Чешуекрылые - Lepidoptera
| Чешуекрылые | |
|---|---|
 | |
| Павлинья бабочка (Аглаис ио ) | |
 | |
| Бабочка сфинкс (Adhemarius gannascus ) | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Класс: | Насекомое |
| (без рейтинга): | Амфиесменоптеры |
| Порядок: | Чешуекрылые Линней, 1758 |
| Подзаказы | |
Чешуекрылые (/ˌлɛпɪˈdɒптərə/ LEP-я-DOP-tər-ə, от Древнегреческий лепис «Масштаб» + птерон «Крыло») является порядок из насекомые это включает бабочки и моль (оба называются чешуекрылые). Около 180 000 виды Lepidoptera описаны в 126 семьи[1] и 46 надсемейства,[2] 10 процентов от общего числа описанных видов живых организмов.[2][3] Это один из самых распространенных и узнаваемых отрядов насекомых в мире.[4] Чешуекрылые демонстрируют множество вариаций основной структуры тела, которые эволюционировали, чтобы получить преимущества в образе жизни и распространении. Недавние оценки показывают, что в этом отряде может быть больше видов, чем считалось ранее.[5] и входит в четверку самых особенный заказы, наряду с Перепончатокрылые, Двукрылые, и Жесткокрылые.[4]
Виды чешуекрылых характеризуются более чем тремя производными признаками. Наиболее очевидным является наличие напольные весы которые охватывают тела, крылья, а хоботок. Чешуя представляет собой видоизмененные, сплющенные «волоски», придающие бабочкам и мотылькам их самые разнообразные цвета и узоры. Почти все виды имеют ту или иную форму перепончатых крыльев, за исключением некоторых, у которых крылья уменьшены или они бескрылые. Спаривание откладка яиц осуществляется взрослыми особями, как правило, вблизи или на растениях-хозяевах личинки. Как и большинство других насекомых, бабочки и моль находятся голометаболический, то есть они проходят полная метаморфоза. Личинок обычно называют гусеницы, и полностью отличаются от их взрослых форм бабочки или бабочки, имея цилиндрическое тело с хорошо развитой головой, ротовыми частями нижней челюсти, тремя парами грудных ног и отсутствием до пяти пар бездельники. По мере роста эти личинки меняют свой внешний вид, проходя ряд стадий, называемых возрастов. После полного созревания личинка развивается в куколка. Некоторые бабочки и многие виды моли плетут шелковые футляры или кокон до окукливания, в то время как другие этого не делают, а уходят в подполье.[4] Куколка бабочки, называемая куколка, имеет твердую кожу, обычно без кокона. После того, как куколка завершает свою метаморфозу, появляется половозрелый взрослый особь.
Чешуекрылые за миллионы лет развили широкий спектр рисунков крыльев и окраски, начиная от серых бабочек, родственных родственным им отрядам. Трихоптера, к ярким бабочкам со сложным узором.[1] Соответственно, это самый известный и популярный из отрядов насекомых, и многие люди занимаются наблюдением, изучением, сбором, выращиванием и торговлей этими насекомыми. Лицо, собирающее или изучающее этот приказ, называется лепидоптеролог.
Бабочки и мотыльки играют важную роль в естественной экосистеме, поскольку опылители и как пища в пищевой цепи; и наоборот, их личинки считаются очень проблематичными для растительности в сельском хозяйстве, поскольку их основным источником пищи часто является живое растительное вещество. У многих видов самка может производить от 200 до 600 яиц, в то время как у других количество может приближаться к 30 000 яиц за один день. Вылупившиеся из этих яиц гусеницы могут нанести ущерб большому количеству сельскохозяйственных культур. Многие виды мотыльков и бабочек представляют экономический интерес в силу своей роли опылителей. шелк они производят, или как вредитель виды.
Этимология
Термин Lepidoptera использовался в 1746 г. Карл Линней в его Фауна Свечица.[6][7] Слово происходит от Греческий λεπίς, ген. λεπίδος ("масштаб ") и πτερόν ("крыло").[8] Иногда термин Ропалоцера используется для клады всех видов бабочек, происходящих из древнегреческого ῥόπαλον (Ропалон)[9]:4150 и κέρας (Керас)[9]:3993 означающие «дубинка» и «рог» соответственно, происходящие от формы усики бабочек.
Происхождение общих имен «бабочка» и «моль» разнообразно и часто неясно. В английский слово бабочка от Древнеанглийский Buttorfleoge, с множеством вариантов написания. В остальном происхождение неизвестно, хотя это могло быть связано с бледно-желтым цветом крыльев многих видов, что предполагает цвет масла.[10][11] Виды Гетероцера обычно называют моль. Происхождение английского слова moth яснее, оно происходит от древнеанглийского] moððe (ср. Нортумбрийский диалект мохэ) из общегерманского (ср. Древнескандинавский мотти, Голландский мот и Немецкий Motte все означает «моль»). Возможно, его происхождение связано с древнеанглийским maða смысл "личинка "или от корня"мошка ", который до 16 века использовался в основном для обозначения личинки, обычно в отношении пожирающей одежды.[12]
Этимологическое происхождение слова «гусеница», личиночной формы бабочек и мотыльков, восходит к началу 16 века, от Средний английский катирпель, катирпеллер, вероятно, переделка Старый северный французский катепелоза (от латинского каттус, "кот" + пилозь, «волосатая»).[13]
Распространение и разнообразие
Чешуекрылые - одна из самых успешных групп насекомых. Встречаются на всех континентах, кроме Антарктида, и населяют все наземные среды обитания, от пустыни до тропических лесов, от низинных лугов до горных плато, но почти всегда связаны с высшими растениями, особенно покрытосеменными (цветущие растения ).[14] Среди самых северных обитающих видов бабочек и мотыльков - арктический Аполлон (Parnassius arcticus ), который находится за Полярным кругом на северо-востоке Якутия, на высоте 1500 м над уровнем моря.[15] в Гималаи, различные виды Аполлонов, такие как Парнасий эпаф были зарегистрированы на высоте до 6000 м над уровнем моря.[16]:221
Некоторые виды чешуекрылых проявляют симбиотический, форетический, или паразитический образ жизни, населяющий тела организмов, а не окружающую среду. Копрофаг пиралид виды бабочек, называемые ленивец, такие как Bradipodicola hahneli и Криптозы choloepi, необычны тем, что они обитают исключительно в мехе ленивцы, млекопитающие, обнаруженные в Центральная и Южная Америка.[17][18]Два вида Tinea Было зарегистрировано, что бабочки питаются роговой тканью и были выведены из рогов крупного рогатого скота. Личинка Зенодохий кокциворелла является внутренним паразитом кокцид Кермес виды. Многие виды были зарегистрированы как размножающиеся в природных материалах или отходах, таких как совиные гранулы, пещеры летучих мышей, соты или больные плоды.[18]
По состоянию на 2007 год было описано около 174 250 видов чешуекрылых, из них бабочки и шкиперы, по оценкам, составляли около 17 950, а мотыльки составляли остальное.[2][19] Подавляющее большинство чешуекрылых обитает в тропиках, но значительное разнообразие существует на большинстве континентов. В Северной Америке обитает более 700 видов бабочек и более 11 000 видов моли,[20][21] в то время как в Австралии зарегистрировано около 400 видов бабочек и 14 000 видов моли.[22] Разнообразие чешуекрылых в каждом фаунистический регион был оценен Джоном Хеппнером в 1991 году на основании частично фактических расчетов из литературы, частично на основе карточных индексов в Музей естественной истории (Лондон) и Национальный музей естественной истории (Вашингтон), и частично по оценкам:[5]
| Палеарктика | Неарктический | Неотропный | Афротропный | Индо-австралийский (включая Индомалайский, Австралийский, и Океанские царства ) | |
|---|---|---|---|---|---|
| Предполагаемое количество видов | 22,465 | 11,532 | 44,791 | 20,491 | 47,287 |
Внешняя морфология
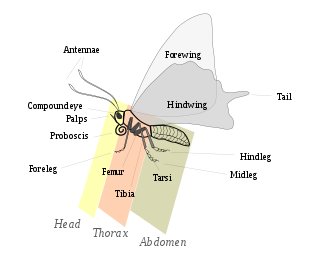
Lepidoptera морфологически отличается от других отрядов главным образом наличием напольные весы на внешних частях тела и придатках, особенно на крылья. Бабочки и мотыльки различаются по размеру от микролепидоптеры всего несколько миллиметров в длину, заметным животным с размахом крыльев более 25 сантиметров, таким как Птичье крыло королевы Александры и Атласский мотылек.[23]:246Чешуекрылые проходят четырехступенчатую жизненный цикл: яйцо; личинка или гусеница; куколка или куколка; и имаго (множественное число: воображает) / взрослые и демонстрируют множество вариаций основной структуры тела, которые дают этим животным преимущества для различных стилей жизни и окружающей среды.
Голова

Голова - это место, где находятся многие органы чувств и части рта. Как и у взрослой особи, личинка также закалена, или склеротизированный головная капсула.[24] Здесь два сложные глаза, и хетоземавстречаются выпуклые пятна или скопления сенсорных щетинок, уникальные для чешуекрылых, хотя многие таксоны утратили одно или оба этих пятна. Антенны имеют широкую вариацию по форме у разных видов и даже у разных полов. Усики бабочек обычно нитевидные и имеют форму булав, у шкиперов крючковатые, а у бабочек членики жгутиков могут быть увеличены или разветвлены. У некоторых бабочек есть увеличенные усики или усики сужающиеся и крючковатые на концах.[25]:559–560
В верхнечелюстные галеи видоизменяются и образуют удлиненный хоботок. Хоботок состоит из одного-пяти сегментов, обычно свернутых под головой небольшими мышцами, когда он не используется для всасывания нектара из цветов или других жидкостей. Немного базальный мотыльки все еще есть челюсти, или отдельные подвижные челюсти, как у их предков, и они образуют семью Micropterigidae.[24][25]:560[26]
Личинки, называемые гусеницы, имеют упрочненную головную капсулу. Гусеницы лишены хоботка и имеют раздельное жевание. ротовые органы.[24] Эти ротовые аппараты, называемые челюсти, используются для пережевывания растительного вещества, которое поедают личинки. Нижняя челюсть или губа слабая, но может нести фильера, орган, используемый для создания шелка. Голова состоит из больших боковых долей, каждая из которых имеет до шести простых глазков в форме эллипса.[25]:562–563
Грудная клетка
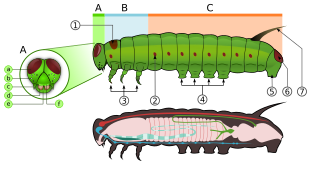
Грудная клетка состоит из трех сросшихся сегментов: переднегрудь, среднегрудь, и заднегрудь, каждый с парой ног. Первый сегмент содержит первую пару ног. У некоторых самцов семейства бабочек Нимфалиды, передние конечности сильно уменьшены и не используются для ходьбы или сидения.[25]:586 Три пары ног покрыты чешуей. У чешуекрылых также есть органы обоняния на ногах, которые помогают бабочке «пробовать» или «нюхать» пищу.[27] У личиночной формы есть 3 пары настоящих ножек, до 11 пар брюшных ножек (обычно восемь) и крючков, называемых апикальными крючками.[14]
Две пары крыльев находятся на среднем и третьем сегментах, или среднегрудь и заднегрудь соответственно. У более поздних родов крылья второго сегмента гораздо более выражены, хотя некоторые более примитивные формы имеют крылья обоих сегментов одинакового размера. Крылья покрыты чешуей, напоминающей черепицу, которые образуют необычайное разнообразие цветов и рисунков. Среднегрудь имеет более мощные мускулы, чтобы двигать моль или бабочку по воздуху, при этом крыло этого сегмента (переднее крыло) имеет более прочную структуру вен.[25]:560 Самое крупное суперсемейство Совки крылья изменены, чтобы действовать как барабанные перепонки или органы слуха.[28]
Гусеница имеет удлиненное мягкое тело, которое может иметь волосовидные или другие выступы, три пары настоящих ног, от одной до 11 пар брюшных ног (обычно восемь) и крючки, называемые апикальными крючками.[14] Грудная клетка обычно имеет по паре ножек на каждом сегменте. Грудная клетка также выстлана множеством дыхалец как на среднегруди, так и на заднегруди, за исключением нескольких водных видов, которые вместо этого имеют форму жабры.[25]:563
Брюшная полость

Брюшная полость, которая менее склеротизирована, чем грудная клетка, состоит из 10 сегментов с перепонками между ними, обеспечивающими шарнирные движения. Грудина на первом сегменте у одних семей мала, а в других полностью отсутствует. Последние два или три сегмента образуют внешние части половых органов вида. В гениталии Lepidoptera очень разнообразны и часто являются единственным средством различения видов. Мужские гениталии включают вальва, который обычно большого размера, так как используется для захвата самки во время спаривания. Женские гениталии состоят из трех отдельных частей.
У самок базальных бабочек только один половой орган, который используется для совокупление и как яйцеклад, или орган яйцекладки. Около 98% видов бабочек имеют отдельный орган для спаривания и внешний проток, по которому идет сперма самца.[25]:561
Брюшко гусеницы имеет четыре пары ложноножек, обычно расположенных на третьем-шестом сегментах брюшка, и отдельную пару ложноножек у ануса, на которых есть пара крошечных крючков, называемых крючками. Они помогают в захвате и ходьбе, особенно у видов, у которых отсутствует много ложноногих (например, личинок Geometridae ). У некоторых базальных бабочек эти ложноножки могут быть на каждом сегменте тела, в то время как ложноножки могут полностью отсутствовать в других группах, которые более приспособлены к свертыванию и жизни в песке (например, Prodoxidae и Nepticulidae соответственно).[25]:563
Напольные весы

Крылья, голова, а также части грудной клетки и брюшка чешуекрылых покрыты мелкой чешуей, от чего отряд получил свое название. Большинство шкал пластинчатый, или лезвие, и прикреплены к ножке, в то время как другие формы могут быть похожими на волосы или специализироваться как вторичные половые признаки.[29]
Просвет или поверхность ламели имеет сложное строение. Придает цвет либо цветным пигменты он содержит или через структурная окраска с механизмами, которые включают фотонные кристаллы и дифракционные решетки.[30]
Весы служат для изоляции, терморегуляции, производства феромоны (только у мужчин ),[31] и помощь в планирующем полете, но самое главное - большое разнообразие ярких или нечетких узоры они обеспечивают, которые помогают организму защитить себя камуфляж или мимикрия, и которые действуют как сигналы другим животным, включая соперников и потенциальные партнеры.[29]




