МикроРНК - MicroRNA

А микроРНК (сокращенно miRNA) - небольшой некодирующая РНК молекула (содержащая около 22 нуклеотиды ) найдены в растениях, животных и некоторых вирусах, которые функционируют в Подавление РНК и посттранскрипционный регуляция экспрессии генов.[1] miRNA функционируют через спаривание оснований с дополнительными последовательностями внутри мРНК молекулы.[2] В результате эти молекулы мРНК становятся замолчать одним или несколькими из следующих процессов: (1) расщепление цепи мРНК на две части, (2) дестабилизация мРНК за счет укорочения ее поли (А) хвост и (3) Менее эффективный перевод мРНК в белки рибосомы.[2][3]
миРНК напоминают малые интерферирующие РНК (миРНК) из РНК-интерференция (РНКи) пути, за исключением того, что миРНК происходят из областей транскриптов РНК, которые складываются сами по себе, образуя короткие шпильки, тогда как миРНК происходят из более длинных областей двухцепочечная РНК.[4] В человеческий геном может кодировать более 1900 miRNA,[5] хотя более свежий анализ показывает, что это число ближе к 600.[6]
miRNAs в изобилии во многих типах клеток млекопитающих[7][8] и, по-видимому, нацелены на около 60% генов человека и других млекопитающих.[9][10] Многие miRNA эволюционно консервативны, что означает, что они выполняют важные биологические функции.[6][1] Например, 90 семейств miRNA были законсервированы, по крайней мере, со времен общего предка млекопитающих и рыб, и большинство из этих консервативных miRNAs имеют важные функции, как показали исследования, в которых были выбиты гены для одного или нескольких членов семейства. у мышей.[1]
История
Первая миРНК была открыта в начале 1990-х годов.[11] Однако miRNA не были признаны отдельным классом биологических регуляторов до начала 2000-х годов.[12][13][14][15][16] Исследование miRNA выявило различные наборы miRNA, экспрессируемые в разных типах клеток и ткани[8][17] и множественные роли miRNA в развитии растений и животных и во многих других биологических процессах.[18][19][20][21][22][23][24] Аберрантная экспрессия miRNA вовлечена в болезненные состояния. Терапия на основе MiRNA находится на стадии исследования.[25][26][27][28]
Первая miRNA была открыта в 1993 г. группой во главе с Амброс и в том числе Ли и Фейнбаум. Однако для дополнительного понимания механизма его действия потребовались одновременно опубликованные работы Рувкун команда, включая Вайтмана и Ха.[11][29] Эти группы последовательно публиковали статьи о лин-4 ген, который, как известно, контролирует время C. elegans личиночное развитие путем подавления лин-14 ген. Когда Ли и др. изолировал лин-4 miRNA, они обнаружили, что вместо того, чтобы производить мРНК, кодирующую белок, она производит короткие некодирующие РНК, одна из которых была ~ 22-нуклеотидной РНК, которая содержала последовательности, частично комплементарные множественным последовательностям в 3 'UTR из лин-14 мРНК.[11] Эта комплементарность была предложена, чтобы препятствовать трансляции лин-14 мРНК в белок LIN-14. В то время лин-4 малая РНК считалась нематода идиосинкразия.
В 2000 году была охарактеризована вторая малая РНК: let-7 РНК, которая подавляет лин-41 способствовать более позднему переходу в развитии C. elegans.[12] В let-7 Было обнаружено, что РНК сохраняется у многих видов, что позволяет предположить, что let-7 РНК и дополнительные «малые временные РНК» могут регулировать время развития у различных животных, включая человека.[13]
Год спустя лин-4 и let-7 Было обнаружено, что РНК являются частью большого класса малых РНК, присутствующих в C. elegans, Дрозофила и человеческие клетки.[14][15][16] Многие РНК этого класса напоминали лин-4 и let-7 РНК, за исключением паттернов их экспрессии, обычно не соответствовали роли в регуляции времени развития. Это предполагает, что большинство из них может функционировать в других типах регуляторных путей. С этого момента исследователи начали использовать термин «микроРНК» для обозначения этого класса малых регуляторных РНК.[14][15][16]
Первым заболеванием человека, связанным с нарушением регуляции miRNA, было хронический лимфолейкоз.
Номенклатура
В соответствии со стандартной системой номенклатуры имена присваиваются экспериментально подтвержденным miRNA перед публикацией.[30][31] За префиксом «miR» следует тире и число, последнее часто указывает порядок именования. Например, miR-124 был назван и, вероятно, обнаружен до miR-456. «MiR-» с заглавной буквы относится к зрелой форме miRNA, а «mir-» без заглавной буквы относится к pre-miRNA и pri-miRNA.[32] Гены, кодирующие миРНК, также называются с использованием того же трехбуквенного префикса в соответствии с соглашениями номенклатуры генов организма. Например, официальные названия генов miRNA у некоторых организмов:мир-1 в C. elegans и Дрозофила, Мир-1 в Rattus norvegicus и МИР-25 в человеке.
miRNA с почти идентичными последовательностями, за исключением одного или двух нуклеотидов, помечаются дополнительной строчной буквой. Например, miR-124a тесно связана с miR-124b. Например:
- hsa-miR-181a: aacauucaACgcugucggugAgu
- hsa-miR-181b: aacauucaUUgcugucggugGgu
Пре-миРНК, при-миРНК и гены, которые приводят к 100% идентичным зрелым миРНК, но которые расположены в разных местах генома, обозначаются дополнительным суффиксом в виде тире. Например, пре-miRNA hsa-mir-194-1 и hsa-mir-194-2 приводят к идентичной зрелой miRNA (hsa-miR-194), но происходят из генов, расположенных в разных областях генома.
Виды происхождения обозначаются трехбуквенным префиксом, например, hsa-miR-124 - это человек (Homo sapiens) miRNA и oar-miR-124 овечья (Овис Овен) miRNA. Другие распространенные префиксы включают «v» для вируса (miRNA, кодируемая вирусным геномом) и «d» для Дрозофила miRNA (плодовая муха, обычно изучаемая в генетических исследованиях).
Когда две зрелые микроРНК происходят из противоположных ветвей одной и той же пре-миРНК и обнаруживаются в примерно одинаковых количествах, они обозначаются суффиксом -3p или -5p. (В прошлом это различие также проводилось с помощью "s" (смысл ) и «как» (антисмысловой)). Однако зрелых микроРНК, обнаруженных на одном плече шпильки, обычно намного больше, чем на другом плече.[4] в этом случае звездочка после названия указывает на зрелые виды, обнаруженные на низких уровнях от противоположного плеча шпильки. Например, miR-124 и miR-124 * имеют общую шпильку пре-miRNA, но в клетке обнаруживается гораздо больше miR-124.
Цели
МикроРНК растений обычно имеют почти идеальное спаривание со своими мРНК-мишенями, что вызывает репрессию генов через расщепление транскриптов-мишеней.[18] Напротив, миРНК животных способны распознавать свои мРНК-мишени, используя всего 6-8 нуклеотидов (затравочная область) на 5'-конце миРНК,[9][33][34] которого недостаточно для того, чтобы вызвать расщепление целевых мРНК.[2] Комбинаторная регуляция - это особенность регуляции miRNA у животных.[2][35] Данная miRNA может иметь сотни различных мишеней мРНК, и данная мишень может регулироваться множеством miRNA.[10][36]
Оценки среднего количества уникальных информационных РНК, которые являются мишенями для репрессии типичной miRNA, варьируются в зависимости от метода оценки.[37] но множественные подходы показывают, что miRNA млекопитающих могут иметь множество уникальных мишеней. Напр., Анализ miRNAs, высококонсервативных у позвоночных, показывает, что каждая имеет в среднем примерно 400 консервативных мишеней.[10] Сходным образом эксперименты показывают, что один вид miRNA может снижать стабильность сотен уникальных информационных РНК.[38] Другие эксперименты показывают, что один вид miRNA может подавлять продукцию сотен белков, но эта репрессия часто бывает относительно мягкой (гораздо меньше, чем в 2 раза).[39][40] Первое заболевание человека, связанное с нарушением регуляции miRNA, было хроническим. лимфоцитарный лейкоз. Последовали и другие В-клеточные злокачественные новообразования.
Биогенез

До 40% генов miRNA могут находиться в интроны или даже экзоны других генов.[41] Обычно, хотя и не исключительно, они обнаруживаются в смысловой ориентации,[42][43] и, таким образом, обычно регулируются вместе с их генами-хозяевами.[41][44][45]
Матрица ДНК - не последнее слово в производстве зрелой миРНК: 6% человеческих миРНК показывают редактирование РНК (IsomiRs ), сайт-специфическая модификация последовательностей РНК для получения продуктов, отличных от продуктов, кодируемых их ДНК. Это увеличивает разнообразие и объем действия miRNA, выходящий за рамки того, что подразумевается только геномом.
Транскрипция
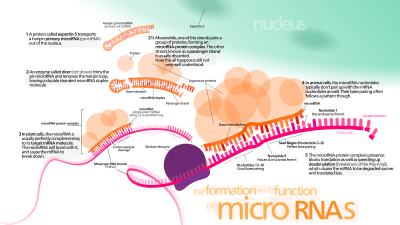
гены miRNA обычно транскрибируются РНК-полимераза II (Pol II).[46][47] Полимераза часто связывается с промотором, обнаруженным рядом с последовательностью ДНК, кодируя то, что станет петлей шпильки пре-миРНК. Итоговая стенограмма закрытый со специально модифицированным нуклеотидом на 5'-конце, полиаденилированный с несколькими аденозины (поли (А) хвост),[46][42] и сращенный. МикроРНК животных первоначально транскрибируются как часть одного плеча ∼80 нуклеотидной РНК. стебель-петля которая, в свою очередь, является частью предшественника miRNA длиной в несколько сотен нуклеотидов, называемого первичной miRNA (pri-miRNA).[46][42] Когда предшественник «стебель-петля» обнаруживается в 3'-UTR, транскрипт может служить в качестве при-миРНК и мРНК.[42] РНК-полимераза III (Pol III) транскрибирует некоторые miRNA, особенно те, которые расположены выше по течению. Последовательности Alu, переносить РНК (тРНК) и промоторные единицы с широким вкраплением повторов (MWIR) млекопитающих.[48]
Ядерная обработка

Одна при-миРНК может содержать от одного до шести предшественников миРНК. Эти структуры петли шпильки состоят примерно из 70 нуклеотидов каждая. Каждая шпилька окружена последовательностями, необходимыми для эффективной обработки.
Структура двухцепочечной РНК (дцРНК) шпилек в при-миРНК распознается ядерным белком, известным как Синдром ДиДжорджи Критическая область 8 (DGCR8 или «Паша» у беспозвоночных), названный в честь его ассоциации с Синдром ДиДжорджи. DGCR8 связывается с ферментом Дроша, белок, который разрезает РНК, чтобы сформировать Микропроцессорный комплекс.[49][50] В этом комплексе DGCR8 ориентирует каталитический домен РНКазы III Drosha для освобождения шпилек от pri-miRNAs путем расщепления РНК примерно на одиннадцать нуклеотидов от основания шпильки (одна спиральная дцРНК превращается в стержень).[51][52] Полученный продукт имеет двухнуклеотидный выступ на 3'-конце; он имеет 3 'гидроксильные и 5' фосфатные группы. Ее часто называют пре-миРНК (миРНК-предшественница). Были идентифицированы мотивы последовательностей ниже пре-miRNA, которые важны для эффективного процессинга.[53][54][55]
Пре-миРНК, которые сращенный непосредственно из интронов, минуя микропроцессорный комплекс, известны как "Миртроны. "Изначально считалось, что существует только в Дрозофила и C. elegansмиртроны теперь обнаружены у млекопитающих.[56]
До 16% пре-миРНК могут быть изменены посредством ядерной Редактирование РНК.[57][58][59] Наиболее часто, ферменты известный как аденозиндезаминазы действуя на РНК (ADAR), катализируют аденозин к инозин (От A к I) переходы. Редактирование РНК может останавливать ядерный процессинг (например, pri-miR-142, что приводит к деградации рибонуклеазой Tudor-SN) и изменять последующие процессы, включая процессинг цитоплазматической miRNA и специфичность мишени (например, путем изменения зародышевой области miR-376 в центральной нервной системе).[57]
Ядерный экспорт

Шпильки пре-миРНК экспортируются из ядра в процессе, включающем ядерно-цитоплазматический челнок. Экспортин-5. Этот белок, входящий в состав кариоферин семейство, распознает двухнуклеотидный выступ, оставленный ферментом РНКазы III Drosha на 3'-конце шпильки пре-миРНК. Опосредованный экспортином-5 транспорт в цитоплазму зависит от энергии, используя GTP привязанный к Ран белок.[60]
Цитоплазматическая обработка
в цитоплазма шпилька пре-миРНК расщепляется ферментом РНКаза III Дайсер.[61] Эта эндорибонуклеаза взаимодействует с 5 'и 3' концами шпильки.[62] и отрезает петлю, соединяющую 3 'и 5' плечи, давая несовершенный дуплекс miRNA: miRNA * длиной около 22 нуклеотидов.[61] Общая длина шпильки и размер петли влияют на эффективность обработки дайсером. Несовершенная природа спаривания miRNA: miRNA * также влияет на расщепление.[61][63] Некоторые из G-богатых пре-миРНК потенциально могут принимать G-квадруплекс структура как альтернатива канонической структуре стержень-петля. Например, человеческая пре-miRNA 92b принимает G-квадруплекс структура, которая устойчива к опосредованному Дайсером расщеплению в цитоплазма.[64] Хотя любая цепь дуплекса потенциально может действовать как функциональная миРНК, только одна цепь обычно включается в РНК-индуцированный комплекс сайленсинга (RISC), где взаимодействуют миРНК и ее мРНК-мишень.
Хотя большинство miRNA расположены внутри клетки, некоторые miRNA, обычно известные как циркулирующие miRNA или внеклеточные miRNA, также были обнаружены во внеклеточной среде, включая различные биологические жидкости и среды для культивирования клеток.[65][66]
Биогенез в растениях
Биогенез miRNA у растений отличается от биогенеза животных, главным образом, стадиями ядерной обработки и экспорта. Вместо того, чтобы расщепляться двумя разными ферментами, один раз внутри ядра и один раз за его пределами, оба расщепления миРНК растения выполняются гомологом Дайсера, называемым Дайсер-подобный1 (DL1). DL1 экспрессируется только в ядре растительных клеток, что указывает на то, что обе реакции происходят внутри ядра. До того, как дуплексы miRNA: miRNA * переносятся из ядра, его 3'-выступы метилируются с помощью РНК метилтрансферазапротеин называется Hua-Enhancer1 (HEN1). Затем дуплекс переносится из ядра в цитоплазму с помощью белка, называемого Hasty (HST), гомолога Exportin 5, где они разбираются, и зрелая миРНК включается в RISC.[67]
РНК-индуцированный комплекс сайленсинга
Зрелая miRNA является частью активного комплекса RNA-индуцированного сайленсинга (RISC), содержащего Dicer и многие связанные с ним белки.[68] RISC также известен как комплекс рибонуклеопротеидов микроРНК (miRNP);[69] RISC со встроенной miRNA иногда называют miRISC.
