ Sætre, Glenn-Peter (2012). "Reinforcement". eLS. Дои:10.1002/9780470015902.a0001754.pub3. ISBN 978-0470016176. Отсутствует или пусто
| название =(помощь) - ^ Ollerton, Jeff (September 2005). "Speciation: Flowering time and the Wallace Effect" (PDF). Heridity. 95 (3): 181–182. Дои:10.1038/sj.hdy.6800718. PMID 16077739. S2CID 13300641. Архивировано из оригинал (PDF) на 2007-06-05. Получено 2015-09-07.
- ^ Howard D. Rundle and Patrik Nosil (2005), "Ecological speciation", Письма об экологии, 8 (3): 336–352, Дои:10.1111 / j.1461-0248.2004.00715.x
- ^ Dolph Schluter (2001), "Ecology and the origin of species", Тенденции в экологии и эволюции, 16 (7): 372–380, Дои:10.1016/S0169-5347(01)02198-X, PMID 11403870
- ^ Jeffrey S. McKinnon; и другие. (2004), "Evidence for ecology's role in speciation", Природа, 429 (6989): 294–298, Bibcode:2004Natur.429..294M, Дои:10.1038/nature02556, PMID 15152252, S2CID 2744267
- ^ а б Dolph Schluter (2009), "Evidence for Ecological Speciation and Its Alternative", Наука, 326 (5915): 737–740, Bibcode:2009Sci...323..737S, Дои:10.1126/science.1160006, PMID 19197053, S2CID 307207
- ^ а б Panhuis, Tami M.; Butlin, Roger; Zuk, Marlene; и другие. (Июль 2001 г.). "Sexual selection and speciation" (PDF). Тенденции в экологии и эволюции. 16 (7): 364–371. Дои:10.1016/s0169-5347(01)02160-7. PMID 11403869.
- ^ Дарвин, Чарльз; А. Р. Уоллес (1858). "On the Tendency of Species to form Varieties; and on the Perpetuation of Varieties and Species by Natural Means of Selection" (PDF). Journal of the Proceedings of the Linnean Society of London. Зоология. 3 (9): 46–50. Дои:10.1111 / j.1096-3642.1858.tb02500.x.
- ^ Дарвин, Чарльз (1859). "4". О происхождении видов (1-е изд.). п. 89.
- ^ Эберхард, В. Г. (1985). Половой отбор и гениталии животных. Издательство Гарвардского университета, Кембридж, Массачусетс
- ^ Gould, Stephen Jay (1980). A Quahog is a Quahog. The Panda's thumb. More reflections in natural history. Нью-Йорк: W. W. Norton & Company. стр.204–213. ISBN 978-0-393-30023-9.
- ^ а б Miller 2013, pp. 177, 395–396
- ^ Rundell, Rebecca J.; Price, Trevor D. (2009-07-01). "Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation". Тенденции в экологии и эволюции. 24 (7): 394–399. Дои:10.1016/j.tree.2009.02.007. ISSN 0169-5347. PMID 19409647.
- ^ Czekanski-Moir, Jesse E.; Rundell, Rebecca J. (2019-05-01). "The Ecology of Nonecological Speciation and Nonadaptive Radiations". Тенденции в экологии и эволюции. 34 (5): 400–415. Дои:10.1016/j.tree.2019.01.012. ISSN 0169-5347. PMID 30824193.
- ^ Новак 1999
- ^ Hiendleder, Stefan; Kaupe, Bernhard; Wassmuth, Rudolf; и другие. (7 мая 2002 г.). "Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies". Труды Королевского общества B. 269 (1494): 893–904. Дои:10.1098/rspb.2002.1975. ЧВК 1690972. PMID 12028771.
- ^ Rice, William R.; Salt, George W. (June 1988). "Видообразование посредством разрушительного отбора по предпочтению среды обитания: экспериментальные данные". Американский натуралист. 131 (6): 911–917. Дои:10.1086/284831.
