Серпин - Serpin
| Серпин (ингибитор сериновой протеазы) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 Серпин (белый) с его «петлей реактивного центра» (синий), привязанный к протеаза (серый). Как только протеаза пытается катализ это будет необратимо подавленный. (PDB: 1К9О) | |||||||||||
| Идентификаторы | |||||||||||
| Символ | Серпин, СЕРПИН (корневой символ семьи) | ||||||||||
| Pfam | PF00079 | ||||||||||
| ИнтерПро | IPR000215 | ||||||||||
| PROSITE | PDOC00256 | ||||||||||
| SCOP2 | 1хле / Объем / СУПФАМ | ||||||||||
| CDD | cd00172 | ||||||||||
| |||||||||||
Серпины площадь надсемейство из белки с подобными структурами, которые впервые были идентифицированы для их ингибирование протеазы активности и встречаются во всех королевства жизни.[1] Аббревиатура серпин была первоначально придумана, потому что первые идентифицированные серпины действуют на химотрипсин-подобные сериновые протеазы (серине протация вгибиторы).[2][3] Они отличаются необычным механизмом действия, в котором они необратимо подавлять их цель протеаза пройдя большой конформационное изменение нарушить его активный сайт.[4][5] Это контрастирует с более распространенными конкурентный механизм для ингибиторов протеазы, которые связываются с активным сайтом протеазы и блокируют доступ к нему.[5][6]
Ингибирование протеазы серпинами контролирует множество биологических процессов, включая коагуляция и воспаление, и, следовательно, эти белки являются мишенью для медицинские исследования.[7] Их уникальное конформационное изменение также делает их интересными для структурная биология и сворачивание белка исследовательские сообщества.[4][5] Механизм конформационных изменений дает определенные преимущества, но он также имеет недостатки: серпины уязвимы для мутации что может привести к серпинопатиям, таким как неправильная упаковка белка и образование неактивных длинноцепочечных полимеры.[8][9] Серпин полимеризация не только снижает количество активного ингибитора, но также приводит к накоплению полимеров, вызывая смерть клетки и органная недостаточность.[7]
Хотя большинство серпинов контролируют протеолитический каскадов, некоторые белки со структурой серпина не ингибиторы ферментов, но вместо этого выполняют различные функции, такие как место хранения (как в Яичный белок —овальбумин ), транспорт как гормон белки-носители (тироксин-связывающий глобулин, кортизол-связывающий глобулин ) и молекулярный шаперон (HSP47 ).[6] Период, термин серпин также используется для описания этих членов, несмотря на их не ингибирующую функцию, поскольку они эволюционно связаны.[1]
История
Об активности протеазы в плазме крови впервые сообщили в конце 1800-х гг.[10] но только в 1950-х годах серпины антитромбин и альфа-1-антитрипсин были изолированы.[11] Первоначальные исследования были сосредоточены на их роли в заболеваниях человека: дефицит альфа-1-антитрипсина один из самых распространенных генетические нарушения, вызывая эмфизема,[8][12][13] и дефицит антитромбина приводит к тромбоз.[14][15]
В 1980-х годах стало ясно, что эти ингибиторы являются частью надсемейство из связанные с белки, которые включали оба ингибитора протеазы (например, альфа-1-антитрипсин ) и не ингибирующие члены (например, овальбумин ).[16] Название «серпин» было придумано на основе наиболее распространенной активности надсемейства (серине протация вгибиторы).[16] Примерно в то же время первый структуры были решены для серпиновых белков (сначала в релаксированной, а затем в стрессовой конформации).[17][18] Структуры показали, что механизм ингибирования включает необычные конформационные изменения и вызвал последующие структурный фокус серпиновых исследований.[5][18]
В настоящее время идентифицировано более 1000 серпинов, в том числе 36 человеческих белков, а также молекулы в целом. королевства жизни-животные, растения, грибы, бактерии, и археи -и немного вирусы.[19][20][21] В 2000-х годах была введена систематическая номенклатура для категоризации членов суперсемейства серпинов на основе их эволюционных отношений.[1] Таким образом, серпины являются самым большим и разнообразным суперсемейством ингибиторов протеаз.[22]
Мероприятия

Большинство серпинов протеаза ингибиторы, нацеленные на внеклеточные, химотрипсин -подобно сериновые протеазы. Эти протеазы обладают нуклеофильный серин остаток в каталитическая триада в их активный сайт. Примеры включают тромбин, трипсин, и эластаза нейтрофилов человека.[23] Серпины действуют как необратимый, ингибиторы суицида путем захвата промежуточного звена каталитического механизма протеазы.[24]
Некоторые серпины ингибируют другие классы протеаз, обычно цистеиновые протеазы, и называются «ингибиторами перекрестного класса». Эти ферменты отличаются от серинпротеаз тем, что они используют нуклеофильный цистеин остаток, а не серин, на их активном сайте.[25] Тем не менее, химический состав ферментов схож, и механизм ингибирования серпинами одинаков для обоих классов протеаз.[26] Примеры ингибирующих серпинов перекрестного класса включают: серпин В4 а плоскоклеточная карцинома антиген 1 (SCCA-1) и птичий серпин белок, специфичный для стадии терминации ядра миелоида и эритроида (MENT), которые подавляют папаин -подобно цистеиновые протеазы.[27][28][29]
Биологическая функция и локализация
Ингибирование протеазы
Примерно две трети серпинов человека выполняют внеклеточные роли, ингибируя протеазы в кровотоке, чтобы модулировать их активность. Например, внеклеточные серпины регулируют протеолитические каскады, лежащие в основе свертывание крови (антитромбин), воспалительный и иммунные ответы (антитрипсин, антихимотрипсин, и С1-ингибитор ) и ремоделирование тканей (PAI-1).[6] Подавляя сигнальный каскад протеазы, они также могут влиять на разработка.[30][31] В таблице серпинов человека (ниже) приведены примеры ряда функций, выполняемых серпином человека, а также некоторые заболевания, возникающие в результате дефицита серпина.
Протеазные мишени внутриклеточных ингибирующих серпинов трудно идентифицировать, поскольку многие из этих молекул, по-видимому, выполняют перекрывающиеся роли. Кроме того, многие серпины человека не имеют точных функциональных эквивалентов в модельных организмах, таких как мышь. Тем не менее, важной функцией внутриклеточных серпинов может быть защита от несоответствующей активности протеаз внутри клетки.[32] Например, один из наиболее охарактеризованных внутриклеточных серпинов человека - это Серпин Б9, что подавляет цитотоксический гранула протеаза гранзим B. При этом Serpin B9 может защищать от непреднамеренного высвобождения гранзима B и преждевременной или нежелательной активации смерть клетки пути.[33]
Немного вирусы используют серпины для нарушения функций протеазы в своем хозяине. В коровья оспа вирусный серпин CrmA (модификатор цитокинового ответа A) используется для предотвращения воспалительного процесса и апоптотический ответы инфицированных клеток-хозяев. CrmA увеличивает инфекционность, подавляя воспалительную реакцию своего хозяина путем ингибирования Ил-1 и Ил-18 обработка цистеиновой протеазой каспаза -1.[34] В эукариоты, серпин растения подавляет оба метакаспасы[35] и папаин-подобная цистеиновая протеаза.[36]
Неограничивающие роли
Неингибирующие внеклеточные серпины также выполняют широкий спектр важных ролей. Тироксин-связывающий глобулин и транскортин транспортировать гормоны тироксин и кортизол, соответственно.[37][38] Не ингибирующий серпин овальбумин это самый распространенный белок в Яичный белок. Его точная функция неизвестна, но считается, что это запасной белок для развивающийся плод.[39] Серпин 47 тепловой удар это сопровождающий, необходим для правильного складывание из коллаген. Он действует, стабилизируя коллаген тройная спираль пока он обрабатывается в эндоплазматический ретикулум.[40]
Некоторые серпины являются ингибиторами протеаз и выполняют дополнительные функции. Например, ядерный ингибитор цистеиновой протеазы MENT, в птицы также действует как ремоделирование хроматина молекула в птичьем красные кровяные тельца.[28][41]
Структура
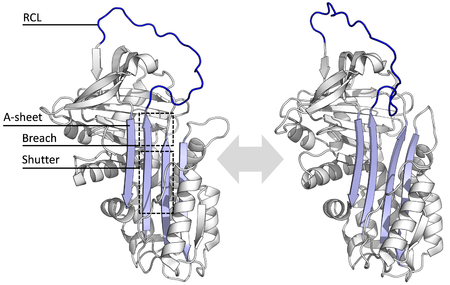
Все серпины имеют общий структура (или складывать), несмотря на их разнообразные функции. У всех обычно по три β-листы (названы A, B и C) и восемь или девять α-спирали (названы hA – hI).[17][18] Наиболее важными областями для функции серпина являются A-лист и петля реактивного центра (RCL). А-лист включает два β-тяжи которые находятся в параллельной ориентации с областью между ними, называемой «заслонкой», и верхней областью, называемой «проломом». RCL формирует начальное взаимодействие с протеазой-мишенью в ингибирующих молекулах. Были решены структуры, показывающие, что RCL полностью открыт или частично вставлен в A-лист, и предполагается, что серпины находятся в динамическое равновесие между этими двумя государствами.[5] RCL также только временно взаимодействует с остальной структурой и, следовательно, очень гибок и подвержен воздействию растворителя.[5]
Определенные серпиновые структуры охватывают несколько различных конформаций, что было необходимо для понимания их многоступенчатого механизма действия. Структурная биология поэтому играет центральную роль в понимании функции и биологии серпинов.[5]
Конформационное изменение и тормозной механизм
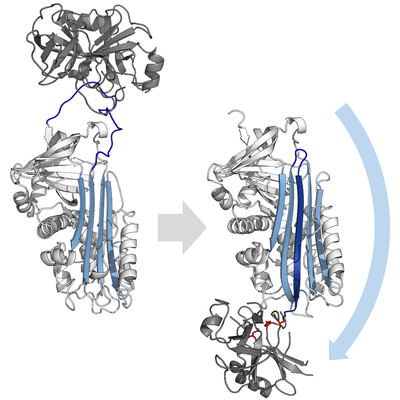
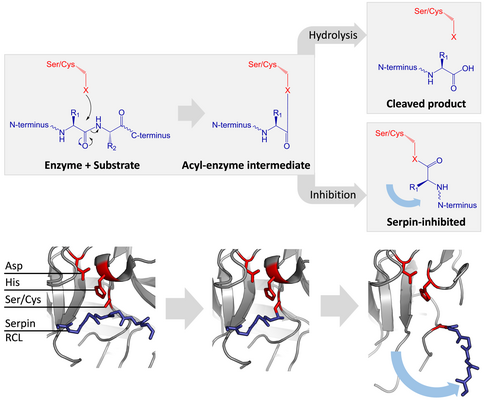
Тормозящие серпины не ингибируют свои целевые протеазы типичным конкурентный (замок и ключ) механизм, используемый большинством мелких ингибиторы протеазы (например. Ингибиторы типа Куница ). Вместо этого серпины используют необычный конформационное изменение, что нарушает структуру протеазы и препятствует ее завершению катализа. Конформационное изменение включает перемещение RCL к противоположному концу белка и его вставку в β-лист A, образуя дополнительный антипараллельный β-цепь. Это переводит серпин из напряженного состояния в расслабленное состояние с более низкой энергией (переход из S в R).[4][5][44]
Серин и цистеиновые протеазы катализировать разрыв пептидной связи с помощью двухэтапного процесса. Первоначально каталитический остаток активного центра триада выполняет нуклеофильный атака на пептидную связь субстрата. Это выпускает новый N-конец и образует ковалентный сложный эфир -связь между ферментом и субстратом.[4] Этот ковалентный комплекс между ферментом и субстратом называется промежуточное соединение ацил-фермента. Для стандартных субстраты, сложноэфирная связь гидролизованный и новый C-конец выпускается для полного катализа. Однако, когда серпин расщепляется протеазой, он быстро претерпевает переход от S к R до того, как промежуточный ацил-фермент гидролизуется.[4] Эффективность торможения зависит от того, что относительный кинетическая скорость конформационного изменения происходит на несколько порядков быстрее, чем гидролиз протеазой.
Поскольку RCL все еще ковалентно присоединен к протеазе через сложноэфирную связь, переход от S к R вытягивает протеазу сверху вниз серпина и искажает каталитическую триаду. Искаженная протеаза может гидролизовать промежуточный ацил-фермент исключительно медленно, поэтому протеаза остается ковалентно связанной от нескольких дней до недель.[24] Серпины классифицируются как необратимые ингибиторы и, как ингибиторы суицида поскольку каждый серпиновый белок навсегда инактивирует отдельную протеазу и может функционировать только один раз.[4]
Аллостерическая активация

В конформационная мобильность серпинов обеспечивает ключевое преимущество перед статическими ингибиторами протеазы типа "замок и ключ".[45] В частности, функция ингибирующих серпинов может быть регулируемый к аллостерический взаимодействие с конкретными кофакторы. В Рентгеновские кристаллические структуры из антитромбин, кофактор гепарина II, MENT и мышиный антихимотрипсин показывают, что эти серпины принимают конформацию, в которой первые две аминокислоты RCL вставлены в верхнюю часть A β-лист. Частично вставленная конформация важна, потому что кофакторы способны конформационно переключать определенные частично вставленные серпины в полностью вытесненную форму.[46][47] Эта конформационная перестройка делает серпин более эффективным ингибитором.