Внутренняя морфология
Репродуктивная система
в репродуктивная система бабочек и моли, мужчина гениталии сложны и неясны. У самок три типа гениталий основаны на соответствующих таксонах: «монотризный», «экзопорический» и «дитризовый». В монотризном типе есть отверстия на сросшихся сегментах 9 и 10 грудины, которые служат для осеменения и откладки яиц. В экзопорианском типе (в Гепиалоидея и Mnesarchaeoidea ) - это два разных места для осеменения и откладки яиц, оба они находятся на той же груди, что и монотризный тип, то есть 9 и 10.[23] У дитризиевых групп есть внутренний канал, по которому проходит сперма, с отдельными отверстиями для совокупления и откладки яиц.[4] У большинства видов гениталии окружены двумя мягкими лопастями, хотя у некоторых видов они могут быть специализированы и склеротизированы для откладки яиц в таких областях, как щели и внутренняя ткань растений.[23] Гормоны и железы, которые их вырабатывают, управляют развитием бабочек и мотыльков по мере их жизненного цикла, называемого эндокринная система. Первый гормон насекомых проторакотропный гормон (PTTH) управляет жизненным циклом вида и диапауза.[32] Этот гормон вырабатывается Корпуса аллата и Корпуса Cardiaca, где он также хранится. Некоторые железы специализируются на выполнении определенных задач, таких как производство шелка или слюноотделение в пальпах.[33]:65, 75 В то время как сердечные тела производят ПТТГ, аллатные тела также производят ювенильные гормоны, а проторозные железы производят гормоны линьки.
Пищеварительная система
в пищеварительная система, передняя часть передней кишки была изменена, чтобы сформировать глоточный всасывающий насос, так как он нужен им для еды, которую они едят, которая по большей части является жидкостью. An пищевод следует и ведет к задней части глотки и у некоторых видов образует форму урожая. Средняя кишка короткая и прямая, а задняя длиннее и извилистая.[23] Предки чешуекрылых, происходящие от Перепончатокрылые, имел слепые кишки средней кишки, хотя они теряются у современных бабочек и мотыльков. Вместо этого все пищеварительные ферменты, кроме начального пищеварения, иммобилизуются на поверхности клеток средней кишки. У личинок длинношеие и черешковые бокаловидные клетки находятся в переднем и заднем отделах средней кишки соответственно. У насекомых бокаловидные клетки выделяют положительный калий ионы, которые поглощаются листьями, попадающими в организм личинок. Большинство бабочек и мотыльков демонстрируют обычный пищеварительный цикл, но виды с разными диетами требуют адаптации, чтобы соответствовать этим новым требованиям.[25]:279

Сердечно-сосудистая система
в сердечно-сосудистая система, гемолимфа, или кровь насекомых, используется для циркуляции тепла в форме терморегуляция, где сокращение мышц производит тепло, которое передается остальному телу в неблагоприятных условиях.[34]У чешуекрылых гемолимфа циркулирует по венам крыльев с помощью пульсирующего органа в той или иной форме, либо через сердце, либо за счет поступления воздуха в дыхательные пути. трахея.[33]:69
Дыхательная система
Воздух всасывается через дыхальца по бокам брюшной полости и грудной клетки, снабжая трахею кислородом, когда он проходит через чешуйки. дыхательная система. Три разных трахеи поставляют и распространяют кислород по телу вида. Дорсальные трахеи снабжают кислородом спинную мускулатуру и сосуды, в то время как брюшные трахеи снабжают вентральную мускулатуру и нервный шнур, а висцеральные трахеи снабжают кишечник, жировые тела и гонады.[33]:71, 72
Полиморфизм


Полиморфизм - это появление форм или «морфов», которые различаются по цвету и количеству признаков в пределах одного вида.[14]:163[35] У чешуекрылых полиморфизм можно наблюдать не только между особями в популяции, но и между полами как половой диморфизм, между географически разделенными популяциями в географический полиморфизм, и между поколениями, летающими в разное время года (сезонный полиморфизм или полифенизм ). У некоторых видов полиморфизм ограничен одним полом, обычно самкой. Это часто включает в себя феномен мимикрия когда миметические морфы летают вместе с немиметическими морфами в популяции определенного вида. Полиморфизм возникает как на определенном уровне с наследственными вариациями в общих морфологических адаптациях особей, так и в определенных специфических морфологических или физиологических признаках внутри вида.[14]
Полиморфизм окружающей среды, в котором черты не передаются по наследству, часто называют полифенизмом, который у чешуекрылых обычно проявляется в виде сезонных морфов, особенно в семействе бабочек Нимфалиды и Pieridae. Пирожная бабочка Старого Света, обычная трава желтая (Eurema Hecabe ) имеет более темную летнюю взрослую форму, вызываемую длинным днем, продолжительностью более 13 часов, в то время как более короткий дневной период в 12 часов или меньше вызывает более бледную форму в послемуссонный период.[36] Полифенизм также встречается у гусениц, например, у пяденицы, Biston Betularia.[37]
Географическая изоляция вызывает расхождение вида в разные морфы. Хороший пример - индийский белый адмирал Прокрисный лимфит, который имеет пять форм, каждая из которых географически отделена от другой большими горными хребтами.[38]:26 Еще более впечатляющим примером географического полиморфизма является Бабочка Аполлон (Парнасий аполлон). Поскольку аполлоны живут небольшими местными популяциями, не контактируя друг с другом, в сочетании с их сильной стенотопической природой и слабой миграционной способностью, скрещивание между популяциями одного вида практически не происходит; благодаря этому они образуют более 600 различных морфов, размер пятен на крыльях которых сильно варьируется.[39]


Половой диморфизм - это проявление различий между самцами и самками у одного вида. У чешуекрылых она широко распространена и почти полностью определяется генетической детерминацией.[36] Половой диморфизм присутствует во всех семействах Papilionoidea и более заметен у Lycaenidae, Pieridae, и некоторые таксоны Нимфалиды. Помимо цветовых вариаций, которые могут отличаться от незначительных до совершенно разных сочетаний цветовых узоров, также могут присутствовать вторичные половые признаки.[38]:25 Различные генотипы, поддерживаемые естественным отбором, также могут экспрессироваться одновременно.[36] Полиморфные и / или миметические самки встречаются у некоторых таксонов в Papilionidae в первую очередь для получения уровня защиты, недоступного самцам их вида. Наиболее явным случаем полового диморфизма является диморфизм взрослых самок многих Psychidae виды, у которых есть только рудиментарные крылья, ноги и ротовой аппарат, по сравнению со взрослыми самцами, которые являются сильными летчиками с хорошо развитыми крыльями и перистыми усиками.[40]
Размножение и развитие

Виды Lepidoptera подвергаются голометаболизм или «полная метаморфоза». Их жизненный цикл обычно состоит из яйцо, а личинка, а куколка, и имаго или взрослый.[14] Личинок обычно называют гусеницы, а куколки моли, заключенные в шелк, называются коконы, а непокрытые куколки бабочек называют хризалиды.
Чешуекрылые в диапаузе
Если вид не размножается круглый год, бабочка или моль могут проникать диапауза, состояние покоя, позволяющее насекомому выживать в неблагоприятных условиях окружающей среды.
Спаривание
Самцы обычно начинают восторг (вылет) раньше самок и пик численности раньше самок. К моменту эклозии оба пола половозрелы.[25]:564 Бабочки и мотыльки обычно не общаются друг с другом, за исключением мигрирующих видов, оставаясь относительно асоциальными. Спаривание начинается с того, что взрослый (самка или самец) привлекает партнера, обычно используя зрительные стимулы, особенно в дневной вид как большинство бабочек. Однако самки большинства ночных видов, в том числе почти всех видов бабочек, используют феромоны для привлечения самцов, иногда с больших расстояний.[14] Некоторые виды участвуют в акустических ухаживаниях или привлекают партнеров с помощью звука или вибрации, например, осиновая моль в горошек, Syntomeida epilais.[41]
Адаптации включают прохождение одного, двух или даже более сезонных поколений, называемых вольтинизм (Унивольтизм, бивольтизм и поливольтизм соответственно). Большинство чешуекрылых в умеренный климат моновольтинные, тогда как в тропическом климате у большинства из них есть два сезонных выводка. Некоторые другие могут воспользоваться любой возможностью и спариваться постоянно в течение года. Эти сезонные адаптации контролируются гормонами, и эти задержки в воспроизводстве называются диапауза.[25]:567 Многие виды чешуекрылых после спаривания и откладывания яиц вскоре умирают, прожив всего несколько дней после эклозии. Другие могут оставаться активными в течение нескольких недель, а затем перезимовать и снова стать сексуально активными, когда погода станет более благоприятной или диапаузой. Сперма самца, который в последний раз спаривался с самкой, скорее всего, оплодотворила яйцеклетку, но сперма от предыдущего спаривания все еще может преобладать.[25]:564
Жизненный цикл

Яйца
Чешуекрылые обычно размножаются половым путем и яйцекладущий (откладка яиц), хотя некоторые виды демонстрируют живорождение в процессе, называемом яйцеклады. Разнообразие различий в яйцо -кладка и количество отложенных яиц. Некоторые виды просто бросают яйца в полете (у этих видов обычно есть многоядные личинки, то есть они едят различные растения, например, гепиалиды и немного нимфалиды )[42] в то время как большинство откладывают яйца рядом с растением-хозяином, которым питаются личинки, или на нем. Количество отложенных яиц может варьироваться от нескольких до нескольких тысяч.[14] Самки бабочек и мотыльков инстинктивно выбирают растение-хозяин, в первую очередь, с помощью химических сигналов.[25]:564
Яйца получены из материалов, проглоченных личинкой, а у некоторых видов - из сперматофоров, полученных от самцов во время спаривания.[43] Яйцо может составлять только 1/1000 массы самки, но она может откладывать яйца до своей собственной массы. Самки с возрастом откладывают яйца меньшего размера. Самки большего размера откладывают яйца большего размера.[44] Яйцо покрыто жестким защитным внешним слоем скорлупы, называемым хорион. Он покрыт тонким слоем воск, что предотвращает высыхание яйца. Каждое яйцо содержит несколько микропиле, или крошечные отверстия в форме воронки на одном конце, цель которых - позволить сперме проникать в яйцеклетку и оплодотворять ее. Яйца бабочек и моли сильно различаются по размеру у разных видов, но все они либо сферические, либо яйцевидные.
Стадия яйца у большинства бабочек длится несколько недель, но яйца откладываются до зимы, особенно у бабочек. умеренные регионы, проходить через диапауза, а вылупление может быть отложено до весны. Другие бабочки могут откладывать яйца весной и вылупляться летом. Эти бабочки обычно относятся к умеренным видам (например, Nymphalis antiopa ).
Личинки

Личинки или гусеницы - первая стадия жизненного цикла после вылупления. Гусеницы, "характерные" полипод личинки с цилиндрическим телом, короткими грудными ногами и брюшными ложноножками (ложноножками) ».[45] У них закаленные (склеротизированный ) головная капсула с адфронтальным швом, образованным медиальным сращением склеритов, нижние челюсти (ротовой аппарат) для жевания, и мягкое трубчатое сегментированное тело, которое может иметь волосовидные или другие выступы, три пары настоящих ножек и дополнительные бездельники (до пяти пар).[46] Тело состоит из тринадцати сегментов, из которых три грудных и десять брюшных.[47] Большинство личинок травоядные животные, но некоторые из них плотоядные животные (некоторые едят муравьев или других гусениц) и детритофаги.[46]
Различные травоядные виды приспособились питаться каждой частью растения и обычно считаются вредителями своих растений-хозяев; Было обнаружено, что некоторые виды откладывают яйца на плоды, а другие виды откладывают яйца на одежду или мех (например, Тинеола Бисселлиелла, моль обыкновенная). Некоторые виды плотоядны, а другие даже паразитируют. Немного Lycaenid такие виды, как Phengaris rebeli находятся социальные паразиты из Мирмика муравьиные гнезда.[48] Вид Geometridae от Гавайи есть плотоядные личинки, которые ловят и поедают мух.[49] Некоторые гусеницы пиралид водные.[50]
Личинки быстро развиваются, несколько поколений в год; однако для развития некоторых видов может потребоваться до 3 лет, и исключительные примеры, такие как Gynaephora groenlandica займет целых семь лет.[14] На личиночной стадии происходят стадии питания и роста, а личинки периодически подвергаются гормонально-индуцированной стадии. шелушение, развиваясь с каждым возраст, пока они не пройдут заключительную личино-кукольную линьку.
Личинки как бабочек, так и мотыльков демонстрируют мимику, чтобы отпугивать потенциальных хищников. У некоторых гусениц есть способность надувать части головы, чтобы казаться змееподобными. У многих есть ложные пятна на глазах, чтобы усилить этот эффект. У некоторых гусениц есть особые конструкции, называемые осметерия (семья Papilionidae ), которые подвергаются воздействию дурно пахнущих химикатов, используемых в обороне. В растениях-хозяевах часто содержатся токсичные вещества, и гусеницы способны улавливать эти вещества и удерживать их во взрослой стадии. Это делает их неприятными для птиц и других хищников. Такая неприятность рекламируется с использованием ярких красных, оранжевых, черных или белых предупреждающих цветов. Ядовитые химические вещества в растениях часто вырабатываются специально, чтобы предотвратить их поедание насекомыми. Насекомые, в свою очередь, разрабатывают контрмеры или используют эти токсины для собственного выживания. Эта «гонка вооружений» привела к совместной эволюции насекомых и растений-хозяев.[51]
Развитие крыла
Форма крыльев личинки не видна снаружи, но когда личинки рассекаются, развивающиеся крылья можно увидеть как диски, которые можно найти на втором и третьем грудных сегментах, вместо дыхальцев, которые видны на брюшных сегментах. Диски крыльев развиваются вместе с трахеей, которая проходит вдоль основания крыла и окружена тонкой периподиальной мембраной, которая связана с внешним эпидермисом личинки крошечным протоком. Диски крыльев очень малы до последнего возраста личинок, когда они резко увеличиваются в размерах, и не затрагиваются ветвлением. трахеи от основания крыла, которые предшествуют образованию жилок крыла, и начинают развиваться узоры, связанные с несколькими ориентирами крыла.[52]
Вблизи окукливания крылья вытесняются за пределы эпидермиса под давлением гемолимфа, и хотя они изначально довольно гибкие и хрупкие, к тому времени, когда куколка отрывается от кутикулы личинки, они плотно прилегают к внешней кутикуле куколки (в куколках obtect). В течение нескольких часов крылья образуют кутикулу, настолько прочную и хорошо прилегающую к телу, что куколки можно брать и брать в руки, не повреждая крылья.[52]
Куколка