- ^ Rice, William R.; Hostert, Ellen E. (December 1993). "Laboratory Experiments on Speciation: What Have We Learned in 40 Years?". Эволюция. 47 (6): 1637–1653. Дои:10.2307/2410209. JSTOR 2410209. PMID 28568007.
- ^ Gavrilets, Sergey (Октябрь 2003 г.). "Perspective: Models of Speciation: What Have We Learned in 40 Years?". Эволюция. 57 (10): 2197–2215. Дои:10.1554/02-727. PMID 14628909. S2CID 198158082.
- ^ Dodd, Diane M. B. (September 1989). "Reproductive Isolation as a Consequence of Adaptive Divergence in Drosophila pseudoobscura". Эволюция. 43 (6): 1308–1311. Дои:10.2307/2409365. JSTOR 2409365. PMID 28564510.
- ^ Киркпатрик, Марк; Ravigné, Virginie (March 2002). "Видообразование путем естественного и полового отбора: модели и эксперименты". Американский натуралист. 159 (S3): S22 – S35. Дои:10.1086/338370. ISSN 0003-0147. PMID 18707367. S2CID 16516804.
- ^ Koukou, Katerina; Pavlikaki, Haris; Kilias, George; и другие. (Январь 2006 г.). "Influence of Antibiotic Treatment and Вольбахия Curing on Sexual Isolation Among Drosophila melanogaster Cage Populations". Эволюция. 60 (1): 87–96. Дои:10.1554/05-374.1. PMID 16568634. S2CID 198153238.
- ^ Symons 1979
- ^ а б Langlois, Judith H.; Roggman, Lori A. (March 1990). "Attractive Faces Are Only Average". Психологическая наука. 1 (2): 115–121. Дои:10.1111/j.1467-9280.1990.tb00079.x. S2CID 18557871.
- ^ Phadnis, Nitin; Orr, H. Allen (16 января 2009 г.). "A Single Gene Causes Both Male Sterility and Segregation Distortion in Дрозофила Hybrids". Наука. 323 (5912): 376–379. Bibcode:2009Sci...323..376P. Дои:10.1126/science.1163934. ЧВК 2628965. PMID 19074311.
- ^ Ramsey, Justin; Schemske, Douglas W. (November 1998). "Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants". Ежегодный обзор экологии и систематики. 29: 467–501. Дои:10.1146/annurev.ecolsys.29.1.467.
- ^ Otto, Sarah P.; Whitton, Jeannette (December 2000). "Polyploid Incidence and Evolution" (PDF). Ежегодный обзор генетики. 34: 401–437. CiteSeerX 10.1.1.323.1059. Дои:10.1146/annurev.genet.34.1.401. PMID 11092833.
- ^ а б Comai, Luca (November 2005). «Преимущества и недостатки полиплоидности». Природа Обзоры Генетика. 6 (11): 836–846. Дои:10.1038 / nrg1711. PMID 16304599. S2CID 3329282.
- ^ Wendel, Jonathan F. (January 2000). «Эволюция генома полиплоидов». Молекулярная биология растений. 42 (1): 225–249. Дои:10.1023 / А: 1006392424384. PMID 10688139. S2CID 14856314.
- ^ Sémon, Marie; Wolfe, Kenneth H. (December 2007). «Последствия дупликации генома». Текущее мнение в области генетики и развития. 17 (6): 505–512. Дои:10.1016 / j.gde.2007.09.007. PMID 18006297.
- ^ Солтис, Памела С.; Soltis, Douglas E. (June 20, 2000). «Роль генетических и геномных атрибутов в успехе полиплоидов». PNAS. 97 (13): 7051–7057. Bibcode:2000PNAS ... 97.7051S. Дои:10.1073 / pnas.97.13.7051. ЧВК 34383. PMID 10860970.
- ^ Mavarez, Jesús; Salazar, Camilo A.; Bermingham, Eldredge; и другие. (June 15, 2006). "Speciation by hybridization in Геликоний butterflies". Природа. 441 (7095): 868–871. Bibcode:2006Natur.441..868M. Дои:10.1038/nature04738. PMID 16778888. S2CID 2457445.