Типичным примером этой ситуации является антитромбин, который циркулирует в плазме в частично введенном относительно неактивном состоянии. Остаток, определяющий первичную специфичность (аргинин P1), указывает на тело серпина и недоступен для протеазы. При связывании высокоаффинной пентасахаридной последовательности в длинноцепочечной гепарин, антитромбин претерпевает конформационные изменения, изгнание RCL и воздействие аргинина P1. Таким образом, форма антитромбина, связанная с пентасахаридом гепарина, является более эффективным ингибитором тромбин и фактор Ха.[48][49] Кроме того, обе эти протеазы свертывания также содержат сайты связывания (называемые экзо-места ) для гепарина. Гепарин, таким образом, также действует как матрица для связывания как протеазы, так и серпина, еще более резко ускоряя взаимодействие между двумя сторонами. После начального взаимодействия образуется окончательный серпиновый комплекс и высвобождается гепариновый фрагмент. Это взаимодействие физиологически важно. Например, после повреждения стенки кровеносного сосуда гепарин подвергается воздействию, и антитромбин активируется, чтобы контролировать реакцию свертывания. Понимание молекулярной основы этого взаимодействия позволило разработать Фондапаринукс, синтетическая форма пентасахарида гепарина, используемого в качестве препарат, препятствующий свертыванию крови.[50][51]
Скрытая форма

Некоторые серпины спонтанно претерпевают переход из S в R, не будучи расщепленными протеазой, с образованием конформации, называемой латентным состоянием. Скрытые серпины не могут взаимодействовать с протеазами и поэтому больше не являются ингибиторами протеаз. Конформационное изменение латентности не совсем то же самое, что переход с S на R расщепленного серпина. Поскольку RCL все еще не поврежден, первая нить C-листа должна отслоиться, чтобы обеспечить полное введение RCL.[52]
Регулирование латентного перехода может действовать как механизм управления в некоторых серпинах, таких как PAI-1. Хотя PAI-1 продуцируется в ингибирующей S-конформации, он «автоинактивируется», переходя в латентное состояние, если он не связан с кофактором. витронектин.[52] Точно так же антитромбин также может спонтанно переходить в латентное состояние в качестве дополнительного механизма модуляции его аллостерической активации гепарином.[53] Наконец, N-конец tengpin, серпин из Thermoanaerobacter tengcongensis, требуется, чтобы заблокировать молекулу в нативном ингибирующем состоянии. Нарушение взаимодействий, осуществляемых N-концевой областью, приводит к спонтанному конформационному изменению этого серпина на латентную конформацию.[54][55]
Конформационное изменение не тормозящих функций
Некоторые неингибирующие серпины также используют конформационные изменения серпинов как часть своей функции. Например, родная (S) форма тироксин-связывающий глобулин имеет высокое сродство к тироксину, тогда как расщепленная (R) форма имеет низкое сродство. По аналогии, транскортин имеет более высокое сродство к кортизолу в нативном (S) состоянии, чем в расщепленном (R) состоянии. Таким образом, в этих серпинах расщепление RCL и переход с S на R были предприняты, чтобы позволить высвобождение лиганда, а не ингибирование протеазы.[37][38][56]
В некоторых серпинах переход с S на R может активировать клеточная сигнализация События. В этих случаях серпин, который образовал комплекс со своей протеазой-мишенью, затем распознается рецептором. Событие связывания затем приводит к передаче сигналов нижестоящим рецептором.[57] Поэтому переход от S к R используется для предупреждения клеток о наличии протеазной активности.[57] Это отличается от обычного механизма, при котором серпины влияют на передачу сигналов просто путем ингибирования протеаз, участвующих в каскаде передачи сигналов.[30][31]
Деградация
Когда серпин ингибирует целевую протеазу, он образует постоянный комплекс, от которого необходимо избавиться. Что касается внеклеточных серпинов, конечные комплексы серпин-фермент быстро выводятся из кровотока. Один из механизмов, с помощью которого это происходит у млекопитающих, - это белок, связанный с рецептором липопротеинов низкой плотности (LRP ), который связывается с ингибирующими комплексами, образованными антитромбином, PA1-1 и нейросерпином, вызывая клеточное поглощение.[57][58] Точно так же Дрозофила серпин, некротический, деградирует в лизосома после попадания в клетку рецептором липофорина-1 (гомологичен клеткам млекопитающих Рецептор ЛПНП семья).[59]
Заболевания и серпинопатии
Серпины участвуют в широком спектре физиологических функций, поэтому мутации в кодирующих их генах могут вызывать ряд заболеваний. Мутации, которые изменяют активность, специфичность или агрегационные свойства серпинов, влияют на то, как они функционируют. Большинство заболеваний, связанных с серпинами, являются результатом полимеризации серпинов в агрегаты, хотя встречаются и несколько других типов мутаций, связанных с заболеванием.[5][60] Расстройство дефицит α-антитрипсина - одно из самых распространенных наследственных заболеваний.[8][13]
Бездействие или отсутствие
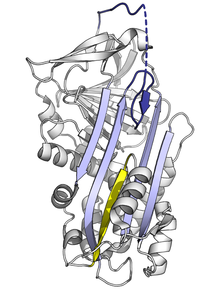
Поскольку напряженная серпиновая складка имеет высокую энергию, мутации могут привести к их неправильному изменению в более низкоэнергетические конформации (например, расслабленные или латентные) до того, как они правильно выполнят свою ингибирующую роль.[7]
Мутации, которые влияют на скорость или степень вставки RCL в A-лист, могут привести к тому, что серпин претерпит конформационное изменение S на R до того, как вступит в контакт с протеазой. Поскольку серпин может произвести это конформационное изменение только один раз, полученный серпин с пропуском осечки неактивен и не может должным образом контролировать свою целевую протеазу.[7][61] Точно так же мутации, которые способствуют несоответствующему переходу в мономерное латентное состояние, вызывают заболевание за счет уменьшения количества активного ингибирующего серпина. Например, варианты антитромбина, связанные с заболеванием покачиваться и колебаться,[62] оба способствуют формированию скрытое состояние.
Структура связанного с заболеванием мутанта антихимотрипсина (L55P) выявила другую, неактивную «δ-конформацию». В δ-конформации четыре остатка RCL вставлены в верхнюю часть β-листа A. Нижняя половина листа заполнена в результате частичного переключения одной из α-спиралей (F-спирали) на Конформация β-цепи, завершающая водородную связь β-слоя.[63] Неясно, могут ли другие серпины принимать этот конформер, и играет ли эта конформация функциональную роль, но предполагается, что δ-конформация может приниматься тироксин-связывающим глобулином во время высвобождения тироксина.[38] Не ингибирующие белки, относящиеся к серпинам, также могут вызывать заболевания при мутации. Например, мутации в SERPINF1 вызывают несовершенный остеогенез тип VI у человека.[64]
В отсутствие необходимого серпина протеаза, которую он обычно регулирует, чрезмерно активна, что приводит к патологиям.[7] Следовательно, простой недостаток серпина (например, нулевая мутация ) может привести к болезни.[65] Джин нокауты, особенно в мышей, используются экспериментально для определения нормальных функций серпинов по эффекту их отсутствия.[66]
Изменение специфики
В некоторых редких случаях изменение одной аминокислоты в RCL серпина изменяет его специфичность, чтобы воздействовать на неправильную протеазу. Например, мутация Антитрипсина-Питтсбурга (M358R) вызывает α1-антитрипсин серпин для ингибирования тромбина, вызывая кровотечение беспорядок.[67]
Полимеризация и агрегация


Большинство серпиновых заболеваний вызвано белковая агрегация и называются «серпинопатиями».[9][63] Серпины уязвимы для болезнетворных мутаций, которые способствуют образованию неправильно свернутых полимеров из-за их изначально нестабильных структур.[63] Хорошо охарактеризованные серпинопатии включают: дефицит α1-антитрипсина (альфа-1), который может вызвать семейную эмфизема, а иногда и печень цирроз, некоторые семейные формы тромбоз относится к дефицит антитромбина, типы 1 и 2 наследственный ангионевротический отек (HAE), связанные с дефицитом С1-ингибитор, и семейная энцефалопатия с телец включения нейросерпина (ФЕНИБ; редкий вид слабоумие вызванные полимеризацией нейросерпина).[8][9][68]
Каждый мономер серпинового агрегата существует в неактивной расслабленной конформации (с RCL, вставленным в A-лист). Следовательно, полимеры гиперстабильны к температуре и не могут ингибировать протеазы. Следовательно, серпинопатии вызывают патологии так же, как и другие протеопатии (например. прион болезней) через два основных механизма.[8][9] Во-первых, отсутствие активного серпина приводит к неконтролируемой активности протеаз и разрушению тканей. Во-вторых, сами сверхстабильные полимеры забивают эндоплазматический ретикулум клеток, которые синтезируют серпины, что в конечном итоге приводит к гибели клеток и повреждению тканей. В случае дефицита антитрипсина полимеры антитрипсина вызывают гибель печень клеток, что иногда приводит к повреждению печени и цирроз. Внутри клетки серпиновые полимеры медленно удаляются через деградацию в эндоплазматическом ретикулуме.[69] Однако детали того, как полимеры серпина вызывают гибель клеток, еще предстоит полностью понять.[8]
Считается, что физиологические серпиновые полимеры образуются замена домена события, когда сегмент одного серпинового белка вставляется в другой.[70] Обмен доменов происходит, когда мутации или факторы окружающей среды мешают заключительным стадиям сворачивания серпина до нативного состояния, вызывая неправильную укладку высокоэнергетических промежуточных продуктов.[71] Обе димер и тример структуры подкачки доменов были решены. В димере (антитромбина) RCL и часть A-листа включаются в A-лист другой молекулы серпина.[70] Тример с замененным доменом (антитрипсина) образуется посредством обмена совершенно другой областью структуры, B-листом (с RCL каждой молекулы, вставленным в свой собственный A-лист).[72] Также было высказано предположение, что серпины могут образовывать перестановки доменов, вставляя RCL одного белка в A-лист другого (полимеризация A-листа).[68][73] Эти димерные и тримерные структуры с замененными доменами считаются строительными блоками вызывающих заболевание полимерных агрегатов, но точный механизм до сих пор неясен.[70][71][72][74]
Лечебные стратегии
Несколько терапевтических подходов используются или исследуются для лечения наиболее распространенной серпинопатии: дефицита антитрипсина.[8] Антитрипсиновая терапия одобрена для лечения тяжелой эмфиземы, связанной с недостаточностью антитрипсина.[75] В этой терапии антитрипсин очищается из плазмы доноров крови и вводится внутривенно (сначала продается как Проластин ).[8][76] Для лечения тяжелых заболеваний легких и печени, связанных с недостаточностью антитрипсина. трансплантация доказал свою эффективность.[8][77] На животных моделях нацеливание генов в индуцированные плюрипотентные стволовые клетки успешно использовался для исправления дефекта полимеризации антитрипсина и восстановления способности печени млекопитающих секретировать активный антитрипсин.[78] Также были разработаны небольшие молекулы, которые блокируют полимеризацию антитрипсина. in vitro.[79][80]
Эволюция
Серпины являются наиболее широко распространенным и самым крупным суперсемейством ингибиторов протеаз.[1][22] Первоначально считалось, что они ограничены эукариот организмов, но с тех пор были обнаружены в бактерии, археи и немного вирусы.[19][20][81] Остается неясным, являются ли гены прокариот потомками предкового прокариотического серпина или продуктом горизонтальный перенос генов от эукариот. Большинство внутриклеточных серпинов относятся к одному филогенетический clade, независимо от того, происходят ли они от растений или животных, что указывает на то, что внутриклеточные и внеклеточные серпины могли разойтись раньше растений и животных.[82] Исключения включают внутриклеточный серпин теплового шока HSP47, который является шапероном, необходимым для правильного сворачивания коллаген, и циклы между цис-Гольджи и эндоплазматический ретикулум.[40]
Считается, что ингибирование протеазы является наследственной функцией, а не ингибирующие члены - результатами эволюционного развития. неофункционализация конструкции. Конформационное изменение S на R также было адаптировано некоторыми связывающими серпинами для регулирования сродства к своим мишеням.[38]
Распределение
Животное
Человек
Геном человека кодирует 16 серпиновых клад, называемых от serpinA до serpinP, включая 29 ингибирующих и 7 неингибирующих серпиновых белков.[6][66] Система именования человеческих серпинов основана на филогенетический анализ примерно 500 серпинов из 2001 г. с белками, названными serpinXY, где X - кладовая белка, а Y - количество белков в этой кладе.[1][19][66] Функции серпинов человека были определены комбинацией биохимический исследования, человек генетические нарушения, и нокаут модели мыши.[66]
| Таблица серпинов человека | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Специализированные змеи млекопитающих
Много млекопитающее Были идентифицированы серпины, которые не имеют очевидной ортологии с аналогом серпина человека. Примеры включают многочисленные грызун змеи (особенно некоторые из мышиный внутриклеточные серпины), а также маточные змеи. Термин маточный серпин относится к членам клады серпина А, которые кодируются геном SERPINA14. Серпины матки производятся эндометрий ограниченной группы млекопитающих в Лавразиатерия клады под влиянием прогестерон или же эстроген.[167] Они, вероятно, не являются функциональными ингибиторами протеиназ и могут действовать во время беременности, подавляя иммунные реакции матери против концептус или участвовать в трансплацентарной транспортировке.[168]
Насекомое
В Drosophila melanogaster геном содержит 29 генов, кодирующих серпины. Анализ аминокислотной последовательности поместил 14 из этих серпинов в серпиновую кладу Q и три - в серпиновую кладу K, а остальные двенадцать классифицированы как сирпины, не принадлежащие ни к какой кладе.[169] Систему классификации клад трудно использовать для Дрозофила серпинов, и вместо этого была принята система номенклатуры, которая основана на положении серпиновых генов на Дрозофила хромосомы. Тринадцать из Дрозофила серпины встречаются в геноме как изолированные гены (включая Serpin-27A, см. ниже), а остальные 16 организованы в пять кластеров генов, которые расположены в хромосомных положениях 28D (2 серпина), 42D (5 серпинов), 43A (4 серпина), 77B (3 серпина) и 88E (2 серпина).[169][170][171]
Исследования по Дрозофила серпины показывают, что Serpin-27A ингибирует протеазу Easter (последняя протеаза в протеолитическом каскаде Nudel, Gastrulation Defective, Snake и Easter) и, таким образом, контролирует формирование дорсовентрального паттерна. Пасхальная функция расщепляет Spätzle (лиганд хемокинового типа), что приводит к платный сигнализация. Помимо своей центральной роли в формировании эмбрионального паттерна, передача сигналов toll также важна для врожденный иммунный ответ у насекомых. Соответственно, serpin-27A также контролирует иммунный ответ насекомых.[31][172][173] В Тенебрио Молитор (большой жук), белок (SPN93), содержащий два дискретных тандемных серпиновых домена, функционирует, чтобы регулировать протеолитический каскад толл.[174]
Нематода
Геном нематода червь C. elegans содержит 9 серпинов, все из которых не имеют сигнальных последовательностей и поэтому, вероятно, являются внутриклеточными.[175] Однако только 5 из этих серпинов, по-видимому, действуют как ингибиторы протеазы.[175] Один, SRP-6, выполняет защитную функцию и защищает от стресса, вызванного Кальпаин -ассоциированное нарушение лизосом. Кроме того, SRP-6 ингибирует лизосомальные цистеиновые протеазы, высвобождаемые после разрыва лизосомы. Соответственно, черви, лишенные SRP-6, чувствительны к стрессу. В частности, черви с нокаутом SRP-6 погибают при помещении в воду (летальный фенотип при гипоосмотическом стрессе или Osl). Поэтому было высказано предположение, что лизосомы играют общую и контролируемую роль в определении судьбы клеток.[176]
Растение
Растение серпины были одними из первых идентифицированных членов суперсемейства.[177] Серпиновый протеин Z ячменя содержится в большом количестве в зерне ячменя и является одним из основных белковых компонентов пива. Геном модельного растения, Arabidopsis thaliana содержат 18 серпиноподобных генов, хотя только 8 из них представляют собой полноразмерные последовательности серпинов.