Примерно через пять-семь возрастов[53]:26–28 или линьки, определенные гормоны, такие как PTTH, стимулируют выработку экдизон, которая инициирует линьку насекомых. Личинка начинает развиваться в куколка: части тела, характерные для личинки, такие как ножки живота, дегенерируют, в то время как другие, такие как ноги и крылья, подвергаются росту. Найдя подходящее место, животное сбрасывает свою последнюю личиночную кутикулу, обнажая кутикулу куколки под ней.
В зависимости от вида куколка может быть покрыта шелковым коконом, прикреплена к разным типам субстратов, закопана в землю или может быть совсем не покрыта. Особенности имаго внешне узнаваемы в куколке. Все придатки на голове и грудной клетке взрослого человека находятся внутри кутикулы (усики, ротовые органы и т. д.) с обернутыми крыльями, прилегающими к усикам.[25]:564 Куколки одних видов имеют функциональные мандибулы, тогда как куколки других видов не функционируют.[24]
Хотя кутикула куколки сильно склеротизирована, некоторые из нижних сегментов брюшной полости не слиты и могут двигаться с помощью небольших мышц, находящихся между мембранами. Перемещение может помочь куколке, например, избежать солнца, которое в противном случае убило бы ее. Куколка Мексиканская прыгающая фасоль моль (Cydia saltitans ) Является ли это. Личинки прорезают люк в фасоли (виды Себастьяния ) и использовать фасоль как укрытие. При резком повышении температуры куколка внутри дергается и дергается, натягивая нити внутри. Покачивание также может помочь сдерживать паразитоид осы от откладывания яиц на куколку. Другие виды бабочек способны отпугивать хищников щелчками.[25]:564, 566
Время до того, как куколка вырастет (выйдет), сильно варьируется. Бабочка-монарх может оставаться в куколке в течение двух недель, в то время как другим видам может потребоваться оставаться в диапаузе более 10 месяцев. Взрослая особь выходит из куколки либо с помощью брюшных крючков, либо из выступов, расположенных на голове. Челюсти наиболее примитивных семейств моли используются для выхода из кокона (например, Micropterigoidea ).[14][25]:564
Взрослый
Большинство видов чешуекрылых не живут долго после эклозии, им требуется всего несколько дней, чтобы найти себе пару и затем отложить яйца. Другие могут оставаться активными в течение более длительного периода (от одной до нескольких недель) или переживать диапаузу и зимовать, как бабочки-монархи, или переждать экологический стресс. Некоторые взрослые виды микролепидоптер проходят стадию, когда не происходит никакой репродуктивной активности, которая длится летом и зимой с последующим спариванием и откладкой яиц ранней весной.[25]:564
В то время как большинство бабочек и мотыльков земной, многие виды Pyralidae действительно водный все стадии, кроме взрослой особи, находятся в воде. Многие виды из других семейств, такие как Erebidae, Nepticulidae, Космоптеригиды, Tortricidae, Olethreutidae, Совки, Cossidae, и Сфингиды водные или полуводный.[54]:22
Поведение
Рейс
Полет - важный аспект жизни бабочек и мотыльков, он используется для уклонения от хищников, поиска пищи и своевременного поиска партнеров, поскольку большинство видов чешуекрылых не живут долго после эклозии. Это основная форма передвижения у большинства видов. У чешуекрылых передние и задние крылья механически связаны и взмахивают синхронно. Полет антеромоторный или движется в основном за счет движения передних крыльев. Хотя, как сообщается, виды чешуекрылых все еще могут летать, когда им отрезаны задние крылья, это снижает их возможности линейного полета и поворота.[55]
Виды чешуекрылых, чтобы летать, должны быть теплыми, примерно от 77 до 79 ° F (от 25 до 26 ° C). Они зависят от достаточно высокой температуры их тела, и, поскольку они не могут регулировать ее самостоятельно, это зависит от окружающей их среды. Бабочки, живущие в более прохладном климате, могут использовать свои крылья, чтобы согреть свое тело. Они будут греться на солнышке, расправив крылья, чтобы получить максимальное воздействие солнечного света. В более жарком климате бабочки могут легко перегреться, поэтому они обычно активны только в более прохладные части дня, рано утром, поздно вечером или рано вечером. В жару днем отдыхают в тени. Некоторые более крупные бабочки с толстым телом (например, Sphingidae) могут в ограниченной степени генерировать собственное тепло, вибрируя крыльями. Тепло, выделяемое летательными мышцами, нагревает грудную клетку, в то время как температура живота не имеет значения для полета. Чтобы избежать перегрева, некоторые бабочки полагаются на волосатую чешую, внутренние воздушные мешочки и другие структуры, которые разделяют грудную клетку и живот и поддерживают прохладу в животе.[56]
Некоторые виды бабочек могут достигать высоких скоростей, например, южный дротик, который может развивать скорость до 48,4 км / ч. Сфингиды - одни из самых быстро летающих насекомых, некоторые из них способны летать со скоростью более 50 км / ч (30 миль / ч), имея размах крыльев 35–150 мм.[1][57] У некоторых видов иногда существует планирующий компонент их полета. Полет происходит либо в режиме зависания, либо в виде движения вперед или назад.[58] У видов бабочек и моли, таких как ястребиная моль, парение важно, так как они должны сохранять определенную стабильность над цветами при питании нектаром.[1]

Навигация важен для видов Lepidoptera, особенно для мигрирующих. Было показано, что бабочки, у которых больше мигрирующих видов, ориентируются при помощи солнечных компасов с временной компенсацией. Они могут видеть поляризованный свет, поэтому может ориентироваться даже в пасмурную погоду. Поляризованный свет в области, близкой к ультрафиолетовому спектру, считается особенно важным.[59] Большинство мигрирующих бабочек - это те, которые живут в полупустынных районах, где сезон размножения короткий.[60] История жизни их растений-хозяев также влияет на стратегии бабочек.[61] Другие теории включают использование пейзажей. Чешуекрылые могут ориентироваться по берегам, горам и даже дорогам. Над морем направление полета гораздо точнее, если берег все еще виден.[62]
Многие исследования также показали, что бабочки умеют ориентироваться. Одно исследование показало, что многие бабочки могут использовать Магнитное поле Земли ориентироваться, как исследование мотылька сердце и дротик предлагает.[63] Другое исследование миграционного поведения серебро Y, показал, что даже на больших высотах этот вид может корректировать свой курс при изменении ветра и предпочитает летать с попутным ветром, предполагающим отличное чувство направления.[64][65] Африсса статира в Панама теряет свои навигационные возможности при воздействии магнитного поля, что говорит о том, что он использует магнитное поле Земли.[66]
Бабочки проявляют тенденцию многократно кружить вокруг искусственного света. Это говорит о том, что они используют технику небесная навигация называется поперечная ориентация. Поддерживая постоянное угловое отношение к яркому небесному свету, например, Луна, они могут летать по прямой. Небесные объекты находятся так далеко, даже после путешествия на большие расстояния, изменение угла между молью и источником света незначительно; далее, Луна всегда будет в верхней части поля зрения или на горизонт. Когда моль встречает более близкий искусственный свет и использует его для навигации, угол заметно меняется уже после небольшого расстояния, в дополнение к тому, что он часто находится ниже горизонта. Мотылек инстинктивно пытается исправить это, поворачиваясь к свету, заставляя летающих бабочек падать вниз и с близкого расстояния, что приводит к спиралевидному полету, который приближается к источнику света.[67] Были предложены другие объяснения, такие как идея о том, что моль может быть нарушена визуальным искажением, называемым Полоса Маха Генри Сяо в 1972 году. Он заявил, что они летят к самой темной части неба в поисках безопасности, поэтому склонны кружить окружающие объекты в области диапазона Маха.[68]
Миграция

Миграция чешуекрылых обычно сезонный, поскольку насекомые перемещаются, спасаясь от засухи или других неблагоприятных условий. Большинство мигрирующих чешуекрылых - бабочки, и расстояние, которое они преодолевают, варьируется. Некоторые мигрирующие бабочки включают траурный плащ, нарисованная дама, Американская леди, красный адмирал, а обыкновенный конский глаз.[53]:29–30 Известный вид бабочек, который мигрирует на большие расстояния, - это богонг моль.[69] Самые известные миграции - это миграции восточного населения бабочка монарх от Мексики до севера Соединенных Штатов и юга Канады, расстояние примерно 4 000–4 800 км (2 500–3 000 миль). Среди других хорошо известных мигрирующих видов - нарисованная дама и несколько Данаин бабочки. Эффектные и масштабные миграции, связанные с муссоны видны в полуостровной Индии.[70] В последнее время миграции изучались с использованием меток крыльев и стабильных изотопов водорода.[71][72]
Мотыльки также совершают миграции, примером которых является уранииды. Урания фулгенс подвергается популяционным взрывам и массовым миграциям, которых не может превзойти никакое другое насекомое в Неотропы. В Коста-Рика и Панама первые перемещения населения могут начаться в июле и начале августа и, в зависимости от года, могут быть очень массовыми, не ослабевая в течение пяти месяцев.[73]
Общение

Феромоны обычно участвуют в брачных ритуалах между видами, особенно мотыльками, но они также являются важным аспектом других форм общения. Обычно феромоны производятся самцом или самкой и обнаруживаются представителями противоположного пола своими антеннами.[74] У многих видов железа между восьмым и девятым сегментами живота у самок вырабатывает феромоны.[14] Общение также может происходить посредством стрижки или издавать звуки при трении различных частей тела друг о друга.[65]
Мотыльки, как известно, участвуют в акустических формах общения, чаще всего в виде ухаживания, привлекая партнеров с помощью звука или вибрации. Как и большинство других насекомых, бабочки улавливают эти звуки с помощью барабанных перепонок в брюшной полости.[75] Примером может служить оса моль в горошек (Syntomeida epilais), который издает звуки с частотой, превышающей обычную для человека (около 20 кГц). Эти звуки также функционируют как тактильное общение или общение посредством прикосновения, поскольку они стрижут или вибрируют субстрат, такой как листья и стебли.[41]
Большинству бабочек не хватает ярких цветов, так как многие виды используют окраску, как камуфляж Бабочки участвуют в визуальном общении. женский капустные бабочки например, используют ультрафиолетовый свет для связи с чешуей, окрашенной в этом диапазоне на спинной поверхности крыла. Когда они летят, каждый взмах крыла вниз создает короткую вспышку ультрафиолетового света, которую самцы, по-видимому, распознают как признак полета потенциального партнера. Эти вспышки крыльев могут привлечь несколько самцов, которые демонстрируют ухаживания в воздухе.[75]
Экология
Мотыльки и бабочки важны в естественной экосистеме. Они являются неотъемлемыми участниками пищевой цепи; эволюционировав вместе с цветковыми растениями и хищниками, виды чешуекрылых сформировали сеть трофический отношения между автотрофы и гетеротрофы, которые входят в стадии личинок, куколок и взрослых чешуекрылых. Личинки и куколки - звенья в рационе птиц и паразитов. энтомофаг насекомые. Взрослые особи включены в пищевые сети гораздо более широкого круга потребителей (включая птиц, мелких млекопитающих, рептилий и т. Д.).[25]:567
Защита и хищничество

Виды чешуекрылых имеют мягкое тело, хрупкие и почти беззащитные, в то время как незрелые стадии движутся медленно или неподвижны, поэтому все стадии подвержены воздействию хищничество. Взрослые бабочки и моль охотятся на птицы, летучие мыши, ящерицы, амфибии, стрекозы, и пауки. Гусеницы и куколки становятся добычей не только птиц, но и беспозвоночных хищников и мелких млекопитающих, а также грибов и бактерий. Паразитоид и паразитический осы и мухи может откладывать яйца в гусенице, которые в конечном итоге убивают ее, поскольку они вылупляются внутри ее тела и поедают ее ткани. Птицы-насекомоядные, вероятно, являются самыми крупными хищниками. Чешуекрылые, особенно неполовозрелые, являются экологически важной пищей для многих насекомоядных птиц, таких как большая синица в Европе.
An "эволюционная гонка вооружений «можно увидеть между хищниками и жертвами. Чешуекрылые разработали ряд стратегий защиты и защиты, включая эволюцию морфологических признаков и изменения экологического образа жизни и поведения. К ним относятся апосематизм, мимикрия, камуфляж, а также разработка шаблонов угроз и их отображения.[76] Только несколько птиц, таких как козодои, охотятся на чешуекрылых. Их главные хищники - летучие мыши. Опять же, существует «эволюционная раса», которая привела к многочисленным эволюционным приспособлениям бабочек, чтобы убежать от своих основных хищников, таких как способность слышать ультразвуковые звуки или даже в некоторых случаях издавать звуки. Охотниками также являются яйца чешуекрылых. Некоторые гусеницы, такие как Зебра Махаон бабочка личинки, каннибалисты.
Некоторые виды чешуекрылых ядовиты для хищников, например, бабочка-монарх в Америке, Атрофаневра виды (розы, ветряные мельницы и др.) в Азии, а также Папилио антимахус, а птичьи крылья, самые большие бабочки в Африке и Азии соответственно. Они получают свою токсичность, улавливая химические вещества из растений, которые они едят, в свои собственные ткани. Некоторые Lepidoptera вырабатывают собственные токсины. Хищники, которые едят ядовитых бабочек и мотыльков, могут заболеть и сильно рвать, научившись не есть эти виды. Хищник, который ранее ел ядовитого чешуекрылого, может в будущем избегать других видов с аналогичными отметинами, тем самым спасая и многие другие виды.[76][77] Токсичные бабочки и личинки, как правило, приобретают яркую окраску и яркие узоры, что является индикатором их токсичности для хищников. Это явление известно как апосематизм.[78] Некоторые гусеницы, особенно представители Papilionidae, содержат осметерий, Y-образный выступ железа найдено в переднегрудной сегмент личинки.[76] При угрозе гусеница издает неприятный запах из органа, чтобы отпугнуть хищников.[79][80]
Камуфляж также является важной защитной стратегией, которая включает использование окраски или формы для слияния с окружающей средой. Некоторые виды чешуекрылых сливаются с окружающей средой, что затрудняет их обнаружение хищниками. Гусеницы могут иметь оттенки зеленого, соответствующие цвету растения-хозяина. Другие выглядят как несъедобные предметы, например, веточки или листья. Например, траурный плащ исчезает на фоне деревьев, когда складывает назад крылья. Личинки некоторых видов, например обыкновенного мормона (Papilio polytes ) и Западный тигровый ласточкин хвост похожи на птичий помет.[76][81] Например, взрослый Sesiidae виды (также известные как яснокрылые мотыльки) имеют общий вид, достаточно похожий на оса или шершень чтобы уменьшить вероятность того, что бабочки станут хищниками Бейтсовская мимикрия.[82] Пятна для глаз являются разновидностью автомимикрия используется некоторыми бабочками и мотыльками. У бабочек пятна состоят из концентрических колец чешуек разного цвета. Предполагаемая роль глазных пятен - отвлекать внимание хищников. Их сходство с глазами вызывает у хищника инстинкт атаковать эти узоры крыльев.[83]
Батезиан и Мюллериан комплексы мимикрии обычно встречаются у чешуекрылых. Генетический полиморфизм и естественный отбор приводят к появлению съедобных видов (мимик), которые получают преимущество в выживании за счет сходства с несъедобными видами (модель). Такой комплекс мимикрии называется Бейтсианом и наиболее широко известен в примере между лименитидин наместник бабочка в отношении несъедобного Данаин монарх. В действительности вице-король более токсичен, чем монарх, и это сходство следует рассматривать как случай мимикрии Мюллера.[84] В мюллеровской мимикрии несъедобные виды, обычно в рамках таксономического порядка, находят выгодным походить друг на друга, чтобы снизить частоту выборки хищниками, которым необходимо узнать о несъедобности насекомых. Таксоны из токсичного рода Геликоний образуют один из самых известных комплексов Мюллера.[85] Взрослые особи различных видов теперь настолько похожи друг на друга, что виды невозможно отличить без тщательного морфологического наблюдения и, в некоторых случаях, вскрытия или генетического анализа.
Бабочки, очевидно, могут слышать расстояние, излучаемое летучими мышами, что, по сути, заставляет летающих бабочек совершать маневры уклонения, потому что летучие мыши являются основными хищниками бабочек. Ультразвуковые частоты вызывают рефлекторное действие в совок мотылек, который заставляет его опускаться на несколько дюймов в полете, чтобы уклониться от атаки.[86] Тигровые мотыльки при защите издают щелчки в пределах того же диапазона, что и летучие мыши, которые мешают летучим мышам и препятствуют их попыткам эхолокации.[87]
Опыление