Считается, что процессинг пре-миРНК дайсером связан с раскручиванием дуплекса. Как правило, в miRISC включается только одна цепь, выбранная на основании ее термодинамической нестабильности и более слабого спаривания оснований на 5'-конце по сравнению с другой цепью.[70][71][72] Положение стержня-петли также может влиять на выбор пряди.[73] Другая цепь, называемая пассажирской цепью из-за ее более низких уровней в устойчивом состоянии, обозначена звездочкой (*) и обычно деградирует. В некоторых случаях обе нити дуплекса жизнеспособны и становятся функциональными miRNA, которые нацелены на разные популяции мРНК.[74]
Члены Аргонавт Семейство белков (Ago) занимает центральное место в функции RISC. Аргонавты необходимы для индуцированного miRNA сайленсинга и содержат два консервативных домена связывания РНК: домен PAZ, который может связывать одноцепочечный 3'-конец зрелой miRNA, и PIWI домен, структурно напоминающий рибонуклеаза-H и функционирует для взаимодействия с 5 'концом направляющей нити. Они связывают зрелую миРНК и ориентируют ее для взаимодействия с целевой мРНК. Некоторые аргонавты, например Ago2 человека, непосредственно расщепляют транскрипты-мишени; аргонавты также могут привлекать дополнительные белки для достижения репрессии трансляции.[75] Геном человека кодирует восемь белков аргонавтов, разделенных по сходству последовательностей на два семейства: AGO (четыре члена присутствуют во всех клетках млекопитающих и называются E1F2C / hAgo у людей) и PIWI (обнаруживаются в зародышевой линии и гематопоэтических стволовых клетках).[69][75]
Дополнительные компоненты RISC включают TRBP [связывающий белок, связывающий трансактивирующую РНК (TAR) вируса иммунодефицита человека (ВИЧ)],[76] ПАКТ (белковый активатор интерферон -индуцированный протеинкиназа ), комплекс SMN, хрупкий X протеин умственной отсталости (FMRP), белок, содержащий домен стафилококковой нуклеазы Tudor (Tudor-SN), предполагаемая ДНК геликаза MOV10, и мотив узнавания РНК, содержащий белок TNRC6B.[60][77][78]
Режим сайленсинга и регуляторные петли
Молчание генов может происходить либо за счет деградации мРНК, либо за счет предотвращения трансляции мРНК. Например, miR16 содержит последовательность, комплементарную AU-богатому элементу, обнаруженному в 3'UTR многих нестабильных мРНК, таких как TNF альфа или GM-CSF.[79] Было продемонстрировано, что при полной комплементарности между микроРНК и последовательностью мРНК-мишени Ago2 может расщеплять мРНК и приводить к прямой деградации мРНК. В отсутствие комплементарности молчание достигается за счет предотвращения трансляции.[38] Связь miRNA и ее мРНК-мишени может быть основана на простой отрицательной регуляции мРНК-мишени, но кажется, что распространенным сценарием является использование «когерентного прямая связь петля »,« петля взаимной отрицательной обратной связи »(также называемая двойной отрицательной петлей) и« петля положительной обратной связи / прямой связи ». Некоторые miRNA работают как буферы случайных изменений экспрессии генов, возникающих из-за стохастических событий в транскрипции, трансляции и стабильности белка. Такая регуляция обычно достигается за счет петель отрицательной обратной связи или некогерентной петли прямой связи, разобщающей выход белка с транскрипцией мРНК.
Оборот
Оборот зрелой miRNA необходим для быстрых изменений в профилях экспрессии miRNA. Считается, что во время созревания miRNA в цитоплазме поглощение белком Argonaute стабилизирует направляющую цепь, тогда как противоположная (* или «пассажирская») цепь предпочтительно разрушается. В том, что было названо стратегией «Используй или потеряй», Argonaute может предпочтительно сохранять miRNAs со многими мишенями, чем miRNAs с небольшим количеством мишеней или без них, что приводит к деградации не нацеленных молекул.[80]
Распад зрелых миРНК в Caenorhabditis elegans опосредуется 5'-к-3 ' экзорибонуклеаза XRN2, также известный как Rat1p.[81] У растений члены семейства SDN (малые РНК-деградирующие нуклеазы) разрушают miRNAs в противоположном (3'-к-5 ') направлении. Подобные ферменты кодируются в геномах животных, но их роль не описана.[80]
Некоторые модификации miRNA влияют на стабильность miRNA. На что указывает работа в модельном организме Arabidopsis thaliana (кресс-салат), миРНК зрелых растений, по-видимому, стабилизируются добавлением метильных групп на 3'-конце. 2'-O-сопряженные метильные группы блокируют добавление урацил (U) остатки по уридилтрансфераза ферменты, модификация, которая может быть связана с деградацией miRNA. Однако уридилирование может также защищать некоторые miRNA; последствия этой модификации до конца не изучены. Сообщалось об уридилировании некоторых микроРНК животных. Как растения, так и животные миРНК могут быть изменены путем добавления остатков аденина (А) к 3'-концу miРНК. В конце слова "млекопитающее" добавлен дополнительный А miR-122, обогащенная печенью миРНК, важная для гепатит С, стабилизирует молекулу, а миРНК растений, заканчивающиеся остатком аденина, имеют более медленную скорость распада.[80]
Клеточные функции

Функция miRNA, по-видимому, заключается в регуляции генов. Для этого miRNA дополнительный к части одного или нескольких информационные РНК (мРНК). MiРНК животных обычно комплементарны сайту в 3 'UTR тогда как миРНК растений обычно комплементарны кодирующим областям мРНК.[83] Идеальное или почти идеальное спаривание оснований с целевой РНК способствует расщеплению РНК.[84] Это основной вид миРНК растений.[85] У животных совпадения несовершенные.
Чтобы частично комплементарные микроРНК распознавали свои мишени, нуклеотиды 2-7 миРНК (ее «затравочная область»)[9][33]) должны идеально дополнять друг друга.[86] МикроРНК животных ингибируют трансляцию белка целевой мРНК[87] (это присутствует, но реже у растений).[85] Частично комплементарные микроРНК также могут ускорить деаденилирование, что приводит к более ранней деградации мРНК.[88] Хотя деградация мРНК, нацеленная на miRNA, хорошо задокументирована, происходит ли репрессия трансляции за счет деградации мРНК, ингибирования трансляции или их комбинации, горячо обсуждается. Последние работы над miR-430 у рыбок данио, а также на bantam-miRNA и miR-9 в Дрозофила культивируемых клеток, показывает, что репрессия трансляции вызвана нарушением инициирование перевода, независимо от деаденилирования мРНК.[89][90]
miRNAs иногда также вызывают гистоновая модификация и Метилирование ДНК из промоутер сайтов, что влияет на экспрессию целевых генов.[91][92]
Девять механизмов действия miRNA описаны и собраны в единую математическую модель:[82]
- Ингибирование инициации Cap-40S;
- 60S ингибирование присоединения рибосомных единиц;
- Замедление удлинения;
- Выпадение рибосом (преждевременное прекращение);
- Ко-трансляционная деградация растущего белка;
- Секвестрация в П-телах;
- распад мРНК (дестабилизация);
- расщепление мРНК;
- Ингибирование транскрипции посредством реорганизации хроматина, опосредованной микроРНК, с последующим подавлением генов.
Эти механизмы часто невозможно различить, используя экспериментальные данные о стационарных скоростях реакций. Тем не менее они дифференцированы по динамике и имеют разные кинетические сигнатуры.[82]
В отличие от растительных микроРНК, животные микроРНК нацелены на различные гены.[33] Однако гены, участвующие в функциях, общих для всех клеток, таких как экспрессия генов, имеют относительно меньше участков-мишеней микроРНК и, по-видимому, подвергаются отбору, чтобы избежать нацеливания микроРНК.[93]
дцРНК также может активировать экспрессия гена, механизм, который был назван «активацией гена, индуцированной малой РНК» или РНКа. дцРНК, нацеленные на промоторы генов, могут индуцировать мощную активацию транскрипции ассоциированных генов. Это было продемонстрировано на клетках человека с использованием синтетических дцРНК, называемых малыми активирующими РНК (saRNAs ),[94] но также было продемонстрировано для эндогенной микроРНК.[95]
Взаимодействие между микроРНК и комплементарными последовательностями генов и даже псевдогены эта доля гомология последовательностей считаются обратным каналом связи, регулирующим уровни экспрессии между паралогическими генами. Получив название «конкурирующие эндогенные РНК» (цРНК ), эти микроРНК связываются с «элементами ответа микроРНК» на генах и псевдогенах и могут служить другим объяснением устойчивости некодирующей ДНК.[96]
Некоторые исследования показывают, что груз мРНК экзосом может играть роль в имплантации, они могут нарушать адгезию между трофобластом и эндометрием или поддерживать адгезию путем понижающей или повышающей регуляции экспрессии генов, участвующих в адгезии / инвазии.[97]
Эволюция
миРНК хорошо консервированный как у растений, так и у животных, и они считаются жизненно важным и эволюционно древним компонентом регуляции генов.[98][99][100][101][102] Хотя основные компоненты пути микроРНК сохраняются между растения и животные, репертуары miRNA в двух царствах, по-видимому, возникли независимо с разными первичными способами действия.[103][104]
микроРНК полезны филогенетический маркеры из-за их явно низкой скорости эволюции.[105] Происхождение микроРНК как регуляторный механизм развился из предыдущего механизма РНКи, который первоначально использовался в качестве защиты от экзогенного генетического материала, такого как вирусы.[106] Их происхождение могло способствовать развитию морфологических инноваций и, сделав экспрессию генов более специфичной и `` настраиваемой '', разрешило возникновение сложных органов.[107] и, возможно, в конечном итоге, сложная жизнь.[102] Быстрые всплески морфологических инноваций обычно связаны с высокой скоростью накопления микроРНК.[105][107]
Новые микроРНК создаются несколькими способами. Новые микроРНК могут возникать в результате случайного образования шпилек в «некодирующих» участках ДНК (то есть в интронах или межгенных областях), но также в результате дупликации и модификации существующих микроРНК.[108] микроРНК могут также образовываться из перевернутых дупликаций последовательностей, кодирующих белок, что позволяет создавать складную структуру шпильки.[109] Скорость эволюции (т. Е. Нуклеотидного замещения) в недавно возникших микроРНК сравнима с таковой в других частях некодирующей ДНК, что подразумевает эволюцию за счет нейтрального дрейфа; однако более старые микроРНК имеют гораздо более низкую скорость изменения (часто менее одной замены за сто миллионов лет),[102] предполагая, что как только микроРНК получает функцию, она подвергается очищающей селекции.[108] Отдельные области в пределах гена miRNA сталкиваются с различным эволюционным давлением, где области, которые жизненно важны для процессинга и функционирования, имеют более высокие уровни сохранения.[110] На этом этапе микроРНК редко теряется из генома животного,[102] хотя более новые микроРНК (таким образом, предположительно нефункциональные) часто теряются.[108] В Arabidopsis thaliana чистый поток генов miRNA был предсказан между 1,2 и 3,3 генами на миллион лет.[111] Это делает их ценным филогенетическим маркером, и они рассматриваются как возможное решение выдающихся филогенетических проблем, таких как взаимоотношения членистоногие.[112] С другой стороны, во многих случаях микроРНК плохо коррелируют с филогенезом, и возможно, что их филогенетическое соответствие в значительной степени отражает ограниченный выбор микроРНК.[113]
микроРНК входят в состав геномы большинства эукариотических организмов, от бурые водоросли[114] животным. Однако разница в том, как эти микроРНК функционируют и как они обрабатываются, предполагает, что микроРНК возникли независимо у растений и животных.[115]
Ориентируясь на животных, геном Mnemiopsis leidyi[116] по-видимому, отсутствуют узнаваемые микроРНК, а также ядерные белки Дроша и Паша, которые имеют решающее значение для биогенеза канонических микроРНК. На данный момент это единственное животное, пропавшее без вести Дроша. МикроРНК играют жизненно важную роль в регуляции экспрессии генов у всех исследованных животных без гребневиков, за исключением Trichoplax adhaerens, единственный известный член филума Placozoa.[117]
У всех видов к марту 2010 было идентифицировано более 5000 различных miRNA.[118] В то время как короткие последовательности РНК (50 - сотни пар оснований) с широко сопоставимой функцией встречаются у бактерий, у бактерий отсутствуют настоящие микроРНК.[119]
Экспериментальное обнаружение и манипуляции
В то время как исследователи сосредоточили свое внимание на экспрессии miRNA в физиологических и патологических процессах, возникли различные технические переменные, связанные с выделением микроРНК. Стабильность хранимых образцов miRNA была поставлена под сомнение.[66] микроРНК разлагаются гораздо легче, чем мРНК, частично из-за их длины, но также из-за повсеместного присутствия РНКазы. Это приводит к необходимости охлаждать образцы на льду и использовать РНКаза -бесплатное оборудование.[120]
Экспрессию микроРНК можно количественно оценить в два этапа. полимеразной цепной реакции процесс изменения ОТ-ПЦР с последующим количественная ПЦР. Варианты этого метода позволяют получить абсолютную или относительную количественную оценку.[121] миРНК также можно гибридизировать с микрочипы, слайды или чипы с зондами к сотням или тысячам мишеней miRNA, так что относительные уровни miRNA могут быть определены в различных образцах.[122] микроРНК могут быть как обнаружены, так и профилированы методами высокопроизводительного секвенирования (секвенирование микроРНК ).[123] Активность miRNA можно экспериментально ингибировать с помощью заблокированная нуклеиновая кислота (LNA) олиго, а Морфолино олиго[124][125] или олиго 2'-O-метил РНК.[126] Конкретная miRNA может подавляться комплементарным антагомир. Созревание микроРНК может ингибироваться в нескольких точках стерически блокирующими олигонуклеотидами.[127][128] Сайт-мишень miRNA транскрипта мРНК также может быть заблокирован стерически-блокирующим олиго.[129] Для обнаружения miRNA in situ, LNA[130] или морфолино[131] могут использоваться зонды. Закрытая конформация LNA приводит к улучшенным свойствам гибридизации и увеличивает чувствительность и селективность, что делает ее идеальной для обнаружения коротких miRNA.[132]
Количественная оценка miRNA с высокой пропускной способностью подвержена ошибкам из-за большей дисперсии (по сравнению с мРНК ), что сопряжено с методологическими проблемами. мРНК -экспрессия поэтому часто анализируется, чтобы проверить эффекты miRNA на их уровнях (например, в[133]). Базы данных можно использовать для создания пары мРНК - и данные miRNA, которые предсказывают мишени miRNA на основе их последовательности оснований.[134][135] Хотя это обычно делается после обнаружения представляющих интерес miRNA (например, из-за высокого уровня экспрессии), идеи для инструментов анализа, которые объединяют мРНК - и была предложена информация об экспрессии miRNA.[136][137]
Болезнь
Подобно тому, как миРНК участвует в нормальном функционировании эукариотических клеток, нарушение регуляции миРНК связано с заболеванием. Созданная вручную общедоступная база данных miR2Disease документирует известную взаимосвязь между нарушением регуляции miRNA и заболеванием человека.[138]
Наследственные болезни
Мутация в области семян miR-96 вызывает наследственную прогрессирующую потерю слуха.[139]
Мутация в семенной области miR-184 вызывает наследственный кератоконус с передней полярной катарактой.[140]
Делеция кластера miR-17 ~ 92 вызывает дефекты скелета и роста.[141]
Рак
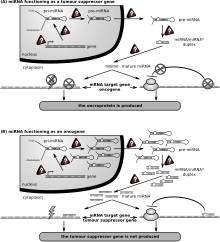
Первым заболеванием человека, связанным с нарушением регуляции miRNA, был хронический лимфолейкоз. Многие другие микроРНК также связаны с раком и, соответственно, их иногда называют «онкомиры ". В злокачественных В-клетках миРНК участвуют в основных путях развития В-клеток, таких как В-клеточный рецептор (BCR) передача сигналов, миграция / адгезия B-клеток, межклеточные взаимодействия в иммунных нишах, а также производство и переключение классов иммуноглобулинов. MiRNA влияют на созревание В-клеток, образование пре-, маргинальной зоны, фолликулярных, В1, плазменных В-клеток и В-клеток памяти.
Другая роль miRNA при раке заключается в использовании уровня их экспрессии для прогноза. В НМРЛ образцы, низкие miR-324 Уровни могут служить индикатором плохой выживаемости.[142] Либо высокий уровень miR-185, либо низкий уровень miR-133b могут коррелировать с метастаз и плохая выживаемость в колоректальный рак.[143]
Кроме того, специфические miRNA могут быть связаны с определенными гистологическими подтипами колоректального рака. Например, было показано, что уровни экспрессии miR-205 и miR-373 увеличиваются при муцинозном колоректальном раке и связанном с муцином язвенном колите раке толстой кишки, но не при спорадической аденокарциноме толстой кишки, в которой отсутствуют муцинозные компоненты.[144] Исследования in vitro показали, что miR-205 и miR-373 могут функционально индуцировать различные признаки опухолевого прогрессирования, связанного с муцином, в эпителиальных клетках кишечника.[144]
Пролиферация клеток гепатоцеллюлярной карциномы может возникать в результате взаимодействия miR-21 с MAP2K3, геном-репрессором опухоли.[145] Оптимальное лечение рака включает точное определение пациентов для терапии со стратификацией риска. Те, у кого есть быстрый ответ на начальное лечение, могут получить пользу от сокращенных схем лечения, что свидетельствует о ценности точных показателей реакции на заболевание. Бесклеточные миРНК очень стабильны в крови, сверхэкспрессируются при раке и поддаются количественной оценке в диагностической лаборатории. В классической Лимфома Ходжкина miR-21, miR-494 и miR-1973 в плазме являются многообещающими биомаркерами ответа на заболевание.[146] Циркулирующие миРНК могут способствовать принятию клинических решений и интерпретации позитронно-эмиссионная томография в сочетании с компьютерная томография. Их можно проводить на каждой консультации для оценки реакции на заболевание и выявления рецидива.