Серпины растений являются мощными ингибиторами химотрипсиноподобных сериновых протеаз млекопитающих. in vitroНаиболее изученным примером является серпин Zx ячменя (BSZx), который способен ингибировать трипсин и химотрипсин, а также несколько факторов свертывания крови.[178] Однако близкие родственники химотрипсиноподобных сериновых протеаз у растений отсутствуют. RCL нескольких серпинов из зерна пшеницы и ржи содержат последовательности повторов поли-Q, подобные тем, которые присутствуют в проламин запасные белки эндосперма.[179][180] Поэтому было высказано предположение, что серпины растений могут подавлять протеазы насекомых или микробов, которые в противном случае переваривали бы запасные белки зерна. В поддержку этой гипотезы в соке флоэмы тыквы были идентифицированы специфические серпины растений (CmPS-1).[181] и огурцы.[182][183] Хотя наблюдалась обратная корреляция между повышающей регуляцией экспрессии CmPS-1 и выживаемостью тли, in vitro эксперименты по кормлению показали, что рекомбинантный CmPS-1 не влияет на выживаемость насекомых.[181]
Были предложены альтернативные роли и мишени протеаз для серпинов растений. В Арабидопсис серпин, АтСерпин1 (At1g47710; 3LE2), Опосредует контроль заданного значения над запрограммированной гибелью клеток, воздействуя на папаин-подобную цистеинпротеазу «Responsive to Desiccation-21» (RD21).[36][184] AtSerpin1 также подавляет метакаспаза -подобные протеазы in vitro.[35] Два других Арабидопсис серпины, AtSRP2 (At2g14540) и AtSRP3 (At1g64030), по-видимому, участвуют в ответах на повреждение ДНК.[185]
Грибковые
Один грибковый Серпин на сегодняшний день охарактеризован: целпин из Пиромицеты виды штамм E2. Пиромицеты это род анаэробных грибов, обнаруженных в кишечнике жвачных животных, и важен для переваривания растительного материала. Предполагается, что Celpin обладает ингибирующим действием и содержит два N-концевых докерин домены в дополнение к его серпиновому домену. Докерины обычно содержатся в белках, которые локализуются в грибковых целлюлосома, большой внеклеточный мультипротеиновый комплекс, расщепляющий целлюлозу.[21] Поэтому предполагается, что целпин может защищать целлюлосому от протеаз растений. Определенные бактериальные серпины локализуются аналогично целлюлосоме.[186]
Прокариотический
Предполагаемые гены серпинов спорадически распространены в прокариоты. В пробирке исследования некоторых из этих молекул показали, что они способны ингибировать протеазы, и предполагается, что они действуют как ингибиторы in vivo. Несколько прокариотных серпинов обнаружены в экстремофилы. Соответственно, и в отличие от серпинов млекопитающих, эти молекулы обладают повышенной устойчивостью к тепловой денатурации.[187][188] Точная роль большинства бактериальных серпинов остается неясной, хотя Clostridium thermocellum Серпин располагается в целлюлосома. Предполагается, что роль серпинов, связанных с целлюлосомой, может заключаться в предотвращении нежелательной активности протеаз против целлюлосомы.[186]
Популярный
Серпины также выражаются вирусы как способ уклониться от иммунной защиты хозяина.[189] В частности, серпины, экспрессируемые вирусы оспы, включая коровья оспа (коровьей оспы) и кроличья оспа (миксома), представляют интерес из-за их потенциального использования в качестве новых терапевтических средств при иммунных и воспалительных расстройствах, а также в терапии трансплантатов.[190][191] Serp1 подавляет TLR-опосредованный врожденный иммунный ответ и допускает неопределенное сердечное аллотрансплантат выживаемость у крыс.[190][192] Crma и Serp2 являются ингибиторами перекрестного класса и нацелены как на серин (гранзим B; хотя и слабо), так и на цистеиновые протеазы (каспаза 1 и каспаза 8).[193][194] По сравнению со своими аналогами у млекопитающих вирусные серпины содержат значительные делеции элементов вторичной структуры. В частности, crmA лишена D-спирали, а также значительных частей A- и E-спиралей.[195]
Смотрите также
Рекомендации
- ^ а б c d е Сильверман Г.А., Берд П.И., Каррелл Р.В., Черч ФК, Кафлин П.В., Геттинс П.Г., Ирвинг Д.А., Ломас Д.А., Люк С.Дж., Мойер Р.В., Пембертон П.А., Ремольд-О'Доннелл Е., Салвесен Г.С., Трэвис Дж., Уиссток Дж. 2001). «Серпины представляют собой расширяющееся суперсемейство структурно похожих, но функционально разнообразных белков. Эволюция, механизм ингибирования, новые функции и пересмотренная номенклатура». Журнал биологической химии. 276 (36): 33293–6. Дои:10.1074 / jbc.R100016200. PMID 11435447.
- ^ Сильверман Г.А., Виссток Дж. К., Боттомли С. П., Хантингтон Дж. А., Кайзерман Д., Люк С. Дж., Пак С. К., Райххарт Дж. М., Берд П.И. (август 2010 г.). «Серпины напрягают мышцы: I. Ограничивают протеолиз в различных биологических системах». Журнал биологической химии. 285 (32): 24299–305. Дои:10.1074 / jbc.R110.112771. ЧВК 2915665. PMID 20498369.
- ^ Whisstock JC, Silverman GA, Bird PI, Bottomley SP, Kaiserman D, Luke CJ, Pak SC, Reichhart JM, Huntington JA (август 2010 г.). «Серпины напрягают мышцы: II. Структурные сведения о распознавании, полимеризации и транспортных функциях целевой пептидазы». Журнал биологической химии. 285 (32): 24307–12. Дои:10.1074 / jbc.R110.141408. ЧВК 2915666. PMID 20498368.
- ^ а б c d е ж Gettins PG (декабрь 2002 г.). «Строение, механизм и функции Серпина». Химические обзоры. 102 (12): 4751–804. Дои:10.1021 / cr010170. PMID 12475206.
- ^ а б c d е ж грамм час я Whisstock JC, Bottomley SP (декабрь 2006 г.). «Молекулярная гимнастика: серпиновая структура, складывание и неправильная складка». Текущее мнение в структурной биологии. 16 (6): 761–8. Дои:10.1016 / j.sbi.2006.10.005. PMID 17079131.
- ^ а б c d е ж Ло Р. Х., Чжан К., Макгоуэн С., Пряжка А. М., Сильверман Г. А., Вонг В., Росадо С. Дж., Лангендорф К. Г., Пайк Р. Н., Берд П. И., Виссток Дж. К. (2006). «Обзор надсемейства серпинов». Геномная биология. 7 (5): 216. Дои:10.1186 / gb-2006-7-5-216. ЧВК 1779521. PMID 16737556.
- ^ а б c d е Stein PE, Carrell RW (февраль 1995 г.). «Что дисфункциональные серпины говорят нам о молекулярной подвижности и болезни?». Структурная биология природы. 2 (2): 96–113. Дои:10.1038 / nsb0295-96. PMID 7749926. S2CID 21223825.
- ^ а б c d е ж грамм час я j Янчаускене С.М., Балс Р., Кочулла Р., Фогельмайер С., Кёнляйн Т., Велте Т. (август 2011 г.). «Открытие α1-антитрипсина и его роль в здоровье и болезнях». Респираторная медицина. 105 (8): 1129–39. Дои:10.1016 / j.rmed.2011.02.002. PMID 21367592.
- ^ а б c d Каррелл Р.В., Ломас Д.А. (июль 1997 г.). «Конформационное заболевание». Ланцет. 350 (9071): 134–8. Дои:10.1016 / S0140-6736 (97) 02073-4. PMID 9228977. S2CID 39124185.
- ^ Ферми С., Персоны Л. (1984). "Фермент Untersuchungen uber die, Vergleichende Studie" [Исследования фермента, Сравнительное исследование]. Z Hyg Infektionskr (на немецком языке) (18): 83–89.
- ^ Шульц Х, Гульден I, Хайде К., Шененбергер М, Швик Г (1955). "Zur Kenntnis der alpha-globulin des menschlichen normal serum" [Для ознакомления с альфа-глобулином нормальных сывороток человека]. Zeitschrift für Naturforschung B (на немецком). 10 (8): 463. Дои:10.1515 / znb-1955-0810. S2CID 95960716.
- ^ Лорелл CB, Эрикссон S (2013). «Электрофоретический профиль α1-глобулина в сыворотке крови при дефиците α1-антитрипсина. 1963». ХОБЛ. 10 Дополнение 1: 3–8. Дои:10.3109/15412555.2013.771956. PMID 23527532. S2CID 36366089.
- ^ а б de Serres FJ (1 ноября 2002 г.). «Расовое и этническое распространение дефицита альфа-антитрипсина во всем мире». Грудь. 122 (5): 1818–1829. Дои:10.1378 / сундук.122.5.1818. PMID 12426287.
- ^ Эгеберг О. (июнь 1965 г.). «Унаследованный дефицит антитромбина, вызывающий тромбофилию». Тромбоз и геморрагический диатез. 13 (2): 516–30. Дои:10.1055 / с-0038-1656297. PMID 14347873.
- ^ а б Патнаик М.М., Молл С. (ноябрь 2008 г.). «Унаследованный дефицит антитромбина: обзор». Гемофилия. 14 (6): 1229–39. Дои:10.1111 / j.1365-2516.2008.01830.x. PMID 19141163. S2CID 20768425.
- ^ а б Хант, LT, Дейхофф, Миссури (июль 1980 г.). «Удивительное новое суперсемейство белков, содержащее овальбумин, антитромбин-III и ингибитор альфа-1-протеиназы». Сообщения о биохимических и биофизических исследованиях. 95 (2): 864–71. Дои:10.1016 / 0006-291X (80) 90867-0. PMID 6968211.
- ^ а б Лёберманн Х., Токуока Р., Дайзенхофер Дж., Хубер Р. (август 1984 г.). «Ингибитор альфа-1-протеиназы человека. Анализ кристаллической структуры двух модификаций кристаллов, молекулярная модель и предварительный анализ последствий для функции». Журнал молекулярной биологии. 177 (3): 531–57. Дои:10.1016/0022-2836(84)90298-5. PMID 6332197.
- ^ а б c Stein PE, Leslie AG, Finch JT, Turnell WG, McLaughlin PJ, Carrell RW (сентябрь 1990 г.). «Кристаллическая структура овальбумина как модель реактивного центра серпинов». Природа. 347 (6288): 99–102. Bibcode:1990Натура.347 ... 99С. Дои:10.1038 / 347099a0. PMID 2395463. S2CID 4342263.