Большинство видов чешуекрылых участвуют в той или иной форме энтомофилия (точнее, психофилия и фаленофилия для бабочек и мотыльков соответственно), или опыление цветов.[88] Большинство взрослых бабочек и мотыльков питаются нектар внутри цветов, используя свои хоботки, чтобы добраться до нектара, спрятанного у основания лепестков. При этом взрослые особи касаются цветов. тычинки, на котором репродуктивная пыльца производится и хранится. Пыльца переносится на придатки взрослых особей, которые летят к следующему цветку, чтобы питаться, и невольно откладывают пыльцу на клеймо следующего цветка, где пыльца прорастает и удобряет семена.[25]:813–814
Цветы, опыляемые бабочками, обычно большие и яркие, розового или бледно-лилового цвета, часто имеют место посадки и обычно ароматные, поскольку бабочки обычно летают днем. Поскольку бабочки не переваривать пыльца (кроме виды геликонидов,[88]) предлагается больше нектара, чем пыльцы. У цветов есть простые проводники нектара, причем нектарники обычно спрятаны в узких трубочках или шпорах, к которым может добраться длинный «язык» бабочек. Такие бабочки как Thymelicus flavus были замечены участвовать в цветочное постоянство, что означает, что они с большей вероятностью передают пыльцу другим растениям того же вида. Это может быть полезно для опыляемых растений, так как постоянство цветков предотвращает потерю пыльцы во время различных полетов, а опылители не забивают рыльца пыльцой других видов цветов.[89]
К наиболее важным группам опылителей бабочек относятся: ястребиная моль из семья Sphingidae. Их поведение похоже на колибри, то есть используя быстрые взмахи крыльев, чтобы парить перед цветами. Большинство ястребиных моли ночной образ жизни или сумеречный, поэтому цветки, опыленные молью (например, Silene latifolia ), как правило, белые, открывающиеся на ночь, большие и эффектные с трубчатыми венчики и сильный сладкий запах вечером, ночью или ранним утром. Производится много нектара, чтобы подпитывать скорость метаболизма необходим для их полета.[90] Другие бабочки (например, совки, геометрические фигуры, пиралиды ) летите медленно и садитесь на цветок. Им не требуется столько нектара, как быстро летающим ястребиным мотылькам, а цветы, как правило, маленькие (хотя они могут собираться в головах).[91]
Мутуализм

Мутуализм это форма биологическое взаимодействие при этом каждый индивидуум получает выгоду в той или иной степени. Примером мутуалистических отношений могут быть отношения между мотыльки юкки (Tegeculidae) и их хозяин, цветы юкки (Спаржевые). Самки моли юкки проникают в цветки хозяина, собирают пыльцу в клубок с помощью специальных верхнечелюстных щупиков, затем перемещаются к вершине пестика, где пыльца оседает на рыльце, и откладывают яйца в основание пестика, где будут развиваться семена. Личинки развиваются в стручках плодов и питаются частью семян. Таким образом, и насекомые, и растения получают выгоду, образуя очень мутуалистические отношения.[25]:814 Другая форма мутуализма возникает между некоторыми личинками бабочек и некоторыми видами бабочек. муравьи (например. Lycaenidae ). Личинки общаются с муравьями с помощью вибраций, передаваемых через субстрат, например древесину дерева или стебли, а также с помощью химических сигналов.[92] Муравьи обеспечивают некоторую степень защиты этих личинок, и они, в свою очередь, собирают выделения медвяной росы.[93]
Паразитизм

Всего 42 вида паразитоид чешуекрылые известны (1 Pyralidae; 40 Epipyropidae ).[25]:748 Личинки больше и малая восковая моль питаться соты внутри пчела гнезда и могут стать вредители; они также находятся в шмель и оса гнезда, хотя и в меньшей степени. В Северной Европе восковая моль считается самым серьезным паразитоидом шмелей и встречается только в гнездах шмелей. В некоторых районах на юге Англии можно уничтожить до 80% гнезд.[94] Известно, что другие паразитические личинки охотятся на цикады и листовые воронки.[95]

И наоборот, мотыльки и бабочки могут быть подвержены паразитические осы и мухи, который может откладывать яйца на гусеницах, которые вылупляются и питаются внутри его тела, что приводит к гибели. Хотя в форме паразитизма, называемого идиобионтом, взрослый парализует хозяина, чтобы не убить его, а чтобы он прожил как можно дольше, чтобы паразитические личинки получили наибольшую пользу. При другой форме паразитизма, коинобионтах, виды живут за счет своих хозяев, находясь внутри (эндопаразит). Эти паразиты живут внутри гусеницы-хозяина на протяжении всего ее жизненного цикла или могут повлиять на нее позже, во взрослом возрасте. В других отрядах к коинобионтам относятся мухи, большинство жесткокрылые, и много перепончатокрылые паразитоиды.[25]:748–749 Некоторые виды могут быть подвержены различным паразитам, например, непарный мотылек (Lymantaria dispar), который подвергается нападению серией из 13 видов в шести различных таксонах на протяжении своего жизненного цикла.[25]:750
В ответ на яйцо или личинку паразитоида в теле гусеницы плазматоциты, или просто клетки-хозяева могут образовывать многослойную капсулу, которая в конечном итоге заставляет эндопаразит задыхаться. Этот процесс, называемый инкапсуляцией, является одним из единственных средств защиты гусениц от паразитоидов.[25]:748
Другие биологические взаимодействия
Некоторые виды чешуекрылых являются вторичными потребителями или хищники. Эти виды обычно охотятся на яйца других насекомых, тлей, щитовок или личинок муравьев.[25]:567 Некоторые гусеницы - каннибалы, а другие охотятся на гусениц других видов (например, гавайских Евпитеция ). Те из 15 видов в Евпитеция Эти зеркальные черви - единственный известный вид бабочек и мотыльков, которые устраивают засады.[96] Известно, что улиток поедают четыре вида. Например, гусеница гавайская (Гипосмокома моллюсков ) использует шелковые ловушки, похожие на пауков, для ловли определенных видов улиток (обычно Торнателлиды ).[95]
Личинки некоторых видов моли в Tineidae, Гелехиоиды, и Совки (семья / суперсемейство / семьи соответственно), кроме других, питаются детрит, или мертвый органический материал, такой как опавшие листья и фрукты, грибы и продукты животного происхождения, и превратить его в перегной.[25]:567 Хорошо известные виды включают тканевая моль (Тинеола Бисселлиелла, Опоясывающий лишай пеллионелла, и Trichophaga tapetzella ), которые питаются детритом, содержащим кератин, в том числе волосы, перья, паутина, птичьи гнезда (особенно домашние голуби, Columba livia domestica) и фрукты или овощи. Эти виды важны для экосистем, поскольку они удаляют вещества, разложение которых в противном случае заняло бы много времени.[97]
В 2015 году сообщалось, что оса браковирус ДНК присутствовала в чешуекрылых, таких как бабочки-монархи, тутовые шелкопряды и мотыльки.[98] Они были описаны в некоторых газетных статьях как примеры естественного генно-инженерный насекомые.[99]
Эволюция и систематика
История учебы

Линней в Systema Naturae (1758) признал три подразделения Lepidoptera: Папилио, Сфинкс и Phalaena, с семью подгруппами в Phalaena.[100] Они сохраняются сегодня как 9 суперсемейств Lepidoptera. Другие работы по классификации, в том числе Майкл Денис & Игнац Шиффермюллер (1775), Йохан Кристиан Фабрициус (1775) и Пьер Андре Латрей (1796). Якоб Хюбнер описали многие роды, а роды чешуекрылых были каталогизированы Фердинанд Оксенхаймер и Георг Фридрих Трейчке в серии томов о фауне чешуекрылых Европы, опубликованных между 1807 и 1835 годами.[100] Готлиб Август Вильгельм Херрих-Шеффер (несколько томов, 1843–1856 гг.) и Эдвард Мейрик (1895) основывали свою классификацию прежде всего на жилковании крыльев. Сэр Джордж Фрэнсис Хэмпсон работали над микроклепидоптерами в этот период и Филипп Кристоф Целлер опубликовано Естественная история Tineinae также на микролепидоптерах (1855).
Одним из первых энтомологов, изучавших ископаемых насекомых и их эволюцию, был Сэмюэл Хаббард Скаддер (1837–1911), работавший над бабочками.[101] Он опубликовал исследование Флориссантских отложений Колорадо, включая исключительно сохранившиеся Продряс Персефона. Андреас В. Мартынов (1879–1938) признал тесную связь между чешуекрылыми и Trichoptera в своих исследованиях филогении.[101]
Главный вклад в 20-й век включал создание Монотризии и дитризии (основанной на структуре женских гениталий) Борнером в 1925 и 1939 годах.[100] Вилли Хенниг (1913–1976) разработал кладистический методологии и применили ее к филогении насекомых. Нильс П. Кристенсен, Э. С. Нильсен и Д. Р. Дэвис изучали отношения между монотризский семьи и Кристенсен в целом работали над насекомыми филогения а также чешуекрылые высшие.[100][101] Хотя часто обнаруживается, что филогения, основанная на ДНК, отличается от филогении, основанной на морфология это не относится к Lepidoptera; Филогенез ДНК в значительной степени соответствует филогенезу, основанному на морфологии.[101]
Было сделано много попыток сгруппировать надсемейства чешуекрылых в естественные группы, большинство из которых терпят неудачу, потому что одна из двух групп не является монофилетический: Microlepidoptera и Macrolepidoptera, Heterocera и Rhopalocera, Jugatae и Frenatae, Monotrysia и Ditrysia.[100]
Окаменелости

Летопись окаменелостей чешуекрылых отсутствует по сравнению с другими крылатыми видами и, как правило, не так распространена, как некоторые другие насекомые, в местах обитания, наиболее способствующих окаменению, таких как озера и пруды; их ювенильная стадия имеет только головную капсулу как твердую часть, которую можно сохранить. Расположение и численность наиболее распространенных видов бабочек указывает на то, что массовые миграции бабочек происходили в палеогене. Северное море, поэтому окаменелостей моли очень мало.[102] Тем не менее, есть окаменелости, некоторые из которых сохранились в янтаре, а некоторые - в очень мелких отложениях. Листовые мины также встречаются в ископаемых листьях, хотя их интерпретация непроста.[101]
Предполагаемые ископаемые представители стволовой группы Амфиесменоптеры (клада, включающая Trichoptera и Lepidoptera) известна из Триасовый.[25]:567 Самые ранние известные окаменелости чешуекрылых представляют собой окаменелые чешуйки из Граница триаса и юры. Они были найдены как редкие палинологический элементов в отложениях границы триаса и юры из керновой скважины Шанделах-1, пробуренной вблизи Брауншвейг на севере Германия. Это отодвигает назад летопись окаменелостей и происхождение глоссатан чешуекрылых примерно на 70 миллионов лет, подтверждая молекулярные оценки Norian (около 212 миллионов лет) расхождение глоссатановых и неглоссатских чешуекрылых. Результаты были опубликованы в 2018 году в журнале. Достижения науки. Авторы исследования предположили, что чешуекрылые развили хоботок как приспособление к питью из капель и тонких пленок воды для поддержания своего состояния. баланс жидкости в жарком и засушливом климат из Триасовый.[103]
Самый ранний из названных таксонов чешуекрылых - Грива археолеписа, примитивный вид бабочек из Раннеюрский период, датированный примерно 190 миллион лет назад, и известен только по трем крыльям, найденным в Чармут Мадстоун из Дорсет, Великобритания. На крыльях видны чешуйки с параллельными бороздками под растровым электронным микроскопом и характерный рисунок жилкования крыла, общий с Трихоптера (ручейники).[104][105] Были обнаружены еще только два набора окаменелостей юрских чешуекрылых, а также 13 наборов из Меловой, которые все принадлежат к примитивным семействам мотыльков.[101]
Еще много окаменелостей найдено из третичного периода, и особенно эоцен Балтийский янтарь. Самые старые настоящие бабочки надсемейства Papilionoidea были найдены в Палеоцен MoClay или Формирование меха из Дания. Лучше всего сохранившиеся ископаемые чешуекрылые относятся к эоцену. Продряс Персефона от Флориссант ископаемые кровати.
Филогения
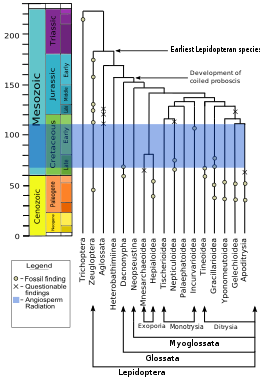
Чешуекрылые и Трихоптера (ручейники) сестринские группы, разделяя много общего, чего не хватает у других; например, женщины обоих отрядов гетерогаметный, то есть у них есть два разных половые хромосомы, в то время как у большинства видов самцы гетерогаметны, а самки имеют две идентичные половые хромосомы. Взрослые особи обоих отрядов демонстрируют особый рисунок жилкования передних крыльев. Личинки в двух отрядах имеют структуры рта и железы, с помощью которых они производят и управляют шелк. Вилли Хенниг сгруппировали два заказа в суперзаказ Амфиесменоптеры; вместе они сестры вымершего ордена Тарахоптеры.[106] Чешуекрылые произошли от общего предка, похожего на дневную моль, который питался либо мертвыми, либо живыми растениями.[107]
В кладограмма, на основе 2008 г. ДНК и белок анализ, показывает порядок как клады, сестра Trichoptera, и более дальняя родственница Двукрылые (настоящие мухи) и Mecoptera (скорпионы).[108][109][110][111]
| частьЭндоптеригот |
| ||||||||||||||||||||||||||||||||||||

Micropterigidae, Agathiphagidae и Heterobathmiidae самые старые и самые базальный линии Lepidoptera. У взрослых этих семей нет загнутого языка или хоботок, которые встречаются у большинства членов отряда, но вместо этого челюсти адаптирован для специальной диеты. Личинки Micropterigidae питаются уходит, грибы, или печеночники (очень похоже на Трихоптера ).[100] Взрослые особи Micropterigidae жуют пыльцу или споры папоротников. У Agathiphagidae личинки живут внутри каури сосны и питаются семенами. У Heterobathmiidae личинки питаются листьями Нотофагус, бук южный. У этих семей также есть нижние челюсти на стадии куколки, которые помогают куколке выйти из семени или кокона после метаморфоза.[100]
В Eriocraniidae имеют короткий спиральный хоботок на взрослой стадии, и хотя они сохраняют свои куколочные мандибулы, с помощью которых они вышли из кокона, их нижние челюсти после этого нефункциональны.[100] Большинство из этих недитризиевых семейств в первую очередь листовые шахтеры в личиночной стадии. Помимо хоботка, среди этих базальных ветвей наблюдается изменение чешуи, при этом более поздние линии показывают более сложные перфорированные чешуйки.[101]
С развитием Дитрисия в середине мелового периода произошли серьезные репродуктивные изменения. Ditrysia, которые составляют 98% чешуекрылых, имеют два отдельных отверстия для размножения у самок (а также третье отверстие для выделения), одно для спаривания и одно для откладки яиц. Эти двое изнутри связаны семенным протоком. (В других базальных линиях есть одна клоака, или позже, два отверстия и внешний семенной канал.) Из ранних линий Ditrysia, Gracillarioidea и Гелехиоидея в основном это листовые комбайны, но более поздние линии питаются извне. в Tineoidea, большинство видов питаются растительным и животным детритом и грибами и строят укрытия на личиночной стадии.[101]
В Yponomeutoidea это первая группа, в которой имеется значительное количество видов, личинки которых питаются травянистыми растениями, а не древесными.[101] Они возникли примерно в то время, когда цветущие растения подверглись обширному адаптивное излучение в середине-Меловой, и Gelechioidea, которые развились в это время, также имеют большое разнообразие. Были ли вовлечены процессы коэволюция или последовательная эволюция, разнообразие чешуекрылых и покрытосеменных увеличивалось вместе.
В так называемом "макролепидоптеры ", который составляет около 60% видов чешуекрылых, наблюдалось общее увеличение в размерах, лучшая летная способность (за счет изменения формы крыльев и сцепления передних и задних крыльев), уменьшение челюстей взрослых особей и изменение расположения крючки на ножках личинки, возможно, для улучшения сцепления с растением-хозяином.[101] Многие также имеют барабанные органы, которые позволяют им слышать. Эти органы эволюционировали по крайней мере восемь раз, потому что они расположены на разных частях тела и имеют структурные различия.[101]Основными линиями макролепидоптера являются Noctuoidea, Bombycoidea, Lasiocampidae, Mimallonoidea, Геометроидеа и Ропалоцера. Bombycoidea плюс Lasiocampidae плюс Mimallonoidea могут быть монофилетический группа.[101] Rhopalocera, включая Papilionoidea (бабочки), Геспериоидея (шкиперы), а Hedyloidea (бабочки-мотыльки), эволюционировали совсем недавно.[100] Об этой группе сохранилась довольно хорошая летопись окаменелостей, самый старый шкипер датируется 56 миллион лет назад.[101]
Таксономия
Таксономия - это классификация видов в выбранных таксонах, при этом процесс наименования называется номенклатура. У чешуекрылых насчитывается более 120 семейств, от 45 до 48 надсемейств. Исторически чешуекрылые всегда подразделялись на пять подотрядов, один из которых - примитивные бабочки, которые никогда не теряли морфологических особенностей своих предков. Остальные мотыльки и бабочки составляют девяносто восемь процентов других таксонов, делая Дитрисия. Совсем недавно находки новых таксонов, личинок и куколок помогли детализировать взаимоотношения примитивных таксонов, филогенетический анализ показал, что примитивные линии парафилетический по сравнению с остальными линиями Lepidoptera. В последнее время лепидоптерологи отказались от кладов, таких как подклассы, а также клады между отрядами и надсемействами.[25]:569
- Zeugloptera клад с Micropterigoidea являясь его единственным надсемейством, содержащим единственное семейство Micropterigidae. Виды Micropterigoidea - это практически живые ископаемые, являющиеся одной из самых примитивных групп чешуекрылых, все еще сохраняющие жевательные части рта (челюсти) у взрослых особей, в отличие от других клад бабочек и моли. Во всем мире известно около 120 видов, более половины из которых входят в этот род. Микроптерикс в Палеарктике. В Северной Америке известны всего два (Эпимартирия ), причем гораздо больше встречается в Азии и юго-западной части Тихого океана, особенно в Новой Зеландии, где обитает около 50 видов.[25]:569
- Aglossata второй по примитивности родословная Lepidoptera; впервые описан в 1952 г. Лайонел Джек Дамблтон. Agathiphagidae является единственным семейством Aglossata и включает два вида в своем единственном роде, Агатифага.[25]:569[112] Agathiphaga queenslandensis и Agathiphaga vitiensis находятся вдоль северо-восточного побережья Квинсленд, Австралия, И в Фиджи к Вануату и Соломоновы острова соответственно.[113][114]
- Heterobathmiina был впервые описан Кристенсеном и Нильсеном в 1979 году. Heterobathmiidae является единственным семейством и включает около 10 видов дневных металлических бабочек, обитающих в южной части Южной Америки, взрослые особи поедают пыльцу Нотофагус или южный бук, и личинки добывают листья.[25]:569[115]
- Глоссата содержит большинство видов, с наиболее очевидной разницей в том, что нижняя челюсть не функционирует, а также удлиненные верхнечелюстные галеи или хоботок. В базальные клады все еще сохраняющие некоторые наследственные черты крыльев, такие как передние и задние крылья аналогичной формы с относительно полным жилкованием. Глоссата также содержит раздел Дитрисия, который содержит 98% всех описанных видов Lepidoptera.[25]:569
Отношение к людям
Культура

Художественные изображения бабочек использовались во многих культурах, в том числе еще 3500 лет назад в египетских иероглифах.[116] Сегодня бабочки широко используются в различных предметах искусства и ювелирных изделиях: в рамах, залитых смолой, в бутылках, ламинированных бумагой, а также в некоторых произведениях искусства и мебели в смешанной технике.[117] Бабочки также вдохновили "бабочка фея «как художественный и вымышленный персонаж.
Во многих культурах душа умершего ассоциируется с бабочкой, например, в Древняя Греция, где слово для бабочки ψυχή (психика) также означает душа и дыхание. В латинский, как и в Древней Греции, слово "бабочка" папилио ассоциировался с душой мертвых.[118] Знак в виде черепа на грудной клетке мертвая голова ястреба помог этим мотылькам, особенно A. atropos, заработайте отрицательную репутацию, например, ассоциации со сверхъестественным и злом. Мотылек широко используется в искусстве и фильмах, таких как Un Chien Andalou (от Buñuel и Дали ) и Молчание ягнят, и в художественном оформлении японской металлической группы Вздох альбом Приветствую Ужас Град. Согласно с Квайдан: Истории и исследования странных вещей, от Лафкадио Хирн, бабочка была замечена в Японии как персонификация души человека; живы ли они, умирают или уже мертвы. Одно японское суеверие гласит, что если бабочка входит в вашу комнату и садится за бамбук экран, к вам придет человек, которого вы больше всего любите. Однако большое количество бабочек считается плохим приметы. Когда Тайра-но Масакадо тайно готовился к своему знаменитому восстанию, в Киото такой огромный рой бабочек, что люди были напуганы, думая, что привидение является предзнаменованием грядущего зла.[119]
В древнем Мезоамериканец город Теотиуакан, ярко окрашенное изображение бабочки было вырезано во многих храмах, зданиях, украшениях и украшено курильницы особенно. Иногда бабочку изображали с пастью ягуар а некоторые виды считались реинкарнациями душ мертвых воинов. Тесная связь бабочек с Огонь и война продолжалась до Цивилизация ацтеков и доказательства подобных изображений ягуара-бабочки были найдены среди Сапотек, и Цивилизации майя.[120]

Вредители
В личинки многих видов чешуекрылых являются основными вредителями сельское хозяйство. Некоторые из основных вредителей включают: Tortricidae, Совки, и Pyralidae. Личинки рода Noctuidae Spodoptera (армейские черви), Helicoverpa (кукурузный червь), или Pieris brassicae может нанести значительный ущерб определенным культурам.[100] Helicoverpa zea личинки (хлопковые совки или плодовые черви томатов) многоядный, то есть они едят разные культуры, в том числе помидоры и хлопок.[121] Peridroma saucia (пестрые совки) описываются как один из самых вредных вредителей для садов, способный уничтожить целые сады и поля за считанные дни.[122]
Бабочки и моль являются одними из крупнейших таксонов, которые питаются исключительно живыми растениями и зависят от них, с точки зрения количества видов, и они входят во многие экосистемы, составляя для этого самую большую биомассу. У многих видов самка может производить от 200 до 600 яиц, а у некоторых других - до 30 000 яиц за один день. Это может создать множество проблем для сельского хозяйства, где многие гусеницы могут поражать акры растительности. По некоторым оценкам, на одном дубе питалось более 80 000 гусениц нескольких различных таксонов. В некоторых случаях личинки фитофагов могут привести к гибели целых деревьев за относительно короткие промежутки времени.[25]:567
Экологические способы уничтожения вредителей Виды чешуекрылых становятся более экономически выгодными, поскольку исследования показали, что такие способы, как интродукция паразитических ос и мух. Например, Саркофага альдрихи, муха, отложившая личинки, питается куколками лесная палатка гусеница моль. Пестициды могут влиять на другие виды, кроме тех, на уничтожение которых они нацелены, нанося ущерб естественной экосистеме.[123] Еще один хороший метод биологической борьбы с вредителями - это использование феромоновые ловушки. Феромоновая ловушка - это разновидность ловушка для насекомых который использует феромоны чтобы заманить насекомые. Чаще всего используются половые феромоны и агрегирующие феромоны. Приманка, пропитанная феромоном, помещается в обычную ловушку, такую как ловушка Delta, ловушка с водяным поддоном или воронкообразная ловушка.[124]
Виды бабочек, которые детритофаги естественно есть детрит содержащий кератин, такие как волосы или перья. Хорошо известные виды суконная моль (T. bisselliella, Т. pellionella, и T. Tapetzella ), питаясь продуктами питания, которые люди считают экономически важными, такими как хлопок, белье, шелк и шерсть ткани а также меха; кроме того, они были найдены в сарае перья и волосы, отруби, манная крупа и порошок (возможно, предпочитая пшеница порошок), печенье, казеин, и насекомое образцы в музеи.[97]
Полезные насекомые
Хотя большинство бабочек и мотыльков отрицательно влияют на экономику, некоторые виды являются ценным экономическим ресурсом. Самый яркий пример - это одомашненный шелкопряд (Bombyx mori), личинки которых делают коконы из шелк, который можно прядить в ткань. Шелк был и был важным экономическим ресурсом на протяжении всей истории. Виды Bombyx mori был одомашнен до такой степени, что его выживание полностью зависит от человечества.[125] Ряд диких бабочек, таких как Бомбикс мандарина, и Антерея виды, помимо других, обеспечивают коммерчески важные шелка.[126]
Предпочтение личинок большинства видов чешуекрылых питаться одним видом или ограниченным кругом растений используется в качестве механизма биологического контроля над ними. сорняки вместо гербицидов. В пиралид кактусовая моль был завезен из Аргентины в Австралию, где успешно подавил миллионы акров кактус опунция.[25]:567 Другой вид Pyralidae, названный бурильный молоток из аллигатора (Аркола маллой), использовался для управления водное растение известный как аллигатор (Альтернантера филоксероидес) в сочетании с аллигатор сорняк, блошка; в этом случае два насекомых работают в синергия и сорняк редко восстанавливается.[127]
Разведение бабочек и моли, или бабочка садоводство / выращивание, стало экологически жизнеспособным процессом интродукции видов в экосистему, чтобы принести ей пользу. Разведение бабочек в Папуа-Новой Гвинее позволяет гражданам этой страны «выращивать» экономически ценные виды насекомых для рынка коллекционеров экологически устойчивым образом.[128]
Еда

