ТИМ ствол - TIM barrel
| ТИМ-цилиндр альдолазного типа | |
|---|---|
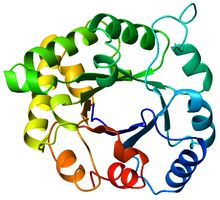 Вид сверху на триозофосфатизомераза (ТИМ) ствол (PDB: 8TIM), Окрашенные от синего (N-конец) до красного (C-конец). | |
| Идентификаторы | |
| Символ | Альдолаза_TIM |
| Pfam клан | CL0036 |
| ИнтерПро | IPR013785 |
| CATH | 8тим |
| SCOP2 | 8тим / Объем / СУПФАМ |
В ТИМ ствол, также известный как α / β ствол,[1]:252 это консервированный белковая складка состоящий из восьми α-спирали и восемь параллельных β-тяжи которые чередуются пептидный каркас.[2] Структура названа в честь триозофосфат изомераза, консервативный метаболический фермент.[3] Цилиндры TIM встречаются повсеместно, примерно 10% всех ферменты приняв эту складку.[4] Далее 5 из 7 комиссия по ферментам (EC) классы ферментов включают белки бочонка TIM.[5][6] Ствол TIM является эволюционно древним, многие из его элементов мало сходство сегодня,[7] вместо этого попадая в зона сумерек сходства последовательностей.[8][9]
Внутренний β-ствол во многих случаях стабилизирован сложным сети соляных мостов.[10] Петли на C-терминал концы β-ствола отвечают за каталитическую активность[11][12] в то время как N-концевые петли важны для устойчивости TIM-цилиндров. Конструкционные вставки от удлиненных петель до независимых домены могут быть вставлены вместо этих петель или в N / C-терминалы. Стволы TIM, кажется, эволюционировали дупликация гена и слияние доменов события полубочковых белков,[13] при этом большинство стволов TIM происходит из общий предок. Это привело к тому, что многие стволы TIM обладают внутренней симметрией.[14] Дальнейшие события дупликации генов этого предкового ствола TIM приводят к расхождению ферментов, обладающих функциональным разнообразием, наблюдаемым сегодня. Стволы TIM также были давней целью для протеиновые дизайнеры. Успешные конструкции цилиндров TIM включают как домены слияния существующих белков, так и de novo конструкции. Эксперименты по слиянию доменов привели ко многим успешным проектам,[15][16][17][18][19][20][21] в то время как de novo конструкции принесли успех только после 28 лет постепенного развития.[22]
Структура


Топология
Ствол ТИМ получил свое название от фермента триозофосфатизомеразы (ТИМ), который был первым белком, имеющим складку, который кристаллизовался.[3] Бочки ТИМ содержат 200-250 аминокислотных остатков,[2] сложены в 8 α-спиралей и 8 β-нитей. Β-тяжи расположены параллельно β-ствол, и окружены 8 α-спиралями. Определяющим свойством β-стволов TIM является то, что они всегда обладают числом сдвига 8.[2] Число сдвига определяется путем выбора остатка x на β-нити-1 и движения вдоль β-цилиндра в перпендикулярном направлении к направлению нитей до тех пор, пока не будет достигнут остаток y на исходной β-нити-1. Количество остатков между начальным и конечным положениями (| y-x |) является числом сдвига.[24] Поскольку количество нитей равно числу сдвига, боковые цепи поочередно направлены к поре и сердцевине, обеспечивая 4-кратную симметрию. Α-спирали окружают и полностью закрывают внутренний β-цилиндр. Короткие петли обычно соединяют вторичные структуры α и β, образуя (βα)8 повторить топологию. В некоторых случаях структуры от расширенных петель до независимых доменов могут быть вставлены вместо этих петель или могут быть присоединены к N / C-концам. Все ферменты TIM-цилиндра обладают каталитическими сайтами на С-конце β-цилиндра,[25] и структурные вставки, присутствующие рядом с этим концом, могут способствовать каталитической активности.
Сердцевина и поры
Цилиндры TIM содержат два разных похороненный области, где аминокислотные остатки полностью окружены своими соседями и не имеют доступа к растворителю. Термин «пора» используется неправильно, поскольку в этой области нет каналов для растворителя. Центральная область состоит из всех остатков, составляющих интерфейс α-β, и находится вне центральной β-бочки. Область пор состоит из всех внутренних остатков β-цилиндра, которые окружены и заключены в основу цепи β-цилиндра.