- ^ а б c Ирвинг Дж. А., Пайк Р. Н., Леск А. М., Виссток Дж. К. (декабрь 2000 г.). «Филогения суперсемейства серпинов: последствия закономерностей сохранения аминокислот для структуры и функции». Геномные исследования. 10 (12): 1845–64. Дои:10.1101 / гр. GR-1478R. PMID 11116082.
- ^ а б Ирвинг Дж. А., Стинбаккерс П. Дж., Леск А. М., Оп ден Кэмп Х. Дж., Пайк Р. Н., Виссток Дж. К. (ноябрь 2002 г.). «Серпины у прокариот». Молекулярная биология и эволюция. 19 (11): 1881–90. Дои:10.1093 / oxfordjournals.molbev.a004012. PMID 12411597.
- ^ а б Стинбаккерс П.Дж., Ирвинг Дж.А., Харханги Х.Р., Свинкелс В.Дж., Ахманова А., Дейкерман Р., Джеттен М.С., ван дер Дрифт С., Виссток Дж.С., Оп ден Кэмп HJ (август 2008 г.) «Серпин в целлюлосоме анаэробного гриба Piromyces sp. Штамм E2». Микологические исследования. 112 (Pt 8): 999–1006. Дои:10.1016 / j.mycres.2008.01.021. PMID 18539447.
- ^ а б Роулингс Н.Д., Толле Д.П., Барретт А.Дж. (март 2004 г.). «Эволюционные семейства ингибиторов пептидазы». Биохимический журнал. 378 (Pt 3): 705–16. Дои:10.1042 / BJ20031825. ЧВК 1224039. PMID 14705960.
- ^ Барретт А.Дж., Роулингс Н.Д. (апрель 1995 г.). «Семейства и кланы сериновых пептидаз». Архивы биохимии и биофизики. 318 (2): 247–50. Дои:10.1006 / abbi.1995.1227. PMID 7733651.
- ^ а б Хантингтон Дж. А., Рид Р. Дж., Каррелл Р. В. (октябрь 2000 г.). «Структура комплекса серпин-протеаза ингибируется деформацией». Природа. 407 (6806): 923–6. Bibcode:2000Натурал.407..923H. Дои:10.1038/35038119. PMID 11057674. S2CID 205009937.
- ^ Барретт А.Дж., Роулингс Н.Д. (май 2001 г.). «Эволюционные линии цистеинпептидаз». Биологическая химия. 382 (5): 727–33. Дои:10.1515 / BC.2001.088. PMID 11517925. S2CID 37306786.
- ^ Ирвинг Дж. А., Пайк Р. Н., Дай В., Бремме Д., Уорролл Д. М., Сильверман Г. А., Кутцер Т. Х., Деннисон С., Боттомли С. П., Виссток Дж. К. (апрель 2002 г.). «Доказательства того, что архитектура серпина внутренне поддерживает ингибирование папаин-подобной цистеиновой протеазы: разработка альфа (1) -антитрипсина для ингибирования протеаз катепсина». Биохимия. 41 (15): 4998–5004. Дои:10.1021 / bi0159985. PMID 11939796.
- ^ а б Шик С., Бремме Д., Бартуски А. Дж., Уэмура Ю., Шехтер Н. М., Сильверман Г. А. (ноябрь 1998 г.). «Петля реактивного сайта серпина SCCA1 необходима для ингибирования цистеиновой протеиназы». Труды Национальной академии наук Соединенных Штатов Америки. 95 (23): 13465–70. Bibcode:1998PNAS ... 9513465S. Дои:10.1073 / пнас.95.23.13465. ЧВК 24842. PMID 9811823.
- ^ а б Макгоуэн С., Бакл А.М., Ирвинг Дж. А., Онг П. К., Баштанник-Пухалович Т. А., Кан В. Т., Хендерсон К. Н., Булинко Ю. А., Попова Е. Ю., Смит А. И., Боттомли С. П., Россджон Дж., Григорьев С. А., Пайк Р. Н., Уиссток Дж. . «Рентгеновская кристаллическая структура MENT: свидетельство наличия функциональных петлевых полимеров при конденсации хроматина». Журнал EMBO. 25 (13): 3144–55. Дои:10.1038 / sj.emboj.7601201. ЧВК 1500978. PMID 16810322.
- ^ Онг П.С., Макгоуэн С., Пирс М.К., Ирвинг Дж. А., Кан В. Т., Григорьев С. А., Терк Б., Сильверман Г. А., Брикс К., Боттомли С. П., Уиссток Дж. К., Пайк Р. Н. (декабрь 2007 г.). «ДНК ускоряет ингибирование катепсина V человека серпинами». Журнал биологической химии. 282 (51): 36980–6. Дои:10.1074 / jbc.M706991200. PMID 17923478.
- ^ а б Акоста Х, Илиев Д., Гран Т.Х., Гуиньяр Н., Маккарана М., Грисбах Дж., Херцманн С., Сага М., Климент М., Пера Э.М. (март 2015 г.). «Серпин PN1 является регулятором обратной связи передачи сигналов FGF в зародышевом листе и формировании первичной оси». Разработка. 142 (6): 1146–58. Дои:10.1242 / dev.113886. PMID 25758225.
- ^ а б c Хашимото С., Ким Д.Р., Вайс Л.А., Миллер Д.В., Морисато Д. (декабрь 2003 г.). «Пространственная регуляция онтогенетической сигнализации серпином». Клетка развития. 5 (6): 945–50. Дои:10.1016 / S1534-5807 (03) 00338-1. PMID 14667416.
- ^ Птица П.И. (февраль 1999 г.). «Регулирование проапоптотических сериновых протеиназ гранул лейкоцитов с помощью внутриклеточных серпинов». Иммунология и клеточная биология. 77 (1): 47–57. Дои:10.1046 / j.1440-1711.1999.00787.x. PMID 10101686. S2CID 44268106.
- ^ Bird CH, Sutton VR, Sun J, Hirst CE, Novak A, Kumar S, Trapani JA, Bird PI (ноябрь 1998 г.). «Селективная регуляция апоптоза: цитотоксический ингибитор серпиновой протеиназы лимфоцитов 9 защищает от апоптоза, опосредованного гранзимом B, не нарушая путь гибели клеток Fas». Молекулярная и клеточная биология. 18 (11): 6387–98. Дои:10.1128 / mcb.18.11.6387. ЧВК 109224. PMID 9774654.
- ^ Ray CA, Black RA, Kronheim SR, Greenstreet TA, Sleath PR, Salvesen GS, Pickup DJ (май 1992). «Вирусное подавление воспаления: вирус коровьей оспы кодирует ингибитор фермента, преобразующего бета интерлейкин-1». Клетка. 69 (4): 597–604. Дои:10.1016 / 0092-8674 (92) 90223-У. PMID 1339309. S2CID 7398844.
- ^ а б Веркаммен Д., Беленги Б., ван де Котт Б., Буненс Т., Гавиган Дж. А., Де Рик Р., Бракениер А., Инзе Д., Харрис Д. Л., Ван Брейзегем Ф (декабрь 2006 г.). «Серпин1 Arabidopsis thaliana является суицидным ингибитором метакаспазы 9». Журнал молекулярной биологии. 364 (4): 625–36. Дои:10.1016 / j.jmb.2006.09.010. PMID 17028019.
- ^ а б Лампл Н., Будай-Адриан О., Давыдов О., Джосс ТВ, Харроп С.Дж., Курми П.М., Робертс Т.Х., Флур Р. (апрель 2010 г.). «Arabidopsis AtSerpin1, кристаллическая структура и взаимодействие in vivo с его протеазой-мишенью, реагирующей на высыхание (RD21)». Журнал биологической химии. 285 (18): 13550–60. Дои:10.1074 / jbc.M109.095075. ЧВК 2859516. PMID 20181955.
- ^ а б c Клибер М.А., Андерхилл С., Хаммонд Г.Л., Мюллер Ю.А. (октябрь 2007 г.). «Кортикостероид-связывающий глобулин, структурная основа для транспорта стероидов и высвобождения, запускаемого протеиназой». Журнал биологической химии. 282 (40): 29594–603. Дои:10.1074 / jbc.M705014200. PMID 17644521.
- ^ а б c d е Чжоу А., Вэй З., Рид Р.Дж., Каррелл Р.В. (сентябрь 2006 г.). «Структурный механизм переноса и выброса тироксина в кровь». Труды Национальной академии наук Соединенных Штатов Америки. 103 (36): 13321–6. Bibcode:2006PNAS..10313321Z. Дои:10.1073 / pnas.0604080103. ЧВК 1557382. PMID 16938877.
- ^ Хантингтон Дж. А., Штейн П. Е. (май 2001 г.). «Состав и свойства яичного альбумина». Журнал хроматографии B. 756 (1–2): 189–98. Дои:10.1016 / S0378-4347 (01) 00108-6. PMID 11419711.
- ^ а б c Мала Дж. Дж., Роуз С. (ноябрь 2010 г.). «Взаимодействие белка теплового шока 47 с коллагеном и реакция на стресс: нетрадиционная модель шаперона?». Науки о жизни. 87 (19–22): 579–86. Дои:10.1016 / j.lfs.2010.09.024. PMID 20888348.
- ^ Григорьев С.А., Беднар Дж., Вальдшнеп К.Л. (февраль 1999 г.). «MENT, гетерохроматиновый белок, который обеспечивает сворачивание хроматина более высокого порядка, является новым членом семейства серпинов». Журнал биологической химии. 274 (9): 5626–36. Дои:10.1074 / jbc.274.9.5626. PMID 10026180.
- ^ Elliott PR, Lomas DA, Carrell RW, Abrahams JP (август 1996 г.). «Тормозная конформация реактивной петли альфа-1-антитрипсина». Структурная биология природы. 3 (8): 676–81. Дои:10.1038 / nsb0896-676. PMID 8756325. S2CID 22976014.
- ^ Хорват А. Дж., Ирвинг Дж. А., Россджон Дж., Ло Р. Х., Боттомли С. П., Куинси Н. С., Пайк Р. Н., Кафлин П. Б., Уиссток Дж. С. (декабрь 2005 г.). "Мышиный ортолог человеческого антихимотрипсина: структурная парадигма серпинов клады A3". Журнал биологической химии. 280 (52): 43168–78. Дои:10.1074 / jbc.M505598200. PMID 16141197.
- ^ Whisstock JC, Скиннер R, Каррелл RW, Леск AM (февраль 2000 г.). «Конформационные изменения в серпинах: I. Нативные и расщепленные конформации альфа (1) -антитрипсина». Журнал молекулярной биологии. 296 (2): 685–99. Дои:10.1006 / jmbi.1999.3520. PMID 10669617.
- ^ а б Хантингтон Дж. А. (август 2006 г.). «Змеевики-фигурки - преимущества подвижного механизма». Тенденции в биохимических науках. 31 (8): 427–35. Дои:10.1016 / j.tibs.2006.06.005. PMID 16820297.
- ^ Джин Л., Абрахамс Дж. П., Скиннер Р., Петиту М., Пайк Р. Н., Каррелл Р. В. (декабрь 1997 г.). «Антикоагулянтная активация антитромбина гепарином». Труды Национальной академии наук Соединенных Штатов Америки. 94 (26): 14683–8. Bibcode:1997PNAS ... 9414683J. Дои:10.1073 / pnas.94.26.14683. ЧВК 25092. PMID 9405673.
- ^ Whisstock JC, Pike RN, Jin L, Skinner R, Pei XY, Carrell RW, Lesk AM (сентябрь 2000 г.). «Конформационные изменения серпинов: II. Механизм активации антитромбина гепарином». Журнал молекулярной биологии. 301 (5): 1287–305. Дои:10.1006 / jmbi.2000.3982. PMID 10966821.
- ^ Ли В., Джонсон Д. Д., Эсмон СТ, Хантингтон Дж. А. (сентябрь 2004 г.). «Структура тройного комплекса антитромбин-тромбин-гепарин раскрывает антитромботический механизм гепарина». Структурная и молекулярная биология природы. 11 (9): 857–62. Дои:10.1038 / nsmb811. PMID 15311269. S2CID 28790576.
- ^ Джонсон Д. Д., Ли В., Адамс Т. Е., Хантингтон Д. А. (май 2006 г.). «Структура фактора Ха-гепарина антитромбин-S195A раскрывает аллостерический механизм активации антитромбина». Журнал EMBO. 25 (9): 2029–37. Дои:10.1038 / sj.emboj.7601089. ЧВК 1456925. PMID 16619025.
- ^ Валенга Дж. М., Джеске В. П., Самама М. М., Frapaise FX, Бик Р. Л., Фарид Дж. (Март 2002 г.). «Фондапаринукс: синтетический пентасахарид гепарина как новое антитромботическое средство». Заключение эксперта по исследуемым препаратам. 11 (3): 397–407. Дои:10.1517/13543784.11.3.397. PMID 11866668. S2CID 24796086.
- ^ Petitou M, van Boeckel CA (июнь 2004 г.). «Синтетический пентасахарид, связывающий антитромбин III, теперь является лекарством! Что будет дальше?». Angewandte Chemie. 43 (24): 3118–33. Дои:10.1002 / anie.200300640. PMID 15199558.
- ^ а б Линдаль Т.Л., Сигурдардоттир О., Виман Б. (сентябрь 1989 г.). «Стабильность ингибитора активатора плазминогена 1 (PAI-1)». Тромбоз и гемостаз. 62 (2): 748–51. Дои:10.1055 / с-0038-1646895. PMID 2479113.