Lepidoptera занимают видное место в энтомофагия как продукты питания почти на всех континентах. Хотя в большинстве случаев коренные жители едят взрослых особей, личинок или куколок в качестве основных продуктов питания, Beondegi или тутовый шелкопряд куколки едят как закуску в Корейская кухня[129] в то время как Червь Магуи считается деликатесом в Мексика.[130] В некоторых частях Уастека, шелковые гнезда Мадронская бабочка поддерживаются на краю крыш домов для потребления.[131] в Карния регион Италии, дети ловят и едят слюнки токсичных Zygaena бабочки в начале лета. Инглувис, несмотря на очень низкое содержание цианогена, служит удобным дополнительным источником сахара для детей, которые могут включать этот ресурс в сезонный деликатес с минимальным риском.[132]
Здоровье
Некоторые личинки как моли, так и бабочек имеют форму волос, которая, как известно, является причиной проблем со здоровьем человека. Волосы гусеницы иногда содержат токсины, и виды из примерно 12 семейств бабочек или бабочек во всем мире могут нанести серьезные травмы человеку (крапивница дерматит и атопический астма к остеохондрит, потребление коагулопатия, почечный неудача, и внутримозговый кровоизлияние ).[133] Кожные высыпания являются наиболее распространенным явлением, но бывают случаи со смертельным исходом.[134] Лономия является частой причиной отравлений у людей в Бразилии: в период с 1989 по 2005 гг. было зарегистрировано 354 случая. Летальность достигает 20%, причем смерть чаще всего вызвана внутричерепным кровоизлиянием.[135]
Эти волосы также вызывают кератоконъюнктивит. Острые зазубрины на концах волосков гусеницы могут застревать в мягких тканях и слизистые оболочки такие как глаза. Как только они попадают в такие ткани, их трудно извлечь, что часто усугубляет проблему, поскольку они мигрируют через мембрану.[136] Это становится особой проблемой в помещении. Волосы легко проникают в здания через системы вентиляции и накапливаются в помещениях из-за своего небольшого размера, что затрудняет их удаление. Это накопление увеличивает риск контакта с людьми в помещениях.[137]
Смотрите также
- Сравнение бабочек и мотыльков
- Lepidoptera в 10-м издании Systema Naturae
- Центр McGuire по чешуекрылым и биоразнообразию, Университет Флориды
- Societas Europaea Lepidopterologica
Списки
использованная литература
- ^ а б c d Капинера, Джон Л. (2008). «Бабочки и мотыльки». Энциклопедия энтомологии. 4 (2-е изд.). Springer. С. 626–672. ISBN 9781402062421.
- ^ а б c Маллет, Джим (12 июня 2007 г.). «Таксономия чешуекрылых: масштаб проблемы». Проект таксома Lepidoptera. Университетский колледж, Лондон. В архиве из оригинала 5 июня 2011 г.. Получено 8 февраля 2011.
- ^ «Проект таксома чешуекрылых». Проект таксома Lepidoptera. В архиве из оригинала 18 августа 2011 г.. Получено 25 февраля 2015.
- ^ а б c d Пауэлл, Джерри А. (2009). «Чешуекрылые». In Resh, Vincent H .; Cardé, Ring T. (ред.). Энциклопедия насекомых (2 (иллюстрировано) изд.). Академическая пресса. С. 557–587. ISBN 978-0-12-374144-8.
- ^ а б Kristensen, Niels P .; Скобл, М. Дж .; Карсхольт, Оле (2007). «Филогения и систематика чешуекрылых: состояние инвентаризации разнообразия моли и бабочек» (PDF). В Z.-Q. Чжан; W. A. Shear (ред.). Трехсотлетие Линнея: прогресс в таксономии беспозвоночных (Zootaxa: 1668). Magnolia Press. С. 699–747. ISBN 978-0-12-690647-9. Получено 2 марта 2010.
- ^ Линней, Карл (1746). Fauna Svecica: sistens animalia Sveciæ regni: квадрупедия, авес, амфибия, рыбы, насекомые, вермесы, дистрибьюторы по классам и обычным, родам и видам. Cum diffis specierum, synonymis autorum, nominibus incolarum, locis Hubationum, descriptionibus инсекториум (на латыни). Лугдуни Батаворум [Лейден]: C. Wishoff et G.J. Wishoff. п. 232.
- ^ Энгель, Майкл С .; Кристенсен, Нильс П. (2013). «История энтомологической классификации». Ежегодный обзор энтомологии. 58 (1): 585–607. Дои:10.1146 / annurev-ento-120811-153536. ISSN 0066-4170. PMID 23317047.
- ^ Харпер, Дуглас. «Чешуекрылые». Интернет-словарь этимологии. В архиве из оригинала 10 июля 2011 г.. Получено 8 февраля 2011.
- ^ а б Партридж, Эрик (2009). Origins: этимологический словарь современного английского языка. Рутледж. ISBN 978-0-203-42114-7.
- ^ Харп, Дуглас; Дэн МакКормак (ноябрь 2001 г.). «Интернет-этимологический словарь». LogoBee. п. 1. В архиве из оригинала 3 сентября 2012 г.. Получено 6 декабря 2009.
- ^ Арнетт, Росс Х. (28 июля 2000 г.). «Часть I: 27». Американские насекомые: справочник насекомых Америки к северу от Мексики (2-е изд.). CRC Press. п. 631. ISBN 978-0-8493-0212-1.
- ^ Харпер, Дуглас. "моль". Интернет-словарь этимологии. В архиве из оригинала от 6 июня 2011 г.. Получено 31 марта 2011.
- ^ "Гусеница". Dictionary.com. В архиве из оригинала от 9 сентября 2011 г.. Получено 5 октября 2011.
- ^ а б c d е ж г час я j k Gullan, P.J .; П. С. Крэнстон (13 сентября 2004 г.). "7". Насекомые: очерк энтомологии (3-е изд.). Вили-Блэквелл. стр.198–199. ISBN 978-1-4051-1113-3.
- ^ Штумпе, Феликс. "Parnassius arctica Eisner, 1968". Russian-Insects.com. Архивировано из оригинал 15 июля 2011 г.. Получено 9 ноября 2010.
- ^ Мани, М. С. (1968). Экология и биогеография высотных насекомых. Том 4 серии entomologica. Springer. п. 530. ISBN 978-90-6193-114-0. Получено 9 ноября 2010.
- ^ Шерман, Ли (2008). "Ученый OSU бросает вызов неизведанному тропическому лесу в поисках редких и исчезающих видов" (PDF). Terra. Государственный университет Орегона. 3 (2). Архивировано из оригинал (PDF) 19 сентября 2011 г.. Получено 14 февраля 2011.
- ^ а б Рау, П. (1941).«Наблюдения за некоторыми чешуекрылыми и перепончатокрылыми паразитами ос Polistes». Анналы энтомологического общества Америки. 34 (2): 355–366(12). Дои:10.1093 / aesa / 34.2.355.
- ^ Маллет, Джим (12 июня 2007 г.). «Таксономия бабочек: масштаб проблемы». Проект таксома Lepidoptera. Университетский колледж, Лондон. В архиве из оригинала 14 мая 2011 г.. Получено 8 февраля 2011.
- ^ Eaton, Eric R .; Кауфман, Кенн (2007). Полевой справочник Кауфмана по насекомым Северной Америки. Houghton Mifflin Harcourt. п. 391. ISBN 978-0-618-15310-7. В архиве из оригинала на 24 июля 2019 г.. Получено 22 сентября 2016.
- ^ Таскес, Пол М .; Таттл, Джеймс П .; Коллинз, Майкл М. (1996). Дикие шелковые мотыльки Северной Америки: естественная история Saturniidae в Соединенных Штатах и Канаде. Серия Корнелла по биологии членистоногих (иллюстрированная ред.). Издательство Корнельского университета. п. 250. ISBN 978-0-8014-3130-2. Получено 12 февраля 2011.
- ^ Грин, Кен; Осборн, Уильям С. (1994). Дикая природа снежной страны Австралии: подробный путеводитель по альпийской фауне (иллюстрированный ред.). Рид. п. 200. ISBN 978-0-7301-0461-2.
- ^ а б c d Гилло, К. (1995). «Бабочки и мотыльки». Энтомология (2-е изд.). С. 246–266. ISBN 978-0-306-44967-3. Получено 14 ноября 2010.
- ^ а б c d Скобл (1995). Раздел Голова взрослого - питание и ощущение, (стр. 4–22).
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах Реш, Винсент Х .; Ринг Т. Карде (1 июля 2009 г.). Энциклопедия насекомых (2-е изд.). США: Academic Press. ISBN 978-0-12-374144-8.
- ^ Кристофер, О'Тул (2002). Энциклопедия насекомых и пауков Firefly (1-е изд.). ISBN 978-1-55297-612-8.
- ^ Хеппнер, Дж. Б. (2008). «Бабочки и мотыльки». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Виртуальная справочная библиотека Gale. 4 (2-е изд.). Ссылка Springer. п. 4345. ISBN 978-1-4020-6242-1. В архиве из оригинала 24 июня 2016 г.. Получено 22 сентября 2016.
- ^ Скобл, MJ. (1992). Чешуекрылые: форма, функции и разнообразие. Oxford Univ. Нажмите. ISBN 978-1-4020-6242-1.
- ^ а б Скобл (1995). Раздел Напольные весы, (стр. 63–66).
- ^ Вукусич, П. (2006). «Структурная окраска чешуекрылых» (PDF). Текущая биология. 16 (16): R621–3. Дои:10.1016 / j.cub.2006.07.040. PMID 16920604. S2CID 52828850. Архивировано из оригинал (PDF) 26 октября 2010 г.. Получено 11 ноября 2010.
- ^ Холл, Джейсон П. У .; Харви, Дональд Дж. (2002). «Обследование андрокониальных органов у Riodinidae (Lepidoptera)» (PDF). Зоологический журнал Линнеевского общества. 136 (2): 171–197. Дои:10.1046 / j.1096-3642.2002.00003.x. Архивировано из оригинал (PDF) 5 марта 2012 г.. Получено 4 июн 2012.
- ^ Уильямс, К. М. (1947). «Физиология диапаузы насекомых. II. Взаимодействие между мозгом куколки и переднегрудными железами в процессе метаморфоза гигантского тутового шелкопряда. Platysamia cecropia". Биологический бюллетень. 93 (2): 89–98. Дои:10.2307/1538279. JSTOR 1538279. PMID 20268135.
- ^ а б c Gullan, P.J .; П. С. Крэнстон (22 марта 2010 г.). Насекомые: очерк энтомологии (4-е изд.). Оксфорд: Wiley, John & Sons, Incorporated. ISBN 978-1-4443-3036-6.
- ^ Lighton J.R.B .; Лавгроув Б. Г. (1990). «Переключение с диффузной на конвективную вентиляцию у пчел, вызванное температурой» (PDF). Журнал экспериментальной биологии. 154 (1): 509–516.
- ^ Форд, Э. Б. (1965). «Генетический полиморфизм». Труды Лондонского королевского общества. Серия B, Биологические науки. Oxford University Press. 164 (995): 350–61. Дои:10.1098 / rspb.1966.0037. PMID 4379524. S2CID 202575235.
- ^ а б c Гуллан и Крэнстон (2005). «Полиморфизм и полифенизм». Насекомые: очерк энтомологии. стр.163–164.
- ^ Нур, Мохамед А. Ф .; Парнелл, Робин С .; Грант, Брюс С. (2008). «Обратимый цветной полифенизм у американской берёзовой мотылька (Бистон бетулярия когнатариа) гусеницы ». PLoS ONE. 3 (9): e3142. Bibcode:2008PLoSO ... 3.3142N. Дои:10.1371 / journal.pone.0003142. ЧВК 2518955. PMID 18769543.
- ^ а б Кунте, Крушнамег (2000). Бабочки полуостровной Индии. Часть жизненного цикла проекта. Ориент Блэксвэн. ISBN 81-7371-354-5, ISBN 978-81-7371-354-5.
- ^ Айви И.Г., Моргун Д.В., Довгайло К.Э., Рубин Н.И., Солодовников И.А. Дневные бабочки (Hesperioidea и Papilionoidea, Lepidoptera) Восточной Европы. «Определитель компакт-дисков, база данных и программный комплекс« Лисандра ». Минск, Киев, Москва: 2005. На русском языке
- ^ "Psychidae at Bug Guide". Государственный университет Айовы. В архиве из оригинала 10 февраля 2012 г.. Получено 19 января 2010.
- ^ а б Sanderford, M. V .; У. Э. Коннер (июль 1990 г.). "Ухаживающие звуки осиной мотылька в горошек, Syntomeida epilais". Naturwissenschaften. 77 (7): 345–347. Bibcode:1990NW ..... 77..345S. Дои:10.1007 / BF01138395. S2CID 1359112.
- ^ Виклунд, Кристер (июль 1984 г.). «Образцы откладки яиц у бабочек в зависимости от их фенологии, а также внешнего вида и обилия их растений-хозяев». Oecologia. 63 (1): 23–29. Bibcode:1984 Oecol..63 ... 23 Вт. Дои:10.1007 / BF00379780. PMID 28311161. S2CID 29210301.
- ^ Оберхаузер, Карен С.; Соленский, Мишель Дж (2004). Бабочка-монарх: биология и сохранение (Первое изд.). Итака, штат Нью-Йорк: Издательство Корнельского университета. ISBN 978-0801441882.
- ^ Оберхаузер 2009, п. 3.
- ^ Gullan, P.J .; Крэнстон, П. С. (2010). «Образцы и фазы жизненного цикла». Насекомые: очерк энтомологии (4-е изд.). Wiley-Blackwell. С. 156–164. ISBN 978-1-4443-3036-6.
- ^ а б Дагдейл, Дж. С. (1996). "Естественная история и идентификация питающихся подстилкой личинок чешуекрылых (Insecta) в буковых лесах, долина Оронго-Ронго, Новая Зеландия, с особым упором на рацион мышей (Mus musculus)" (PDF). Журнал Королевского общества Новой Зеландии. 26 (4): 251–274. Дои:10.1080/03014223.1996.9517513. Получено 14 ноября 2010.[постоянная мертвая ссылка ]
- ^ Триплхорн, Чарльз А .; Джонсон, Норман Ф. (2005). Введение Боррора и Делонга в изучение насекомых. Белмонт, Калифорния: Томсон Брукс / Коул. ISBN 978-0-03-096835-8.
- ^ Elmes, G.W .; Wardlaw, J.C .; Schönrogge, K .; Thomas, J.A .; Кларк, Р. (2004). «Пищевой стресс вызывает различную выживаемость социально паразитических гусениц Maculinea rebeli, интегрированных в колонии видов муравьев Myrmica, являющихся хозяевами и не являющихся хозяевами». Entomologia Experimentalis et Applicata. 110 (1): 53–63. Дои:10.1111 / j.0013-8703.2004.00121.x. S2CID 86427983.
- ^ Арнетт, Росс Х. мл. (28 июля 2000 г.). Американские насекомые. Справочник насекомых Америки к северу от Мексики (2-е изд.). ООО "ЦРК пресс". С. 631–632. ISBN 978-0-8493-0212-1.
- ^ Берг, Клиффорд О. (1950). «Биология некоторых водных гусениц (Pyralididae: Нимфула spp.), которые питаются Потамогетон". Труды Американского микроскопического общества. 69 (3): 254–266. Дои:10.2307/3223096. JSTOR 3223096.
- ^ Эрлих, П. Р .; Рэйвен, П. Х. (1964). «Бабочки и растения: исследование коэволюции». Эволюция. 18 (4): 586–608. Дои:10.2307/2406212. JSTOR 2406212.
- ^ а б Ниджут, Х. Фредерик (17 августа 1991 г.). Развитие и эволюция рисунка крыльев бабочек (Смитсоновская серия в сравнительной эволюционной биологии) (1-е изд.). Scholarly Press Смитсоновского института. С. 2–4. ISBN 978-0-87474-917-5.
- ^ а б Доул, Клэр Хаген (28 мая 2003 г.). Руководство садовника бабочек. Бруклинский ботанический сад. п.26. ISBN 978-1-889538-58-7.
метаморфоза бабочки.
- ^ Уорд, Джеймс V .; Уорд, Питер Э. (1992). Экология, биология и среда обитания водных насекомых. Джон Вили и сыновья. ISBN 978-0-471-55007-5.
- ^ Бенджамин Янцен; Томас Эйснер (28 июля 2008 г.). «Задние крылья не нужны для полета, но необходимы для выполнения обычного уклончивого полета Lepidoptera». Труды Национальной академии наук. 105 (43): 16636–16640. Bibcode:2008PNAS..10516636J. Дои:10.1073 / pnas.0807223105. ЧВК 2575472. PMID 18936482.
- ^ «Шкиперы бабочки и мотыльки: чешуекрылые - поведение и размножение». Net Industries и ее лицензиары. 2011. Архивировано с оригинал 26 июля 2011 г.. Получено 20 февраля 2011.
- ^ Рейснер, Алекс. «Скорость животных». speedofanimals.com. Архивировано из оригинал 25 февраля 2011 г.. Получено 20 февраля 2011.
- ^ Скобл, Малкольм (1995). Чешуекрылые: форма, функции и разнообразие. Oxford University Press, 1995. С. 66–67. ISBN 978-0-19-854952-9.
- ^ Сауман, Иво; Адриана Д. Бриско; Хайсун Чжу; Диндин Ши; Орен Фрой; Юлия Сталлейкен; Цюань Юань; Эми Кассельман; Стивен М. Репперт (5 мая 2005 г.). «Подключение навигационных часов к входу солнечного компаса в мозге бабочки-монарха». Нейрон. 46 (3): 457–467. Дои:10.1016 / j.neuron.2005.03.014. PMID 15882645. S2CID 17755509.
- ^ Саутвуд, Т. Р. Э. (1962). «Миграция наземных членистоногих по отношению к среде обитания». Биологические обзоры. 37 (2): 171–211. Дои:10.1111 / j.1469-185X.1962.tb01609.x. S2CID 84711127.
- ^ Деннис, Роджер Л. Х .; Тим Г. Шрив; Генри Р. Арнольд; Дэвид Б. Рой (сентябрь 2005 г.). «Контролирует ли широта диеты размер распространения травоядных насекомых? История жизни и источники ресурсов для бабочек-специалистов». Журнал сохранения насекомых. 9 (3): 187–200. Дои:10.1007 / s10841-005-5660-х. S2CID 20605146.
- ^ Made, J. G. van der; Йозеф Блаб; Руди Хольцбергер; Х. ван ден Бийтель (1989). Actie voor Vlinders, zo kunnen we ze redden (на голландском). Верт: M&P Cop. п. 192. ISBN 978-90-6590-303-7.
- ^ Бейкер, Р. Робин (февраль 1987 г.). "Комплексное использование луны и магнитных компасов мотыльком" сердце и дротик ", Восклицательный агротис". Поведение животных. 35 (1): 94–101. Дои:10.1016 / S0003-3472 (87) 80214-2. S2CID 54314728.
- ^ Брин, Аманда (7 мая 2008 г.). «Ученые открывают компас у мигрирующих бабочек». Гринвичский университет в Медуэе. п. 1. Архивировано из оригинал 30 мая 2012 г.. Получено 9 декабря 2009.
- ^ а б Чепмен, Джейсон У .; Reynolds, Don R .; Муритсен, Хенрик; Хилл, Джейн К .; Райли, Джо Р .; Сивелл, Дункан; Смит, Алан Д .; Войвод, Ян П. (8 апреля 2008 г.). «Выбор ветра и компенсация сноса оптимизируют пути миграции высоко летающей бабочки» (PDF). Текущая биология. 18 (7): 514–518. Дои:10.1016 / j.cub.2008.02.080. PMID 18394893. S2CID 3236142.
- ^ Srygley, Роберт Б .; Oliveira, Evandro G .; Риверос, Андре Дж. (2005). «Экспериментальные доказательства магнитного чутья у неотропических мигрирующих бабочек (Lepidoptera: Pieridae)» (PDF). Британский журнал поведения животных. 71 (1): 183–191. Дои:10.1016 / j.anbehav.2005.04.013. S2CID 25203050. Архивировано из оригинал (PDF) 4 ноября 2009 г.
- ^ Эллиот, Дебби; Беренбаум, май (18 августа 2007 г.). «Почему моль привлекает пламя? (Аудио)». Национальное общественное радио. п. 1. В архиве из оригинала от 8 января 2009 г.. Получено 12 декабря 2009.
- ^ Сяо, Генри С. (1972). Привлечение моли к свету и инфракрасному излучению. Сан-Франциско Пресс. ISBN 978-0-911302-21-9.
- ^ Ордер, Эрик; Фрост, Барри; Грин, Кен; Муритсен, Хенрик; Дрейер, Дэвид; Адден, Андреа; Браубургер, Кристина; Хайнце, Стэнли (2016). "Австралийский богонгский мотылек Агротис инфуса: Дальний ночной навигатор ». Границы поведенческой нейробиологии. 10: 77. Дои:10.3389 / fnbeh.2016.00077. ЧВК 4838632. PMID 27147998.
- ^ Уильямс, К. Б. (1927). «Исследование миграции бабочек на юге Индии и Цейлона, основанное в основном на записях гг. Эвершеда, Э. Э. Грина, Дж. К. Фрайера и У. Ормистона». Труды Лондонского королевского энтомологического общества. 75 (1): 1–33. Дои:10.1111 / j.1365-2311.1927.tb00054.x.
- ^ Urquhart, F.A .; Н. Р. Уркхарт (1977). «Районы зимовки и миграционные пути бабочки Монарх (Данаус п. плексипп, Lepidoptera: Danaidae) в Северной Америке с особым упором на западную популяцию ". Канадский энтомолог. 109 (12): 1583–1589. Дои:10.4039 / ent1091583-12.
- ^ Вассенаар Л. И .; К. А. Хобсон (1998). «Натальное происхождение мигрирующих бабочек-монархов в зимующих колониях в Мексике: новые изотопные доказательства». Труды Национальной академии наук. 95 (26): 15436–15439. Bibcode:1998PNAS ... 9515436W. Дои:10.1073 / пнас.95.26.15436. ЧВК 28060. PMID 9860986.
- ^ Смит, Н. Г. (1983). Янзен, Д. Х. (ред.). Урания фулгенс (Калипато Верде, Зеленая Урания). Коста-Рика естественная история. Чикаго: Издательство Чикагского университета. п. 816.
- ^ Чепмен, Р. Ф. (1998). Насекомые: строение и функции (4-е изд.). Нью-Йорк: Издательство Кембриджского университета. п.715. ISBN 978-0-521-57890-5.
Насекомые.
- ^ а б Мейер, Джон Р. (2006). «Акустическая коммуникация». Кафедра энтомологии, C Государственного университета. Архивировано из оригинал 20 июля 2011 г.. Получено 25 февраля 2011.
- ^ а б c d «Защитные механизмы гусеницы и бабочки». EnchantedLearning.com. Архивировано из оригинал 28 марта 2009 г.. Получено 7 декабря 2009.
- ^ Кричер, Джон (16 августа 1999 г.). "6". Неотропический компаньон. Princeton University Press. С. 157–158. ISBN 978-0-691-00974-2.
- ^ Santos, J.C .; Cannatella, D.C .; Канателла, округ Колумбия (2003). «Множественные, повторяющиеся причины апосематизма и специализации диеты на ядовитых лягушках». Труды Национальной академии наук. 100 (22): 12792–12797. Bibcode:2003ПНАС..10012792С. Дои:10.1073 / pnas.2133521100. ЧВК 240697. PMID 14555763. (Абстрактные).
- ^ «осметерий». Merriam-Webster, Incorporated. В архиве из оригинала 20 февраля 2010 г.. Получено 9 декабря 2009.
- ^ Хэдли, Дебби. «Осметериум». Руководство About.com. Архивировано из оригинал 23 июля 2008 г.. Получено 9 декабря 2009.
- ^ Латимер, Джонатан П .; Карен Стрэй Нолтинг (30 мая 2000 г.). Бабочки. Houghton Mifflin Harcourt Trade & Reference Publishers. п.12. ISBN 978-0-395-97944-0.
Махаон тигровый.
- ^ Насекомые и пауки мира. 10. Корпорация Маршалла Кавендиша. Маршалл Кавендиш. Январь 2003. С. 292–293. ISBN 978-0-7614-7344-2.CS1 maint: другие (ссылка на сайт)
- ^ Кэрролл, Шон (2005). Бесконечные формы - самые прекрасные: новая наука evodev и создание царства животных. W. W. Norton & Co., стр.205 –210. ISBN 978-0-393-06016-4.
Защита глазных пятен бабочки.
- ^ Ritland, D. B .; Л. П. Брауэр (1991). «Бабочка наместника не имитирует Бейтса». Природа. 350 (6318): 497–498. Bibcode:1991Натура.350..497р. Дои:10.1038 / 350497a0. S2CID 28667520.
Наместники так же неприятны, как монархи, и значительно более неприятны, чем королевы из репрезентативных популяций Флориды.
- ^ Мейер, А. (2006). «Повторяющиеся образцы мимикрии». PLOS Биология. 4 (10): e341. Дои:10.1371 / journal.pbio.0040341. ЧВК 1617347. PMID 17048984.
- ^ Джонс, Дж; D. Уотерс (2000). «Слух мотылька в ответ на эхолокационные крики летучей мыши регулируется независимо по времени и частоте». Труды Королевского общества B: биологические науки. 267 (1453): 1627–32. Дои:10.1098 / rspb.2000.1188. ЧВК 1690724. PMID 11467425.
- ^ Рэтклифф, Джон М .; Фуллард, Джеймс Н .; Артур, Бенджамин Дж .; Хой, Рональд Р. (2009). «Тигровая моль и угроза летучих мышей: принятие решений на основе активности одного сенсорного нейрона» (PDF). Письма о биологии. 5 (3): 368–371. Дои:10.1098 / рсбл.2009.0079. ЧВК 2679932. PMID 19324625. Архивировано из оригинал (PDF) 19 июля 2011 г.. Получено 11 февраля 2011.
- ^ а б Гилберт, Л. Э. (1972). «Питание пыльцой и репродуктивная биология Геликоний бабочки ". Труды Национальной академии наук. 69 (6): 1402–1407. Bibcode:1972ПНАС ... 69.1403Г. Дои:10.1073 / пнас.69.6.1403. ЧВК 426712. PMID 16591992.
- ^ Goulson, D .; Ollerton, J .; Слуман, К. (1997). "Стратегии поиска пищи у маленькой шкиперной бабочки, Thymelicus flavus: когда переключаться? ". Поведение животных. 53 (5): 1009–1016. Дои:10.1006 / anbe.1996.0390. S2CID 620334.
- ^ Хелен Дж. Янг; Лорен Гравиц (2002). "Влияние возраста стигмы на восприимчивость Silene alba (Caryophyllaceae) ". Американский журнал ботаники. 89 (8): 1237–1241. Дои:10.3732 / ajb.89.8.1237. PMID 21665724.
- ^ Oliveira PE; ЧП Гиббс; А.А. Барбоза (2004). «Опыление молью древесных пород в Серрадосе в Центральной Бразилии: случай, когда так много всего из-за столь немногих?». Систематика и эволюция растений. 245 (1–2): 41–54. Дои:10.1007 / s00606-003-0120-0. S2CID 21936259.
- ^ Деврис, П. Дж. (1988). "Личиночные муравьиные органы Thisbe irenea (Lepidoptera: Riodinidae) и их влияние на муравьев ". Зоологический журнал Линнеевского общества. 94 (4): 379–393. Дои:10.1111 / j.1096-3642.1988.tb01201.x.
- ^ Деврис, Пи (июнь 1990 г.). «Усиление симбиоза между гусеницами-бабочками и муравьями с помощью вибрационной коммуникации». Наука. 248 (4959): 1104–1106. Bibcode:1990Sci ... 248.1104D. Дои:10.1126 / science.248.4959.1104. PMID 17733373. S2CID 35812411.
- ^ Бентон, Франк (1895). Медоносная пчела: инструкция по пчеловодству [Самые известные бабочки Европы. Описание наиболее важных видов и инструкции по распознаванию и сбору бабочек и гусениц.]. 1–6, 33. Oestergaard Verlag. С. 113–114.
- ^ а б Рубинофф, Даниэль; Хейнс, Уильям П. (2005). «Паутина гусеница преследует улиток». Наука. 309 (5734): 575. Дои:10.1126 / наука.1110397. PMID 16040699. S2CID 42604851.
- ^ Пирс, Н. Э. (1995). «Хищные и паразитические чешуекрылые: хищники, живущие на растениях». Журнал общества лепидоптерологов. 49 (4): 412–453.
- ^ а б Грабе, Альберт (1942). Eigenartige Geschmacksrichtungen bei Kleinschmetterlingsraupen («Странные вкусы среди гусениц микромони») (PDF). 27 (на немецком языке). С. 105–109.
- ^ Гасми, Лайла; Булен, Элен; Готье, Джереми; Хуа-Ван, Орели; Мюссе, Карин; Якубовска, Агата К .; Ори, Жан-Марк; Волкова, Энн-Натали; Huguet, Элизабет (17 сентября 2015 г.). «Повторяющееся одомашнивание чешуекрылых генов от их паразитов, опосредованных браковирусами». PLOS Genet. 11 (9): e1005470. Дои:10.1371 / journal.pgen.1005470. ЧВК 4574769. PMID 26379286.
