IDH3A - IDH3A




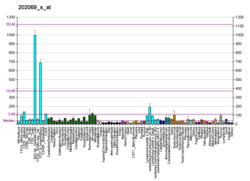
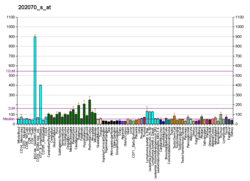
Субъединица альфа изоцитратдегидрогеназы [НАД], митохондриальная (IDH3α) является фермент что у людей кодируется IDH3A ген.[5][6]
Изоцитратдегидрогеназы (IDH) катализировать окислительный декарбоксилирование из изоцитрат к 2-оксоглутарат. Эти ферменты принадлежат к двум разным подклассам, один из которых использует НАД (+) как акцептор электронов а другой НАДФ (+). Сообщалось о пяти изоцитратдегидрогеназах: о трех НАД (+) - зависимых изоцитратдегидрогеназах, которые локализуются в митохондриальном матриксе, и о двух. НАДФ (+) - зависимые изоцитратдегидрогеназы, одна из которых митохондриальный а другой преимущественно цитозольный. НАД (+) - зависимые изоцитратдегидрогеназы катализируют аллостерически регулируемый ограничивающий шаг из цикл трикарбоновых кислот. Каждый изофермент представляет собой гетеротетрамер который состоит из двух альфа подразделения, одна бета-субъединица и одна гамма-субъединица. Белок, кодируемый этим геном, представляет собой альфа-субъединицу одного изофермента НАД (+) - зависимой изоцитратдегидрогеназы. [предоставлено RefSeq, июль 2008 г.][6]
Структура
IDH3 - один из трех изоферментов изоцитратдегидрогеназы, два других - IDH1 и IDH2, и кодируется одним из пяти генов изоцитратдегидрогеназы, которые IDH1, IDH2, IDH3A, IDH3B, и IDH3G.[7] Гены IDH3A, IDH3B, и IDH3G кодируют субъединицы IDH3, который является гетеротетрамер состоит из двух α-субъединиц 37 кДа (IDH3α), одной β-субъединицы 39 кДа (IDH3β) и одной γ-субъединицы 39 кДа (IDH3γ), каждая из которых имеет различные изоэлектрические точки.[8][9][10] Выравнивание их аминокислотные последовательности выявляет ~ 40% идентичности между IDH3α и IDH3β, ~ 42% идентичности между IDH3α и IDH3γ и еще более близкую идентичность 53% между IDH3β и IDH3γ, что составляет 34% идентичности и 23% сходства для всех трех типов субъединиц.[9][10][11][12] В частности, Arg 88 в IDH3α важен для каталитической активности IDH3, тогда как эквивалентные Arg99 в IDH3β и Arg97 в IDH3γ в значительной степени участвуют в аллостерической регуляции фермента с помощью ADP и NAD.[11] Таким образом, возможно, что эти субъединицы произошли от дупликация гена общего предкового гена и исходного каталитического Arg остаток были адаптированы к аллостерическим функциям в β- и γ-субъединицах.[9][11] Точно так же Жерех 181 в IDH3α необходим для катализа, в то время как эквивалентные Asp192 в IDH3β и Asp190 в IDH3γ усиливают НАД- и Mn2+-обвязка.[9] Поскольку окислительное декарбоксилирование, катализируемое IDH3, требует связывания НАД, Mn2+, а субстрат isocitrate, все три субъединицы участвуют в каталитической реакции.[10][11] Более того, исследования фермента в сердце свиньи показывают, что димеры αβ и αγ составляют два сайта связывания для каждого из его лиганды, включая изоцитрат, Mn2 + и NAD, в одном тетрамере IDH3.[9][10]
Функция
Как изоцитратдегидрогеназа, IDH3 катализирует необратимое окислительное декарбоксилирование изоцитрата с образованием α-кетоглутарат (α-KG) и CO2 как часть Цикл TCA в метаболизме глюкозы.[5][8][9][10][11] Этот шаг также учитывает сопутствующие снижение NAD + в NADH, который затем используется для генерации АТФ сквозь электронная транспортная цепь. Примечательно, что IDH3 полагается на NAD + в качестве своего акцептор электронов, в отличие от NADP +, такого как IDH1 и IDH2.[8][9] Активность IDH3 регулируется потребностями клетки в энергии: когда клетке требуется энергия, IDH3 активируется АДФ; а когда энергия больше не требуется, IDH3 ингибируется АТФ и NADH.[9][10] Эта аллостерическая регуляция позволяет IDH3 функционировать как лимитирующая ступень в цикле TCA.[5][13] Внутри клеток было обнаружено, что IDH3 и его субъединицы локализовать к митохондрии.[5][9][10]
Клиническое значение