- ^ Sherwood, Jonathan (September 8, 2006). "Genetic Surprise Confirms Neglected 70-Year-Old Evolutionary Theory" (Пресс-релиз). Университет Рочестера. Получено 2015-09-10.
- ^ Masly, John P.; Jones, Corbin D.; Mohamed, A. F. Noor; и другие. (8 сентября 2006 г.). "Gene Transposition as a Cause of Hybrid Sterility in Дрозофила". Наука. 313 (5792): 1448–1450. Bibcode:2006Sci...313.1448M. Дои:10.1126/science.1128721. PMID 16960009. S2CID 23462115.
- ^ Minkel, J. R. (September 8, 2006). "Wandering Fly Gene Supports New Model of Speciation". Scientific American. Получено 2015-09-11.
- ^ а б c Gould, Stephen Jay; Элдридж, Найлз (Spring 1977). "Punctuated equilibria: the tempo and mode of evolution reconsidered" (PDF). Палеобиология. 3 (2): 115–151. Дои:10.1017/s0094837300005224. JSTOR 2400177. Архивировано из оригинал (PDF) on 2014-06-24. Получено 2015-09-15.
- ^ Laws 2010, стр. 210–215
- ^ Уильямс 1992, гл. 9
- ^ а б Eldredge & Gould 1972, chpt. 5
- ^ Mayr 1954, стр.157–180
- ^ Maynard Smith 1989, п. 281
- ^ Gould 1980, пт. 4, chpt. 18
- ^ Уильямс 1974
- ^ Мейнард Смит, Джон (March 14, 1964). "Group Selection and Kin Selection". Природа. 201 (4924): 1145–1147. Bibcode:1964Natur.201.1145S. Дои:10.1038/2011145a0. S2CID 4177102.
- ^ Dawkins 1995, chpt. 4
- ^ Докинз, Ричард (Декабрь 1994 г.). "Burying the Vehicle". Поведенческие науки и науки о мозге. 17 (4): 616–617. Дои:10.1017/S0140525X00036207. ISSN 0140-525X. Архивировано из оригинал on 2006-09-15. Получено 2015-09-15. "Remarks on an earlier article by [Elliot] Sober [sic] и Дэвид Слоан Уилсон, who made a more extended argument in their recent book Unto Others : The Evolution and Psychology of Unselfish Behavior"
- ^ Деннет, Дэниел С. (Декабрь 1994 г.). "E Pluribus Unum?". Поведенческие науки и науки о мозге. 17 (4): 617–618. Дои:10.1017/S0140525X00036219. Архивировано из оригинал 27 декабря 2007 г. "Commentary on Wilson & Sober: Group Selection."
- ^ Пинкер, Стивен (18 июня 2012 г.). "The False Allure of Group Selection". edge.org. Edge Foundation, Inc. Получено 2015-09-15.
- ^ а б c Кэмпбелл 1990, pp. 450–451, 487–490, 499–501
- ^ а б Ayala 1982, pp. 73–83, 182–190, 198–215
- ^ а б McCarthy & Rubidge 2005
Библиография
- Ayala, Francisco J. (1982). Population and Evolutionary Genetics. Benjamin/Cummings Series in the Life Sciences. Menlo Park, CA: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-0315-5. LCCN 81021623. OCLC 8034790.
- Berlocher, Stewart H. (1998). "Origins: A Brief History of Research on Speciation". In Howard, Daniel J.; Berlocher, Stewart H. (eds.). Endless Forms: Species and Speciation. Нью-Йорк: Oxford University Press. ISBN 978-0-19-510901-6. LCCN 97031461. OCLC 37545522.
- Бернштейн, Кэрол; Bernstein, Harris (1991). Aging, Sex, and DNA Repair. San Diego, CA: Академическая пресса. ISBN 978-0-12-092860-6. LCCN 90014467. OCLC 22542921.
- Кэмпбелл, Нил А. (1990). Биология (2-е изд.). Redwood City, CA: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-1800-5. LCCN 89017952. OCLC 20352649.