МикроРНК могут быть использованы в качестве инструментов или мишеней для лечения различных видов рака.[147] В нескольких исследованиях было обнаружено, что специфическая микроРНК, miR-506, действует как антагонист опухоли. Было обнаружено, что в значительном количестве образцов рака шейки матки экспрессия miR-506 подавлена. Кроме того, miR-506 способствует апоптозу клеток рака шейки матки посредством своего прямого транскрипционного фактора пути хэджхог, Gli3.[148][149]
Ремонт ДНК и рак
Рак вызывается накоплением мутации от повреждения ДНК или неисправленных ошибок в Репликация ДНК.[150] Дефекты в Ремонт ДНК вызывают накопление мутаций, которые могут привести к раку.[151] Некоторые гены, участвующие в репарации ДНК, регулируются микроРНК.[152]
Линия зародыша мутации в генах репарации ДНК вызывают только 2–5% рак толстой кишки случаи.[153] Однако измененная экспрессия микроРНК, вызывающая дефицит репарации ДНК, часто связана с раком и может быть важным фактором. причинный фактор. Среди 68 спорадических случаев рака толстой кишки со сниженной экспрессией Ремонт несоответствия ДНК белок MLH1, большинство из них были признаны недостаточными из-за эпигенетическое метилирование из CpG остров MLH1 ген.[154] Однако до 15% дефицита MLH1 при спорадическом раке толстой кишки, по-видимому, обусловлены сверхэкспрессией микроРНК miR-155, которая подавляет экспрессию MLH1.[155]
В 29–66%[156][157] из глиобластомы, Репарация ДНК недостаточна из-за эпигенетического метилирования MGMT ген, который снижает экспрессию белка MGMT. Однако для 28% глиобластом белок MGMT недостаточен, но промотор MGMT не метилирован.[156] В глиобластомах без метилированных промоторов MGMT уровень микроРНК miR-181d составляет обратно коррелированный с экспрессией белка MGMT и прямой мишенью miR-181d является MGMT мРНК 3'UTR ( три основных непереведенных региона мРНК MGMT).[156] Таким образом, в 28% глиобластом повышенная экспрессия miR-181d и пониженная экспрессия фермента репарации ДНК MGMT могут быть причинным фактором.
HMGA белки (HMGA1a, HMGA1b и HMGA2) участвуют в развитии рака, и экспрессия этих белков регулируется микроРНК. Экспрессия HMGA почти не обнаруживается в дифференцированных тканях взрослого человека, но повышена во многих случаях рака. Белки HMGA являются полипептиды из ~ 100 аминокислотных остатков, характеризующихся модульной организацией последовательностей. Эти белки имеют три высоко положительно заряженных области, называемых Крючки AT, которые связывают малую бороздку участков ДНК, богатых АТ, в определенных областях ДНК. Новообразования человека, включая карциномы щитовидной железы, предстательной железы, шейки матки, колоректального тракта, поджелудочной железы и яичников, демонстрируют сильное увеличение белков HMGA1a и HMGA1b.[158] У трансгенных мышей с HMGA1, нацеленным на лимфоидные клетки, развивается агрессивная лимфома, что показывает, что высокая экспрессия HMGA1 связана с раком и что HMGA1 может действовать как онкоген.[159] Белок HMGA2 специфически нацелен на промотор ERCC1, тем самым снижая экспрессию этого гена репарации ДНК.[160] Экспрессия белка ERCC1 была недостаточной в 100% из 47 оцененных случаев рака толстой кишки (хотя степень участия HGMA2 неизвестна).[161]
Сердечное заболевание
Глобальная роль функции miRNA в сердце была решена путем условного ингибирования созревания miRNA в мышиный сердце. Это показало, что miRNAs играют важную роль во время его развития.[162][163] Исследования профилей экспрессии miRNA демонстрируют, что уровни экспрессии определенных miRNA изменяются в пораженном сердце человека, указывая на их участие в кардиомиопатии.[164][165][166] Более того, исследования специфических miRNA на животных идентифицировали различные роли miRNA как во время развития сердца, так и при патологических состояниях, включая регуляцию ключевых факторов, важных для кардиогенеза, реакции гипертрофического роста и сердечной проводимости.[163][167][168][169][170][171] Другая роль miRNA в сердечно-сосудистых заболеваниях - использование уровней их экспрессии для диагностики, прогноза или стратификации риска.[172] miRNA в животных моделях также были связаны с метаболизмом и регуляцией холестерина.
miRNA-712
Мышиный микроРНК-712 является потенциальным биомаркером (т.е. предиктором) для атеросклероз, сердечно-сосудистое заболевание артериальной стенки, связанное с задержкой липидов и воспалением.[173] Неламинарный кровоток также коррелирует с развитием атеросклероза, поскольку механосеноры эндотелиальных клеток реагируют на силу сдвига нарушенного кровотока (d-поток).[174] Ряд проатерогенных генов, включая матричные металлопротеиназы (MMP) активируются d-потоком,[174] опосредуют провоспалительные и проангиогенные сигналы. Эти данные наблюдались в перевязанных сонных артериях мышей, чтобы имитировать эффекты d-потока. В течение 24 часов ранее существовавшая незрелая miR-712 образовывала зрелую miR-712, что позволяет предположить, что miR-712 является чувствительной к потоку.[174] Соответствуя этим результатам, miR-712 также активируется в эндотелиальных клетках, подверженных естественному d-потоку в большей кривизне дуги аорты.[174]
Происхождение
Последовательность пре-мРНК miR-712 генерируется из гена рибосомных RN45 мыши на уровне внутренняя расшифрованная прокладка регион 2 (ИТС2).[174] XRN1 представляет собой экзонуклеазу, которая разрушает область ITS2 во время процессинга RN45.[174] Снижение XRN1 при d-потокеТаким образом, условия приводит к накоплению miR-712.[174]
Механизм
MiR-712 нацелен на тканевый ингибитор металлопротеиназы 3 (TIMP3).[174] ТИМП обычно регулируют активность матриксных металлопротеиназ (ММП), которые разрушают внеклеточный матрикс (ЕСМ). Артериальный ECM в основном состоит из коллаген и эластин волокна, обеспечивающие структурную поддержку и свойства отдачи артерий.[175] Эти волокна играют решающую роль в регуляции воспаления и проницаемости сосудов, которые важны для развития атеросклероза.[176] TIMP3, экспрессируемый эндотелиальными клетками, является единственным TIMP, связанным с ECM.[175] Снижение экспрессии TIMP3 приводит к увеличению деградации ECM в присутствии d-потока. В соответствии с этими находками, ингибирование pre-miR712 увеличивает экспрессию TIMP3 в клетках даже при воздействии турбулентного потока.[174]
TIMP3 также снижает экспрессию TNFα (провоспалительного регулятора) во время турбулентного потока.[174] Активность TNFα в турбулентном потоке измеряли по экспрессии TNFα-превращающего фермента (TACE) в крови. TNFα снижается, если miR-712 ингибируется или TIMP3 сверхэкспрессируется,[174] предполагая, что miR-712 и TIMP3 регулируют активность TACE в условиях турбулентного потока.
Анти-miR-712 эффективно подавляет экспрессию miR-712, индуцированную d-потоком, и увеличивает экспрессию TIMP3.[174] Анти-miR-712 также ингибирует повышенную проницаемость сосудов, тем самым значительно снижая развитие атеросклероза и инфильтрацию иммунных клеток.[174]
Человеческий гомолог микроРНК-205
Человеческий гомолог miR-712 был обнаружен в гене гомолога RN45s, который поддерживает miRNA, подобные мышам.[174] MiR-205 человека имеет сходные последовательности с miR-712 мышей и сохраняется у большинства позвоночных.[174] MiR-205 и miR-712 также разделяют более 50% мишеней передачи сигналов клеток, включая TIMP3.[174]
При тестировании d-поток снижал экспрессию XRN1 у людей, как и в эндотелиальных клетках мышей, что указывает на потенциально общую роль XRN1 у людей.[174]
Болезнь почек
Целевое удаление Dicer в FoxD1 -производные почечные клетки-предшественники на мышиной модели привели к сложному почечному фенотипу, включая увеличение нефрон прародители, меньше ренин клетки, гладкие мышцы артериолы, прогрессивный мезангиальный выпадение и гломерулярные аневризмы.[177] Высокая пропускная способность в целом транскриптом профилирование модели мышей с нокаутом FoxD1-Dicer выявило эктопическую активацию проапоптотического гена, Bcl2L11 (Bim) и нарушение регуляции p53 пути с увеличением эффекторных генов p53, включая Bax, Trp53inp1, Июн, Cdkn1a, Mmp2, и Arid3a. Уровни белка p53 остались неизменными, что позволяет предположить, что стромальные miRNA FoxD1 непосредственно репрессируют гены-эффекторы p53. Используя подход отслеживания происхождения, за которым следует Флуоресцентно-активируемая сортировка клеток, профилирование miRNA клеток, происходящих из FoxD1, не только всесторонне определили транскрипционный ландшафт miRNAs, которые являются критическими для развития сосудов, но также идентифицировали ключевые miRNAs, которые, вероятно, модулируют почечный фенотип в его отсутствие. Эти miRNA включают miRs-10a, 18a, 19b, 24, 30c, 92a, 106a, 130a, 152, 181a, 214, 222, 302a, 370 и 381, которые регулируют Bcl2L11 (Bim) и miRs-15b, 18a, 21, 30c, 92a, 106a, 125b-5p, 145, 214, 222, 296-5p и 302a, которые регулируют гены эффекторов р53. В соответствии с результатами профилирования, эктопический апоптоз наблюдали в клеточных производных линии предков, происходящих от FoxD1, и подтверждают важность почечных стромальных miRNAs в клеточном гомеостазе.[177]
Нервная система
miRNAs, по-видимому, регулируют развитие и функцию нервная система.[178] Нервные миРНК участвуют на различных стадиях синаптического развития, включая дендритогенез (включая miR-132, miR-134 и miR-124 ), синапс формирование[179] и созревание синапсов (где, как полагают, участвуют miR-134 и miR-138).[180] Некоторые исследования обнаруживают измененную экспрессию miRNA в Болезнь Альцгеймера,[181] а также шизофрения, биполярное расстройство, большая депрессия и тревожные расстройства.[182][183][184]
Инсульт
По данным Центра по контролю и профилактике заболеваний, инсульт является одной из основных причин смерти и длительной инвалидности в Америке. 87% случаев связаны с ишемическим инсультом, который возникает в результате закупорки артерии головного мозга, по которой проходит богатая кислородом кровь. Нарушение кровотока означает, что мозг не может получать необходимые питательные вещества, такие как кислород и глюкозу, и удалять отходы, такие как углекислый газ.[185][186] miRNA играют роль в посттрансляционном подавлении генов, воздействуя на гены в патогенезе церебральной ишемии, такие как воспалительный, ангиогенез и апоптотический путь.[187]
Алкоголизм
Жизненно важная роль miRNA в экспрессии генов важна для зависимость в частности алкоголизм.[188] Хроническое злоупотребление алкоголем приводит к стойким изменениям функции мозга, частично опосредованным изменениями в экспрессия гена.[188] miRNA глобальная регуляция многих нижестоящих генов считается важной в отношении реорганизации или синаптических связей или долгосрочных нейронных адаптаций, включающих изменение поведения от потребления алкоголя к вывод и / или зависимость.[189] Было обнаружено, что в посмертном мозге алкоголика изменено до 35 различных miRNA, все из которых нацелены на гены, которые включают регуляцию клеточный цикл, апоптоз, клеточная адгезия, развитие нервной системы и клеточная сигнализация.[188] Измененные уровни миРНК были обнаружены в медиальном префронтальная кора алкогользависимых мышей, что предполагает роль miRNA в управлении трансляционным дисбалансом и создании дифференциально экспрессируемых белков в той области мозга, где сложное когнитивное поведение и принятие решений скорее всего происходят.[190]
miRNAs могут быть либо активированы, либо подавлены в ответ на хроническое употребление алкоголя. Экспрессия miR-206 увеличивалась в префронтальной коре головного мозга алкогольных крыс, нацеливаясь на фактор транскрипции нейротрофический фактор мозга (BDNF ) и, в конечном итоге, уменьшит его выраженность. BDNF играет критически важную роль в формировании и созревании новых нейронов и синапсов, что предполагает возможное участие в росте синапсов /синаптическая пластичность у лиц, злоупотребляющих алкоголем.[191] miR-155, важная для регуляции вызванного алкоголем нейровоспаление ответов, было обнаружено, что он активирован, что предполагает роль микроглия и воспалительный цитокины в патофизиологии алкоголя.[192] Подавление miR-382 было обнаружено в прилежащее ядро, структура в базальный передний мозг важно в регулировании чувств награда это сила мотивационных привычек. miR-382 - цель для дофаминовый рецептор D1 (DRD1), и его сверхэкспрессия приводит к усилению регуляции DRD1 и дельта fosB, фактор транскрипции, который активирует серию событий транскрипции в ядре прилежащий что в конечном итоге приводит к аддиктивному поведению.[193] С другой стороны, сверхэкспрессия miR-382 приводила к ослаблению питья и ингибированию DRD1 и дельта fosB усиление регуляции в моделях алкоголизма на крысах, демонстрируя возможность использования miRNA-target фармацевтические препараты в лечении.[193]
Ожирение
miRNA играют решающую роль в регуляции стволовая клетка предшественники дифференцируются в адипоциты.[194] Исследования для определения роли плюрипотентные стволовые клетки играть в адипогенез, были исследованы в увековеченном человеческом Костный мозг -производный стромальная клетка линия hMSC-Tert20.[195] Снижение экспрессии miR-155, miR-221 и miR-222 было обнаружено во время адипогенного программирования как иммортализованных, так и первичных hMSC, что позволяет предположить, что они действуют как негативные регуляторы дифференцировки. Наоборот, эктопическое выражение miRNA 155, 221 и 222 значительно ингибировали адипогенез и подавляли индукцию главных регуляторов PPARγ и CCAAT / связывающий энхансер белок альфа (CEBPA ).[196] Это открывает путь к возможному лечению генетического ожирения.
Другой класс миРНК, регулирующих резистентность к инсулину, ожирение, и сахарный диабет, это let-7 семья. Let-7 накапливается в тканях человека в процессе старение.[197] Когда let-7 подвергалась эктопической сверхэкспрессии, чтобы имитировать ускоренное старение, мыши становились инсулинорезистентными и, следовательно, более склонными к ожирению, вызванному диетой с высоким содержанием жиров, и сахарный диабет.[198] Напротив, когда let-7 подавлялась инъекциями let-7-специфичного антагомиры, мыши становятся более чувствительными к инсулину и значительно устойчивы к ожирению и диабету, вызванному диетой с высоким содержанием жиров. Подавление let-7 может не только предотвратить ожирение и диабет, но и обратить вспять и вылечить это состояние.[199] Эти экспериментальные данные предполагают, что ингибирование let-7 может представлять собой новую терапию для ожирение и диабет 2 типа.