Из-за гофрированной природы β-тяжей альтернативные остатки вдоль цепи почти равномерно разделены между порами (53%) и сердцевиной (47%). Для β-стволов 95% остатков керна захоронены. Только 11% их основных остатков являются полярный, обладающие сродством к воде и обладающие способностью образовывать водородные связи или солевые мостики.[10] Точно так же 84% остатков пор β-нити захоронены. Однако 42% их поровых остатков полярны. Эти остатки образуют сложные сети солевых мостиков, чтобы компенсировать отсутствие доступа к растворителям.
Элементы стабилизации ствола TIM

Соляные мостики внутри поры ствола TIM, как полагают, вносят вклад в общую стабильность складки. Пример большой сети соляных мостов можно найти в 2-дезоксирибозо-5-фосфатальдолаза. Было обнаружено, что эта сеть сохраняется в семействе альдолаз класса I.
Точная причина чрезмерной представленности полярных остатков и солевых мостиков в поре остается неясной. Одно исследование предлагает улучшить складываемостьа не термодинамическая стабильность стволов TIM. Вовремя складывание В процессе, остатки внутренних пор на β-нитях будут подвергаться воздействию воды. Частично свернутые модули βαβα, называемые фолдонами, должны быть энергетически стабилизированы полярными остатками пор во время этой стадии сворачивания.
В другом исследовании с участием S. solfataricus индол-3-глицеринфосфатсинтаза Было обнаружено, что бочкообразный белок TIM, консервативный модуль βαβαβ, является важным шаблоном сворачивания, который направляет сворачивание других вторичных структур. Закрытие β-ствола происходило только в конце процесса складывания. Однако в этом случае авторы приписали разветвленные алифатические аминокислоты (валин, лейцин и изолейцин) в стабильности фолдона.
Еще одним стабилизирующим элементом стволов TIM является зажим β-шпильки. Доноры Н-связи боковой цепи на N-концах β-цепей с четными номерами часто образуют Н-связи с атомами водорода амидов основной цепи в предшествующих β-цепях с нечетными номерами. Эти зажимы (или аналоги гидрофобных мостиков боковой цепи) законсервированы в ортологах ствола TIM 3 индол-3-глицеринфосфатсинтазы из царства бактерий и архей, что означает, что они возникли в их последнем общем предке и сохранялись более миллиарда лет.
Конструкционные вставки

N / C-концевые и петлевые области на бочкообразных белках TIM способны содержать структурные вставки, начиная от простых вторичные структурные мотивы завершить домены. Эти домены способствуют распознаванию субстрата и каталитической активности. Ниже обсуждаются четыре различных примера цилиндров TIM, содержащих дополнительные мотивы и домены.
Bacillus subtilis Оротидин 5'-фосфатдекарбоксилаза (PDB: 1DBT) Представляет собой бочкообразный белок TIM, имеющий 4 α-спирали вместо петель βα, обычно присутствующих на С-конце β-ствола (остатки 35-42, 89-91, 126-133 и 215-219). Одна из этих спиралей (R215 → K219) содержит консервативный аргинин остаток (R215), необходимый для взаимодействия с фосфатной составляющей оротидин-5'-монофосфата. Было обнаружено, что другие спирали не содержат остатков, критических для каталитической активности, и могут выполнять структурные роли.
Бифункциональная гистидин-изомераза биосинтеза триптофана (PriA) Mycobacterium tuberculosis (PDB: 2Y85) Обладает способностью катализировать две реакции: (i) реакция HisA: превращение N - [(5-фосфорибозил) формимино] -5-аминоимидазол-4-карбоксамидрибонуклеотида (ProFAR) в N - [(5-фосфорибулозил) формимино] -5-аминоимидазол-4-карбоксамид рибонуклеотид (PRFAR) и (ii) реакция TrpF: N- (5'-фосфорибозил) антранилат (PRA) в 1- (O-карбоксифениламино) -1'-дезоксирибулозу-5 '-фосфат (CdRP). PriA представляет собой фермент в форме цилиндра TIM, который вмещает оба субстрата с помощью петель активного сайта (петли 1, 5 и 6, удлиненные петли βα на С-конце β-цилиндра), которые изменяют конформацию в зависимости от присутствующего реагента. Петля 1 охватывает активный сайт только в присутствии ProFAR. Loop5 наматывается на активный сайт, принимая конформацию β-листа в присутствии CdRP или узловую конформацию в присутствии ProFAR. Петля 6 охватывает активный центр для всех реагентов.
Lactococcus lactis Дигидрооротатдегидрогеназа А (ДХОДА) (PDB: 2DOR) Является примером ствола TIM, имеющего β-листы и удлиненные петли на С-конце концов β-ствола. ДГОДА катализирует окисление дигидрооротата до оротата, который является частью de novo Путь синтеза уридин-5'-монофосфата (UMP). Это окисление опосредуется флавинмононуклеотидом (FMN). Здесь β-листы и расширенные петли окружают активный центр, формируя полость, а также содержат несколько каталитических остатков.
В Methylophilus methylotrophus триметиламиндегидрогеназа (PDB: 2ТМД) TIM баррель - это пример полной вставки домена. Здесь Россманн фолд домен вставлен на C-конце TIM-ствола. Триметиламиндегидрогеназа катализирует превращение триметиламина в формальдегид. Для этой реакции требуется как восстановленный кофактор мононуклеотида 6-S-цистеинилфлавина (FMN), так и восстановленный железо-сера ([4Fe-4S]+) центр. FMN ковалентно связан в C-концевой области β-ствола. [4Fe-4S]+ центр слишком велик, чтобы поместиться в ствол ТИМ, и вместо этого расположен в непосредственной близости, на расстоянии 7 Å, на границе раздела между стволом ТИМ и доменами складки Россмана.
Складные механизмы
Консервация стволовой складки TIM отражается в сохранении ее равновесных и кинетических механизмов складывания у бактериальных паралогов с филогенетически отличными клонами. Химическая денатурация нескольких природных[27][28] и 2 варианта ствола TIM[28] неизменно включает густонаселенный промежуточный продукт равновесия. Кинетические интермедиаты, которые появляются после разбавления из сильно денатурирующих растворов, включают в себя ранние неправильно свернутые виды, которые должны, по крайней мере, частично разворачиваться, чтобы получить доступ к продуктивному пути складывания.[27][28] Ограничивающим скорость этапом сворачивания является закрытие 8-нитевого β-цилиндра, при этом предшествующая форма открытого бочонка соответствует равновесному промежуточному продукту.[29] Моделирование молекулярной динамики, ориентированное на естественных людей, резюмирует экспериментальные результаты и указывает путь к тестируемым вычислительным моделям для сложных механизмов сворачивания.[30]
Сохраненные фитнес-ландшафты
Бочковые белки TIM обладают необычно высокой пластичностью последовательностей, образуя большие семейства ортологичных и паралоговых ферментов в широко различных организмах. Эта пластичность предполагает ландшафт последовательностей, который позволяет адаптировать белок к множеству условий окружающей среды, в значительной степени независимо от филогенетической истории, при сохранении функции. Глубокое мутационное сканирование[31] подход и конкурентная проба[32] был использован для определения пригодности всех возможных аминокислотных мутантов по положениям в 3-х гипертермофильных ферментах TIM-бочки с гипертермофильной индол-3-глицеролфосфатсинтазой (IGPS) в поддержании роста дрожжевого хозяина, лишенного IGPS. Хотя 2 бактериальных и 1 архейный фермент IGPS были только на 30-40% идентичны по последовательности, их пригодность сильно коррелировала: одни и те же аминокислоты в одних и тех же положениях в трех разных белках имели очень похожую приспособленность. Корреляцию можно рассматривать как сохранение ландшафта приспособленности для фермента ствола TIM в течение эволюционного времени.
Области петли
Из примерно 200 остатков, необходимых для полного формирования ствола TIM, примерно 160 считаются структурно эквивалентными для разных белков, разделяющих эту складку. Остальные остатки расположены на участках петель, которые связывают спирали и нити; петли на C-терминал конец прядей имеет тенденцию содержать активный сайт, что является одной из причин, по которой эта складка настолько распространена: остатки, необходимые для поддержания структуры, и остатки, которые влияют на ферментативный Катализ - это по большей части отдельные подмножества:[33] Фактически, связывающие петли могут быть настолько длинными, что содержат другие белковые домены. Недавно было продемонстрировано, что каталитические петли могут обмениваться между различными ферментами TIM-барабанчика как полуавтономные единицы функциональных групп.[34]
Эволюция и происхождение


Преобладающая теория эволюции ствола TIM включает дупликацию и слияние генов, начиная с полубочки, которая в конечном итоге формирует полный ствол TIM. Множественные исследования подтверждают теорию дивергентная эволюция от одного предка и обсуждаются ниже.
Эволюция от общего предка
В начале 1990-х годов было отмечено, что все стволовые структуры ТИМ, решенные в то время, были ферментами, что указывало на расхождение от общего предка.[11][12] Кроме того, все цилиндры TIM обладают активными сайтами на С-конце β-цилиндров. предположили, что общий сайт связывания фосфата, образованный маленькой α-спиралью и цилиндрическими петлями TIM-7/8, сильно указывает на дивергентную эволюцию.[35] Дальнейшие исследования этих фосфатных групп показали, что 12 из 23 SCOP Семейства стволов TIM произошли от общего предка.[36] Точно так же были намеки на общее происхождение 17 из 21 CATH Семейства стволов TIM.[7] На основании этих сообщений считается правдоподобным, что большинство стволовых белков TIM произошли от общего предка.
