Цикл Кори - Cori cycle
В Цикл Кори (также известный как цикл молочной кислоты), названный в честь его первооткрывателей, Карл Фердинанд Кори и Герти Кори,[1] это метаболический путь, в котором лактат производится анаэробным гликолиз в мышцах транспортируется в печень и превращается в глюкозу, которая затем возвращается в мышцы и циклически метаболизируется обратно в лактат.[2]
Процесс

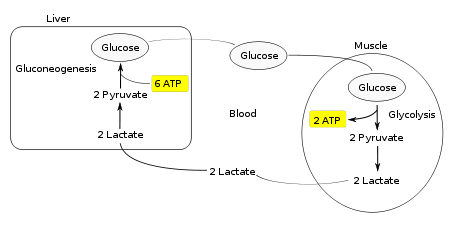
Мышечная активность требует АТФ, что обеспечивается разбивкой гликоген в скелетные мышцы. Распад гликогена, известный как гликогенолиз, релизы глюкоза в виде глюкозо-1-фосфат (G1P). G1P преобразован в G6P к фосфоглюкомутаза. G6P легко вводится в гликолиз, (или можно перейти в пентозофосфатный путь если концентрация G6P высока) процесс, который обеспечивает АТФ мышечные клетки как источник энергии. Во время мышечной активности необходимо постоянно пополнять запасы АТФ. Когда поставка кислород достаточно, эта энергия поступает от кормления пируват, один продукт гликолиза, в цикл лимонной кислоты, который в конечном итоге производит АТФ через кислородзависимую окислительного фосфорилирования.
Когда подача кислорода недостаточна, обычно во время интенсивной мышечной активности, энергия должна выделяться через анаэробный метаболизм. Молочнокислое брожение превращает пируват в лактат к лактатдегидрогеназа. Самое главное, ферментация восстанавливает НАД+, поддерживая его концентрацию, поэтому могут происходить дополнительные реакции гликолиза. Стадия ферментации окисляет НАДН производится путем гликолиза обратно в НАД+, переводя два электрона из НАДН для уменьшения пирувата в лактат. (См. Основные статьи на гликолиз и ферментация для подробностей.)
Вместо того, чтобы накапливаться внутри мышечных клеток, лактат, производимый анаэробной ферментацией, поглощается печень. Это инициирует вторую половину цикла Кори. В печени, глюконеогенез происходит. С интуитивной точки зрения глюконеогенез обращает вспять как гликолиз, так и ферментацию, превращая лактат сначала в пируват, а затем обратно в глюкозу. Затем глюкоза поступает в мышцы через кровоток; он готов к дальнейшим реакциям гликолиза. Если мышечная активность остановилась, глюкоза используется для пополнения запасов гликогена через гликогенез.[3]
В целом, этапы гликолиза цикла производят 2 молекулы АТФ при затратах 6 молекул АТФ, потребляемых на этапах глюконеогенеза. Каждая итерация цикла должна поддерживаться чистым потреблением 4 молекул АТФ. В результате цикл не может продолжаться бесконечно. Интенсивное потребление молекул АТФ в цикле Кори сдвигает метаболический нагрузка с мышц на печень.
Значимость
Важность цикла основана на предотвращении лактоацидоз во время анаэробных условий в мышцах. Однако обычно до того, как это произойдет, молочная кислота перемещается из мышц в печень.[3]
Кроме того, этот цикл важен для производства АТФ, источника энергии, во время мышечной нагрузки. Окончание мышечной нагрузки позволяет циклу Кори работать более эффективно. Это погашает кислородный долг, поэтому и цепь переноса электронов, и цикл лимонной кислоты могут производить энергию с оптимальной эффективностью.[3]
Цикл Кори - гораздо более важный источник субстрата для глюконеогенез чем еда.[4][5] Вклад лактата цикла Кори в общее производство глюкозы увеличивается с увеличением голодание продолжительность до выхода на плато.[6] В частности, после 12, 20 и 40 часов голодания добровольцев-людей на глюконеогенез приходится 41%, 71% и 92% продукции глюкозы, но вклад лактата цикла Кори в глюконеогенез составляет 18%, 35% и 36%. %, соответственно.[6] Оставшееся производство глюкозы происходит за счет распада белка,[6] мышечный гликоген,[6] и глицерин из липолиз.[7]
Наркотик метформин может вызвать лактоацидоз у пациентов с почечная недостаточность потому что метформин ингибирует печеночный глюконеогенез цикла Кори, особенно комплекс митохондриальной дыхательной цепи 1.[8] Накопление лактата и его субстратов для производства лактата, пирувата и аланина, приводит к избытку лактата.[9] Обычно избыток лактата выводится почками, но у пациентов с почечной недостаточностью почки не могут справиться с избытком молочной кислоты.
Смотрите также
Рекомендации
- ^ «Карл и Герти Кори и углеводный обмен». Американское химическое общество. 2004. Получено 12 мая 2020.
- ^ Нельсон, Дэвид Л. и Кокс, Майкл М. (2005) Принципы биохимии Ленингера, четвертое издание. Нью-Йорк: W.H. Фримен и компания, стр. 543.
- ^ а б c "Цикл Кори В архиве 2008-04-23 на Wayback Machine ". Получено 3 мая 2008 г. из Elmhurst, стр. 1–3.
- ^ Герих Дж. Э., Мейер С., Верле Х. Дж., Штумволл М. (2001). «Почечный глюконеогенез: его значение в гомеостазе глюкозы человека». Уход за диабетом. 24 (2): 382–391. Дои:10.2337 / diacare.24.2.382. PMID 11213896.
- ^ Nuttall FQ, Ngo A, Gannon MC (2008). "Регулирование производства глюкозы в печени и роль глюконеогенеза у человека: постоянна ли скорость глюконеогенеза?". Исследования и обзоры диабета / метаболизма. 24 (6): 438–458. Дои:10.1002 / дмрр.863. PMID 18561209. S2CID 24330397.
- ^ а б c d Кац Дж, Тайек Дж.А. (1998). «Глюконеогенез и цикл Кори у людей, голодавших в течение 12, 20 и 40 часов». Американский журнал физиологии. 275 (3, часть 1): E537 – E542. Дои:10.1152 / ajpendo.1998.275.3.E537. PMID 9725823.
- ^ Кэхилл Г.Ф. (2006). «Топливный обмен при голодании» (PDF). Ежегодный обзор питания. 26: 1–22. Дои:10.1146 / annurev.nutr.26.061505.111258. PMID 16848698.
- ^ Vecchio, S. et al. «Накопление метформина: лактоацидоз и высокие уровни метформина в плазме в серии ретроспективных клинических случаев у 66 пациентов, получающих хроническую терапию», Clin Toxicol. 2014; 52 (2).
- ^ Sirtori CR, Pasik C. «Повторная оценка бигуанида, метформина: механизм действия и переносимость». Pharmacol Res 1994; 30.
Источники
- Смит, А. Д., Датта, С. П., Смит, Г. Ховард, Кэмпбелл, П. Н., Бентли, Р. (редакторы) и др. (1997) Оксфордский словарь биохимии и молекулярной биологии. Нью-Йорк: Издательство Оксфордского университета.
Эта статья нужны дополнительные цитаты для проверка. (Апрель 2007 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |