Триозофосфат изомераза - Triosephosphate isomerase
| триозофосфат изомераза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
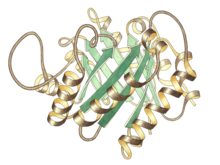 Вид сбоку мономера триозоп-изомеразы, активный центр вверху в центре | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 5.3.1.1 | ||||||||
| Количество CAS | 9023-78-3 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Триозофосфат изомераза (TPI или же ТИМ) является фермент (EC 5.3.1.1 ) который катализирует обратимое взаимопревращение триоза фосфат изомеры дигидроксиацетонфосфат и D-глицеральдегид-3-фосфат.
| Дигидроксиацетонфосфат | триозофосфат изомераза | D-глицеральдегид-3-фосфат | |
 |  | ||
| триозофосфат изомераза | |||
Сложный C00111 в КЕГГ База данных Pathway.Фермент 5.3.1.1 в КЕГГ База данных Pathway.Сложный C00118 в КЕГГ База данных Pathway.
TPI играет важную роль в гликолиз и необходим для эффективного производства энергии. ТПИ был обнаружен почти в каждом организме, в котором проводился поиск фермента, включая таких животных, как млекопитающие и насекомые а также в грибы, растения, и бактерии. Однако некоторые бактерии, которые не осуществляют гликолиз, например уреаплазма, отсутствие TPI.
У людей дефицит TPI связан с прогрессирующим тяжелым неврологическим расстройством, называемым дефицит триозофосфатизомеразы. Дефицит триозофосфатизомеразы характеризуется хроническим гемолитическая анемия. Хотя есть разные мутации , которые вызывают это заболевание, большинство из них включают мутацию глутаминовой кислоты в положении 104 на аспарагиновую кислоту.[1]
Триозофосфатизомераза - это высокоэффективный фермент, который в миллиарды раз выполняет реакцию быстрее, чем это происходило бы в растворе в естественных условиях. Реакция настолько эффективна, что ее называют каталитически совершенный: Это ограничено только скоростью, которую может размытый внутрь и из активного центра фермента.[2][3]
Механизм
Механизм включает промежуточное образование «эндиол». Относительная свободная энергия каждого основного состояния и переходного состояния была определена экспериментально и показана на рисунке.[2]

Структура TPI способствует превращению дигидроксиацетонфосфата (DHAP) в глицеральдегид-3-фосфат (GAP). В нуклеофильный глутамат 165 остатка TPI депротонирует субстрат,[4] и электрофильный остаток гистидина 95 отдает протон с образованием промежуточного эндиола.[5][6] При депротонировании ендиолат затем коллапсирует и, отрывая протон от протонированного глутамата 165, образует продукт GAP. Катализ обратной реакции протекает аналогично, образуя тот же ендиол, но с коллапсом ендиолятов из кислорода на C2.[7]
TPI имеет ограниченное распространение. С точки зрения термодинамики, образование DHAP в соотношении 20: 1 предпочтительнее образования GAP.[8] Однако при гликолизе использование GAP на последующих этапах метаболизма приводит к реакции, направленной на его производство. сульфат, фосфат, и арсенат ионы, которые связываются с активный сайт.[9] Другие ингибиторы включают 2-фосфогликолят, a аналог переходного состояния, и D-глицерин-1-фосфат, a аналог субстрата.[10]

Структура
| Триозофосфат изомераза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ТИМ | ||||||||
| Pfam | PF00121 | ||||||||
| Pfam клан | CL0036 | ||||||||
| ИнтерПро | IPR000652 | ||||||||
| PROSITE | PDOC00155 | ||||||||
| SCOP2 | 1 т / ч / Объем / СУПФАМ | ||||||||
| |||||||||
Триозофосфат-изомераза представляет собой димер идентичных подразделения, каждый из которых состоит примерно из 250 аминокислота остатки. Трехмерная структура субъединицы содержит восемь α-спирали снаружи и восемь параллельных β-тяжи изнутри. На иллюстрации ленточный остов каждой субъединицы окрашен от синего до красного от N-конца к C-концу. Этот структурный мотив называется αβ-бочкой или ТИМ-ствол, и является наиболее часто наблюдаемым белковая складка. В активный сайт этого фермента находится в центре ствола. А глютаминовая кислота остаток и гистидин участвуют в каталитический механизм. Последовательность вокруг остатков активного центра сохраняется во всех известных триозофосфат-изомеразах.
Структура триозофосфат-изомеразы способствует ее функции. Помимо точно размещенных остатков глутамата и гистидина для образования эндиола, цепь TPI из десяти или одиннадцати аминокислот действует как петля для стабилизации промежуточного продукта. Петля, образованная остатками от 166 до 176, замыкается и образует водородная связь к фосфатной группе субстрата. Это действие стабилизирует промежуточный эндиол и другие переходные состояния на пути реакции.[7]
В дополнение к тому, что реакция становится кинетически возможной, петля TPI изолирует реакционноспособный промежуточный эндиол, чтобы предотвратить разложение до метилглиоксаль и неорганический фосфат. Водородная связь между ферментом и фосфатной группой субстрата делает такое разложение стереоэлектронно невыгодным.[7] Метилглиоксаль - это токсин, который в случае образования выводится через глиоксалазная система.[11]Потеря высокоэнергетической фосфатной связи и субстрата для остального гликолиза делает образование метилглиоксаля неэффективным.