Гемостаз
miRNA также играют решающую роль в регуляции сложных ферментных каскадов, включая систему свертывания крови.[200] Крупномасштабные исследования функционального нацеливания miRNA недавно обнаружили рациональные терапевтические мишени в системе гемостаза.[201][202]
Некодирующие РНК
Когда проект генома человека нанес на карту свой первый хромосома в 1999 году было предсказано, что геном будет содержать более 100 000 генов, кодирующих белок. Однако в конечном итоге было идентифицировано только около 20 000 человек.[203] С тех пор появление биоинформатических подходов в сочетании с исследованиями разбиения генома на транскриптом,[204] систематическое секвенирование полной длины кДНК библиотеки[205] и экспериментальная проверка[206] (включая создание антисмысловых олигонуклеотидов на основе miRNA, называемых антагомиры ) показали, что многие транскрипты представляют собой небелковые РНК, в том числе несколько snoRNAs и миРНК.[207]
Вирусы
Вирусные микроРНК играют важную роль в регуляции экспрессии генов вируса и / или генов хозяина, принося пользу вирусу. Следовательно, miRNAs играют ключевую роль во взаимодействиях хозяин-вирус и патогенезе вирусных заболеваний.[208][209] Экспрессия активаторов транскрипции вирус герпеса человека-6 Считается, что ДНК регулируется вирусной miRNA.[210]
Прогнозирование цели
miRNA могут связываться с транскриптами целевой информационной РНК (мРНК) генов, кодирующих белок, и отрицательно контролировать их трансляцию или вызывать деградацию мРНК. Ключевое значение имеет точное определение мишеней miRNA.[211] Сравнение прогностической эффективности восемнадцати in silico алгоритмы доступны.[212] Крупномасштабные исследования функционального нацеливания miRNA показывают, что многие функциональные miRNA могут быть упущены с помощью алгоритмов прогнозирования мишеней.[201]
Смотрите также
- Олигонуклеотиды против miRNA
- Экспрессия гена
- Список инструментов для прогнозирования генов miRNA
- Список инструментов для прогнозирования мишеней miRNA
- МикроДНК
- miR-324-5p
- РНК-интерференция
- Малая интерферирующая РНК
- МикроРНК, происходящая из малой ядрышковой РНК
использованная литература
- ^ а б c Бартель Д.П. (март 2018 г.). «МикроРНК многоклеточных животных». Ячейка. 173 (1): 20–51. Дои:10.1016 / j.cell.2018.03.006. ЧВК 6091663. PMID 29570994.
- ^ а б c d Бартель Д.П. (январь 2009 г.). «МикроРНК: распознавание мишеней и регуляторные функции». Ячейка. 136 (2): 215–33. Дои:10.1016 / j.cell.2009.01.002. ЧВК 3794896. PMID 19167326.
- ^ Фабиан М.Р., Зоненберг Н., Филипович В. (2010). «Регулирование трансляции и стабильности мРНК с помощью микроРНК». Ежегодный обзор биохимии. 79: 351–79. Дои:10.1146 / annurev-biochem-060308-103103. PMID 20533884.
- ^ а б Бартель Д.П. (январь 2004 г.). «МикроРНК: геномика, биогенез, механизм и функции». Ячейка. 116 (2): 281–97. Дои:10.1016 / S0092-8674 (04) 00045-5. PMID 14744438.
- ^ Homo sapiens миРНК в miRBase в Манчестерский университет
- ^ а б Фромм Б., Биллипп Т., Пек Л.Е., Йохансен М., Тарвер Дж. Э., Кинг Б.Л. и др. (2015). «Единая система аннотации генов микроРНК позвоночных и эволюция микроРНКома человека». Ежегодный обзор генетики. 49: 213–42. Дои:10.1146 / annurev-genet-120213-092023. ЧВК 4743252. PMID 26473382.
- ^ Lim LP, Lau NC, Weinstein EG, Abdelhakim A, Yekta S, Rhoades MW, Burge CB, Bartel DP (апрель 2003 г.). «МикроРНК Caenorhabditis elegans». Гены и развитие. 17 (8): 991–1008. Дои:10.1101 / gad.1074403. ЧВК 196042. PMID 12672692.
- ^ а б Лагос-Кинтана М., Раухут Р., Ялчин А., Мейер Дж., Лендекель В., Тушл Т. (апрель 2002 г.). «Идентификация тканеспецифичных микроРНК мыши». Текущая биология. 12 (9): 735–9. Дои:10.1016 / S0960-9822 (02) 00809-6. PMID 12007417.
- ^ а б c Льюис Б.П., Burge CB, Bartel DP (январь 2005 г.). «Консервативное спаривание семян, часто фланкированное аденозинами, указывает на то, что тысячи человеческих генов являются мишенями для микроРНК». Ячейка. 120 (1): 15–20. Дои:10.1016 / j.cell.2004.12.035. PMID 15652477.
- ^ а б c Friedman RC, Farh KK, Burge CB, Bartel DP (январь 2009 г.). «Большинство мРНК млекопитающих являются консервативными мишенями для микроРНК». Геномные исследования. 19 (1): 92–105. Дои:10.1101 / гр.082701.108. ЧВК 2612969. PMID 18955434.
- ^ а б c Ли Р.К., Фейнбаум Р.Л., Амброс V (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК с антисмысловой комплементарностью lin-14». Ячейка. 75 (5): 843–54. Дои:10.1016 / 0092-8674 (93) 90529-У. PMID 8252621.
- ^ а б Рейнхарт Б.Дж., Слэк Ф.Дж., Бассон М., Паскинелли А.Э., Беттингер Д.С., Ругви А.Э., Хорвиц Х.Р., Рувкун Г. (февраль 2000 г.). «21-нуклеотидная РНК let-7 регулирует время развития у Caenorhabditis elegans». Природа. 403 (6772): 901–6. Bibcode:2000Натура.403..901р. Дои:10.1038/35002607. PMID 10706289. S2CID 4384503.
- ^ а б Паскинелли А.Е., Рейнхарт Б.Дж., Слак Ф., Мартиндейл М.К., Курода М.И., Маллер Б., Хейворд, округ Колумбия, Болл Э. , Дэвидсон Э., Рувкун Г. (ноябрь 2000 г.). «Сохранение последовательности и временной экспрессии гетерохронной регуляторной РНК let-7». Природа. 408 (6808): 86–9. Bibcode:2000Натура 408 ... 86П. Дои:10.1038/35040556. PMID 11081512. S2CID 4401732.
- ^ а б c Лагос-Кинтана М., Раухут Р., Лендекель В., Тушл Т. (октябрь 2001 г.). «Идентификация новых генов, кодирующих малые экспрессированные РНК». Наука. 294 (5543): 853–8. Bibcode:2001Sci ... 294..853L. Дои:10.1126 / science.1064921. HDL:11858 / 00-001M-0000-0012-F65F-2. PMID 11679670. S2CID 18101169.
- ^ а б c Лау NC, Лим LP, Вайнштейн Э.Г., Бартель Д.П. (октябрь 2001 г.). «Обильный класс крошечных РНК с вероятной регуляторной ролью у Caenorhabditis elegans». Наука. 294 (5543): 858–62. Bibcode:2001Наука ... 294..858Л. Дои:10.1126 / science.1065062. PMID 11679671. S2CID 43262684.
- ^ а б c Ли RC, Амброс V (октябрь 2001 г.). «Обширный класс малых РНК у Caenorhabditis elegans». Наука. 294 (5543): 862–4. Bibcode:2001Sci ... 294..862L. Дои:10.1126 / science.1065329. PMID 11679672. S2CID 33480585.
- ^ Винхолдс Э., Клоостерман В.П., Миска Э., Альварес-Сааведра Э., Березиков Э., де Брейн Э., Хорвиц Х.Р., Кауппинен С., Пластерк Р.Х. (июль 2005 г.). «Экспрессия микроРНК в эмбриональном развитии рыбок данио». Наука. 309 (5732): 310–1. Bibcode:2005Наука ... 309..310Вт. Дои:10.1126 / science.1114519. PMID 15919954. S2CID 38939571.
- ^ а б Джонс-Роудс MW, Бартель Д.П., Бартель Б (2006). «MicroRNAS и их регуляторные роли в растениях». Ежегодный обзор биологии растений. 57: 19–53. Дои:10.1146 / annurev.arplant.57.032905.105218. PMID 16669754.
- ^ Бреннеке Дж., Хипфнер Д.Р., Старк А., Рассел Р. Б., Коэн С. М. (апрель 2003 г.). «бантам кодирует регулируемую в процессе развития микроРНК, которая контролирует пролиферацию клеток и регулирует проапоптотический ген hid у дрозофилы». Ячейка. 113 (1): 25–36. Дои:10.1016 / S0092-8674 (03) 00231-9. PMID 12679032.
- ^ Куэльяр Т.Л., Макманус М.Т. (декабрь 2005 г.). «МикроРНК и эндокринная биология». Журнал эндокринологии. 187 (3): 327–32. Дои:10.1677 / joe.1.06426. PMID 16423811.
- ^ Пой М.Н., Элиассон Л., Круцфельдт Дж., Куваджима С., Ма Х, Макдональд П.Е., Пфеффер С., Тушл Т., Раевски Н., Рорсман П., Стоффель М. (ноябрь 2004 г.). «МикроРНК, специфичная для островков поджелудочной железы, регулирует секрецию инсулина». Природа. 432 (7014): 226–30. Bibcode:2004Натура.432..226П. Дои:10.1038 / природа03076. PMID 15538371. S2CID 4415988.
- ^ Чен Ч.З., Ли Л., Лодиш Х.Ф., Бартель Д.П. (январь 2004 г.). «МикроРНК модулируют дифференцировку гемопоэтических клонов». Наука. 303 (5654): 83–6. Bibcode:2004 Наука ... 303 ... 83C. Дои:10.1126 / science.1091903. HDL:1721.1/7483. PMID 14657504. S2CID 7044929.
- ^ Уилфред Б. Р., Ван В. X., Нельсон П. Т. (июль 2007 г.). «Активизирующие исследования miRNA: обзор роли miRNA в метаболизме липидов с предсказанием, что miR-103/107 регулирует метаболические пути человека». Молекулярная генетика и метаболизм. 91 (3): 209–17. Дои:10.1016 / j.ymgme.2007.03.011. ЧВК 1978064. PMID 17521938.
- ^ Harfe BD, McManus MT, Mansfield JH, Hornstein E, Tabin CJ (август 2005 г.). «Дайсер фермента РНКазы III необходим для морфогенеза, но не для формирования паттерна конечности позвоночного». Труды Национальной академии наук Соединенных Штатов Америки. 102 (31): 10898–903. Bibcode:2005PNAS..10210898H. Дои:10.1073 / pnas.0504834102. ЧВК 1182454. PMID 16040801.
- ^ Транг П., Вейдхаас Дж. Б., Slack FJ (декабрь 2008 г.). «МикроРНК как потенциальные лекарственные средства против рака». Онкоген. 27 Дополнение 2: S52–7. Дои:10.1038 / onc.2009.353. PMID 19956180.
- ^ Ли Ч, Фэн Й, Кукос Дж, Чжан Л. (декабрь 2009 г.). «Терапевтические стратегии микроРНК при раке человека». Журнал AAPS. 11 (4): 747–57. Дои:10.1208 / с12248-009-9145-9. ЧВК 2782079. PMID 19876744.
- ^ Фазанаро П., Греко С., Иван М., Капогросси М.С., Мартелли Ф. (январь 2010 г.). «МикроРНК: новые терапевтические мишени при острых ишемических заболеваниях». Фармакология и терапия. 125 (1): 92–104. Дои:10.1016 / j.pharmthera.2009.10.003. PMID 19896977.
- ^ Hydbring P, Badalian-Very G (август 2013 г.). «Клиническое применение микроРНК». F1000 Исследования. 2: 136. Дои:10.12688 / f1000research.2-136.v2. ЧВК 3917658. PMID 24627783.
- ^ Вайтман Б., Ха И., Рувкун Г. (декабрь 1993 г.). «Посттранскрипционная регуляция гетерохронного гена lin-14 с помощью lin-4 опосредует формирование временного паттерна у C. elegans». Ячейка. 75 (5): 855–62. Дои:10.1016/0092-8674(93)90530-4. PMID 8252622.
- ^ Амброс В., Бартель Б., Бартель Д. П., Бердж С. Б., Кэррингтон Дж. К., Чен Х, Дрейфус Г., Эдди С. Р., Гриффитс-Джонс С., Маршалл М., Мацке М., Рувкун Г., Тушл Т. (март 2003 г.). «Единая система аннотации микроРНК». РНК. 9 (3): 277–9. Дои:10.1261 / rna.2183803. ЧВК 1370393. PMID 12592000.
- ^ Гриффитс-Джонс С., Грокок Р. Дж., Ван Донген С., Бейтман А., Энрайт А. Дж. (Январь 2006 г.). «miRBase: последовательности микроРНК, мишени и номенклатура генов». Исследования нуклеиновых кислот. 34 (Выпуск базы данных): D140–4. Дои:10.1093 / nar / gkj112. ЧВК 1347474. PMID 16381832.
- ^ Райт М.В., Бруфорд Е.А. (январь 2011 г.). «Обозначение« мусора »: номенклатура генов небелковой кодирующей РНК (нкРНК) человека». Геномика человека. 5 (2): 90–8. Дои:10.1186/1479-7364-5-2-90. ЧВК 3051107. PMID 21296742.
- ^ а б c Льюис Б.П., Ши И.Х., Джонс-Роудс М.В., Бартель Д.П., Бердж CB (декабрь 2003 г.). «Прогнозирование мишеней микроРНК млекопитающих». Ячейка. 115 (7): 787–98. Дои:10.1016 / S0092-8674 (03) 01018-3. PMID 14697198.
- ^ Элвангер, округ Колумбия, Бюттнер Ф.А., Мевес Х.В., Штумпфлен V (май 2011 г.). «Достаточный минимальный набор типов семян miRNA». Биоинформатика. 27 (10): 1346–50. Дои:10.1093 / биоинформатика / btr149. ЧВК 3087955. PMID 21441577.
- ^ Раевский Н. (июнь 2006 г.). «Предсказания мишеней микроРНК у животных». Природа Генетика. 38 Дополнение (6с): S8–13. Дои:10,1038 / ng1798. PMID 16736023. S2CID 23496396.
- ^ Крек А., Грюн Д., Пой М.Н., Вольф Р., Розенберг Л., Эпштейн Э.Дж., МакМенамин П., да Пьедаде И., Гунсалус К.С., Стоффель М., Раевский Н. (май 2005 г.). «Комбинаторные прогнозы мишеней микроРНК». Природа Генетика. 37 (5): 495–500. Дои:10,1038 / ng1536. PMID 15806104. S2CID 22672750.
- ^ Томсон Д.У., Бракен С.П., Гудолл Г.Дж. (сентябрь 2011 г.). «Экспериментальные стратегии идентификации мишеней микроРНК». Исследования нуклеиновых кислот. 39 (16): 6845–53. Дои:10.1093 / нар / gkr330. ЧВК 3167600. PMID 21652644.
- ^ а б Лим LP, Лау NC, Гарретт-Энгеле П., Гримсон А., Шелтер Дж. М., Касл Дж., Бартель Д. П., Линсли П. С., Джонсон Дж. М. (февраль 2005 г.). «Анализ микроматрицы показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа. 433 (7027): 769–73. Bibcode:2005Натура.433..769L. Дои:10.1038 / природа03315. PMID 15685193. S2CID 4430576.
- ^ Зельбах М., Шванхойссер Б., Тирфельдер Н., Фанг З., Ханин Р., Раевский Н. (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белков, вызванные микроРНК». Природа. 455 (7209): 58–63. Bibcode:2008Натура.455 ... 58С. Дои:10.1038 / природа07228. PMID 18668040. S2CID 4429008.
- ^ Бэк Д., Виллен Дж., Шин С., Камарго Ф. Д., Гиги С. П., Бартель Д. П. (сентябрь 2008 г.). «Влияние микроРНК на выход белка». Природа. 455 (7209): 64–71. Bibcode:2008Натура.455 ... 64Б. Дои:10.1038 / природа07242. ЧВК 2745094. PMID 18668037.
- ^ а б Родригес А., Гриффитс-Джонс С., Ашерст Дж. Л., Брэдли А. (октябрь 2004 г.). «Идентификация генов-хозяев микроРНК млекопитающих и единиц транскрипции». Геномные исследования. 14 (10A): 1902–10. Дои:10.1101 / гр. 2722704. ЧВК 524413. PMID 15364901.
- ^ а б c d Кай Икс, Хагедорн СН, Каллен Б.Р. (декабрь 2004 г.). «Человеческие микроРНК процессируются из кэпированных полиаденилированных транскриптов, которые также могут функционировать как мРНК». РНК. 10 (12): 1957–66. Дои:10.1261 / rna.7135204. ЧВК 1370684. PMID 15525708.
- ^ Вебер MJ (январь 2005 г.). «Новые гены микроРНК человека и мыши, найденные путем поиска гомологии». Журнал FEBS. 272 (1): 59–73. Дои:10.1111 / j.1432-1033.2004.04389.x. PMID 15634332. S2CID 32923462.
- ^ Ким Ю.К., Ким В.Н. (февраль 2007 г.). «Обработка интронных микроРНК». Журнал EMBO. 26 (3): 775–83. Дои:10.1038 / sj.emboj.7601512. ЧВК 1794378. PMID 17255951.