- Clapham, Arthur Roy; Tutin, Thomas G.; Warburg, Edmund F. (1952). Flora of the British Isles. Кембридж, Великобритания: Издательство Кембриджского университета. LCCN 52008880. OCLC 1084058.
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей. LCCN 06017473. OCLC 741260650. The book is available from The Complete Work of Charles Darwin Online. Проверено 12 сентября 2015.
- Докинз, Ричард (1995). River Out of Eden: A Darwinian View of Life. Серия «Мастера науки». Нью-Йорк: Базовые книги. ISBN 978-0-465-01606-8. LCCN 94037146. OCLC 31376584.
- Элдридж, Найлз; Gould, Stephen Jay (1972). "Punctuated Equilibria: An Alternative to Phyletic Gradualism". In Schopf, Thomas J. M. (ed.). Модели в палеобиологии. San Francisco, CA: Freeman Cooper & Co. ISBN 978-0-87735-325-6. LCCN 72078387. OCLC 572084. Перепечатано в Eldredge 1985, pp. 193–223
- Eldredge, Niles (1985). Time Frames: The Rethinking of Darwinian Evolution and the Theory of Punctuated Equilibria. Нью-Йорк: Саймон и Шустер. ISBN 978-0-671-49555-8. LCCN 84023632. OCLC 11443805.
- Endler, John A. (1977). Geographic Variation, Speciation, and Clines. Monographs in Population Biology. 10. Принстон, штат Нью-Джерси: Princeton University Press. С. 1–246. ISBN 978-0-691-08187-8. LCCN 76045896. OCLC 2645720. PMID 409931.
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений в естествознании (1-е изд.). Нью-Йорк: W. W. Norton & Company. ISBN 978-0-393-01380-1. LCCN 80015952. OCLC 6331415.
- Grant, Verne (1971). Вид растений. Нью-Йорк: Columbia University Press. ISBN 978-0-231-03208-7. LCCN 75125620. OCLC 139834.
- Hockey, Phil A. R.; Dean, W. Richard J.; Ryan, Peter G., eds. (2005). Roberts Birds of Southern Africa (7-е изд.). Cape Town, South Africa: Trustees of the J. Voelcker Bird Book Fund. ISBN 978-0-620-34053-3. LCCN 2006376728. OCLC 65978899.
- Laws, Bill (2010). Fifty Plants that Changed the Course of History. Буффало, Нью-Йорк: Книги Светлячка. ISBN 978-1-55407-798-4. LCCN 2011414731. OCLC 711609823.
- Мейнард Смит, Джон (1989). Эволюционная генетика. Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-854215-5. LCCN 88017041. OCLC 18069049.
- Майр, Эрнст (1954). "Change of Genetic Environment and Evolution". В Хаксли, Джулиан; Харди, Алистер С.; Ford, Edmund B. (ред.). Evolution as a Process. Лондон: Аллен и Анвин. LCCN 54001781. OCLC 974739.
- Майр, Эрнст (1982). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Кембридж, Массачусетс: Belknap Press, издательство Harvard University Press. ISBN 978-0-674-36445-5. LCCN 81013204. OCLC 7875904.
- Mayr, Ernst (1988). Toward a New Philosophy of Biology: Observations of an Evolutionist. Кембридж, Массачусетс: Belknap Press of Harvard University Press. ISBN 978-0-674-89665-9. LCCN 87031892. OCLC 17108004.
- Mayr, Ernst (1992). "Speciational Evolution or Punctuated Equilibrium". In Somit, Albert; Peterson, Steven A. (eds.). Dynamics of Evolution: The Punctuated Equilibrium Debate in the Natural and Social Sciences. Итака, штат Нью-Йорк: Издательство Корнельского университета. ISBN 978-0-8014-9763-6. LCCN 91055569. OCLC 24374091.
- McCarthy, Terence; Rubidge, Bruce (2005). The Story of Earth & Life: A Southern African Perspective on a 4.6-Billion-Year Journey. Cape Town, South Africa: Struik Publishers. ISBN 978-1-77007-148-3. LCCN 2006376206. OCLC 62098231.