Происхождение через дупликацию генов и слияние доменов

Многие бочкообразные белки ТИМ обладают 2-кратной, 4-кратной или 8-кратной внутренней симметрией, что позволяет предположить, что стволы ТИМ произошли от предковых (βα)4, (βα)2, или мотивы βα посредством дупликации генов и слияние доменов. Хороший пример 2-кратной внутренней симметрии наблюдается у ферментов ProFAR-изомераза (HisA) и имидазолглицеринфосфатсинтаза (HisF) Thermotoga maritima биосинтез гистидина путь.[13] Они катализируют 2 последовательные реакции в пути, обладают 25% гомологией последовательностей и обладают среднеквадратичными отклонениями (RMSD) в пределах 1,5-2 Å, что свидетельствует о расхождении от общего предка. Что еще интереснее, петли на С-концевых концах HisA и HisF обнаруживают двукратно повторяющийся паттерн, указывая тем самым, что их общий предок также обладал двукратной внутренней симметрией. На основе этих наблюдений была построена модель эволюции стволов TIM.[13] У предкового полубочки произошла бы дупликация и слияние генов, в результате чего получился бы единственный белок, содержащий два полубочковых домена. Структурные адаптации могли бы произойти, приводя к слиянию этих доменов с образованием закрытой β-бочки и формированию предковой бочки TIM. Функциональная адаптация также должна была произойти, что привело бы к развитию новой каталитической активности на С-конце β-цилиндра. На этом этапе общий предок HisA и HisF претерпел бы второе событие дупликации гена. Дивергентная эволюция дуплицированных генов предкового ствола TIM должна была привести к образованию HisA и HisF.
Интересно, что эта эволюционная модель была экспериментально подтверждена с использованием рационального дизайна белка и направленная эволюция. Höcker и другие. сначала слили две C-концевые половины HisF, получив HisF-CC. Затем эта конструкция была стабилизирована вставкой внутреннего соляной мост, давая HisF-C * C.[17] Дальнейшая ступенчатая стабилизация и солюбилизация HisF-C * C была достигнута за счет оптимизации границы раздела полубарабана, генерируя HisF-C ** C и HisF-C *** C, соответственно.[15][16] Кристаллическая структура HisF-C *** C выявила 2-кратно симметричный цилиндр TIM, что подтверждает возможность слияния естественных доменов. Более того, Höcker создал первые химерные стволы HisAF и HisFA TIM, используя полубочки HisA и HisF.[17] Эти эксперименты привели к предложению нового средства диверсификации и эволюции ферментов TIM-баррелей посредством обмена (βα) 4 полубочковых доменов среди уже существующих TIM-бочек. В соответствии с этой идеей установлена высокая каталитическая активность конструкции HisAF.[18] Аналогично химерный βα5-флаводоксин-подобная складка (CheY) / бочки HisF TIM,[19][20] и идеально 2-кратно симметричный ствол TIM на основе HisF[21][28] также были созданы.
Существование 4/8-кратной внутренней симметрии было предположено на основе вычислительного анализа последовательностей стволов TIM.[14] Например, кишечная палочка KDPG альдолаза[37] (PDB: 1FQ0) Было предложено обладать четкой 4-кратной симметрией с заметной 8-кратной симметрией. Конструкция 4-х кратно симметричного ствола ТИМ[22] подтвердили возможность более высоких порядков внутренней симметрии в естественных бочках из ТИМ и будут подробно обсуждены в следующем разделе. На сегодняшний день не поступало никаких экспериментальных доказательств существования 8-кратно симметричных стволов TIM.
De novo Конструкция ствола ТИМ
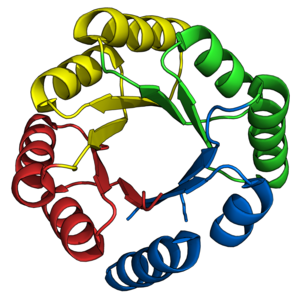
Складывание ствола TIM было давней целью для de novo протеиновые дизайнеры. Как описано ранее, многочисленные стволы TIM были успешно спроектированы на основе уже существующих естественных полубаррелей. Напротив, de novo Проектирование стволов TIM происходило поэтапно в течение 28 лет.[38]
Серия Octarellin[39][40][41][42][43] белков (Octarellin I → VI) были первыми попытками создать de novo Ствол ТИМ. Поскольку область дизайна белков все еще находилась в зачаточном состоянии, эти попытки дизайна имели лишь ограниченный успех. Хотя они отображали круговой дихроизм спектры согласуются с белками αβ и некоторыми характеристиками кооперативного сворачивания, все пептиды серии Octarellin были нерастворимы, и их нужно было реолюбилизировать из органы включения для дальнейшей характеристики. Интересно, что Octarellin V.1[44] показал Россманн -подобная складка в условиях сокристалла.