- ^ Баскервиль С., Бартель Д.П. (март 2005 г.). «Профилирование микроматрицы микроРНК выявляет частую коэкспрессию с соседними миРНК и генами хозяина». РНК. 11 (3): 241–7. Дои:10.1261 / rna.7240905. ЧВК 1370713. PMID 15701730.
- ^ а б c Ли Й, Ким М., Хан Дж., Еом К. Х., Ли С., Пэк Ш., Ким В. Н. (октябрь 2004 г.). «Гены микроРНК транскрибируются РНК-полимеразой II». Журнал EMBO. 23 (20): 4051–60. Дои:10.1038 / sj.emboj.7600385. ЧВК 524334. PMID 15372072.
- ^ Чжоу X, Жуань Дж, Ван Г, Чжан В (март 2007 г.). «Характеристика и идентификация основных промоторов микроРНК у четырех модельных видов». PLOS вычислительная биология. 3 (3): e37. Bibcode:2007PLSCB ... 3 ... 37Z. Дои:10.1371 / journal.pcbi.0030037. ЧВК 1817659. PMID 17352530.
- ^ Фаллер М., Го Ф (ноябрь 2008 г.). «Биогенез микроРНК: снять шкуру с кошки можно разными способами». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов. 1779 (11): 663–7. Дои:10.1016 / j.bbagrm.2008.08.005. ЧВК 2633599. PMID 18778799.
- ^ Ли Й, Ан С, Хан Дж, Чой Х, Ким Дж, Йим Дж, Ли Дж, провост П., Родмарк О, Ким С., Ким В.Н. (сентябрь 2003 г.). «Ядерная РНКаза III Дроша инициирует процессинг микроРНК». Природа. 425 (6956): 415–9. Bibcode:2003Натурал.425..415л. Дои:10.1038 / природа01957. PMID 14508493. S2CID 4421030.
- ^ Грегори Р.И., Чендримада Т.П., Шихаттар Р. (2006). «Биогенез микроРНК: выделение и характеристика микропроцессорного комплекса». Протоколы микроРНК. Методы молекулярной биологии. 342. С. 33–47. Дои:10.1385/1-59745-123-1:33. ISBN 978-1-59745-123-9. PMID 16957365.
- ^ Хан Дж, Ли Й, Йом К. Х., Ким Ю. К., Джин Х, Ким В. Н. (декабрь 2004 г.). «Комплекс Дроша-DGCR8 в процессинге первичной микроРНК». Гены и развитие. 18 (24): 3016–27. Дои:10.1101 / gad.1262504. ЧВК 535913. PMID 15574589.
- ^ Хан Дж., Ли Й., Еом К. Х., Нам Дж. В., Хео И., Ри Дж. К., Сон С. Ю., Чо Й, Чжан Б. Т., Ким В. Н. (июнь 2006 г.). «Молекулярная основа распознавания первичных микроРНК комплексом Drosha-DGCR8». Ячейка. 125 (5): 887–901. Дои:10.1016 / j.cell.2006.03.043. PMID 16751099.
- ^ Конрад Т., Марсико А., Гере М., Ором UA (октябрь 2014 г.). «Активность микропроцессора контролирует дифференциальный биогенез miRNA in Vivo». Отчеты по ячейкам. 9 (2): 542–54. Дои:10.1016 / j.celrep.2014.09.007. PMID 25310978.
- ^ Auyeung VC, Ulitsky I, McGeary SE, Bartel DP (февраль 2013 г.). «Помимо вторичной структуры: детерминанты первичной последовательности лицензируют шпильки при-миРНК для обработки». Ячейка. 152 (4): 844–58. Дои:10.1016 / j.cell.2013.01.031. ЧВК 3707628. PMID 23415231.
- ^ Али PS, Ghoshdastider U, Hoffmann J, Brutschy B, Filipek S (ноябрь 2012 г.). «Распознавание предшественника let-7g miRNA человеческим Lin28B». Письма FEBS. 586 (22): 3986–90. Дои:10.1016 / j.febslet.2012.09.034. PMID 23063642. S2CID 28899778.
- ^ Березиков Э., Чунг В.Дж., Уиллис Дж., Куппен Э., Лай Э.С. (октябрь 2007 г.). "Гены миртрона млекопитающих". Молекулярная клетка. 28 (2): 328–36. Дои:10.1016 / j.molcel.2007.09.028. ЧВК 2763384. PMID 17964270.
- ^ а б Кавахара Й., Мегроу М., Крайдер Э., Иизаса Х., Валенте Л., Хатзигеоргиу А.Г., Нисикура К. (сентябрь 2008 г.). «Частота и судьба редактирования микроРНК в мозге человека». Исследования нуклеиновых кислот. 36 (16): 5270–80. Дои:10.1093 / nar / gkn479. ЧВК 2532740. PMID 18684997.
- ^ Винтер Дж., Юнг С., Келлер С., Грегори Р. И., Дидерикс С. (март 2009 г.). «Многие пути к зрелости: пути биогенеза микроРНК и их регуляция». Природа клеточной биологии. 11 (3): 228–34. Дои:10.1038 / ncb0309-228. PMID 19255566. S2CID 205286318.
- ^ Ohman M (октябрь 2007 г.). «Претендент на редактирование от одного лица к другому или союзник процесса микроРНК». Биохимия. 89 (10): 1171–6. Дои:10.1016 / j.biochi.2007.06.002. PMID 17628290.
- ^ а б Мерчисон EP, Hannon GJ (Июнь 2004 г.). «miRNAs в движении: биогенез miRNA и механизм RNAi». Текущее мнение в области клеточной биологии. 16 (3): 223–9. Дои:10.1016 / j.ceb.2004.04.003. PMID 15145345.

- ^ а б c Лунд Э, Дальберг Дж. Э. (2006). «Субстратная селективность экспортина 5 и Дайсера в биогенезе микроРНК». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 71: 59–66. Дои:10.1101 / sqb.2006.71.050. PMID 17381281.
- ^ Пак Джей, Хео И, Тиан Й, Симаншу Д. К., Чанг Х, Джи Ди, Патель DJ, Ким В. Н. (июль 2011 г.). «Dicer распознает 5'-конец РНК для эффективной и точной обработки». Природа. 475 (7355): 201–5. Дои:10.1038 / природа10198. ЧВК 4693635. PMID 21753850.
- ^ Джи Х (2008). «Механизм действия РНКазы III: как режет кубик». РНК интерференция. Актуальные темы микробиологии и иммунологии. 320. С. 99–116. Дои:10.1007/978-3-540-75157-1_5. ISBN 978-3-540-75156-4. PMID 18268841.
- ^ Mirihana Arachchilage G, Dassanayake AC, Basu S (февраль 2015 г.). «Зависимый от ионов калия структурный переключатель РНК регулирует созревание человеческой пре-миРНК 92b». Химия и биология. 22 (2): 262–72. Дои:10.1016 / j.chembiol.2014.12.013. PMID 25641166.
- ^ Сохель MH (2016). «Внеклеточные / циркулирующие микроРНК: механизмы высвобождения, функции и проблемы». Достижения в области наук о жизни. 10 (2): 175–186. Дои:10.1016 / j.als.2016.11.007.
- ^ а б Бёкель Дж. Н., Рейс С. М., Лайстнер Д., Томе К. Э., Зейхер А. М., Фихтльшерер С., Келлер Т. (апрель 2014 г.). «От сердца до кончиков пальцев: вклад сердца на периферические уровни микроРНК». Международный журнал кардиологии. 172 (3): 616–7. Дои:10.1016 / j.ijcard.2014.01.082. PMID 24508494.
- ^ Леландэ-Бриер С., Сорин С., Деклерк М., Бенслиман А., Креспи М., Хартманн С. (март 2010 г.). «Разнообразие малых РНК в растениях и его влияние на развитие». Текущая геномика. 11 (1): 14–23. Дои:10.2174/138920210790217918. ЧВК 2851111. PMID 20808519.
- ^ Рана TM (январь 2007 г.). «Освещая тишину: понимание структуры и функции малых РНК». Обзоры природы Молекулярная клеточная биология. 8 (1): 23–36. Дои:10.1038 / nrm2085. PMID 17183358. S2CID 8966239.
- ^ а б Шварц Д.С., Заморе П.Д. (май 2002 г.). «Почему miRNA живут в miRNP?». Гены и развитие. 16 (9): 1025–31. Дои:10.1101 / gad.992502. PMID 12000786.
- ^ Krol J, Sobczak K, Wilczynska U, Drath M, Jasinska A, Kaczynska D, Krzyzosiak WJ (октябрь 2004 г.). «Структурные особенности предшественников микроРНК (миРНК) и их значение для биогенеза миРНК и дизайна малых интерферирующих РНК / коротких шпилечных РНК». Журнал биологической химии. 279 (40): 42230–9. Дои:10.1074 / jbc.M404931200. PMID 15292246.
- ^ Хворова А., Рейнольдс А., Джаясена С.Д. (октябрь 2003 г.). «Функциональные миРНК и миРНК демонстрируют смещение цепи». Ячейка. 115 (2): 209–16. Дои:10.1016 / S0092-8674 (03) 00801-8. PMID 14567918.
- ^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (октябрь 2003 г.). «Асимметрия в сборке ферментного комплекса РНКи». Ячейка. 115 (2): 199–208. Дои:10.1016 / S0092-8674 (03) 00759-1. PMID 14567917.
- ^ Лин С.Л., Чанг Д., Ин С.Ю. (август 2005 г.). «Асимметрия интронных пре-миРНК структур в функциональной сборке RISC». Ген. 356: 32–8. Дои:10.1016 / j.gene.2005.04.036. ЧВК 1788082. PMID 16005165.
- ^ Okamura K, Chung WJ, Lai EC (сентябрь 2008 г.). «Длинный и короткий инвертированные повторы генов у животных: микроРНК, миртроны и шпильки РНК». Клеточный цикл. 7 (18): 2840–5. Дои:10.4161 / cc.7.18.6734. ЧВК 2697033. PMID 18769156.
- ^ а б Пратт А.Дж., Макрей И.Дж. (июль 2009 г.). «РНК-индуцированный комплекс подавления звука: универсальная машина подавления генов». Журнал биологической химии. 284 (27): 17897–901. Дои:10.1074 / jbc.R900012200. ЧВК 2709356. PMID 19342379.
- ^ Макрей И.Дж., Ма Э., Чжоу М., Робинсон CV, Дудна Д.А. (январь 2008 г.). «Восстановление RISC-загрузочного комплекса человека in vitro». Труды Национальной академии наук Соединенных Штатов Америки. 105 (2): 512–7. Bibcode:2008ПНАС..105..512М. Дои:10.1073 / pnas.0710869105. ЧВК 2206567. PMID 18178619.
- ^ Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, Rappsilber J, Mann M, Dreyfuss G (март 2002 г.). «miRNPs: новый класс рибонуклеопротеидов, содержащий множество микроРНК». Гены и развитие. 16 (6): 720–8. Дои:10.1101 / gad.974702. ЧВК 155365. PMID 11914277.
- ^ Мейстер Г., Ландталер М., Петерс Л., Чен П.Й., Урлауб Г., Люрманн Р., Тушл Т. (декабрь 2005 г.). «Идентификация новых белков, связанных с аргонавтом». Текущая биология. 15 (23): 2149–55. Дои:10.1016 / j.cub.2005.10.048. PMID 16289642.
- ^ Jing Q, Huang S, Guth S, Zarubin T., Motoyama A, Chen J, Di Padova F, Lin SC, Gram H, Han J (март 2005 г.). «Участие микроРНК в нестабильности мРНК, опосредованной AU-богатыми элементами». Ячейка. 120 (5): 623–34. Дои:10.1016 / j.cell.2004.12.038. PMID 15766526.
- ^ а б c Кай З.С., Паскинелли А.Е. (январь 2010 г.). «Убийцы микроРНК: факторы, регулирующие исчезновение миРНК». Структурная и молекулярная биология природы. 17 (1): 5–10. Дои:10.1038 / nsmb.1762. ЧВК 6417416. PMID 20051982.
- ^ Чаттерджи С., Гроссханс Х (сентябрь 2009 г.). «Активный оборот модулирует активность зрелой микроРНК у Caenorhabditis elegans». Природа. 461 (7263): 546–9. Bibcode:2009Натура.461..546C. Дои:10.1038 / природа08349. PMID 19734881. S2CID 4414841.
- ^ а б c Морозова Н., Зиновьев А., Нонне Н., Причард Л.Л., Горбань А.Н., Харель-Беллан А. (сентябрь 2012 г.). «Кинетические сигнатуры механизмов действия микроРНК». РНК. 18 (9): 1635–55. Дои:10.1261 / rna.032284.112. ЧВК 3425779. PMID 22850425.
- ^ Ван XJ, Рейес JL, Чуа Н.Х., Gaasterland T (2004). «Прогнозирование и идентификация микроРНК Arabidopsis thaliana и их мРНК-мишеней». Геномная биология. 5 (9): R65. Дои:10.1186 / gb-2004-5-9-r65. ЧВК 522872. PMID 15345049.
- ^ Кавасаки Х, Тайра К. (2004). «МикроРНК-196 ингибирует экспрессию HOXB8 при миелоидной дифференцировке клеток HL60». Серия симпозиумов по нуклеиновым кислотам. 48 (1): 211–2. Дои:10.1093 / nass / 48.1.211. PMID 17150553.
- ^ а б Moxon S, Jing R, Szittya G, Schwach F, Rusholme Pilcher RL, Moulton V, Dalmay T. (октябрь 2008 г.). «Глубокое секвенирование коротких РНК томатов позволяет идентифицировать микроРНК, нацеленные на гены, участвующие в созревании плодов». Геномные исследования. 18 (10): 1602–9. Дои:10.1101 / гр.080127.108. ЧВК 2556272. PMID 18653800.
- ^ Mazière P, Enright AJ (июнь 2007 г.). «Прогнозирование мишеней микроРНК». Открытие наркотиков сегодня. 12 (11–12): 452–8. Дои:10.1016 / j.drudis.2007.04.002. PMID 17532529.
- ^ Уильямс А.Е. (февраль 2008 г.). «Функциональные аспекты микроРНК животных». Клеточные и молекулярные науки о жизни. 65 (4): 545–62. Дои:10.1007 / s00018-007-7355-9. PMID 17965831. S2CID 5708394.
- ^ Eulalio A, Huntzinger E, Nishihara T, Rehwinkel J, Fauser M, Izaurralde E (январь 2009 г.). «Деаденилирование - широко распространенный эффект регуляции miRNA». РНК. 15 (1): 21–32. Дои:10.1261 / rna.1399509. ЧВК 2612776. PMID 19029310.
- ^ Баззини А.А., Ли М.Т., Хиральдез А.Дж. (апрель 2012 г.). «Профилирование рибосом показывает, что miR-430 снижает трансляцию перед тем, как вызвать распад мРНК у рыбок данио». Наука. 336 (6078): 233–7. Bibcode:2012Sci ... 336..233B. Дои:10.1126 / наука.1215704. ЧВК 3547538. PMID 22422859.
- ^ Джуранович С., Нахви А., Грин Р. (апрель 2012 г.). «miRNA-опосредованное подавление гена путем репрессии трансляции с последующим деаденилированием и распадом мРНК». Наука. 336 (6078): 237–40. Bibcode:2012Sci ... 336..237D. Дои:10.1126 / science.1215691. ЧВК 3971879. PMID 22499947.
- ^ Тан И, Чжан Б., Ву Т, Скогербё Г., Чжу Х, Го Х, Хе С., Чен Р. (февраль 2009 г.). «Транскрипционное ингибирование экспрессии Hoxd4 с помощью miRNA-10a в клетках рака груди человека». BMC Молекулярная биология. 10 (1): 12. Дои:10.1186/1471-2199-10-12. ЧВК 2680403. PMID 19232136.
- ^ Хокинс П.Г., Моррис К.В. (март 2008 г.). «РНК и транскрипционная модуляция экспрессии генов». Клеточный цикл. 7 (5): 602–7. Дои:10.4161 / cc.7.5.5522. ЧВК 2877389. PMID 18256543.
- ^ Старк А., Бреннеке Дж., Бушати Н., Рассел Р. Б., Коэн С. М. (декабрь 2005 г.). «МикроРНК животных придают устойчивость экспрессии генов и оказывают значительное влияние на эволюцию 3'UTR». Ячейка. 123 (6): 1133–46. Дои:10.1016 / j.cell.2005.11.023. PMID 16337999.