- ^ Мушунже А., Эванс Дж., Бреннан С.О., Каррелл Р.В., Чжоу А. (декабрь 2004 г.). «Скрытый антитромбин и его обнаружение, образование и оборот в кровообращении». Журнал тромбоза и гемостаза. 2 (12): 2170–7. Дои:10.1111 / j.1538-7836.2004.01047.x. PMID 15613023. S2CID 43029244.
- ^ Чжан К., Бакл А.М., Ло Р.Х., Пирс М.К., Кабрита Л.Д., Ллойд Г.Дж., Ирвинг Дж. А., Смит А.И., Рузила К., Россджон Дж., Боттомли С.П., Уиссток Дж. «N-конец серпина, tengpin, функционирует для улавливания метастабильного нативного состояния». Отчеты EMBO. 8 (7): 658–63. Дои:10.1038 / sj.embor.7400986. ЧВК 1905895. PMID 17557112.
- ^ Чжан К., Ло Р. Х., Боттомли С. П., Уиссток Дж. К., Пряжка А. М. (март 2008 г.). «Структурная основа полимеризации петлевых С-листов в серпинах». Журнал молекулярной биологии. 376 (5): 1348–59. Дои:10.1016 / j.jmb.2007.12.050. PMID 18234218.
- ^ Пембертон PA, Stein PE, Pepys MB, Potter JM, Carrell RW (ноябрь 1988 г.). «Гормон-связывающие глобулины претерпевают серпиновые конформационные изменения при воспалении». Природа. 336 (6196): 257–8. Bibcode:1988Натура.336..257П. Дои:10.1038 / 336257a0. PMID 3143075. S2CID 4326356.
- ^ а б c Цао С., Лоуренс Д.А., Ли Й., фон Арним К.А., Херц Дж., Су Е.Дж., Макарова А., Хайман Б.Т., Стрикленд Д.К., Чжан Л. (май 2006 г.). «Эндоцитарный рецептор LRP вместе с tPA и PAI-1 координирует Mac-1-зависимую миграцию макрофагов». Журнал EMBO. 25 (9): 1860–70. Дои:10.1038 / sj.emboj.7601082. ЧВК 1456942. PMID 16601674.
- ^ Дженсен Дж. К., Долмер К., Геттинс П. Г. (июль 2009 г.). «Специфичность связывания белка, связанного с рецептором липопротеинов низкой плотности, с различными конформационными состояниями ингибитора-1 активатора плазминогена серпинов клады E и нексина-1 протеиназы». Журнал биологической химии. 284 (27): 17989–97. Дои:10.1074 / jbc.M109.009530. ЧВК 2709341. PMID 19439404.
- ^ Соукуп С.Ф., Кули Дж., Губб Д. (июнь 2009 г.). Рулифсон Э (ред.). «Поглощение некротического серпина у Drosophila melanogaster через рецептор липофорина-1». PLOS Genetics. 5 (6): e1000532. Дои:10.1371 / journal.pgen.1000532. ЧВК 2694266. PMID 19557185.
- ^ Kaiserman D, Whisstock JC, Bird PI (1 января 2006 г.). «Механизмы дисфункции серпина при болезни». Обзоры экспертов в области молекулярной медицины. 8 (31): 1–19. Дои:10.1017 / S1462399406000184. PMID 17156576.
- ^ Hopkins PC, Carrell RW, Stone SR (август 1993 г.). «Эффекты мутаций в шарнирной области серпинов». Биохимия. 32 (30): 7650–7. Дои:10.1021 / bi00081a008. PMID 8347575.
- ^ Beauchamp NJ, Pike RN, Daly M, Butler L, Makris M, Dafforn TR, Zhou A, Fitton HL, Preston FE, Peake IR, Carrell RW (октябрь 1998 г.). «Antithrombins Wibble and Wobble (T85M / K): архетипические конформационные заболевания с латентным переходом in vivo, тромбозом и активацией гепарина». Кровь. 92 (8): 2696–706. Дои:10.1182 / blood.V92.8.2696. PMID 9763552.
- ^ а б c Гупту Б., Хейз Б., Чанг В.С., Даффорн Т.Р., Каррелл Р.В., Рид Р.Дж., Ломас Д.А. (январь 2000 г.). «Неактивная конформация серпин-альфа (1) -антихимотрипсина указывает на двухэтапное введение реактивной петли: последствия для ингибирующей функции и конформационного заболевания». Труды Национальной академии наук Соединенных Штатов Америки. 97 (1): 67–72. Bibcode:2000PNAS ... 97 ... 67G. Дои:10.1073 / pnas.97.1.67. ЧВК 26617. PMID 10618372.
- ^ а б Homan EP, Rauch F, Grafe I, Lietman C, Doll JA, Dawson B, Bertin T, Napierala D, Morello R, Gibbs R, White L, Miki R, Cohn DH, Crawford S, Travers R, Glorieux FH, Lee B (Декабрь 2011 г.). «Мутации в SERPINF1 вызывают несовершенный остеогенез типа VI». Журнал исследований костей и минералов. 26 (12): 2798–803. Дои:10.1002 / jbmr.487. ЧВК 3214246. PMID 21826736.
- ^ Фэй В.П., Паркер А.С., Кондри Л.Р., Шапиро А.Д. (июль 1997 г.). «Дефицит ингибитора-1 активатора плазминогена человека (PAI-1): характеристика большого родственника с нулевой мутацией в гене PAI-1». Кровь. 90 (1): 204–8. Дои:10.1182 / кровь.V90.1.204. PMID 9207454.
- ^ а б c d е ж Хейт С., Джексон BC, МакЭндрюс М., Райт М. В., Томпсон, округ Колумбия, Сильверман Г. А., Неберт Д. В., Василиу В. (30 октября 2013 г.). «Обновление суперсемейства гена SERPIN человека и мыши». Геномика человека. 7: 22. Дои:10.1186/1479-7364-7-22. ЧВК 3880077. PMID 24172014.
- ^ Оуэн М.С., Бреннан С.О., Льюис Дж. Х., Каррелл Р. У. (сентябрь 1983 г.). «Мутация антитрипсина в антитромбин. Альфа-1-антитрипсин Питтсбург (358 Met приводит к Arg), фатальное нарушение свертываемости крови». Медицинский журнал Новой Англии. 309 (12): 694–8. Дои:10.1056 / NEJM198309223091203. PMID 6604220.
- ^ а б Ломас Д.А., Эванс Д.Л., Финч Д.Т., Каррелл Р.В. (июнь 1992 г.). «Механизм накопления Z альфа 1-антитрипсина в печени». Природа. 357 (6379): 605–7. Bibcode:1992Натура.357..605Л. Дои:10.1038 / 357605a0. PMID 1608473. S2CID 4359543.
- ^ Крегер Х., Миранда Э., Маклауд И., Перес Дж., Кроутер, округ Колумбия, Марчиньяк С.Дж., Ломас Д.А. (август 2009 г.). «Деградация, связанная с эндоплазматическим ретикулумом (ERAD), и аутофагия взаимодействуют для разрушения полимерогенных мутантных серпинов». Журнал биологической химии. 284 (34): 22793–802. Дои:10.1074 / jbc.M109.027102. ЧВК 2755687. PMID 19549782.
- ^ а б c Ямасаки М., Ли В., Джонсон Д. Д., Хантингтон Дж. А. (октябрь 2008 г.). «Кристаллическая структура стабильного димера раскрывает молекулярную основу полимеризации серпина». Природа. 455 (7217): 1255–8. Bibcode:2008 Натур.455.1255Y. Дои:10.1038 / природа07394. PMID 18923394. S2CID 205215121.
- ^ а б Боттомли СП (октябрь 2011 г.). «Структурное разнообразие неправильного фолдинга α1-антитрипсина». EMBO отчеты. 12 (10): 983–4. Дои:10.1038 / embor.2011.187. ЧВК 3185355. PMID 21921939.
- ^ а б Ямасаки М., Сендалл Т.Дж., Пирс М.К., Уиссток Дж. К., Хантингтон Дж. А. (октябрь 2011 г.). «Молекулярная основа дефицита α1-антитрипсина, выявленная структурой тримеров с заменой домена». EMBO отчеты. 12 (10): 1011–7. Дои:10.1038 / embor.2011.171. ЧВК 3185345. PMID 21909074.
- ^ Чанг В.С., Виссток Дж., Хопкинс П.С., Леск А.М., Каррелл Р.В., Уорделл М.Р. (январь 1997 г.). «Важность высвобождения цепи 1C для механизма полимеризации ингибирующих серпинов». Белковая наука. 6 (1): 89–98. Дои:10.1002 / pro.5560060110. ЧВК 2143506. PMID 9007980.
- ^ Миранда Е., Перес Дж., Экеова У.И., Хаджич Н., Калшекер Н., Гупту Б., Портманн Б., Белорги Д., Хилл М., Чемберс С., Текман Дж., Александр Дж. Дж., Марчиньяк С.Дж., Ломас Д.А. (сентябрь 2010 г.). «Новое моноклональное антитело для характеристики патогенных полимеров при заболеваниях печени, связанных с дефицитом альфа1-антитрипсина». Гепатология. 52 (3): 1078–88. Дои:10.1002 / hep.23760. PMID 20583215. S2CID 8188156.
- ^ Sandhaus RA (октябрь 2004 г.). «Дефицит альфа1-антитрипсина. 6: новые и появляющиеся методы лечения дефицита альфа1-антитрипсина». Грудная клетка. 59 (10): 904–9. Дои:10.1136 / thx.2003.006551. ЧВК 1746849. PMID 15454659.
- ^ Льюис ЕС (2012). «Расширение клинических показаний к терапии α (1) -антитрипсином». Молекулярная медицина. 18 (6): 957–70. Дои:10.2119 / молмед.2011.00196. ЧВК 3459478. PMID 22634722.
- ^ Фрегонский Л., Столк Дж. (2008). «Наследственный дефицит альфа-1-антитрипсина и его клинические последствия». Журнал редких заболеваний Orphanet. 3: 16. Дои:10.1186/1750-1172-3-16. ЧВК 2441617. PMID 18565211.
- ^ Юса К., Рашид С.Т., Стрик-Маршан Х., Варела I, Лю П.К., Пашон Д.Е., Миранда Э., Ордоньес А., Ханнан Н.Р., Рухани Ф.Дж., Дарче С., Александр Дж., Марчиньяк С.Дж., Фусаки Н., Хасегава М., Холмс М.С., Ди Санто Дж. П., Ломас Д. А., Брэдли А., Валлиер Л. (октябрь 2011 г.). «Направленная генная коррекция дефицита α1-антитрипсина в индуцированных плюрипотентных стволовых клетках». Природа. 478 (7369): 391–4. Bibcode:2011Натура.478..391л. Дои:10.1038 / природа10424. ЧВК 3198846. PMID 21993621.
- ^ Малля М., Филлипс Р.Л., Салдана С.А., Гупту Б., Браун С.К., Термин Д.Д., Ширвани А.М., Ву Й., Сиферс Р.Н., Абагян Р., Ломас Д.А. (ноябрь 2007 г.). «Маленькие молекулы блокируют полимеризацию Z альфа1-антитрипсина и увеличивают клиренс внутриклеточных агрегатов». Журнал медицинской химии. 50 (22): 5357–63. Дои:10.1021 / jm070687z. ЧВК 2631427. PMID 17918823.
- ^ Госай С.Дж., Квак Дж.Х., Люк С.Дж., Лонг О.С., Кинг Д.Е., Ковач К.Дж., Джонстон П.А., Шун Т.Ю., Лазо Д.С., Перлмуттер Д.Х., Сильверман Г.А., Пак СК (2010). «Автоматизированный скрининг препаратов для животных с высоким содержанием с использованием C. elegans, экспрессирующих склонный к агрегации серпин α1-антитрипсин Z». PLOS ONE. 5 (11): e15460. Bibcode:2010PLoSO ... 515460G. Дои:10.1371 / journal.pone.0015460. ЧВК 2980495. PMID 21103396.
- ^ Кабрита Л.Д., Ирвинг Дж. А., Пирс М. С., Виссток Дж. С., Боттомли С. П. (сентябрь 2007 г.). «Аэропин из экстремофила Pyrobaculum aerophilum обходит ловушку неправильного свертывания серпина». Журнал биологической химии. 282 (37): 26802–9. Дои:10.1074 / jbc.M705020200. PMID 17635906.
- ^ Fluhr R, Lampl N, Roberts TH (май 2012 г.). «Ингибиторы серпиновых протеаз в биологии растений». Physiologia Plantarum. 145 (1): 95–102. Дои:10.1111 / j.1399-3054.2011.01540.x. PMID 22085334.
- ^ Столлер JK, Aboussouan LS (2005). «Дефицит альфа1-антитрипсина» (PDF). Ланцет. 365 (9478): 2225–36. Дои:10.1016 / S0140-6736 (05) 66781-5. PMID 15978931. S2CID 54415934.
- ^ Münch J, Ständker L, Adermann K, Schulz A, Schindler M, Chinnadurai R, Pöhlmann S, Chaipan C, Biet T, Peters T., Meyer B, Wilhelm D, Lu H, Jing W, Jiang S, Forssmann WG, Kirchhoff F (Апрель 2007 г.). «Открытие и оптимизация природного ингибитора проникновения ВИЧ-1, нацеленного на слитый пептид gp41». Клетка. 129 (2): 263–75. Дои:10.1016 / j.cell.2007.02.042. PMID 17448989.