Исследования показывают, что лизин, расположенный рядом с активным центром (в положении 12), также имеет решающее значение для функции фермента. Лизин, протонированный при физиологическом pH, может помочь нейтрализовать отрицательный заряд фосфатной группы. Когда этот лизин превращается в нейтральную аминокислоту, TPI теряет все функции, но мутанты с другой положительно заряженной аминокислотой сохраняют некоторую функцию.[12]
Смотрите также
- ТИМ ствол
- Дефицит триозофосфат-изомеразы
- TPI1
- Триозофосфат изомераза в интерактивном 3D на Протеопедия
- Семейство триозофосфатизомеразы (ТИМ) в PROSITE
Рекомендации
- ^ Орос Ф., Олах Дж., Овади Дж. (Декабрь 2006 г.). «Дефицит триозофосфатизомеразы: факты и сомнения». IUBMB Life. 58 (12): 703–15. Дои:10.1080/15216540601115960. PMID 17424909.
- ^ а б Олбери В.Дж., Ноулз-младший (декабрь 1976 г.). «Профиль свободной энергии реакции, катализируемой триозофосфатизомеразой». Биохимия. 15 (25): 5627–31. Дои:10.1021 / bi00670a031. PMID 999838.
- ^ Роуз И.А., Фунг В.Дж., Warms JV (май 1990 г.). «Диффузия протонов в активном центре триозофосфатизомеразы». Биохимия. 29 (18): 4312–7. Дои:10.1021 / bi00470a008. PMID 2161683.
- ^ Альбер Т., Баннер Д.В., Блумер А.С., Петско Г.А., Филлипс Д., Риверс П.С., Уилсон И.А. (июнь 1981 г.). «О трехмерной структуре и каталитическом механизме триозофосфатизомеразы». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 293 (1063): 159–71. Дои:10.1098 / рстб.1981.0069. PMID 6115415.
- ^ Никбарг Е.Б., Давенпорт Р.К., Петско Г.А., Ноулз-младший (август 1988 г.). «Триозофосфат-изомераза: удаление предположительно электрофильного остатка гистидина приводит к незначительному изменению каталитического механизма». Биохимия. 27 (16): 5948–60. Дои:10.1021 / bi00416a019. PMID 2847777.
- ^ Комивес Э.А., Чанг Л.С., Лолис Э., Тилтон РФ, Петско Г.А., Ноулз-младший (март 1991 г.). «Электрофильный катализ в триозофосфатизомеразе: роль гистидина-95». Биохимия. 30 (12): 3011–9. Дои:10.1021 / bi00226a005. PMID 2007138.
- ^ а б c Ноулз-младший (март 1991 г.). «Ферментный катализ: не иначе, просто лучше». Природа. 350 (6314): 121–4. Дои:10.1038 / 350121a0. PMID 2005961.
- ^ Харрис Т.К., Коул Р.Н., Комер ФИ, Милдван А.С. (ноябрь 1998 г.). «Перенос протона в механизме триозофосфатизомеразы». Биохимия. 37 (47): 16828–38. Дои:10.1021 / bi982089f. PMID 9843453.
- ^ Lambeir AM, Opperdoes FR, Wierenga RK (октябрь 1987 г.). «Кинетические свойства триозофосфатизомеразы из Trypanosoma brucei brucei. Сравнение с мышечными и дрожжевыми ферментами кролика». Европейский журнал биохимии. 168 (1): 69–74. Дои:10.1111 / j.1432-1033.1987.tb13388.x. PMID 3311744.
- ^ Лолис Э., Петско Г.А. (июль 1990 г.). «Кристаллографический анализ комплекса между триозофосфатизомеразой и 2-фосфогликолятом с разрешением 2,5 А: значение для катализа». Биохимия. 29 (28): 6619–25. Дои:10.1021 / bi00480a010. PMID 2204418.
- ^ Крейтон ди-джей, Гамильтон Д.С. (март 2001 г.). «Краткая история глиоксалазы I и то, что мы узнали о зависимой от ионов металлов, ферментативно-катализируемой изомеризации». Архивы биохимии и биофизики. 387 (1): 1–10. Дои:10.1006 / abbi.2000.2253. PMID 11368170.
- ^ Лоди П.Дж., Чанг Л.К., Ноулз Дж. Р., Комивес Е.А. (март 1994 г.). «Триозофосфат-изомеразе требуется положительно заряженный активный сайт: роль лизина-12». Биохимия. 33 (10): 2809–14. Дои:10.1021 / bi00176a009. PMID 8130193.
внешняя ссылка
- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB для триозофосфатизомеразы человека
АТФ ADP АТФ ADP + + 2 ×  2 ×  2 × 3-фосфоглицерат 2 ×  2 × 2-фосфоглицерат 2 ×  2 × Фосфоенолпируват 2 ×  ADP АТФ 2 × Пируват 2 × |