- ^ Ли LC (2008). «Активация генов, опосредованная малой РНК». В Моррис К.В. (ред.). РНК и регуляция экспрессии генов: скрытый уровень сложности. Horizon Scientific Press. ISBN 978-1-904455-25-7.
- ^ Place RF, Li LC, Pookot D, Noonan EJ, Dahiya R (февраль 2008 г.). «МикроРНК-373 индуцирует экспрессию генов с комплементарными промоторными последовательностями». Труды Национальной академии наук Соединенных Штатов Америки. 105 (5): 1608–13. Bibcode:2008ПНАС..105.1608П. Дои:10.1073 / pnas.0707594105. ЧВК 2234192. PMID 18227514.
- ^ Салмена Л., Полисено Л., Тай Й, Кац Л., Пандольфи П. П. (август 2011 г.). "Гипотеза цРНК: Розеттский камень скрытого языка РНК?". Ячейка. 146 (3): 353–8. Дои:10.1016 / j.cell.2011.07.014. ЧВК 3235919. PMID 21802130.
- ^ https://doi.org/10.1016/j.ebiom.2015.09.003}
- ^ Axtell MJ, Bartel DP (июнь 2005 г.). «Древность микроРНК и их мишеней у наземных растений». Растительная клетка. 17 (6): 1658–73. Дои:10.1105 / tpc.105.032185. ЧВК 1143068. PMID 15849273.
- ^ Танзер А., Штадлер П.Ф. (май 2004 г.). «Молекулярная эволюция кластера микроРНК». Журнал молекулярной биологии. 339 (2): 327–35. CiteSeerX 10.1.1.194.1598. Дои:10.1016 / j.jmb.2004.03.065. PMID 15136036.
- ^ Чен К., Раевский Н. (февраль 2007 г.). «Эволюция регуляции генов с помощью факторов транскрипции и микроРНК». Природа Обзоры Генетика. 8 (2): 93–103. Дои:10,1038 / nrg1990. PMID 17230196. S2CID 174231.
- ^ Ли CT, Рисом Т., Штраус В.М. (апрель 2007 г.). «Эволюционная консервация регуляторных цепей микроРНК: изучение сложности генов микроРНК и консервативных взаимодействий микроРНК с мишенями через филогению многоклеточных». ДНК и клеточная биология. 26 (4): 209–18. Дои:10.1089 / dna.2006.0545. PMID 17465887.
- ^ а б c d Петерсон К.Дж., Дитрих М.Р., МакПик М.А. (июль 2009 г.). «МикроРНК и макроэволюция многоклеточных животных: понимание канализации, сложности и кембрийского взрыва». BioEssays. 31 (7): 736–47. Дои:10.1002 / bies.200900033. PMID 19472371. S2CID 15364875.
- ^ Шабалина С.А., Кунин Е.В. (октябрь 2008 г.). «Происхождение и эволюция эукариотической РНК-интерференции». Тенденции в экологии и эволюции. 23 (10): 578–87. Дои:10.1016 / j.tree.2008.06.005. ЧВК 2695246. PMID 18715673.
- ^ Axtell MJ, Westholm JO, Lai EC (2011). "Vive la différence: биогенез и эволюция микроРНК у растений и животных". Геномная биология. 12 (4): 221. Дои:10.1186 / gb-2011-12-4-221. ЧВК 3218855. PMID 21554756.
- ^ а б Уилер Б.М., Хаймберг А.М., Мой В.Н., Сперлинг Е.А., Голштин Т.В., Хебер С., Петерсон К.Дж. (2009). «Глубокая эволюция микроРНК многоклеточных животных». Эволюция и развитие. 11 (1): 50–68. Дои:10.1111 / j.1525-142X.2008.00302.x. PMID 19196333. S2CID 14924603.
- ^ Пашковский П.П., Рязанский С.С. (июнь 2013 г.). «Биогенез, эволюция и функции растительных микроРНК». Биохимия. Биохимия. 78 (6): 627–37. Дои:10.1134 / S0006297913060084. PMID 23980889. S2CID 12025420.
- ^ а б Хаймберг А.М., Семпере Л.Ф., Мой В.Н., Донохью П.С., Петерсон К.Дж. (февраль 2008 г.). «МикроРНК и появление морфологической сложности позвоночных». Труды Национальной академии наук Соединенных Штатов Америки. 105 (8): 2946–50. Bibcode:2008PNAS..105.2946H. Дои:10.1073 / pnas.0712259105. ЧВК 2268565. PMID 18287013.
- ^ а б c Нодзава М., Миура С., Ней М. (июль 2010 г.). «Происхождение и эволюция генов микроРНК у видов дрозофилы». Геномная биология и эволюция. 2: 180–9. Дои:10.1093 / gbe / evq009. ЧВК 2942034. PMID 20624724.
- ^ Аллен Э., Се З., Густафсон А.М., Сунг Г.Х., Спатафора Дж. У., Кэррингтон Дж. К. (декабрь 2004 г.). «Эволюция генов микроРНК путем инвертированной дупликации последовательностей генов-мишеней в Arabidopsis thaliana». Природа Генетика. 36 (12): 1282–90. Дои:10,1038 / ng1478. PMID 15565108. S2CID 11997028.
- ^ Warthmann N, Das S, Lanz C, Weigel D (май 2008 г.). «Сравнительный анализ локуса микроРНК MIR319a у Arabidopsis и родственных Brassicaceae». Молекулярная биология и эволюция. 25 (5): 892–902. Дои:10.1093 / molbev / msn029. PMID 18296705.
- ^ Fahlgren N, Jogdeo S, Kasschau KD, Sullivan CM, Chapman EJ, Laubinger S, Smith LM, Dasenko M, Givan SA, Weigel D, Carrington JC (апрель 2010 г.). «Эволюция гена микроРНК у Arabidopsis lyrata и Arabidopsis thaliana». Растительная клетка. 22 (4): 1074–89. Дои:10.1105 / tpc.110.073999. ЧВК 2879733. PMID 20407027.
- ^ Caravas J, Фридрих M (июнь 2010 г.). «О клещах и многоножках: недавний прогресс в разрешении основания дерева членистоногих». BioEssays. 32 (6): 488–95. Дои:10.1002 / bies.201000005. PMID 20486135. S2CID 20548122.
- ^ Кенни Нью-Джерси, Намигай Е.К., Марлетаз Ф., Хуэй Дж.Х., Шимелд С.М. (декабрь 2015 г.). «Проект геномных сборок и предполагаемые дополнения микроРНК литоральных lophotrochozoans Patella vulgata (Mollusca, Patellogastropoda) и Spirobranchus (Pomatoceros) lamarcki (Annelida, Serpulida)». Морская геномика. 24 (2): 139–46. Дои:10.1016 / j.margen.2015.07.004. PMID 26319627.
- ^ Кок Дж. М., Стерк Л., Рузе П., Скорнет Д., Аллен А. Э., Амуциас Г. и др. (Июнь 2010 г.). «Геном эктокарпа и независимая эволюция многоклеточности бурых водорослей». Природа. 465 (7298): 617–21. Bibcode:2010 Натур.465..617C. Дои:10.1038 / природа09016. PMID 20520714.
- ^ Куперус Дж. Т., Фальгрен Н., Кэррингтон Дж. К. (февраль 2011 г.). «Эволюция и функциональная диверсификация генов MIRNA». Растительная клетка. 23 (2): 431–42. Дои:10.1105 / tpc.110.082784. ЧВК 3077775. PMID 21317375.
- ^ Райан Дж. Ф., Панг К., Шницлер К. Э., Нгуен А. Д., Морленд Р. Т., Симмонс Д. К., Б. Дж. Кох, Фрэнсис В. Р., Хавлак П., Смит С. А., Патнэм Н.Х., Пикша Ш, Данн К.В., Вольфсберг Т.Г., Малликин Дж.С., Мартиндейл М.К., Баксеванис А.Д. (декабрь 2013 г.). «Геном гребневика Mnemiopsis leidyi и его значение для эволюции типов клеток». Наука. 342 (6164): 1242592. Дои:10.1126 / science.1242592. ЧВК 3920664. PMID 24337300.
- ^ Максвелл Е.К., Райан Дж.Ф., Шницлер К.Э., Браун В.Е., Баксеванис А.Д. (декабрь 2012 г.). «МикроРНК и важные компоненты механизма обработки микроРНК не кодируются в геноме гребневика Mnemiopsis leidyi». BMC Genomics. 13 (1): 714. Дои:10.1186/1471-2164-13-714. ЧВК 3563456. PMID 23256903.
- ^ Даймонд П.Ф. (15 марта 2010 г.). «Терапевтический потенциал миРНК». Новости генной инженерии и биотехнологии. 30 (6): 1. Архивировано из оригинал 10 июля 2010 г.. Получено 10 июля 2010.
- ^ Тьяден Б., Гудвин С.С., Опдайк Дж. А., Гийе М., Фу Д. X., Готтесман С., Сторц Г. (2006). «Целевое предсказание малых некодирующих РНК в бактериях». Исследования нуклеиновых кислот. 34 (9): 2791–802. Дои:10.1093 / нар / gkl356. ЧВК 1464411. PMID 16717284.
- ^ Лю К.Г., Калин Г.А., Волиния С., Кроче С.М. (2008). «Профилирование экспрессии микроРНК с использованием микрочипов». Протоколы природы. 3 (4): 563–78. Дои:10.1038 / nprot.2008.14. PMID 18388938. S2CID 2441105.
- ^ Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH, Nguyen JT, Barbisin M, Xu NL, Mahuvakar VR, Andersen MR, Lao KQ, Livak KJ, Guegler KJ (ноябрь 2005 г.). «Количественная оценка микроРНК в режиме реального времени с помощью ОТ-ПЦР« стержень-петля »». Исследования нуклеиновых кислот. 33 (20): e179. Дои:10.1093 / nar / gni178. ЧВК 1292995. PMID 16314309.
- ^ Шингара Дж., Кейгер К., Шелтон Дж., Лаосинчай-Вольф В., Пауэрс П., Конрад Р., Браун Д., Лабурье Е. (сентябрь 2005 г.). «Оптимизированная платформа для выделения и маркировки для точного профилирования экспрессии микроРНК». РНК. 11 (9): 1461–70. Дои:10.1261 / rna.2610405. ЧВК 1370829. PMID 16043497.
- ^ Buermans HP, Ariyurek Y, van Ommen G, den Dunnen JT, 't Hoen PA (декабрь 2010 г.). «Новые методы профилирования экспрессии микроРНК на основе секвенирования нового поколения». BMC Genomics. 11: 716. Дои:10.1186/1471-2164-11-716. ЧВК 3022920. PMID 21171994.
- ^ Клоостерман В.П., Винхолдс Э., Кеттинг РФ, Plasterk RH (2004). «Требования к субстрату для функции let-7 в развивающемся эмбрионе рыбок данио». Исследования нуклеиновых кислот. 32 (21): 6284–91. Дои:10.1093 / нар / гх968. ЧВК 535676. PMID 15585662.
- ^ Флинт А.С., Ли Н., Тэтчер Э.Дж., Солница-Крезель Л., Паттон Дж. Г. (февраль 2007 г.). «Рыбка данио miR-214 модулирует передачу сигналов Hedgehog для определения судьбы мышечных клеток». Природа Генетика. 39 (2): 259–63. Дои:10,1038 / ng1953. ЧВК 3982799. PMID 17220889.
- ^ Мейстер Г., Ландталер М., Дорсетт Ю., Тушл Т. (март 2004 г.). «Последовательно-специфическое ингибирование вызванного микроРНК и миРНК молчания РНК». РНК. 10 (3): 544–50. Дои:10.1261 / rna.5235104. ЧВК 1370948. PMID 14970398.
- ^ Kloosterman WP, Lagendijk AK, Ketting RF, Moulton JD, Plasterk RH (август 2007 г.). «Целенаправленное ингибирование созревания miRNA с помощью морфолино показывает роль miR-375 в развитии островков поджелудочной железы». PLOS Биология. 5 (8): e203. Дои:10.1371 / journal.pbio.0050203. ЧВК 1925136. PMID 17676975.
- ^ Геберт Л.Ф., Ребхан М.А., Кривелли С.Е., Дензлер Р., Стоффель М., Холл Дж. (Январь 2014 г.). «Миравирсен (SPC3649) может ингибировать биогенез miR-122». Исследования нуклеиновых кислот. 42 (1): 609–21. Дои:10.1093 / nar / gkt852. ЧВК 3874169. PMID 24068553.
- ^ Чой В.Й., Хиралдес А.Дж., Шир А.Ф. (октябрь 2007 г.). «Средства защиты мишеней выявляют ослабление и балансировку агониста и антагониста Nodal с помощью miR-430». Наука. 318 (5848): 271–4. Bibcode:2007Наука ... 318..271C. Дои:10.1126 / science.1147535. PMID 17761850. S2CID 30461594.
- ^ You Y, Moreira BG, Behlke MA, Owczarzy R (май 2006 г.). «Дизайн зондов LNA, улучшающих распознавание несовпадений». Исследования нуклеиновых кислот. 34 (8): e60. Дои:10.1093 / нар / gkl175. ЧВК 1456327. PMID 16670427.
- ^ Lagendijk AK, Moulton JD, Bakkers J (июнь 2012 г.). «Выявление деталей: протокол гибридизации цельной микроРНК in situ для эмбрионов рыбок данио и взрослых тканей». Биология Открыть. 1 (6): 566–9. Дои:10.1242 / bio.2012810. ЧВК 3509442. PMID 23213449.
- ^ Каур Х., Арора А., Венгель Дж., Маити С. (июнь 2006 г.). «Термодинамические, противоионные и гидратационные эффекты для включения заблокированных нуклеотидов нуклеиновых кислот в дуплексы ДНК». Биохимия. 45 (23): 7347–55. Дои:10.1021 / bi060307w. PMID 16752924.
- ^ Нильсен Дж. А., Лау П., Марич Д., Баркер Дж. Л., Хадсон Л. Д. (август 2009 г.). «Интеграция профилей экспрессии микроРНК и мРНК нейрональных предшественников для определения регуляторных сетей, лежащих в основе начала кортикального нейрогенеза». BMC Neuroscience. 10: 98. Дои:10.1186/1471-2202-10-98. ЧВК 2736963. PMID 19689821.
- ^ Гримсон А., Фар К.К., Джонстон В.К., Гарретт-Энджеле П., Лим LP, Бартель Д.П. (июль 2007 г.). «Специфичность нацеливания микроРНК у млекопитающих: детерминанты, выходящие за рамки спаривания семян». Молекулярная клетка. 27 (1): 91–105. Дои:10.1016 / j.molcel.2007.06.017. ЧВК 3800283. PMID 17612493.
- ^ Гриффитс-Джонс С., Сайни Х. К., ван Донген С., Энрайт А. Дж. (Январь 2008 г.). «miRBase: инструменты для геномики микроРНК». Исследования нуклеиновых кислот. 36 (Выпуск базы данных): D154–8. Дои:10.1093 / нар / гкм952. ЧВК 2238936. PMID 17991681.
- ^ Нам С., Ли М., Чой К., Балч С., Ким С., Племянник К.П. (июль 2009 г.). «Интегрированный анализ микроРНК и мРНК (MMIA): веб-инструмент для изучения биологических функций экспрессии микроРНК». Исследования нуклеиновых кислот. 37 (Выпуск веб-сервера): W356–62. Дои:10.1093 / nar / gkp294. ЧВК 2703907. PMID 19420067.
- ^ Артманн С., Юнг К., Блекманн А., Байсбарт Т. (2012). Проверо П. (ред.). «Обнаружение одновременных групповых эффектов в экспрессии микроРНК и связанных наборов генов-мишеней». PLOS ONE. 7 (6): e38365. Bibcode:2012 ПРИЛОЖЕНИЕ ... 738365A. Дои:10.1371 / journal.pone.0038365. ЧВК 3378551. PMID 22723856.
- ^ Цзян Цюй, Ван И, Хао И, Цзюань Л., Тэн М., Чжан Икс, Ли М, Ван Г, Лю И (январь 2009 г.). "miR2Disease: вручную созданная база данных по дерегуляции микроРНК при заболеваниях человека". Исследования нуклеиновых кислот. 37. 37 (Выпуск базы данных): D98–104. Дои:10.1093 / nar / gkn714. ЧВК 2686559. PMID 18927107.