- ^ Гупту Б., Диккенс Дж. А., Ломас Д. А. (февраль 2014 г.). «Молекулярная и клеточная патология дефицита α₁-антитрипсина». Тенденции в молекулярной медицине. 20 (2): 116–27. Дои:10.1016 / j.molmed.2013.10.007. PMID 24374162.
- ^ Сейшас С., Суриано Дж., Карвалью Ф., Серука Р., Роча Дж., Ди Риенцо А. (февраль 2007 г.). «Разнообразие последовательностей в проксимальном подкластере 14q32.1 SERPIN: свидетельство естественного отбора в пользу псевдогенизации SERPINA2». Молекулярная биология и эволюция. 24 (2): 587–98. Дои:10.1093 / molbev / msl187. PMID 17135331.
- ^ Калшекер Н.А. (сентябрь 1996 г.). «Альфа-1-антихимотрипсин». Международный журнал биохимии и клеточной биологии. 28 (9): 961–4. Дои:10.1016/1357-2725(96)00032-5. PMID 8930118.
- ^ Сантамария М., Пардо-Саганта А., Альварес-Асиаин Л., Ди Скала М., Цянь С., Прието Дж., Авила М.А. (апрель 2013 г.). «Ядерный α1-антихимотрипсин способствует конденсации хроматина и подавляет пролиферацию клеток гепатоцеллюлярной карциномы человека». Гастроэнтерология. 144 (4): 818–828.e4. Дои:10.1053 / j.gastro.2012.12.029. PMID 23295442.
- ^ Zhang S, Janciauskiene S (апрель 2002 г.). «Многофункциональная способность белков: альфа1-антихимотрипсин и взаимосвязь с болезнью Альцгеймера». Журнал болезни Альцгеймера. 4 (2): 115–22. Дои:10.3233 / JAD-2002-4206. PMID 12214135.
- ^ Чао Дж., Сталлоне Дж. Н., Лян Ю. М., Чен Л. М., Ван Д. З., Чао Л. (июль 1997 г.). «Каллистатин - новый мощный вазодилататор». Журнал клинических исследований. 100 (1): 11–7. Дои:10.1172 / JCI119502. ЧВК 508159. PMID 9202051.
- ^ Мяо Р.К., Агата Дж., Чао Л., Чао Дж. (Ноябрь 2002 г.). «Каллистатин - новый ингибитор ангиогенеза и роста опухолей». Кровь. 100 (9): 3245–52. Дои:10.1182 / кровь-2002-01-0185. PMID 12384424.
- ^ Лю Ю., Бледсо Дж., Хагивара М., Шен Б., Чао Л., Чао Дж. (Октябрь 2012 г.). «Истощение эндогенного каллистатина усугубляет почечный и сердечно-сосудистый окислительный стресс, воспаление и ремоделирование органов». Американский журнал физиологии. Почечная физиология. 303 (8): F1230–8. Дои:10.1152 / ajprenal.00257.2012. ЧВК 3469672. PMID 22811485.
- ^ Гейгер М (март 2007 г.). «Ингибитор протеина С, серпин с функциями внутри и вне сосудистой биологии». Тромбоз и гемостаз. 97 (3): 343–7. Дои:10.1160 / th06-09-0488. PMID 17334499.
- ^ Баумгертнер П., Гейгер М., Цизенисс С., Маллейер Дж., Хантингтон Дж. А., Хохрайнер К., Билек Э., Штокельхубер М., Лаубер К., Шерфельд Д., Швилле П., Вельделе К., Бейер К., Энгельманн Б. (ноябрь 2007 г.). «Фосфатидилэтаноламин критически поддерживает интернализацию ингибитора протеина С, проникающего в клетки». Журнал клеточной биологии. 179 (4): 793–804. Дои:10.1083 / jcb.200707165. ЧВК 2080921. PMID 18025309.
- ^ Uhrin P, Dewerchin M, Hilpert M, Chrenek P, Schöfer C, Zechmeister-Machhart M, Krönke G, Vales A, Carmeliet P, Binder BR, Geiger M (декабрь 2000 г.). «Нарушение гена ингибитора протеина С приводит к нарушению сперматогенеза и мужскому бесплодию». Журнал клинических исследований. 106 (12): 1531–9. Дои:10.1172 / JCI10768. ЧВК 381472. PMID 11120760.
- ^ Хан М.Х., Хван С.И., Рой Д.Б., Лундгрен Д.Х., Прайс СП, Усман С.С., Фернальд Г.Х., Герлитц Б., Робинсон У.Х., Баранзини С.Е., Гриннелл Б.В., Рейн С.С., Собел Р.А., Хан Д.К., Штайнман Л. (февраль 2008 г.). «Протеомный анализ активных очагов рассеянного склероза позволяет выявить терапевтические цели». Природа. 451 (7182): 1076–81. Bibcode:2008 Натур.451.1076H. Дои:10.1038 / природа06559. PMID 18278032. S2CID 4421395.
- ^ Torpy DJ, Ho JT (август 2007 г.). «Полиморфизмы генов глобулина, связывающего кортикостероиды: клинические последствия и связи с идиопатическими расстройствами хронической усталости». Клиническая эндокринология. 67 (2): 161–7. Дои:10.1111 / j.1365-2265.2007.02890.x. PMID 17547679. S2CID 43352358.
- ^ Барталена Л., Роббинс Дж. (1992). «Вариации белков транспорта тироидных гормонов и их клинические последствия». Щитовидная железа. 2 (3): 237–45. Дои:10.1089 / th.1992.2.237. PMID 1422238.
- ^ Персани Л. (сентябрь 2012 г.). «Клинический обзор: Центральный гипотиреоз: патогенные, диагностические и терапевтические проблемы». Журнал клинической эндокринологии и метаболизма. 97 (9): 3068–78. Дои:10.1210 / jc.2012-1616. PMID 22851492.
- ^ Кумар Р., Сингх В.П., Бейкер К.М. (июль 2007 г.). «Внутриклеточная ренин-ангиотензиновая система: новая парадигма». Тенденции в эндокринологии и метаболизме. 18 (5): 208–14. Дои:10.1016 / j.tem.2007.05.001. PMID 17509892. S2CID 24041932.
- ^ Танимото К., Сугияма Ф., Гото Й., Исида Дж., Такимото Э., Ягами К., Фукамизу А., Мураками К. (декабрь 1994 г.). «Мыши с дефицитом ангиотензиногена и гипотонией». Журнал биологической химии. 269 (50): 31334–7. PMID 7989296.
- ^ Jeunemaitre X, Gimenez-Roqueplo AP, Célérier J, Corvol P (1999). «Варианты ангиотензиногена и гипертония человека». Текущие отчеты о гипертонии. 1 (1): 31–41. Дои:10.1007 / s11906-999-0071-0. PMID 10981040. S2CID 42614761.
- ^ Сетхи А.А., Нордестгаард Б.Г., Тибьярг-Хансен А. (июль 2003 г.). «Полиморфизм гена ангиотензиногена, ангиотензиноген плазмы и риск гипертонии и ишемической болезни сердца: метаанализ». Артериосклероз, тромбоз и биология сосудов. 23 (7): 1269–75. Дои:10.1161 / 01.ATV.0000079007.40884.5C. PMID 12805070.
- ^ Dickson ME, компакт-диск Sigmund (июль 2006 г.). «Генетические основы гипертонии: возвращаясь к ангиотензиногену». Гипертония. 48 (1): 14–20. Дои:10.1161 / 01.HYP.0000227932.13687.60. PMID 16754793.
- ^ Фрейзер Дж. К., Джексон Д. Г., Гайярд Дж. П., Люттер М., Лю Ю. Дж., Banchereau J, Capra JD, Pascual V (октябрь 2000 г.). "Идентификация центерина: нового серпина, ограниченного В-клетками герминального центра человека". Европейский журнал иммунологии. 30 (10): 3039–48. Дои:10.1002 / 1521-4141 (200010) 30:10 <3039 :: AID-IMMU3039> 3.0.CO; 2-H. PMID 11069088.
- ^ Патерсон М.А., Хорват А.Дж., Пайк Р.Н., Кафлин П.Б. (август 2007 г.). «Молекулярная характеристика центерина, серпина клетки зародышевого центра». Биохимический журнал. 405 (3): 489–94. Дои:10.1042 / BJ20070174. ЧВК 2267310. PMID 17447896.
- ^ Патерсон М.А., Хоскинг П.С., Кафлин П.Б. (июль 2008 г.). «Экспрессия серпинцентрина определяет фенотип зародышевого центра в B-клеточных лимфомах». Американский журнал клинической патологии. 130 (1): 117–26. Дои:10.1309 / 9QKE68QU7B825A3U. PMID 18550480.
- ^ Эштон-Рикардт PG (апрель 2013 г.). «Новая роль ингибиторов сериновой протеазы в иммунитете Т-лимфоцитов и за его пределами». Письма иммунологии. 152 (1): 65–76. Дои:10.1016 / j.imlet.2013.04.004. PMID 23624075.
- ^ Хан X, Fiehler R, Broze GJ (ноябрь 2000 г.). «Характеристика протеина Z-зависимого ингибитора протеазы». Кровь. 96 (9): 3049–55. Дои:10.1182 / кровь.V96.9.3049. PMID 11049983.
- ^ Хида К., Вада Дж., Эгути Дж., Чжан Х., Баба М., Сейда А., Хашимото И., Окада Т., Ясухара А., Накацука А., Шиката К., Хораи С., Футами Дж., Ватанабэ Е., Мацуки Ю., Хирамацу Р., Акаги С. , Макино Х., Канвар Ю.С. (июль 2005 г.). «Ингибитор сериновой протеазы висцеральной жировой ткани: уникальный адипоцитокин, повышающий чувствительность к инсулину, при ожирении». Труды Национальной академии наук Соединенных Штатов Америки. 102 (30): 10610–5. Bibcode:2005ПНАС..10210610Н. Дои:10.1073 / pnas.0504703102. ЧВК 1180799. PMID 16030142.
- ^ Фэн Р., Ли И, Ван Ц., Ло Ц, Лю Л., Чуо Ф, Ли Ц., Сун Ц. (октябрь 2014 г.). «Более высокие уровни васпина у субъектов с ожирением и сахарным диабетом 2 типа: метаанализ». Исследования и клиническая практика диабета. 106 (1): 88–94. Дои:10.1016 / j.diabres.2014.07.026. PMID 25151227.
- ^ Ремольд-О'Доннелл Э, Чин Дж, Альбертс М. (июнь 1992 г.). «Последовательность и молекулярная характеристика ингибитора эластазы моноцитов / нейтрофилов человека». Труды Национальной академии наук Соединенных Штатов Америки. 89 (12): 5635–9. Bibcode:1992PNAS ... 89.5635R. Дои:10.1073 / пнас.89.12.5635. ЧВК 49347. PMID 1376927.
- ^ Бенарафа К., Прибе Г. П., Ремольд-О'Доннелл Э. (август 2007 г.). «Ингибитор сериновой протеазы нейтрофилов serpinb1 сохраняет защитные функции легких при инфекции Pseudomonas aeruginosa». Журнал экспериментальной медицины. 204 (8): 1901–9. Дои:10.1084 / jem.20070494. ЧВК 2118684. PMID 17664292.
- ^ Antalis TM, La Linn M, Donnan K, Mateo L, Gardner J, Dickinson JL, Buttigieg K, Suhrbier A (июнь 1998 г.). «Ингибитор сериновой протеиназы (серпин), ингибитор активации плазминогена типа 2 защищает от вирусных цитопатических эффектов посредством конститутивного прайминга интерферона альфа / бета». Журнал экспериментальной медицины. 187 (11): 1799–811. Дои:10.1084 / jem.187.11.1799. ЧВК 2212304. PMID 9607921.
- ^ Чжао А., Ян З., Сун Р., Гринчук В., Нетцель-Арнетт С., Энглин И. Е., Дрисбо К. Х., Нотари Л., Бол Дж. А., Мэдден КБ, Урбан Дж. Ф., Antalis TM, Ши-Донохью Т. (июнь 2013 г.). «SerpinB2 имеет решающее значение для иммунитета Th2 против кишечной нематодной инфекции». Журнал иммунологии. 190 (11): 5779–87. Дои:10.4049 / jimmunol.1200293. ЧВК 4068334. PMID 23630350.
- ^ Догерти К.М., Пирсон Дж. М., Ян А. Ю., Вестрик Р. Дж., Бейкер М. С., Гинзбург Д. (январь 1999 г.). «Ген ингибитора активатора плазминогена-2 не требуется для нормального развития или выживания мышей». Труды Национальной академии наук Соединенных Штатов Америки. 96 (2): 686–91. Bibcode:1999ПНАС ... 96..686Д. Дои:10.1073 / пнас.96.2.686. ЧВК 15197. PMID 9892694.
- ^ Такеда А., Ямамото Т., Накамура Ю., Такахаши Т., Хибино Т. (февраль 1995 г.). «Антиген плоскоклеточной карциномы является мощным ингибитором цистеиновой протеиназы катепсина L». Письма FEBS. 359 (1): 78–80. Дои:10.1016 / 0014-5793 (94) 01456-б. PMID 7851535. S2CID 35146299.
- ^ а б Турато С., Понтиссо П. (март 2015 г.). «SERPINB3 (ингибитор серпиновой пептидазы, клад B (овальбумин), член 3)». Атлас генетики и цитогенетики в онкологии и гематологии. 19 (3): 202–209. Дои:10.4267/2042/56413. ЧВК 4430857. PMID 25984243.
- ^ а б Sivaprasad U, Askew DJ, Ericksen MB, Gibson AM, Stier MT, Brandt EB, Bass SA, Daines MO, Chakir J, Stringer KF, Wert SE, Whitsett JA, Le Cras TD, Wills-Karp M, Silverman GA, Khurana Hershey ГК (январь 2011 г.). «Неизбыточная роль мышиного Serpinb3a в индукции выработки слизи при астме». Журнал аллергии и клинической иммунологии. 127 (1): 254–61, 261.e1–6. Дои:10.1016 / j.jaci.2010.10.009. ЧВК 3058372. PMID 21126757.
- ^ Шик С., Камачи Ю., Бартуски А. Дж., Каталтепе С., Шехтер Н. М., Пембертон П. А., Сильверман Г. А. (январь 1997 г.). «Антиген плоскоклеточной карциномы 2 представляет собой новый серпин, который ингибирует химотрипсиноподобные протеиназы, катепсин G и химазу тучных клеток». Журнал биологической химии. 272 (3): 1849–55. Дои:10.1074 / jbc.272.3.1849. PMID 8999871.
- ^ Теох СС, Виссток Дж. К., Берд П.И. (апрель 2010 г.). «Маспин (SERPINB5) - облигатный внутриклеточный серпин». Журнал биологической химии. 285 (14): 10862–9. Дои:10.1074 / jbc.M109.073171. ЧВК 2856292. PMID 20123984.
- ^ Zou Z, Anisowicz A, Hendrix MJ, Thor A, Neveu M, Sheng S, Rafidi K, Seftor E, Sager R (январь 1994). «Маспин, серпин с опухолевой активностью в эпителиальных клетках молочной железы человека». Наука. 263 (5146): 526–9. Bibcode:1994Наука ... 263..526Z. Дои:10.1126 / science.8290962. PMID 8290962.
- ^ а б c Теох С.С., Вьюссо Дж., Пракаш М., Беркович С., Луу Дж., Берд СН, Лоу Р.Х., Росадо С., Прайс Дж. Т., Виссток Дж. С., Берд П.И. «Маспин не требуется для эмбрионального развития или подавления опухоли». Nature Communications. 5: 3164. Bibcode:2014 НатКо ... 5,3164 т. Дои:10.1038 / ncomms4164. ЧВК 3905777. PMID 24445777.
- ^ Гао Ф., Ши Х.Й., Догти С., Селла Н., Чжан М. (апрель 2004 г.). «Маспин играет важную роль в раннем эмбриональном развитии». Разработка. 131 (7): 1479–89. Дои:10.1242 / dev.01048. PMID 14985257.
- ^ Скотт Флорида, Херст С.Е., Сан Дж., Берд С.Х., Боттомли СП, Берд П.И. (март 1999 г.) «Внутриклеточный ингибитор серпиновой протеиназы 6 экспрессируется в моноцитах и гранулоцитах и является мощным ингибитором протеазы азурофильных гранул, катепсина G». Кровь. 93 (6): 2089–97. Дои:10.1182 / blood.V93.6.2089.406k10_2089_2097. PMID 10068683.
- ^ Тан Дж, Пракаш, доктор медицины, Кайзерман Д., Берд П.И. (июль 2013 г.). «Отсутствие SERPINB6A вызывает нейросенсорную тугоухость с множественными гистопатологиями во внутреннем ухе мыши». Американский журнал патологии. 183 (1): 49–59. Дои:10.1016 / j.ajpath.2013.03.009. PMID 23669344.
- ^ Скарфф К.Л., Унг К.С., Нандуркар Х., Crack PJ, Bird CH, Bird PI (май 2004 г.). «Целенаправленное нарушение SPI3 / Serpinb6 не приводит к дефектам развития или роста, дисфункции лейкоцитов или предрасположенности к инсульту». Молекулярная и клеточная биология. 24 (9): 4075–82. Дои:10.1128 / MCB.24.9.4075-4082.2004. ЧВК 387772. PMID 15082799.
- ^ Сирмачи А., Эрбек С., Прайс Дж, Хуанг М., Думан Д., Дженгиз Ф. Б., Бадемчи Г., Токгоз-Йилмаз С., Хисми Б., Оздаг Н., Озтюрк Б., Кулаксизоглу С., Йилдирим Е., Кокотас Н., Григориаду М., Петерсен М. Шахин Х., Канаан М., Кинг М.С., Чен З.Й., Блэнтон С.Х., Лю XZ, Зухнер С., Акар Н., Текин М. (2010). «Усекающая мутация в SERPINB6 связана с аутосомно-рецессивной несиндромной сенсоневральной потерей слуха». Американский журнал генетики человека. 86 (5): 797–804. Дои:10.1016 / j.ajhg.2010.04.004. ЧВК 2869020. PMID 20451170.
- ^ Мията Т., Инаги Р., Нангаку М., Имасава Т., Сато М., Изухара И., Судзуки Д., Ёсино А., Оноги Н., Кимура М., Сугияма С., Курокава К. (март 2002 г.). «Сверхэкспрессия мегсина серпина вызывает прогрессирующую пролиферацию и экспансию мезангиальных клеток». Журнал клинических исследований. 109 (5): 585–93. Дои:10.1172 / JCI14336. ЧВК 150894. PMID 11877466.
- ^ а б Мията Т., Ли М., Ю X, Хираяма Н. (май 2007 г.). «Ген Megsin: его геномный анализ, патобиологические функции и терапевтические перспективы». Текущая геномика. 8 (3): 203–8. Дои:10.2174/138920207780833856. ЧВК 2435355. PMID 18645605.
- ^ Кубо А. (август 2014 г.). «Кератоз ладонно-подошвенного типа по типу Нагашима: распространенный азиатский тип, вызванный дефицитом ингибитора протеазы SERPINB7». Журнал следственной дерматологии. 134 (8): 2076–9. Дои:10.1038 / jid.2014.156. PMID 25029323.
- ^ Дален-младший, Жан Ф., Томас Дж., Фостер, округ Колумбия, Кисиэль В. (январь 1998 г.). «Ингибирование растворимого рекомбинантного фурина ингибитором протеиназы человека 8». Журнал биологической химии. 273 (4): 1851–4. Дои:10.1074 / jbc.273.4.1851. PMID 9442015.
- ^ Сан Дж., Бёрд Ч., Саттон В., Макдональд Л., Кафлин П. Б., Де Йонг Т. А., Трапани Дж. А., Берд П. И. (ноябрь 1996 г.). «В цитотоксических лимфоцитах присутствует цитозольный ингибитор гранзима B, относящийся к вирусному регулятору апоптоза, модификатору цитокинового ответа А». Журнал биологической химии. 271 (44): 27802–9. Дои:10.1074 / jbc.271.44.27802. PMID 8910377.
- ^ Чжан М., Пак С.М., Ван И, Шах Р., Лю Н., Мурманн А.Э., Ван С.Р., Питер М.Э., Эштон-Рикардт П.Г. (апрель 2006 г.). «Ингибитор сериновой протеазы 6 защищает цитотоксические Т-клетки от самоповреждения, обеспечивая целостность цитотоксических гранул». Иммунитет. 24 (4): 451–61. Дои:10.1016 / j.immuni.2006.02.002. PMID 16618603.
- ^ Риззителли А., Мейтер С., Вега Рамос Дж., Берд СН, Минтерн Дж.Д., Манган М.С., Вилладангос Дж., Берд П.И. (октябрь 2012 г.). «У мышей с дефицитом Serpinb9 (Spi6) нарушена перекрестная презентация антигена, опосредованная дендритными клетками». Иммунология и клеточная биология. 90 (9): 841–51. Дои:10.1038 / icb.2012.29. PMID 22801574. S2CID 39276036.
- ^ Ривальд М., Чуанг Т., Нойбауэр А., Рисс Х., Шлиф Р.Р. (февраль 1998 г.). «Экспрессия бомапина, нового серпина человека, при нормальном / злокачественном кроветворении и в линиях моноцитарных клеток THP-1 и AML-193». Кровь. 91 (4): 1256–62. Дои:10.1182 / кровь.V91.4.1256. PMID 9454755.
- ^ а б Askew DJ, Cataltepe S, Kumar V, Edwards C, Pace SM, Howarth RN, Pak SC, Askew YS, Brömme D, Luke CJ, Whisstock JC, Silverman GA (август 2007 г.). «SERPINB11 - это новый неингибиторный внутриклеточный серпин. Общие однонуклеотидные полиморфизмы в каркасе нарушают конформационные изменения». Журнал биологической химии. 282 (34): 24948–60. Дои:10.1074 / jbc.M703182200. PMID 17562709.
- ^ Финно С.Дж., Стивенс С., Янг А., Аффолтер В., Джоши Н.А., Рамзи С., Баннаш Д.Л. (апрель 2015 г.). «Вариант смещения рамки SERPINB11, связанный с новым специфическим фенотипом копыт у пони Коннемара». PLOS Genetics. 11 (4): e1005122. Дои:10.1371 / journal.pgen.1005122. ЧВК 4395385. PMID 25875171.
- ^ Askew YS, Pak SC, Luke CJ, Askew DJ, Cataltepe S, Mills DR, Kato H, Lehoczky J, Dewar K, Birren B, Silverman GA (декабрь 2001 г.). «SERPINB12 - новый член семейства овец-серпинов человека, который широко экспрессируется и ингибирует трипсиноподобные сериновые протеиназы». Журнал биологической химии. 276 (52): 49320–30. Дои:10.1074 / jbc.M108879200. PMID 11604408.
- ^ Велсс Т., Сан Дж., Ирвинг Дж. А., Блюм Р., Смит А. И., Виссток Дж. К., Пайк Р. Н., фон Микец А., Ружичка Т., Берд П. И., Abts HF (июнь 2003 г.). «Hurpin является селективным ингибитором лизосомального катепсина L и защищает кератиноциты от апоптоза, индуцированного ультрафиолетом». Биохимия. 42 (24): 7381–9. Дои:10.1021 / bi027307q. PMID 12809493.
- ^ Исигуро К., Кодзима Т., Кадомацу К., Накаяма И., Такаги А., Сузуки М., Такеда Н., Ито М., Ямамото К., Мацусита Т., Кусугами К., Мурамацу Т., Сайто Х. (октябрь 2000 г.). «Полный дефицит антитромбина у мышей приводит к гибели эмбрионов». Журнал клинических исследований. 106 (7): 873–8. Дои:10.1172 / JCI10489. ЧВК 517819. PMID 11018075.
- ^ Хантингтон Дж. А. (июль 2011 г.). «Строение, функции и дисфункция Серпина». Журнал тромбоза и гемостаза. 9 Дополнение 1: 26–34. Дои:10.1111 / j.1538-7836.2011.04360.x. PMID 21781239. S2CID 1020630.
- ^ Vicente CP, He L, Pavão MS, Tollefsen DM (декабрь 2004 г.). «Антитромботическая активность дерматансульфата у мышей с дефицитом кофактора II гепарина». Кровь. 104 (13): 3965–70. Дои:10.1182 / кровь-2004-02-0598. PMID 15315969.
- ^ Айхара К., Адзума Х, Акаике М, Икеда Й, Сата М, Такамори Н, Яги С., Ивасе Т, Сумитомо Й, Кавано Х, Ямада Т, Фукуда Т, Мацумото Т, Сэкинэ К., Сато Т, Накамичи Й, Ямамото Й. , Ёсимура К., Ватанабэ Т., Накамура Т., Оомидзу А., Цукада М., Хаяси Х., Судо Т., Като С., Мацумото Т. (июнь 2007 г.). «Штамм-зависимая эмбриональная летальность и чрезмерное ремоделирование сосудов у мышей с дефицитом кофактора II гепарина». Журнал клинических исследований. 117 (6): 1514–26. Дои:10.1172 / JCI27095. ЧВК 1878511. PMID 17549254.
- ^ Кейл Дж. М., Лоуренс Д. А. (сентябрь 2007 г.). «Структурно-функциональные отношения ингибитора активатора плазминогена-1 и его потенциал в качестве терапевтического агента». Текущие цели в отношении лекарств. 8 (9): 971–81. Дои:10.2174/138945007781662337. PMID 17896949.
- ^ Лино М.М., Атанасоски С., Кваджо М., Фаярд Б., Морено Е., Бреннер Х. Р., Сутер Ю., Монард Д. (апрель 2007 г.). «У мышей, лишенных протеазы нексин-1, наблюдается замедленное структурное и функциональное восстановление после раздавливания седалищного нерва». Журнал неврологии. 27 (14): 3677–85. Дои:10.1523 / JNEUROSCI.0277-07.2007. ЧВК 6672422. PMID 17409231.
- ^ Murer V, Spetz JF, Hengst U, Altrogge LM, de Agostini A, Monard D (март 2001 г.). «Нарушения фертильности самцов у мышей, лишенных ингибитора сериновой протеазы протеазы нексин-1». Труды Национальной академии наук Соединенных Штатов Америки. 98 (6): 3029–33. Bibcode:2001ПНАС ... 98.3029М. Дои:10.1073 / pnas.051630698. ЧВК 30601. PMID 11248026.
- ^ Люти А., Ван дер Путтен Х., Боттери Ф.М., Мансуй И.М., Мейнс М., Фрей Ю., Сансиг Дж., Портет С., Шмутц М., Шредер М., Нитч С., Лоран Дж. П., Монард Д. (июнь 1997 г.). «Ингибитор эндогенной сериновой протеазы модулирует эпилептическую активность и долгосрочную потенциацию гиппокампа». Журнал неврологии. 17 (12): 4688–99. Дои:10.1523 / JNEUROSCI.17-12-04688.1997. ЧВК 6573330. PMID 9169529.