Серия белков симметрина (Симметрин-1 → 4) показала более благоприятные биофизические характеристики. Симметрин-1 был легко растворим, демонстрировал спектр кругового дихроизма, согласующийся с αβ белками, и демонстрировал превосходные характеристики кооперативного разворачивания и рефолдинга. Несмотря на эти достижения, все белки этого семейства проявляли характеристики расплава при анализе с использованием ЯМР (ядерный магнитный резонанс ), и дальнейших работ по разгадке их структур вести не удалось.
Белки серии sTIM[22] представлял первый успешный de novo Конструкция ствола ТИМ.[45][38] СТИМ-11 (PDB: 5BVL) Был разработан с внутренней 4-кратной симметрией, чтобы упростить вычислительный дизайн с использованием программного пакета Rosetta.[46] Полученные ранее первые принципы[47] были использованы для определения топологии и длины вторичной структуры. sTIM-11 оказался очень термостабильный, кооперативно складывающаяся конструкция, принявшая задуманную конструкцию
Смотрите также
Рекомендации
![]() Эта статья была адаптирована из следующего источника под CC BY 4.0 лицензия (2020 ) (отчеты рецензента ): «Ствол ТИМ» (PDF), WikiJournal of Science, 3 (1): 4, 2020, Дои:10.15347 / WJS / 2020.004, ISSN 2470-6345, Викиданные Q87400003
Эта статья была адаптирована из следующего источника под CC BY 4.0 лицензия (2020 ) (отчеты рецензента ): «Ствол ТИМ» (PDF), WikiJournal of Science, 3 (1): 4, 2020, Дои:10.15347 / WJS / 2020.004, ISSN 2470-6345, Викиданные Q87400003
- ^ Воет Д., Воет Дж. Г. (2011). «Глава 8. Трехмерные структуры белков». Биохимия (4-е изд.). John Wiley & Sons, Inc. ISBN 978-0470-91745-9.
- ^ а б c Веренга РК (март 2001 г.). «Сгиб TIM-бочонка: универсальная основа для эффективных ферментов». Письма FEBS. 492 (3): 193–8. Дои:10.1016 / s0014-5793 (01) 02236-0. PMID 11257493. S2CID 42044123.
- ^ Янсен Р., Герштейн М. (март 2000 г.). «Анализ дрожжевого транскриптома со структурными и функциональными категориями: характеристика высокоэкспрессированных белков». Исследования нуклеиновых кислот. 28 (6): 1481–8. Дои:10.1093 / nar / 28.6.1481. ЧВК 111042. PMID 10684945.
- ^ Нагано Н., Хатчинсон Э. Г., Торнтон Дж. М. (октябрь 1999 г.). «Бочкообразные структуры в белках: автоматическая идентификация и классификация, включая анализ последовательности бочек TIM». Белковая наука. 8 (10): 2072–84. Дои:10.1110 / пс 8.10.2072. ЧВК 2144152. PMID 10548053.
- ^ Уэбб ЕС (1992). Номенклатура ферментов: Рекомендации Номенклатурного комитета Международного союза биохимии и молекулярной биологии по номенклатуре и классификации ферментов. Академическая пресса. ISBN 978-0-12-227164-9.
- ^ а б Нагано Н., Оренго, Калифорния, Торнтон Дж. М. (август 2002 г.). «Одно устройство с множеством функций: эволюционные отношения между семействами стволов TIM, основанные на их последовательностях, структурах и функциях». Журнал молекулярной биологии. 321 (5): 741–65. Дои:10.1016 / с0022-2836 (02) 00649-6. PMID 12206759.
- ^ Livesay DR, La D (май 2005 г.). «Эволюционное происхождение и каталитическое значение консервативных электростатических сетей в белках TIM-баррель». Белковая наука. 14 (5): 1158–70. Дои:10.1110 / пс. 041221105. ЧВК 2253277. PMID 15840824.
- ^ Чунг С.Ю., Суббиа С. (октябрь 1996 г.). «Структурное объяснение сумеречной зоны гомологии белковой последовательности». Структура. 4 (10): 1123–7. Дои:10.1016 / s0969-2126 (96) 00119-0. PMID 8939745.
- ^ а б Виджаябаскар М.С., Вишвешвара С. (2012). «Понимание складчатой организации ствола TIM на основе взаимодействующих сетей структур на основе энергии». PLOS вычислительная биология. 8 (5): e1002505. Bibcode:2012PLSCB ... 8E2505V. Дои:10.1371 / journal.pcbi.1002505. ЧВК 3355060. PMID 22615547.
- ^ а б Фарбер Г.К., Петско Г.А. (июнь 1990 г.). «Эволюция альфа / бета ферментов ствола». Тенденции в биохимических науках. 15 (6): 228–34. Дои:10.1016 / 0968-0004 (90) 90035-А. PMID 2200166.
- ^ а б Рирдон Д., Фарбер Г.К. (апрель 1995 г.). «Структура и эволюция альфа / бета стволовых белков». Журнал FASEB. 9 (7): 497–503. Дои:10.1096 / fasebj.9.7.7737457. PMID 7737457.
- ^ а б c d Ланг Д., Тома Р., Хенн-Сакс М., Стернер Р., Вильманнс М. (сентябрь 2000 г.). «Структурные доказательства эволюции бета / альфа-бочонка каркаса путем дупликации и слияния генов». Наука. 289 (5484): 1546–50. Bibcode:2000Sci ... 289.1546L. Дои:10.1126 / science.289.5484.1546. PMID 10968789.
- ^ а б Сёдинг Дж, Реммерт М., Бигерт А. (июль 2006 г.). «HHrep: обнаружение повторов белка de novo и происхождение бочек TIM». Исследования нуклеиновых кислот. 34 (Проблема с веб-сервером): W137-42. Дои:10.1093 / нар / gkl130. ЧВК 1538828. PMID 16844977.
- ^ а б Зейтц Т., Бокола М., Кларен Дж., Стернер Р. (сентябрь 2007 г.).«Стабилизация 8-цилиндрового (бета-альфа) белка, разработанного из идентичных полубаррелей». Журнал молекулярной биологии. 372 (1): 114–29. Дои:10.1016 / j.jmb.2007.06.036. PMID 17631894.
- ^ а б Хеккер Б., Лохнер А., Зейтц Т., Кларен Дж., Штернер Р. (февраль 2009 г.). «Кристаллическая структура высокого разрешения искусственного (бета-альфа) (8) -баррельного белка, созданного из идентичных полубочек». Биохимия. 48 (6): 1145–7. Дои:10.1021 / bi802125b. PMID 19166324.
- ^ а б c Хеккер Б., Кларен Дж., Стернер Р., Макар А.Б., Макмартин К.Э., Палезе М., Тефли Т.Р. (июнь 1975 г.). «Формиатный тест в биологических жидкостях: применение при отравлении метанолом». Биохимическая медицина. 13 (2): 117–26. Дои:10.1016/0006-2944(75)90147-7. ЧВК 534502. PMID 15539462.
- ^ а б Кларен Дж., Малиси С., Хёкер Б., Стернер Р. (март 2009 г.). «Установление уровней каталитической активности дикого типа на естественных и искусственных (бета-альфа) 8-цилиндровых белковых каркасах». Труды Национальной академии наук Соединенных Штатов Америки. 106 (10): 3704–9. Bibcode:2009PNAS..106.3704C. Дои:10.1073 / pnas.0810342106. ЧВК 2656144. PMID 19237570.
- ^ а б Бхарат Т.А., Айзенбейс С., Зет К., Хёкер Б. (июль 2008 г.). «Бета-альфа-ствол, созданный комбинацией фрагментов из разных складок». Труды Национальной академии наук Соединенных Штатов Америки. 105 (29): 9942–7. Bibcode:2008PNAS..105.9942B. Дои:10.1073 / pnas.0802202105. ЧВК 2481348. PMID 18632584.
- ^ а б Eisenbeis S, Proffitt W, Coles M, Truffault V, Shanmugaratnam S, Meiler J, Höcker B (март 2012 г.). «Возможности рекомбинации фрагментов для рационального дизайна белков». Журнал Американского химического общества. 134 (9): 4019–22. Дои:10.1021 / ja211657k. PMID 22329686.
- ^ а б Fortenberry C, Bowman EA, Proffitt W, Dorr B, Combs S, Harp J и др. (Ноябрь 2011 г.). «Изучение симметрии как путь к вычислительному дизайну больших белковых доменов». Журнал Американского химического общества. 133 (45): 18026–9. Дои:10.1021 / ja210593m. ЧВК 3781211. PMID 21978247.
- ^ а б c Хуанг П.С., Фельдмайер К., Пармеджиани Ф., Веласко Д.А., Хёкер Б., Бейкер Д. (январь 2016 г.). «Дизайн De novo четырехсимметричного белка TIM-барреля с точностью до атомного уровня». Природа Химическая Биология. 12 (1): 29–34. Дои:10.1038 / nchembio.1966. ЧВК 4684731. PMID 26595462.
- ^ Нагараджан Д., Дека Г., Рао М. (август 2015 г.). «Дизайн симметричных стволовых белков TIM из первых принципов». BMC Биохимия. 16 (1): 18. Дои:10.1186 / s12858-015-0047-4. ЧВК 4531894. PMID 26264284.