Экспрессия IDH3α была связана с рак, с высокой базальной экспрессией в нескольких линиях раковых клеток и повышенной экспрессией, что свидетельствует о худшем прогнозе у онкологических больных. Предполагается, что IDH3α способствует росту опухоли в качестве регулятора α-KG, который впоследствии регулирует HIF-1. HIF-1 широко известен тем, что сдвигает метаболизм глюкозы с окислительного фосфорилирования к аэробике гликолиз в раковых клетках ( Эффект варбурга ). Более того, активность IDH3α приводит к ангиогенез и метаболическое перепрограммирование, чтобы обеспечить необходимые питательные вещества для непрерывного роста клеток. Между тем, подавление IDH3α препятствует росту опухоли. Таким образом, IDH3α может оказаться многообещающей терапевтической мишенью при лечении рака.[8]
IDH3α также участвует в психические расстройства. В частности, экспрессия IDH3α в мозжечок наблюдается значительно ниже для биполярное расстройство, сильное депрессивное расстройство, и шизофрения. Аномальные уровни IDH3α могут нарушить функцию митохондрий и способствовать патогенез этих расстройств.[13]
Мутации в этом гене были связаны с аутосомно-рецессивным пигментный ретинит.[14]
Смотрите также
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000166411 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000032279 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ссылка на Mouse PubMed:». Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d Huh TL, Kim YO, Oh IU, Song BJ, Inazawa J (март 1996). «Отнесение гена альфа-субъединицы изоцитратдегидрогеназы (IDH3A) к митохондриальной NAD + человека человека к 15q25.1 -> q25.2 посредством гибридизации in situ». Геномика. 32 (2): 295–6. Дои:10.1006 / geno.1996.0120. PMID 8833160.
- ^ а б «Ген Entrez: IDH3A изоцитратдегидрогеназа 3 (NAD +) альфа».
- ^ Димитров Л., Хонг С.С., Ян С., Чжуан З., Хейсс Д.Д. (2015). «Новые разработки в патогенезе и терапевтическом воздействии мутации IDH1 в глиоме». Международный журнал медицинских наук. 12 (3): 201–13. Дои:10.7150 / ijms.11047. ЧВК 4323358. PMID 25678837.
- ^ а б c d Zeng L, Morinibu A, Kobayashi M, Zhu Y, Wang X, Goto Y, Yeom CJ, Zhao T, Hirota K, Shinomiya K, Itasaka S, Yoshimura M, Guo G, Hammond EM, Hiraoka M, Harada H (сентябрь 2015 г.) ). «Аберрантная экспрессия IDH3α способствует росту злокачественной опухоли путем индукции HIF-1-опосредованного метаболического перепрограммирования и ангиогенеза». Онкоген. 34 (36): 4758–66. Дои:10.1038 / onc.2014.411. PMID 25531325.
- ^ а б c d е ж грамм час я Бзымек КП, Колман РФ (май 2007 г.). «Роль альфа-Asp181, бета-Asp192 и гамма-Asp190 в отличительных субъединицах человеческой НАД-специфической изоцитратдегидрогеназы». Биохимия. 46 (18): 5391–7. Дои:10.1021 / bi700061t. PMID 17432878.
- ^ а б c d е ж грамм Саундар С., О'хаган М., Фомулу К.С., Колман РФ (июль 2006 г.). «Идентификация Mn2 + -связывающих аспартатов из альфа, бета и гамма субъединиц НАД-зависимой изоцитратдегидрогеназы человека». Журнал биологической химии. 281 (30): 21073–81. Дои:10.1074 / jbc.m602956200. PMID 16737955.
- ^ а б c d е Soundar S, Park JH, Huh TL, Colman RF (декабрь 2003 г.). «Оценка с помощью мутагенеза важности 3 аргининов в альфа, бета и гамма субъединицах НАД-зависимой изоцитратдегидрогеназы человека». Журнал биологической химии. 278 (52): 52146–53. Дои:10.1074 / jbc.m306178200. PMID 14555658.
- ^ Данге М., Колман РФ (июль 2010 г.). «Каждый консервативный тир активного сайта в трех субъединицах изоцитратдегидрогеназы человека выполняет разные функции». Журнал биологической химии. 285 (27): 20520–5. Дои:10.1074 / jbc.m110.115386. ЧВК 2898308. PMID 20435888.
- ^ а б Йошими Н., Футамура Т., Берген С.Е., Иваяма Ю., Ишима Т., Селлгрен С., Экман С.Дж., Якобссон Дж., Полссон Е., Какумото К., Охги Ю., Йошикава Т., Ланден М., Хашимото К. (ноябрь 2016 г.). «Метаболомика спинномозговой жидкости определяет ключевую роль изоцитратдегидрогеназы в биполярном расстройстве: доказательства в поддержку гипотезы митохондриальной дисфункции». Молекулярная психиатрия. 21 (11): 1504–1510. Дои:10.1038 / mp.2015.217. ЧВК 5078854. PMID 26782057.