- ^ Шайх-Леско, Рина (17 сентября 2015 г.). "Гены паразитов сохраняются в геномах хозяев | The Scientist Magazine®". Ученый. В архиве из оригинала 21 декабря 2016 г.. Получено 13 июля 2016.
- ^ а б c d е ж г час я j Скобл, Малкольм Дж. (Сентябрь 1995 г.). "2". Чешуекрылые: форма, функции и разнообразие (1-е изд.). Оксфордский университет: Издательство Оксфордского университета. С. 4–5. ISBN 978-0-19-854952-9.
- ^ а б c d е ж г час я j k л м Гримальди, Д.; Энгель, М.С. (2005). Эволюция насекомых. Издательство Кембриджского университета. ISBN 978-0-521-82149-0.
- ^ Ржавчина, шутка (2000). «Палеонтология: летопись окаменелостей массовой миграции бабочек». Природа. 405 (6786): 530–531. Bibcode:2000Натура.405..530р. Дои:10.1038/35014733. PMID 10850702. S2CID 4374137.
- ^ van Eldijk, Timo J. B .; Вапплер, Торстен; Strother, Paul K .; ван дер Вейст, Каролин М. Х .; Раджаи, Хоссейн; Вишер, Хенк; ван де Шутбрюгге, Бас (10 января 2018 г.). "Триасово-юрское окно в эволюцию чешуекрылых". Достижения науки. 4 (1): e1701568. Bibcode:2018SciA .... 4E1568V. Дои:10.1126 / sciadv.1701568. ЧВК 5770165. PMID 29349295.
- ^ Гримальди, Дэвид А .; Майкл С. Энгель (2005). Эволюция насекомых. Издательство Кембриджского университета. п. 561. ISBN 978-0-521-82149-0. В архиве из оригинала 8 января 2014 г.. Получено 15 июля 2011.
- ^ Дэвис, Хейзел; Батлер, Кэрол А. (июнь 2008 г.). Кусают ли бабочки ?: увлекательные ответы на вопросы о бабочках и мотыльках. Издательство Университета Рутгерса. п. 48. ISBN 978-0-8135-4268-3. В архиве из оригинала 8 января 2014 г.. Получено 15 июля 2011.
- ^ Вольфрам Мей; Вилфрид Вичард; Патрик Мюллер; Бо Ван (2017). «Чертеж Amphiesmenoptera - Tarachoptera, новый отряд насекомых из бирманского янтаря (Insecta, Amphiesmenoptera)». Окаменелости. 20 (2): 129–145. Дои:10.5194 / fr-20-129-2017.
- ^ Кайла, Лаури; Марко Мутанен; Томми Найман (27 августа 2011 г.). «Филогения мега-разнообразных Gelechioidea (Lepidoptera): адаптации и детерминанты успеха». Молекулярная филогенетика и эволюция. 61 (3): 801–809. Дои:10.1016 / j.ympev.2011.08.016. PMID 21903172.
- ^ Уайтинг, Майкл Ф .; Whiting, Alison S .; Hastriter, Michael W .; Диттмар, Катарина (2008). «Молекулярная филогения блох (Insecta: Siphonaptera): происхождение и ассоциации хозяев». Кладистика. 24 (5): 677–707. CiteSeerX 10.1.1.731.5211. Дои:10.1111 / j.1096-0031.2008.00211.x. S2CID 33808144.
- ^ Йейтс, Дэвид К .; Вигманн, Брайан. «Насекомые Endopterygota с полным метаморфозом». Древо жизни. В архиве из оригинала 26 мая 2016 г.. Получено 24 мая 2016.
- ^ Уайтинг, Майкл Ф. (2002). «Mecoptera парафилетична: множественные гены и филогения Mecoptera и Siphonaptera». Zoologica Scripta. 31 (1): 93–104. Дои:10.1046 / j.0300-3256.2001.00095.x. S2CID 56100681. Архивировано из оригинал 5 января 2013 г.
- ^ Вигманн, Брайан; Йейтс, Дэвид К. (2012). Эволюционная биология мух. Издательство Колумбийского университета. п. 5. ISBN 978-0-231-50170-5.
- ^ Кристенсен, Н. П. (1999). «Бабочки, не относящиеся к глоссатану». В Н. П. Кристенсен (ред.). Чешуекрылые, мотыльки и бабочки Том 1: Эволюция, систематика и биогеография. Справочник по зоологии. Естественная история типов животного мира. Том IV Arthropoda: Insecta Part 35. Вальтер де Грюйтер. С. 41–49.
- ^ "Виды Agathiphaga queenslandensis Дамблтон, 1952 г. ". Справочник австралийской фауны. Департамент окружающей среды, воды, наследия и искусства. 9 октября 2008 г. В архиве из оригинала 5 апреля 2011 г.. Получено 31 августа 2010.
- ^ Beccaloni, G .; Скобл, М .; Китчинг, I .; Simonsen, T .; Робинсон, G .; Питкин, Б .; Hine, A .; Лял, К., ред. (2003). "Agathiphaga vitiensis". Глобальный указатель названий Lepidoptera. Музей естественной истории. Получено 8 мая 2018.
- ^ Beccaloni, G .; Скобл, М .; Китчинг, I .; Simonsen, T .; Робинсон, G .; Питкин, Б .; Hine, A .; Лял, К., ред. (2003). "Гетеробатмия". Глобальный указатель названий Lepidoptera. Музей естественной истории. Получено 8 мая 2018.
- ^ Ларсен, Торбен Б. (1994). «Бабочки Египта». Saudi Aramco World. 45 (5): 24–27. Архивировано из оригинал 13 января 2010 г.. Получено 18 декабря 2009.
- ^ «Стол с настоящими бабочками в смоле». Mfjoe.com. 18 декабря 2009 г. Архивировано с оригинал 6 мая 2010 г.. Получено 28 апреля 2012.
- ^ Рабуцци, Мэтью (ноябрь 1997 г.). "Этимология бабочки". Дайджест культурной энтомологии 4. Купертино, Калифорния: Bugbios. п. 4. Архивировано из оригинал 3 декабря 1998 г.. Получено 18 декабря 2009.
- ^ Хирн, Лафкадио (1904). Квайдан: Истории и исследования странной вещи. Dover Publications, Inc. ISBN 978-0-486-21901-1.
- ^ Миллер, Мэри (1993). Боги и символы древней Мексики и майя. Темза и Гудзон. ISBN 978-0-500-27928-1.
- ^ Кук, Келли А .; Weinzier, R. (2004). "IPM: Полевые культуры: кукурузный червь (Heliothis Zea)". IPM. п. 1. Архивировано из оригинал 9 февраля 2009 г.. Получено 17 января 2009.
- ^ Капинера, Джон Л. (2008). «Пестролистная совка, Peridroma saucia (Hübner) (Lepidoptera: Noctuidae)». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Springer Нидерланды. С. 4038–4041. Дои:10.1007/978-1-4020-6359-6_3936. ISBN 9781402062421.
- ^ Хан, Джефф (15 июня 2003 г.). «Дружелюбные мухи: хорошие новости, плохие новости». Новости Yard & Garden Line. Университет Миннесоты. 5 (9). Архивировано из оригинал 20 июля 2011 г.
- ^ Weinzierl, R .; Henn, T .; Koehler, P. G .; Такер, К. Л. (июнь 2005 г.). «Аттрактанты и ловушки для насекомых». Альтернативы в борьбе с насекомыми. Отделение энтомологии и нематологии Университета Флориды. Управление сельскохозяйственной энтомологии, Иллинойсский университет в Урбане-Шампейн. Архивировано из оригинал 11 марта 2011 г.
- ^ Goldsmith M. R .; Т. Шимада; Х. Абэ (2005). "Генетика и геномика тутового шелкопряда, Bombyx mori". Ежегодный обзор энтомологии. 50: 71–100. Дои:10.1146 / annurev.ento.50.071803.130456. PMID 15355234.
- ^ Ёситаке, Н. (1968). «Филогенетические аспекты происхождения японской расы тутового шелкопряда, Bombyx mori". Журнал серикологических наук Японии. 37: 83–87.
- ^ Кумбс, Э. М. (2004). Биологический контроль инвазивных растений в США. Корваллис: Издательство государственного университета Орегона. п. 146. ISBN 978-0-87071-029-2.
- ^ Фермы бабочек | Сохранение тропических лесов | Бабочка на ранчо В архиве 22 января 2008 г. Wayback Machine. butterfliesandart.com
- ^ Робинсон, Мартин; Бартлетт, Рэй и Уайт, Роб (2007) Корея. Публикации Lonely Planet, ISBN 978-1-74104-558-1. (стр.63)
- ^ Акунья, Ана Мария; Касо, Лаура; Алифат, Марио М .; Вергара, Карлос Х. (2011). «Съедобные насекомые как часть традиционной пищевой системы города Пополока Лос-Рейес-Метцонтла, Мексика». Журнал этнобиологии. 31 (1): 150–169. Дои:10.2993/0278-0771-31.1.150. S2CID 53963758.
- ^ Рамос-Элордуй, Джульета; Морено, член парламента Хосе; Васкес, Адольфо I .; Ландеро, Ивонн; Олива-Ривера, Эктор; Камачо, Виктор Х.М. (6 января 2011 г.). «Съедобные чешуекрылые в Мексике: географическое распространение, этническая принадлежность, экономическое и пищевое значение для сельских жителей». Журнал этнобиологии и этномедицины. 7: 2. Дои:10.1186/1746-4269-7-2. ISSN 1746-4269. ЧВК 3034662. PMID 21211040.
- ^ Загробельный, Мика; Дреон, Анджело Леандро; Гомьеро, Тициано; Marcazzan, Gian Luigi; Яркий, Миккель Андреас; Мёллер, Биргер Линдберг; Паолетти, Маурицио Г. (2009). «Ядовитая моль: источник действительно безопасного лакомства». Журнал этнобиологии. 29 (1): 64–76. Дои:10.2993/0278-0771-29.1.64. S2CID 73665731.
- ^ Диаз, HJ (2005). «Развитие глобальной эпидемиологии, синдромной классификации, управления и профилактики заражения гусеницами». Американский журнал тропической медицины и гигиены. 72 (3): 347–357. Дои:10.4269 / ajtmh.2005.72.347. PMID 15772333.
- ^ Redd, J .; Voorhees, R .; Торок, Т. (2007). «Вспышка лепидоптеризма в лагере бойскаутов». Журнал Американской академии дерматологии. 56 (6): 952–955. Дои:10.1016 / j.jaad.2006.06.002. PMID 17368636.
- ^ Kowacs, P.A .; Cardoso, J .; Entres, M .; Новак, Э. М .; Вернек, Л. К. (декабрь 2006 г.). «Смертельное внутримозговое кровоизлияние в результате Lonomia obliqua отравление гусеницей: история болезни ". Arquivos de Neuro-Psiquiatria. 64 (4): 1030–2. Дои:10.1590 / S0004-282X2006000600029. PMID 17221019. Бесплатный полный текст.
- ^ Патель Р.Дж., Шанбхаг Р.М. (1973). «Узелковая офтальмия - (история болезни)». Индийский журнал офтальмологии. 21 (4): 208. Архивировано с оригинал 12 сентября 2019 г.. Получено 17 ноября 2018.
- ^ Балит, Коррин Р .; Птолемей, Елена Ц .; Гири, Мерилин Дж .; Рассел, Ричард С .; Исбистер, Джеффри К. (2001). «Вспышка гусеничного дерматита, вызванная переносимыми по воздуху волосками мотылька омелы обыкновенной (Euproctis edwardsi)". Медицинский журнал Австралии. 175 (11–12): 641–3. Дои:10.5694 / j.1326-5377.2001.tb143760.x. PMID 11837874. S2CID 26910462. В архиве из оригинала 4 апреля 2011 г.. Получено 25 февраля 2011.. Бесплатный полный текст.
дальнейшее чтение
- Кристенсен, Н. П. (ред.) 1999. Чешуекрылые, мотыльки и бабочки. Том 1: Эволюция, систематика и биогеография. Handbuch der Zoologie. Eine Naturgeschichte der Stämme des Tierreiches / Справочник по зоологии. Естественная история типов животного мира. Группа / Том IV Arthropoda: Insecta Teilband / Part 35: 491 стр. Вальтер де Грюйтер, Берлин, Нью-Йорк.
- Немос Ф. (ок. 1895 г.). Europas bekannteste Schmetterlinge. Beschreibung der wichtigsten Arten und Anleitung zur Kenntnis und zum Sammeln der Schmetterlinge und Raupen [Самые известные бабочки Европы. Описание наиболее важных видов и инструкции по распознаванию и сбору бабочек и гусениц.] (PDF). Берлин: Oestergaard Verlag. Архивировано из оригинал (PDF) 24 июля 2011 г.
- Най, И. В. Б. и Флетчер, Д. С. 1991. Родовые названия бабочек мира. Том 6: xxix + 368 стр. Попечители Британского музея (естественная история), Лондон.
- О'Тул, Кристофер. 2002 г. Энциклопедия насекомых и пауков Firefly. ISBN 1-55297-612-2.
- Список используемой литературы
- Ламас, Херардо (1990). «Аннотированный список лепидоптерологических журналов» (PDF). Журнал исследований Lepidoptera. 29 (1–2): 92–104. Архивировано из оригинал (PDF) 27 августа 2016 г.. Получено 22 ноября 2012.
внешние ссылки
- Исторические иллюстрации мотылька
- Чешуекрылые у насекомых (Insecta) мира
- «Чешуекрылые». Интегрированная система таксономической информации.
- Между страницами: сокровища из коллекции Франклемона Виртуальная онлайн-выставка с подборкой исторических энтомологических сочинений и изображений из Энтомологической библиотеки Комстока при Корнельском университете.
- Региональные сайты
- Британские бабочки и мотыльки
- Бабочки Болгарии
- Бабочки Канады
- Фотография европейских бабочек и мотыльков
- Чешуекрылые французских Антильских островов
- Бабочки Индии
- Контрольный список бабочек в Индокитае
- Бабочки Ямайки
- Япмот Японская моль. Доступ к изображениям осуществляется с помощью чисел слева.
- Бабочки и мотыльки в Нидерландах
- Бабочки и мотыльки Северной Ирландии
- Бабочки Азиатской России
- Фотографии личинок и куколок бабочек и мотыльков. Испания
- Бабочки Астурии - Испания
- Шведские мотыльки и бабочки Lepidoptera (английский)
- Бабочки Турции
Насекомое заказы | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||
| |||||||||||||||
| |||||||||||||||