- ^ Мурзин А.Г., Леск А.М., Чотия С. (март 1994 г.). «Принципы, определяющие структуру баррелей бета-листов в белках. I. Теоретический анализ». Журнал молекулярной биологии. 236 (5): 1369–81. Дои:10.1016/0022-2836(94)90064-7. PMID 8126726.
- ^ Брандин С. (1991). «Ствол ТИМ - наиболее часто встречающийся мотив сворачивания в белках». Текущее мнение в структурной биологии. 1 (6): 978–983. Дои:10.1016 / 0959-440x (91) 90094-а.
- ^ Барбер MJ, Neame PJ, Lim LW, White S, Matthews FS (апрель 1992 г.). «Корреляция рентгенологических и экспериментальных аминокислотных последовательностей триметиламиндегидрогеназы». Журнал биологической химии. 267 (10): 6611–9. PMID 1551870.
- ^ а б Форсайт В. Р., Билсель О, Гу З., Мэтьюз С. Р. (сентябрь 2007 г.). «Топология и последовательность в сворачивании ствола белка TIM: глобальный анализ подчеркивает разделение между промежуточными промежуточными продуктами сворачивания вне пути и стабильными промежуточными продуктами сворачивания на пути в сложном механизме сворачивания ствола (бета-альфа) 8 с неизвестной функцией из B. subtilis». Журнал молекулярной биологии. 372 (1): 236–53. Дои:10.1016 / j.jmb.2007.06.018. PMID 17619021.
- ^ а б c d Карстенсен Л., Сперл Дж. М., Бокола М., Список F, Шмид FX, Стернер Р. (август 2012 г.). «Сохранение механизма сворачивания между сконструированными первичными (βα) 8-цилиндровыми белками и их современным потомком». Журнал Американского химического общества. 134 (30): 12786–91. Дои:10.1021 / ja304951v. PMID 22758610.
- ^ Гу З., Рао М.К., Форсайт В.Р., Финке Дж.М., Мэтьюз С.Р. (ноябрь 2007 г.). «Структурный анализ кинетических промежуточных продуктов сворачивания для бочкообразного белка TIM, индол-3-глицеринфосфатсинтазы, с помощью масс-спектрометрии с водородным обменом и моделирования Gō». Журнал молекулярной биологии. 374 (2): 528–46. Дои:10.1016 / j.jmb.2007.09.024. ЧВК 2735044. PMID 17942114.
- ^ Halloran KT, Wang Y, Arora K, Chakravarthy S, Irving TC, Bilsel O, et al. (Август 2019 г.). «Расстройство и сворачивание бочкообразного белка TIM». Труды Национальной академии наук Соединенных Штатов Америки. 116 (33): 16378–16383. Дои:10.1073 / pnas.1900880116. ЧВК 6697809. PMID 31346089.
- ^ Чан Ю.Х., Венев С.В., Зельдович К.Б., Мэтьюз С.Р. (март 2017 г.). «Корреляция ландшафтов пригодности для трех ортологичных стволов TIM происходит из-за ограничений последовательности и структуры». Nature Communications. 8: 14614. Bibcode:2017НатКо ... 814614C. Дои:10.1038 / ncomms14614. ЧВК 5343507. PMID 28262665.
- ^ Hietpas RT, Jensen JD, Bolon DN (май 2011 г.). «Экспериментальное освещение фитнес-ландшафта». Труды Национальной академии наук Соединенных Штатов Америки. 108 (19): 7896–901. Дои:10.1073 / pnas.1016024108. ЧВК 3093508. PMID 21464309.
- ^ Очоа-Лейва А., Соберон X, Санчес Ф., Аргуэлло М., Монтеро-Моран Джи, Сааб-Ринкон Джи (апрель 2009 г.). «Дизайн белка посредством систематического каталитического обмена петлей в (бета / альфа) 8 раз». Журнал молекулярной биологии. 387 (4): 949–64. Дои:10.1016 / j.jmb.2009.02.022. PMID 19233201.
- ^ Очоа-Лейва А., Барона-Гомес Ф., Сааб-Ринкон Г, Вердел-Аранда К., Санчес Ф., Соберон X (август 2011 г.). «Изучение адаптируемости петли структура-функция (β / α) (8) -баррельского фермента посредством перестановки петель и вариабельности шарнира». Журнал молекулярной биологии. 411 (1): 143–57. Дои:10.1016 / j.jmb.2011.05.027. PMID 21635898.
- ^ Брандин К.И. (1991). «Ствол ТИМ - наиболее часто встречающийся мотив сворачивания в белках». Текущее мнение в структурной биологии. 1 (6): 978–983. Дои:10.1016 / 0959-440x (91) 90094-а.