- Michod, Richard E. (1995). Eros and Evolution: A Natural Philosophy of Sex. Helix Books. Ридинг, Массачусетс: Эддисон-Уэсли. ISBN 978-0-201-40754-9. LCCN 94013158. OCLC 30625193.
- Michod, Richard E. (1999). Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02699-2. LCCN 98004166. OCLC 38948118.
- Miller, William B., Jr. (2013). The Microcosm Within: Evolution and Extinction in the Hologenome. Бока-Ратон, Флорида: Универсальные издатели. ISBN 978-1-61233-277-2. LCCN 2013033832. OCLC 859168474.
- Новак, Рональд М. (1999). Млекопитающие мира Уокера (6-е изд.). Балтимор, Мэриленд: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-5789-8. LCCN 98023686. OCLC 39045218.
- Саймонс, Дональд (1979). Эволюция человеческой сексуальности. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-502535-4. LCCN 78023361. OCLC 4494283.
- Уильямс, Джордж К. (1974) [Originally published 1966]. Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton Science Library. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02357-1. LCCN 65017164. OCLC 8500898.
- Williams, George C. (1992). Natural Selection: Domains, Levels, and Challenges. Оксфордская серия по экологии и эволюции. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-506933-4. LCCN 91038938. OCLC 228136567.
дальнейшее чтение
- Койн, Джерри А.; Orr, H. Allen (2004). Видообразование. Sunderlands, MA: Sinauer Associates. ISBN 978-0-87893-089-0. LCCN 2004009505. OCLC 55078441.
- Gavrilets, S. (2004). Fitness Landscapes and the Origin of Species. Издательство Принстонского университета. ISBN 978-0691119830.
- Grant, Verne (1981). Вид растений (2-е изд.). Нью-Йорк: Columbia University Press. ISBN 978-0-231-05112-5. LCCN 81006159. OCLC 7552165.
- Marko, Peter B. (2008). "Allopatry". В Jørgensen, Sven Erik; Fath, Brian (eds.). Энциклопедия экологии. 1, A-C (1st ed.). Оксфорд, Великобритания: Эльзевир. С. 131–138. ISBN 978-0-444-52033-3. LCCN 2008923435. OCLC 173240026.
- Майр, Эрнст (1963). Animal Species and Evolution. Кембридж, Массачусетс: Belknap Press, издательство Harvard University Press. ISBN 978-0-674-03750-2. LCCN 63009552. OCLC 899044868.
- Schilthuizen, Menno (2001). Frogs, Flies, and Dandelions: The Making of Species. Оксфорд; Нью-Йорк: Oxford University Press. ISBN 978-0-19-850393-4. LCCN 2001270180. OCLC 46729094.
- Shapiro, J. B.; Leducq, J-B.; Mallet, J. (2016). "What is Speciation?". PLOS Genetics. 12 (3): e1005860. Дои:10.1371/journal.pgen.1005860. ЧВК 4816541. PMID 27030977.
- White, Michael J. D. (1978). Modes of Speciation. A Series of Books in Biology. Сан-Франциско, Калифорния: В. Х. Фриман и компания. ISBN 978-0-7167-0284-9. LCCN 77010955. OCLC 3203453.
внешняя ссылка
- Boxhorn, Joseph (September 1, 1995). "Observed Instances of Speciation". Архив TalkOrigins. Houston, TX: The TalkOrigins Foundation, Inc.
- Hawks, John D. (9 февраля 2005 г.). "Speciation". John Hawks Weblog.
- "Speciation". Калифорнийский университет в Беркли.
| Базовые концепты | ||
|---|---|---|
| Географические режимы | ||
| Изолирующие факторы | ||
| Гибридизация | ||
| Видообразование в таксонах | ||

| Явления |  | ||||
|---|---|---|---|---|---|
| Модели | |||||
| Причины | |||||
| Теории и концепции | |||||
| События вымирания |
| ||||
| Вымершие виды |
| ||||
| Организации | |||||
| Смотрите также | |||||
| |||||