- ^ Mencía A, Modamio-Høybjør S, Redshaw N, Morín M, Mayo-Merino F, Olavarrieta L, Aguirre LA, del Castillo I, Steel KP, Dalmay T, Moreno F, Moreno-Pelayo MA (май 2009 г.). «Мутации в области зародыша miR-96 человека ответственны за несиндромную прогрессирующую потерю слуха». Природа Генетика. 41 (5): 609–13. Дои:10,1038 / нг.355. PMID 19363479. S2CID 11113852.
- ^ Хьюз А.Е., Брэдли Д.Т., Кэмпбелл М., Лехнер Дж., Дэш Д.П., Симпсон Д.А., Уиллоуби CE (ноябрь 2011 г.). «Мутация, изменяющая семенную область miR-184, вызывает семейный кератоконус с катарактой». Американский журнал генетики человека. 89 (5): 628–33. Дои:10.1016 / j.ajhg.2011.09.014. ЧВК 3213395. PMID 21996275.
- ^ de Pontual L, Yao E, Callier P, Faivre L, Drouin V, Cariou S, Van Haeringen A, Geneviève D, Goldenberg A, Oufadem M, Manouvrier S, Munnich A, Vidigal JA, Vekemans M, Lyonnet S, Henrion-Caude А., Вентура А., Амиэль Дж. (Сентябрь 2011 г.). «Делеция зародышевой линии кластера miR-17∼92 вызывает дефекты скелета и роста у людей». Природа Генетика. 43 (10): 1026–30. Дои:10,1038 / нг.915. ЧВК 3184212. PMID 21892160.
- ^ Выза У, Вудер Т., Колде Р., Фишер К., Валк К., Тыниссон Н., Роосипуу Р., Вило Дж., Мецпалу А., Аннило Т. (октябрь 2011 г.). «Идентификация miR-374a как прогностического маркера выживаемости у пациентов с немелкоклеточным раком легкого на ранней стадии». Гены, хромосомы и рак. 50 (10): 812–22. Дои:10.1002 / gcc.20902. PMID 21748820. S2CID 9746594.
- ^ Акчакая П., Экелунд С., Колосенко И., Карамута С., Озата Д.М., Се Х., Линдфорсс Ю., Оливекрона Х., Луи В.О. (август 2011 г.). «Нарушение регуляции miR-185 и miR-133b связано с общей выживаемостью и метастазированием при колоректальном раке». Международный журнал онкологии. 39 (2): 311–8. Дои:10.3892 / ijo.2011.1043. PMID 21573504.
- ^ а б Эйкинг А., Рейс Х., Франк М., Геркен Г., Шмид К.В., Карио Е. (2016). «MiR-205 и MiR-373 связаны с агрессивным муцинозным колоректальным раком человека». PLOS ONE. 11 (6): e0156871. Bibcode:2016PLoSO..1156871E. Дои:10.1371 / journal.pone.0156871. ЧВК 4894642. PMID 27271572.
- ^ МикроРНК-21 способствует пролиферации клеток гепатоцеллюлярной карциномы HepG2 посредством репрессии митоген-активируемой протеинкиназы-киназы 3. Guangxian Xu et al, 2013
- ^ Джонс К., Норс Дж. П., Кин С., Бхатнагар А., Ганди М.К. (январь 2014 г.). «МикроРНК плазмы - биомаркеры реакции на заболевание при классической лимфоме Ходжкина». Клинические исследования рака. 20 (1): 253–64. Дои:10.1158 / 1078-0432.CCR-13-1024. PMID 24222179.
- ^ Хоссейнахли Н., Агапур М., Дуйф PH, Барадаран Б. (август 2018 г.). «Лечение рака с помощью заместительной терапии микроРНК: обзор литературы». Журнал клеточной физиологии. 233 (8): 5574–5588. Дои:10.1002 / jcp.26514. PMID 29521426.
- ^ Лю Г, Сун И, Джи П, Ли Х, Когделл Д., Ян Д., Паркер Керриган BC, Шмулевич И., Чен К., Суд А. К., Сюэ Ф., Чжан В. (июль 2014 г.). «MiR-506 подавляет пролиферацию и вызывает старение, напрямую воздействуя на ось CDK4 / 6-FOXM1 при раке яичников». Журнал патологии. 233 (3): 308–18. Дои:10.1002 / path.4348. ЧВК 4144705. PMID 24604117.
- ^ Вэнь Си, Линь И, Юй Йу, Цао С.Дж., Чжан Р., Ян ХМ, Ли Дж, Чжан Ю.Л., Ван Й.Х., Ма М.З., Сун У.В., Лу XL, Ван Дж.Х., Тэн Ю.С., Чжан З.Г. (февраль 2015 г.). «miR-506 действует как опухолевый супрессор, напрямую воздействуя на фактор транскрипции пути hedgehog Gli3 при раке шейки матки человека». Онкоген. 34 (6): 717–25. Дои:10.1038 / onc.2014.9. PMID 24608427. S2CID 20603801.
- ^ Loeb, Keith R .; Лоэб, Лоуренс А. (март 2000 г.). «Значение множественных мутаций при раке». Канцерогенез. 21 (3): 379–385. Дои:10.1093 / carcin / 21.3.379. PMID 10688858.
- ^ Лодиш, Харви; Берк, Арнольд; Kaiser, Chris A .; Кригер, Монти; Бретчер, Энтони; Плоег, Хидде; Амон, Анжелика; Мартин, Келси К. (2016). Молекулярная клеточная биология (8-е изд.). Нью-Йорк: В. Х. Фриман и компания. п. 203. ISBN 978-1-4641-8339-3.
- ^ Hu, H; Гатти, РА (июнь 2011 г.). «МикроРНК: новые игроки в реакции на повреждение ДНК». J Mol Cell Biol. 3 (3): 151–8. Дои:10.1093 / jmcb / mjq042. ЧВК 3104011. PMID 21183529.
- ^ Джасперсон К.В., Туохи Т.М., Некласон Д.В., Берт Р.В. (июнь 2010 г.). «Наследственный и семейный рак толстой кишки». Гастроэнтерология. 138 (6): 2044–58. Дои:10.1053 / j.gastro.2010.01.054. ЧВК 3057468. PMID 20420945.
- ^ Truninger K, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO, Bannwart F, Yurtsever H, Neuweiler J, Riehle HM, Cattaruzza MS, Heinimann K, Schär P, Jiricny J, Marra G (май 2005 г.). «Иммуногистохимический анализ показывает высокую частоту дефектов PMS2 при колоректальном раке». Гастроэнтерология. 128 (5): 1160–71. Дои:10.1053 / j.gastro.2005.01.056. PMID 15887099.
- ^ Валери Н., Гаспарини П., Фаббри М., Бракони К., Веронезе А., Ловат Ф, Адаир Б., Ваннини И., Фанини Ф., Боттони А., Костинян С., Сандху С.К., Нуово Г.Дж., Ольха Х, Гафа Р., Калор Ф, Феррацин М. , Lanza G, Volinia S, Negrini M, McIlhatton MA, Amadori D, Fishel R, Croce CM (апрель 2010 г.). «Модуляция репарации несовпадений и стабильности генома с помощью miR-155». Труды Национальной академии наук Соединенных Штатов Америки. 107 (15): 6982–7. Bibcode:2010ПНАС..107.6982В. Дои:10.1073 / pnas.1002472107. ЧВК 2872463. PMID 20351277.
- ^ а б c Чжан В., Чжан Дж., Хоадли К., Кушваха Д., Рамакришнан В., Ли С., Кан Ч., Ю Й, Цзян С., Сон С. В., Цзян Т., Чен СС (июнь 2012 г.). «miR-181d: прогнозирующий биомаркер глиобластомы, который подавляет экспрессию MGMT». Нейроонкология. 14 (6): 712–9. Дои:10.1093 / neuonc / nos089. ЧВК 3367855. PMID 22570426.
- ^ Spiegl-Kreinecker S, Pirker C, Filipits M, Lötsch D, Buchroithner J, Pichler J, Silye R, Weis S, Micksche M, Fischer J, Berger W (январь 2010 г.). «Экспрессия белка O6-метилгуанин-ДНК-метилтрансферазы в опухолевых клетках предсказывает исход терапии темозоломидом у пациентов с глиобластомой». Нейроонкология. 12 (1): 28–36. Дои:10.1093 / neuonc / nop003. ЧВК 2940563. PMID 20150365.
- ^ Сгарра Р., Рустиги А., Тессари М.А., Ди Бернардо Дж., Альтамура С., Фуско А., Манфиолетти Дж., Джанкотти В. (сентябрь 2004 г.). «Ядерные фосфопротеины HMGA и их связь со структурой хроматина и раком». Письма FEBS. 574 (1–3): 1–8. Дои:10.1016 / j.febslet.2004.08.013. PMID 15358530. S2CID 28903539.
- ^ Сюй Ю., Самтер Т.Ф., Бхаттачарья Р., Тесфайе А., Фукс Э.Дж., Вуд Л.Дж., Хусо Д.Л., Ресар Л.М. (май 2004 г.). «Онкоген HMG-I вызывает высоко проникающее агрессивное лимфоидное злокачественное новообразование у трансгенных мышей и сверхэкспрессируется при лейкемии человека». Исследования рака. 64 (10): 3371–5. Дои:10.1158 / 0008-5472.CAN-04-0044. PMID 15150086.
- ^ Borrmann L, Schwanbeck R, Heyduk T., Seebeck B, Rogalla P, Bullerdiek J, Wisniewski JR (декабрь 2003 г.). «Белок группы A2 с высокой подвижностью и его производные связывают конкретную область промотора гена репарации ДНК ERCC1 и модулируют его активность». Исследования нуклеиновых кислот. 31 (23): 6841–51. Дои:10.1093 / нар / gkg884. ЧВК 290254. PMID 14627817.
- ^ Фасиста А., Нгуен Х., Льюис С., Прасад А.Р., Рэмси Л., Зейтлин Б., Нфонсам В., Кроуз Р.С., Бернштейн Х., Пейн С.М., Стерн С., Оатман Н., Банерджи Б., Бернштейн С. (апрель 2012 г.). «Недостаточная экспрессия ферментов репарации ДНК при раннем прогрессировании до спорадического рака толстой кишки». Целостность генома. 3 (1): 3. Дои:10.1186/2041-9414-3-3. ЧВК 3351028. PMID 22494821.
- ^ Чен Дж. Ф., Мерчисон EP, Tang R, Callis TE, Tatsuguchi M, Deng Z, Rojas M, Hammond SM, Schneider MD, Selzman CH, Meissner G, Patterson C, Hannon GJ, Ван ДЗ (февраль 2008 г.). «Целенаправленное удаление Дайсера в сердце приводит к дилатационной кардиомиопатии и сердечной недостаточности». Труды Национальной академии наук Соединенных Штатов Америки. 105 (6): 2111–6. Bibcode:2008PNAS..105.2111C. Дои:10.1073 / pnas.0710228105. ЧВК 2542870. PMID 18256189.
- ^ а б Чжао Ю., Рэнсом Дж. Ф., Ли А., Ведантам В., фон Дрехле М., Мут А. Н., Цучихаши Т., Макманус М. Т., Шварц Р. Дж., Шривастава Д. (апрель 2007 г.). «Нарушение регуляции кардиогенеза, сердечной проводимости и клеточного цикла у мышей, лишенных miRNA-1-2». Ячейка. 129 (2): 303–17. Дои:10.1016 / j.cell.2007.03.030. PMID 17397913.
- ^ Thum T, Galuppo P, Wolf C, Fiedler J, Kneitz S, van Laake LW, Doevendans PA, Mummery CL, Borlak J, Haverich A, Gross C., Engelhardt S, Ertl G, Bauersachs J (июль 2007 г.). «МикроРНК в сердце человека: ключ к репрограммированию генов плода при сердечной недостаточности». Тираж. 116 (3): 258–67. Дои:10.1161 / CIRCULATIONAHA.107.687947. PMID 17606841.
- ^ ван Рой Э., Сазерленд Л. Б., Лю Н., Уильямс А. Х., Маканалли Дж., Джерард Р. Д., Ричардсон Дж. А., Олсон Э. Н. (ноябрь 2006 г.). «Фирменный паттерн стресс-чувствительных микроРНК, которые могут вызывать гипертрофию сердца и сердечную недостаточность». Труды Национальной академии наук Соединенных Штатов Америки. 103 (48): 18255–60. Bibcode:2006ПНАС..10318255В. Дои:10.1073 / pnas.0608791103. ЧВК 1838739. PMID 17108080.
- ^ Тацугути М., Сеок Х.Й., Каллис Т.Е., Томсон Дж. М., Чен Дж. Ф., Ньюман М., Рохас М., Хаммонд С. М., Ван Д. З. (июнь 2007 г.). «Экспрессия микроРНК динамически регулируется во время гипертрофии кардиомиоцитов». Журнал молекулярной и клеточной кардиологии. 42 (6): 1137–41. Дои:10.1016 / j.yjmcc.2007.04.004. ЧВК 1934409. PMID 17498736.
- ^ Чжао Ю., Самал Е., Шривастава Д. (июль 2005 г.). «Сывороточный фактор ответа регулирует мышечно-специфическую микроРНК, которая нацелена на Hand2 во время кардиогенеза». Природа. 436 (7048): 214–20. Bibcode:2005Натура.436..214Z. Дои:10.1038 / природа03817. PMID 15951802. S2CID 4340449.
- ^ Сяо Дж, Ло Х, Линь Х, Чжан И, Лю И, Ван Н, Чжан И, Ян Б., Ван З (апрель 2007 г.). «МикроРНК miR-133 подавляет экспрессию K + -каналов HERG, способствуя удлинению интервала QT в сердцах больных диабетом». Журнал биологической химии. 282 (17): 12363–7. Дои:10.1074 / jbc.C700015200. PMID 17344217.
- ^ Ян Б., Линь Х, Сяо Дж, Лу И, Ло Х, Ли Б, Чжан И, Сюй Ц, Бай И, Ван Х, Чен Дж, Ван З (апрель 2007 г.). «Специфическая для мышц микроРНК miR-1 регулирует аритмогенный потенциал сердца, воздействуя на GJA1 и KCNJ2». Природа Медицина. 13 (4): 486–91. Дои:10,1038 / нм 1569. PMID 17401374. S2CID 1935811.
- ^ Каре А., Каталуччи Д., Фелисетти Ф., Бончи Д., Аддарио А., Галло П, Банг М. Л., Сегналини П., Гу И, Далтон Н. Д., Элия Л., Латронико М. В., Хёйдал М., Ауторе С., Руссо М. А., Дорн Г. В., Эллингсен О. , Руис-Лозано П., Петерсон К.Л., Кроче С.М., Пешле С., Кондорелли Дж. (Май 2007 г.). «MicroRNA-133 контролирует гипертрофию сердца». Природа Медицина. 13 (5): 613–8. Дои:10,1038 / нм1582. PMID 17468766. S2CID 10097893.
- ^ ван Рой Э., Сазерленд Л. Б., Ци Х, Ричардсон Дж. А., Хилл Дж., Олсон Э. Н. (апрель 2007 г.). «Контроль стресс-зависимого сердечного роста и экспрессии генов с помощью микроРНК». Наука. 316 (5824): 575–9. Bibcode:2007Наука ... 316..575В. Дои:10.1126 / science.1139089. PMID 17379774. S2CID 1927839.
- ^ Keller T, Boeckel JN, Groß S, Klotsche J, Palapies L, Leistner D, et al. (Июль 2017 г.). «Улучшенная стратификация риска при профилактике с помощью панели выбранных циркулирующих микроРНК». Научные отчеты. 7 (1): 4511. Bibcode:2017НатСР ... 7,45 11K. Дои:10.1038 / s41598-017-04040-w. ЧВК 5495799. PMID 28674420.
- ^ Insull W (январь 2009 г.). «Патология атеросклероза: образование бляшек и реакция бляшек на лечение». Американский журнал медицины. 122 (1 приложение): S3 – S14. Дои:10.1016 / j.amjmed.2008.10.013. PMID 19110086.
- ^ а б c d е ж г час я j k л м п о п q Сон DJ, Кумар С., Такабе В., Ким К.В., Ни К.В., Альбертс-Гриль Н., Чан И.Х., Ким С., Ким В., Вон Кан С., Бейкер А.Х., Вун Со Дж., Феррара К.В., Чо Х (2013). «Атипичная механочувствительная микроРНК-712, полученная из прерибосомальной РНК, вызывает эндотелиальное воспаление и атеросклероз». Nature Communications. 4: 3000. Bibcode:2013НатКо ... 4.3000S. Дои:10.1038 / ncomms4000. ЧВК 3923891. PMID 24346612.
- ^ а б Басу Р., Фан Д., Кандалам Ви, Ли Дж., Дас С.К., Ван Х, Болдуин Т.А., Аудит Г.Ю., Кассири З. (декабрь 2012 г.). «Утрата гена Timp3 приводит к образованию аневризмы брюшной аорты в ответ на ангиотензин II». Журнал биологической химии. 287 (53): 44083–96. Дои:10.1074 / jbc.M112.425652. ЧВК 3531724. PMID 23144462.
- ^ Либби П. (2002). «Воспаление при атеросклерозе». Природа. 420 (6917): 868–74. Bibcode:2002Натурал.420..868Л. Дои:10.1038 / природа01323. PMID 12490960. S2CID 407449.
- ^ а б Phua YL, Chu JY, Marrone AK, Bodnar AJ, Sims-Lucas S, Ho J (октябрь 2015 г.). «Почечные стромальные миРНК необходимы для нормального нефрогенеза и выживания мезангиальных клубочков». Физиологические отчеты. 3 (10): e12537. Дои:10.14814 / phy2.12537. ЧВК 4632944. PMID 26438731.
- ^ Маес О.К., Чертков Х.М., Ван Э., Шиппер Х.М. (май 2009 г.). «МикроРНК: последствия для болезни Альцгеймера и других заболеваний ЦНС человека». Текущая геномика. 10 (3): 154–68. Дои:10.2174/138920209788185252. ЧВК 2705849. PMID 19881909.
- ^ Амин Н.Д., Бай Дж., Клуг Дж. Р., Бонаноми Д., Панкрац М. Т., Гиффорд В. Д., Хинкли, Калифорния, Стернфельд М.Дж., Дрисколл С.П., Домингес Б., Ли К.Ф., Джин Х, Пфафф С.Л. (декабрь 2015 г.). «Потеря мотонейрон-специфической микроРНК-218 вызывает системную нервно-мышечную недостаточность». Наука. 350 (6267): 1525–9. Bibcode:2015Научный ... 350.1525А. Дои:10.1126 / science.aad2509. ЧВК 4913787. PMID 26680198.
- ^ Schratt G (декабрь 2009 г.). «микроРНК в синапсе». Обзоры природы. Неврология. 10 (12): 842–9. Дои:10.1038 / nrn2763. PMID 19888283. S2CID 3507952.
- ^ Хоссейнян С, Арефиан А, Рахш-Хоршид Х (2020). «Мета-анализ данных по экспрессии генов подчеркивает синаптическую дисфункцию в гиппокампе мозга при болезни Альцгеймера». Научные отчеты. 10 (1): 8384. Дои:10.1038 / s41598-020-64452-z. ЧВК 7239885. PMID 32433480.
- ^ Hommers LG, Domschke K, Deckert J (январь 2015 г.). «Неоднородность и индивидуальность: микроРНК при психических расстройствах». Журнал нейронной передачи. 122 (1): 79–97. Дои:10.1007 / s00702-014-1338-4. PMID 25395183. S2CID 25088900.
- ^ Фэн Дж., Сун Дж., Ян Дж., Нолтнер К., Ли В., Бузин С.Х., Лонгмат Дж., Хестон Л.Л., Росси Дж., Соммер СС (июль 2009 г.). Reif A (ред.). «Доказательства Х-хромосомной шизофрении, связанной с изменениями микроРНК». PLOS ONE. 4 (7): e6121. Bibcode:2009PLoSO ... 4.6121F. Дои:10.1371 / journal.pone.0006121. ЧВК 2699475. PMID 19568434.
- ^ Беверидж, штат Нью-Джерси, Гардинер Э, Кэрролл А.П., Туни, штат Пенсильвания, Кэрнс, М.Дж. (декабрь 2010 г.). «Шизофрения связана с увеличением биогенеза кортикальных микроРНК». Молекулярная психиатрия. 15 (12): 1176–89. Дои:10.1038 / mp.2009.84. ЧВК 2990188. PMID 19721432.
- ^ "Факты об инсульте | cdc.gov". www.cdc.gov. 15 марта 2019 г.. Получено 5 декабря 2019.
- ^ Каток, Кэмерон; Ханна, Савита (май 2011 г.). «МикроРНК в этиологии и патологии ишемического инсульта». Физиологическая геномика. 43 (10): 521–528. Дои:10.1152 / физиолгеномика.00158.2010. ISSN 1094-8341. ЧВК 3110894. PMID 20841499.
- ^ Оуян, И-Бин; Старый, Крид М .; Ян, Го-Юань; Жиффар, Рона (1 января 2013 г.). «микроРНК: инновационные мишени для церебральной ишемии и инсульта». Текущие цели в отношении лекарств. 14 (1): 90–101. Дои:10.2174/138945013804806424. ISSN 1389-4501. ЧВК 3673881. PMID 23170800.
- ^ а б c Леволь Дж. М., Нуньес Ю. О., Додд П. Р., Тивари Г. Р., Харрис Р. А., Мэйфилд Р. Д. (ноябрь 2011 г.). «Повышение регуляции микроРНК в мозгу алкоголиков». Алкоголизм, Клинические и экспериментальные исследования. 35 (11): 1928–37. Дои:10.1111 / j.1530-0277.2011.01544.x. ЧВК 3170679. PMID 21651580.
- ^ Тапоцик Дж. Д., Соломон М., Фланиган М., Мейнхардт М., Барбье Э., Шанк Дж. Р., Швандт М., Зоммер У. С., Хайлиг М. (июнь 2013 г.). «Скоординированная дисрегуляция мРНК и микроРНК в медиальной префронтальной коре головного мозга крыс после алкогольной зависимости». Журнал фармакогеномики. 13 (3): 286–96. Дои:10.1038 / tpj.2012.17. ЧВК 3546132. PMID 22614244.
- ^ Горини Г., Нуньес Ю.О., Мэйфилд РД (2013). «Интеграция miRNA и профилирование белков выявляют скоординированные нейроадаптации в мозге зависимых от алкоголя мышей». PLOS ONE. 8 (12): e82565. Bibcode:2013PLoSO ... 882565G. Дои:10.1371 / journal.pone.0082565. ЧВК 3865091. PMID 24358208.
- ^ Tapocik JD, Barbier E, Flanigan M, Solomon M, Pincus A, Pilling A, Sun H, Schank JR, King C, Heilig M (март 2014 г.). «МикроРНК-206 в медиальной префронтальной коре головного мозга крыс регулирует экспрессию BDNF и употребление алкоголя». Журнал неврологии. 34 (13): 4581–8. Дои:10.1523 / JNEUROSCI.0445-14.2014. ЧВК 3965783. PMID 24672003.
- ^ Липпаи Д., Бала С., Чак Т., Курт-Джонс Е.А., Сабо Г. (2013). «Хроническая индуцированная алкоголем микроРНК-155 способствует нейровоспалению TLR4-зависимым образом у мышей». PLOS ONE. 8 (8): e70945. Bibcode:2013PLoSO ... 870945L. Дои:10.1371 / journal.pone.0070945. ЧВК 3739772. PMID 23951048.
- ^ а б Ли Дж, Ли Дж, Лю Х, Цинь С., Гуань И, Лю И, Ченг И, Чен Х, Ли В, Ван С., Сюн М., Кужикандатхил Э.В., Йе Дж. Х., Чжан Ц. (сентябрь 2013 г.). «Профиль экспрессии микроРНК и функциональный анализ показывают, что miR-382 является критически новым геном алкогольной зависимости». EMBO Молекулярная медицина. 5 (9): 1402–14. Дои:10.1002 / emmm.201201900. ЧВК 3799494. PMID 23873704.
- ^ Ромао Дж. М., Джин В., Додсон М. В., Хаусман Г. Дж., Мур СС, Гуань Л. Л. (сентябрь 2011 г.). «Регуляция микроРНК в адипогенезе млекопитающих». Экспериментальная биология и медицина. 236 (9): 997–1004. Дои:10.1258 / ebm.2011.011101. PMID 21844119. S2CID 30646787.
- ^ Skårn M, Namløs HM, Noordhuis P, Wang MY, Meza-Zepeda LA, Myklebost O (апрель 2012 г.). «Дифференцировка адипоцитов стромальных клеток, происходящих из костного мозга человека, модулируется микроРНК-155, микроРНК-221 и микроРНК-222». Стволовые клетки и развитие. 21 (6): 873–83. Дои:10.1089 / scd.2010.0503. HDL:10852/40423. PMID 21756067.
- ^ Цзо И, Цян Л., фермер SR (март 2006 г.). «Активация экспрессии CCAAT / энхансер-связывающего белка (C / EBP) альфа с помощью C / EBP beta во время адипогенеза требует гамма-ассоциированной репрессии HDAC1 на промоторе гена C / ebp альфа, активируемого пролифератором пероксисом». Журнал биологической химии. 281 (12): 7960–7. Дои:10.1074 / jbc.M510682200. PMID 16431920.
- ^ Джун-Хао Э., Гупта Р. Р., Ших-Чанг Н. (март 2016 г.). «Lin28 и let-7 в метаболической физиологии старения». Тенденции в эндокринологии и метаболизме. 27 (3): 132–141. Дои:10.1016 / j.tem.2015.12.006. PMID 26811207. S2CID 3614126.
- ^ Чжу Х., Ших-Чанг Н., Сегре А.В., Шинода Дж., Шах С.П., Эйнхорн В.С., Такеучи А., Энгрейц Дж. М., Хаган Дж. П., Харас М. Г., Урбах А., Торнтон Дж. Е., Трибулет Р., Грегори Р. И., Альтшулер Д., Дейли Г. К. ( Сентябрь 2011 г.). «Ось Lin28 / let-7 регулирует метаболизм глюкозы». Ячейка. 147 (1): 81–94. Дои:10.1016 / j.cell.2011.08.033. ЧВК 3353524. PMID 21962509.
- ^ Frost RJ, Olson EN (декабрь 2011 г.). «Контроль гомеостаза глюкозы и чувствительности к инсулину с помощью семейства микроРНК Let-7». Труды Национальной академии наук Соединенных Штатов Америки. 108 (52): 21075–80. Bibcode:2011PNAS..10821075F. Дои:10.1073 / pnas.1118922109. ЧВК 3248488. PMID 22160727.
- ^ Теруэль-Монтойя Р., Розендал FR, Мартинес С. (февраль 2015 г.). «МикроРНК в гемостазе». Дж. Тромб Хемост. 13 (2): 170–181. Дои:10.1111 / jth.12788. PMID 25400249.
- ^ а б Nourse J, Braun J, Lackner K, Hüttelmaier S, Danckwardt S (сентябрь 2018 г.). «Крупномасштабная идентификация функционального нацеливания на микроРНК выявляет кооперативную регуляцию гемостатической системы». Дж. Тромб Хемост. 16 (11): 2233–2245. Дои:10.1111 / jth.14290. PMID 30207063.
- ^ Nourse J, Danckwardt S (сентябрь 2020 г.). «Новое обоснование для нацеливания на FXI: выводы из гемостатического таргетинга микроРНК для новых антикоагулянтных стратегий». Pharmacol Ther. Дои:10.1016 / j.pharmthera.2020.107676. PMID 32898547.
- ^ Фазан М., Мэттик Дж. С. (сентябрь 2007 г.). «Повышение оценки функциональных последовательностей человека». Геномные исследования. 17 (9): 1245–53. Дои:10.1101 / гр.6406307. PMID 17690206.
- ^ Бертоне П., Столк В., Ройс Т.Э., Розовски Д.С., Урбан А.Е., Чжу Х, Ринн Д.Л., Тонгпразит В., Саманта М., Вайсман С., Герштейн М., Снайдер М. (декабрь 2004 г.). «Глобальная идентификация транскрибируемых последовательностей человека с помощью массивов тайлинга генома». Наука. 306 (5705): 2242–6. Bibcode:2004Научный ... 306.2242B. Дои:10.1126 / science.1103388. PMID 15539566. S2CID 396518.
- ^ Ота Т., Сузуки Ю., Нисикава Т., Оцуки Т., Сугияма Т., Ирие Р. и др. (Январь 2004 г.). «Полное секвенирование и характеристика 21 243 полноразмерных кДНК человека». Природа Генетика. 36 (1): 40–5. Дои:10,1038 / ng1285. PMID 14702039.
- ^ Кун Д.Е., Мартин М.М., Фельдман Д.С., Терри А.В., Нуово Г.Дж., Элтон Т.С. (январь 2008 г.). «Экспериментальная проверка мишеней miRNA». Методы. 44 (1): 47–54. Дои:10.1016 / j.ymeth.2007.09.005. ЧВК 2237914. PMID 18158132.
- ^ Hüttenhofer A, Schattner P, Polacek N (май 2005 г.). «Некодирующие РНК: надежда или шумиха?». Тенденции в генетике. 21 (5): 289–97. Дои:10.1016 / j.tig.2005.03.007. PMID 15851066.
- ^ Куреши А., Такур Н., Монга И., Такур А., Кумар М. (1 января 2014 г.). «VIRmiRNA: исчерпывающий ресурс для экспериментально подтвержденных вирусных miRNA и их мишеней». База данных. 2014: bau103. Дои:10.1093 / база данных / bau103. ЧВК 4224276. PMID 25380780.
- ^ Кумар М. «ВИРмиРНК». Ресурс для экспериментальных вирусных miRNA и их мишеней. Центр биоинформатики, CSIR-IMTECH.
- ^ Тудденхэм Л., Юнг Дж. С., Чане-Вун-Мин Б., Дёлькен Л., Пфеффер С. (февраль 2012 г.). «Глубокое секвенирование малых РНК позволяет идентифицировать микроРНК и другие небольшие некодирующие РНК из вируса герпеса человека 6B». Журнал вирусологии. 86 (3): 1638–49. Дои:10.1128 / JVI.05911-11. ЧВК 3264354. PMID 22114334.
- ^ Чжэн Х, Фу Р., Ван Дж. Т., Лю Цюй, Чен Х, Цзян Ю (апрель 2013 г.). «Достижения в методах прогнозирования мишеней микроРНК». Международный журнал молекулярных наук. 14 (4): 8179–87. Дои:10.3390 / ijms14048179. ЧВК 3645737. PMID 23591837.
- ^ Agarwal V, Bell GW, Nam JW, Bartel DP (август 2015 г.). «Предсказание эффективных сайтов-мишеней микроРНК в мРНК млекопитающих». eLife. 4: e05005. Дои:10.7554 / eLife.05005. ЧВК 4532895. PMID 26267216.
дальнейшее чтение
- Определение и классификация miRNA: Амброс В., Бартель Б., Бартель Д. П., Бердж С. Б., Кэррингтон Дж. К., Чен Х, Дрейфус Г., Эдди С. Р., Гриффитс-Джонс С., Маршалл М., Мацке М., Рувкун Г., Тушл Т. (март 2003 г.). «Единая система аннотации микроРНК». РНК. 9 (3): 277–9. Дои:10.1261 / rna.2183803. ЧВК 1370393. PMID 12592000.
- Наука обзор малых РНК: Баулкомб Д. (сентябрь 2002 г.). «События ДНК. Микромир РНК». Наука. 297 (5589): 2002–3. Дои:10.1126 / science.1077906. PMID 12242426. S2CID 82531727.
- Открытие лин-4, первая обнаруженная miRNA: Ли Р.К., Фейнбаум Р.Л., Амброс V (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК с антисмысловой комплементарностью lin-14». Ячейка. 75 (5): 843–54. Дои:10.1016 / 0092-8674 (93) 90529-У. PMID 8252621.
внешние ссылки
- База данных miRBase
- miRTarBase, экспериментально подтвержденная база данных взаимодействий микроРНК-мишень.
- семирна, Веб-приложение для поиска микроРНК в геноме растений.
- ONCO.IO: Интегративный ресурс для анализа микроРНК и факторов транскрипции при раке.
- МирОБ: MicroRNA нацелена на базу данных и инструмент анализа данных и датавиз.
- База данных ChIPBase: База данных открытого доступа для декодирования факторы транскрипции которые участвовали в транскрипции микроРНК или влияли на нее из данных ChIP-seq.
- Анимационный ролик о процессе биогенеза микроРНК.
- реагенты для модуляции miRNA для активации или подавления функции эндогенной зрелой микроРНК
Типы нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновые кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||
| |||||||