- ^ Копли Р. Р., Борк П. (ноябрь 2000 г.). «Гомология среди (бета-альфа) (8) стволов: значение для эволюции метаболических путей». Журнал молекулярной биологии. 303 (4): 627–41. Дои:10.1006 / jmbi.2000.4152. PMID 11054297.
- ^ Ваймер Н., Бьюкенен Л. В., Хендерсон Д., Мета Н., Боттинг С. Н., Поцивавсек Л. и др. (Январь 2001 г.). «Направленная эволюция нового каталитического сайта в 2-кето-3-дезокси-6-фосфоглюконат альдолазе из Escherichia coli». Структура. 9 (1): 1–9. Дои:10.1016 / S0969-2126 (00) 00555-4. PMID 11342129.
- ^ а б Борман С (2015). «Белковые дизайнеры выкатывают бочку». Новости химии и машиностроения. 93 (47). п. 6.
- ^ Горадж К., Ренард А., Марсьяль Дж. А. (март 1990 г.). «Синтез, очистка и начальная структурная характеристика октареллина, полипептида de novo, смоделированного на основе альфа / бета-стволовых белков». Белковая инженерия. 3 (4): 259–66. Дои:10.1093 / белок / 3.4.259. PMID 2188263.
- ^ Борегар М., Горадж К., Гоффин В., Хереманс К., Гурмагтиг Э., Рюссхарт Дж. М., Марсьяль Дж. А. (октябрь 1991 г.). «Спектроскопическое исследование структуры октареллина (белка de novo, разработанного для упаковки в альфа / бета-цилиндр)». Белковая инженерия. 4 (7): 745–9. Дои:10.1093 / белок / 4.7.745. PMID 1798699.
- ^ Houbrechts A, Moreau B, Abagyan R, Mainfroid V, Préaux G, Lamproye A и др. (Март 1995 г.). «Октареллины второго поколения: два новых полипептида de novo (бета / альфа) 8, разработанные для исследования влияния упаковки бета-остатков на стабильность структуры альфа / бета-ствола». Белковая инженерия. 8 (3): 249–59. Дои:10.1093 / белок / 8.3.249. PMID 7479687.
- ^ Offredi F, Dubail F, Kischel P, Sarinski K, Stern AS, Van de Weerdt C и др. (Январь 2003 г.). «Дизайн остова и последовательности de novo идеализированного альфа / бета-цилиндрического белка: свидетельство стабильной третичной структуры» (PDF). Журнал молекулярной биологии. 325 (1): 163–74. Дои:10.1016 / S0022-2836 (02) 01206-8. PMID 12473459.
- ^ Фигероа М., Оливейра Н., Лежен А., Кауфманн К.В., Дорр Б.М., Матань А. и др. (2013). «Октареллин VI: использование розетки для создания предполагаемого искусственного (β / α) 8 белка». PLOS ONE. 8 (8): e71858. Bibcode:2013PLoSO ... 871858F. Дои:10.1371 / journal.pone.0071858. ЧВК 3747059. PMID 23977165.
- ^ Фигероа М., Слойтель М., Вандевенн М., Парвизи Дж., Аттаут С., Жакин О. и др. (Июль 2016 г.). «Неожиданная структура разработанного белка Octarellin V.1 создает проблему для инструментов прогнозирования структуры белка». Журнал структурной биологии. 195 (1): 19–30. Дои:10.1016 / j.jsb.2016.05.004. PMID 27181418.
- ^ Нанда V (январь 2016 г.). «Протеиновый дизайн: добраться до дна бочки ТИМ». Природа Химическая Биология. 12 (1): 2–3. Дои:10.1038 / nchembio.1987. PMID 26678608.
- ^ Кауфманн К.В., Леммон Г.Х., Делука С.Л., Шихан Дж. Х., Мейлер Дж. (Апрель 2010 г.). «Практически полезно: что может сделать для вас набор для моделирования белков Rosetta». Биохимия. 49 (14): 2987–98. Дои:10.1021 / bi902153g. ЧВК 2850155. PMID 20235548.
- ^ Кога Н., Тацуми-Кога Р., Лю Дж., Сяо Р., Acton TB, Montelione GT, Baker D (ноябрь 2012 г.). «Принципы построения идеальных белковых структур». Природа. 491 (7423): 222–7. Bibcode:2012Натура.491..222K. Дои:10.1038 / природа11600. ЧВК 3705962. PMID 23135467.
внешняя ссылка
- Список SCOP белков, принимающих бочкообразную складку TIM
- Бабу М.М. (1998). «Анализ ствола TIM». Центр биотехнологии Университета Анны.