- ^ Питер В.Г., Никопулос К., Квинодос М., Грансе Л., Фаринелли П., Суперти-Фурга А., Андреассон С., Риволта С. (апрель 2019 г.). «Новый миссенс-вариант IDH3A вызывает аутосомно-рецессивный пигментный ретинит». Офтальмологическая генетика. 40 (2): 177–181. Дои:10.1080/13816810.2019.1605391. PMID 31012789.
дальнейшее чтение
- Андерсон Н.Л., Андерсон Н.Г. (ноябрь 2002 г.). «Протеом плазмы человека: история, характер, диагностические перспективы». Молекулярная и клеточная протеомика. 1 (11): 845–67. Дои:10.1074 / mcp.R200007-MCP200. PMID 12488461.
- Kim YO, Oh IU, Park HS, Jeng J, Song BJ, Huh TL (май 1995 г.). «Характеристика клона кДНК для человеческой альфа-субъединицы NAD (+) - изоцитратдегидрогеназы и структурное сравнение с ее изоферментами разных видов». Биохимический журнал. 308 (Пт 1): 63–8. Дои:10.1042 / bj3080063. ЧВК 1136843. PMID 7755589.
- Маруяма К., Сугано С. (январь 1994 г.). «Олиго-кэппинг: простой метод замены кэп-структуры эукариотических мРНК олигорибонуклеотидами». Ген. 138 (1–2): 171–4. Дои:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Судзуки Ю., Ёситомо-Накагава К., Маруяма К., Суяма А., Сугано С. (октябрь 1997 г.). «Создание и характеристика полноразмерной библиотеки кДНК, обогащенной по 5'-концу». Ген. 200 (1–2): 149–56. Дои:10.1016 / S0378-1119 (97) 00411-3. PMID 9373149.
- Kim YO, Koh HJ, Kim SH, Jo SH, Huh JW, Jeong KS, Lee IJ, Song BJ, Huh TL (декабрь 1999 г.). «Идентификация и функциональная характеристика новой тканеспецифической изоформы бета-субъединицы НАД (+) - зависимой изоцитратдегидрогеназы». Журнал биологической химии. 274 (52): 36866–75. Дои:10.1074 / jbc.274.52.36866. PMID 10601238.
- Вайс К., Цзэн Ю., Хуанг Дж., Собока М.Б., Рашбрук Дж. И. (февраль 2000 г.). «Бычья НАД + -зависимая изоцитратдегидрогеназа: альтернативный сплайсинг и тканезависимая экспрессия субъединицы 1». Биохимия. 39 (7): 1807–16. Дои:10.1021 / bi991691i. PMID 10677231.
- Адкинс Дж. Н., Варнум С. М., Обери К. Дж., Мур Р. Дж., Энджелл Н. Х., Смит Р. Д., Спрингер Д. Л., Паундс Дж. Г. (декабрь 2002 г.). «К протеому сыворотки крови человека: анализ методом многомерного разделения в сочетании с масс-спектрометрией». Молекулярная и клеточная протеомика. 1 (12): 947–55. Дои:10.1074 / mcp.M200066-MCP200. PMID 12543931.
- Soundar S, Park JH, Huh TL, Colman RF (декабрь 2003 г.). «Оценка с помощью мутагенеза важности 3 аргининов в альфа, бета и гамма субъединицах НАД-зависимой изоцитратдегидрогеназы человека». Журнал биологической химии. 278 (52): 52146–53. Дои:10.1074 / jbc.M306178200. PMID 14555658.
- Гуо Д., Хан Дж., Адам Б.Л., Колберн Н.Х., Ван М.Х., Донг З., Эйзирик Д.Л., Ше Дж. Х., Ван Си (декабрь 2005 г.). «Протеомный анализ субстратов SUMO4 в клетках HEK293 в условиях стресса, вызванного сывороточным голоданием». Сообщения о биохимических и биофизических исследованиях. 337 (4): 1308–18. Дои:10.1016 / j.bbrc.2005.09.191. PMID 16236267.
- Саундар С., О'хаган М., Фомулу К.С., Колман РФ (июль 2006 г.). «Идентификация Mn2 + -связывающих аспартатов из альфа, бета и гамма субъединиц НАД-зависимой изоцитратдегидрогеназы человека». Журнал биологической химии. 281 (30): 21073–81. Дои:10.1074 / jbc.M602956200. PMID 16737955.
- Бзымек КП, Колман РФ (май 2007 г.). «Роль альфа-Asp181, бета-Asp192 и гамма-Asp190 в отличительных субъединицах человеческой НАД-специфической изоцитратдегидрогеназы». Биохимия. 46 (18): 5391–7. Дои:10.1021 / bi700061t. PMID 17432878.