- ^ а б Doll JA, Stellmach VM, Bouck NP, Bergh AR, Lee C, Abramson LP, Cornwell ML, Pins MR, Borensztajn J, Crawford SE (июнь 2003 г.). «Фактор пигментного эпителия регулирует сосудистую сеть и массу простаты и поджелудочной железы». Природа Медицина. 9 (6): 774–80. Дои:10,1038 / нм 870. PMID 12740569. S2CID 5967666.
- ^ Бесерра С.П., Перес-Медиавилла Л.А., Велдон Дж. Э., Локателли-Хупс С., Сенанаяке П., Нотари Л., Нотарио В., Холлифилд Дж. Г. (ноябрь 2008 г.). «Фактор, происходящий из пигментного эпителия, связывается с гиалуронаном. Картирование сайта связывания гиалуронана». Журнал биологической химии. 283 (48): 33310–20. Дои:10.1074 / jbc.M801287200. ЧВК 2586245. PMID 18805795.
- ^ Андреу-Агульо С., Моранте-Редолат Ж.М., Дельгадо А.С., Фариньяс I (декабрь 2009 г.). «Фактор сосудистой ниши PEDF модулирует Notch-зависимую стволовость в субэпендимальной зоне взрослых». Природа Неврология. 12 (12): 1514–23. Дои:10.1038 / № 2437. PMID 19898467. S2CID 5332822.
- ^ Виман Б., Коллен Д. (сентябрь 1979 г.). «О механизме реакции между альфа-2-антиплазмином человека и плазмином». Журнал биологической химии. 254 (18): 9291–7. PMID 158022.
- ^ Lijnen HR, Okada K, Matsuo O, Collen D, Dewerchin M (апрель 1999 г.). «Дефицит гена альфа2-антиплазмина у мышей связан с повышенным фибринолитическим потенциалом без явного кровотечения». Кровь. 93 (7): 2274–81. Дои:10.1182 / кровь.V93.7.2274. PMID 10090937.
- ^ Карпентер С.Л., Мэтью П. (ноябрь 2008 г.). «Альфа2-антиплазмин и его дефицит: дисбаланс фибринолиза». Гемофилия. 14 (6): 1250–4. Дои:10.1111 / j.1365-2516.2008.01766.x. PMID 19141165. S2CID 205295156.
- ^ Фавье Р., Аоки Н., Де Мёрлоуз П. (1 июля 2001 г.). «Врожденная недостаточность ингибитора α2-плазмина: обзор». Британский журнал гематологии. 114 (1): 4–10. Дои:10.1046 / j.1365-2141.2001.02845.x. ISSN 1365-2141. PMID 11472338. S2CID 71010865.
- ^ Beinrohr L, Harmat V, Dobó J, Lörincz Z, Gál P, Závodszky P (июль 2007 г.). «Структура домена серпина ингибитора C1 раскрывает вероятный механизм потенцирования гепарина и конформационного заболевания». Журнал биологической химии. 282 (29): 21100–9. Дои:10.1074 / jbc.M700841200. PMID 17488724.
- ^ Моллнес Т.Э., Йокиранта Т.С., Труэдссон Л., Нильссон Б., Родригес де Кордоба С., Киршфинк М. (сентябрь 2007 г.). «Комплементарный анализ в 21 веке». Молекулярная иммунология. 44 (16): 3838–49. Дои:10.1016 / j.molimm.2007.06.150. HDL:10261/61732. PMID 17768101.
- ^ Triggianese P, Chimenti MS, Toubi E, Ballanti E, Guarino MD, Perricone C, Perricone R (август 2015 г.). «Аутоиммунная сторона наследственного ангионевротического отека: сведения о патогенезе». Отзывы об аутоиммунности. 14 (8): 665–9. Дои:10.1016 / j.autrev.2015.03.006. PMID 25827463.
- ^ Нагаи Н., Хосокава М., Итохара С., Адачи Е., Мацусита Т., Хосокава Н., Нагата К. (сентябрь 2000 г.). «Эмбриональная летальность мышей с нокаутом молекулярного шаперона hsp47 связана с дефектами биосинтеза коллагена». Журнал клеточной биологии. 150 (6): 1499–506. Дои:10.1083 / jcb.150.6.1499. ЧВК 2150697. PMID 10995453.
- ^ Марини Дж. К., Райх А., Смит С. М. (август 2014 г.). «Несовершенный остеогенез из-за мутаций в неколлагеновых генах: уроки биологии образования кости». Текущее мнение в педиатрии. 26 (4): 500–7. Дои:10.1097 / MOP.0000000000000117. ЧВК 4183132. PMID 25007323.
- ^ Байерс PH, Pyott SM (1 января 2012 г.). «Рецессивно наследуемые формы несовершенного остеогенеза». Ежегодный обзор генетики. 46: 475–97. Дои:10.1146 / annurev-genet-110711-155608. PMID 23145505.
- ^ Остервальдер Т., Чинелли П., Байчи А., Пеннелла А., Крюгер С.Р., Шримпф С.П., Майнс М., Сондереггер П. (январь 1998 г.). «Ингибитор сериновой протеиназы, секретируемый аксонами, нейросерпин, ингибирует активаторы плазминогена и плазмин, но не тромбин». Журнал биологической химии. 273 (4): 2312–21. Дои:10.1074 / jbc.273.4.2312. PMID 9442076.
- ^ Crowther DC (июль 2002 г.). «Семейные конформационные заболевания и деменции». Человеческая мутация. 20 (1): 1–14. Дои:10.1002 / humu.10100. PMID 12112652. S2CID 22326349.
- ^ Белоргей Д., Хэгглёф П., Карлссон-Ли С., Ломас Д.А. (1 марта 2007 г.). «Неправильный фолдинг белков и серпинопатии». Прион. 1 (1): 15–20. Дои:10.4161 / pri.1.1.3974. ЧВК 2633702. PMID 19164889.
- ^ Одзаки К., Нагата М., Судзуки М., Фудзивара Т., Миёси Ю., Исикава О., Охигаши Х., Имаока С., Такахаши Е., Накамура И. (июль 1998 г.). «Выделение и характеристика нового специфичного для поджелудочной железы гена человека, pancpin, который подавляется в раковых клетках поджелудочной железы». Гены, хромосомы и рак. 22 (3): 179–85. Дои:10.1002 / (SICI) 1098-2264 (199807) 22: 3 <179 :: AID-GCC3> 3.0.CO; 2-T. PMID 9624529.
- ^ Loftus SK, Cannons JL, Incao A, Pak E, Chen A, Zerfas PM, Bryant MA, Biesecker LG, Schwartzberg PL, Pavan WJ (сентябрь 2005 г.). «Апоптоз ацинарных клеток у мышей с дефицитом Serpini2, моделирующих панкреатическую недостаточность». PLOS Genetics. 1 (3): e38. Дои:10.1371 / journal.pgen.0010038. ЧВК 1231717. PMID 16184191.
- ^ Падуя МБ, Ковальски А.А., Каньяс М.Ю., Хансен П.Дж. (февраль 2010 г.). «Молекулярная филогения серпинов матки и ее связь с эволюцией плацентации». Журнал FASEB. 24 (2): 526–37. Дои:10.1096 / fj.09-138453. PMID 19825977. S2CID 9248169.
- ^ Падуя МБ, Хансен П.Дж. (октябрь 2010 г.). «Эволюция и функция серпинов матки (SERPINA14)». Американский журнал репродуктивной иммунологии. 64 (4): 265–74. Дои:10.1111 / j.1600-0897.2010.00901.x. PMID 20678169.
- ^ а б Райххарт JM (декабрь 2005 г.). «Верхушка еще одного айсберга: змеиные дрозофилы». Тенденции в клеточной биологии. 15 (12): 659–65. Дои:10.1016 / j.tcb.2005.10.001. PMID 16260136.
- ^ Тан Х, Камбрис З., Леметр Б., Хашимото С. (октябрь 2008 г.). «Серпин, регулирующий иммунную меланизацию дыхательной системы дрозофилы». Клетка развития. 15 (4): 617–26. Дои:10.1016 / j.devcel.2008.08.017. ЧВК 2671232. PMID 18854145.
- ^ Шерфер С., Танг Х., Камбрис З., Лхосин Н., Хашимото С., Леметр Б. (ноябрь 2008 г.). «Drosophila Serpin-28D регулирует активность фенолоксидазы гемолимфы и пигментацию взрослых». Биология развития. 323 (2): 189–96. Дои:10.1016 / j.ydbio.2008.08.030. PMID 18801354.
- ^ Рашлоу C (январь 2004 г.). "Дорсовентральное формирование паттерна: змеевик наконец-то прижат". Текущая биология. 14 (1): R16–8. Дои:10.1016 / j.cub.2003.12.015. PMID 14711428.
- ^ Ligoxygakis P, Roth S, Reichhart JM (декабрь 2003 г.). «Серпин регулирует формирование дорсально-вентральной оси у эмбриона дрозофилы». Текущая биология. 13 (23): 2097–102. Дои:10.1016 / j.cub.2003.10.062. PMID 14654000.
- ^ Цзян Р., Чжан Б., Курокава К., Со И, Ким Э., Хван Хо, Ли Дж. Х., Сирацучи А., Чжан Дж., Наканиши Ю., Ли Х. С., Ли Б. Л. (октябрь 2011 г.). «93-кДа двухдоменный ингибитор сериновой протеазы (Serpin) выполняет регуляторную функцию в каскаде протеолитической передачи сигналов Toll жука». Журнал биологической химии. 286 (40): 35087–95. Дои:10.1074 / jbc.M111.277343. ЧВК 3186399. PMID 21862574.
- ^ а б Пак С.К., Кумар В., Цу С., Люк С.Дж., Аскью Ю.С., Аскью Д.И., Миллс Д.Р., Бремме Д., Сильверман Г.А. (апрель 2004 г.). «SRP-2 представляет собой межклассовый ингибитор, который участвует в постэмбриональном развитии нематоды Caenorhabditis elegans: начальная характеристика серпинов клады L». Журнал биологической химии. 279 (15): 15448–59. Дои:10.1074 / jbc.M400261200. PMID 14739286.
- ^ Люк С.Дж., Пак С.К., Аскью Ю.С., Навильа Т.Л., Аскью Д.И., Нобар С.М., Ветика А.С., Лонг ОС, Уоткинс СК, Штольц Д.Б., Барстед Р.Дж., Молдер Г.Л., Бремме Д., Сильверман Г.А. (сентябрь 2007 г.). «Внутриклеточный серпин регулирует некроз, подавляя индукцию и последствия лизосомного повреждения». Клетка. 130 (6): 1108–19. Дои:10.1016 / j.cell.2007.07.013. ЧВК 2128786. PMID 17889653.
- ^ Хейгаард Дж, Расмуссен С.К., Брандт А, Свендсени (1985). «Гомология последовательности между протеином Z эндосперма ячменя и ингибиторами протеазы семейства альфа-1-антитрипсинов». FEBS Lett. 180 (1): 89–94. Дои:10.1016/0014-5793(85)80238-6. S2CID 84790014.
- ^ Даль С.В., Расмуссен С.К., Петерсен Л.С., Хейгаард Дж. (Сентябрь 1996 г.). «Ингибирование факторов свертывания рекомбинантным серпином ячменя BSZx». Письма FEBS. 394 (2): 165–8. Дои:10.1016/0014-5793(96)00940-4. PMID 8843156.
- ^ Хейгаард Дж (январь 2001 г.). «Тормозящие серпины из зерна ржи с глутамином в качестве остатков P1 и P2 в реактивном центре». Письма FEBS. 488 (3): 149–53. Дои:10.1016 / S0014-5793 (00) 02425-X. PMID 11163762. S2CID 27933086.
- ^ Ostergaard H, Rasmussen SK, Roberts TH, Hejgaard J (октябрь 2000 г.). «Тормозящие серпины из пшеничного зерна с реактивными центрами, напоминающими богатые глутамином повторы запасных белков проламина. Клонирование и характеристика пяти основных молекулярных форм». Журнал биологической химии. 275 (43): 33272–9. Дои:10.1074 / jbc.M004633200. PMID 10874043.
- ^ а б Yoo BC, Aoki K, Xiang Y, Campbell LR, Hull RJ, Xoconostle-Cázares B, Monzer J, Lee JY, Ullman DE, Lucas WJ (ноябрь 2000 г.). «Характеристика серпина-1 флоэмы тыквы максимума (CmPS-1). Ингибитор эластазы, регулируемый развитием». Журнал биологической химии. 275 (45): 35122–8. Дои:10.1074 / jbc.M006060200. PMID 10960478.
- ^ la Cour Petersen M, Hejgaard J, Thompson GA, Schulz A (декабрь 2005 г.). «Серпины тыквенного флоэмы передаются трансплантатом и, по-видимому, устойчивы к обмену в комплексе ситовых элементов и клеток-компаньонов». Журнал экспериментальной ботаники. 56 (422): 3111–20. Дои:10.1093 / jxb / eri308. PMID 16246856.
- ^ Робертс Т.Х., Хейгаард Дж. (Февраль 2008 г.). «Серпины в растениях и зеленых водорослях». Функциональная и интегративная геномика. 8 (1): 1–27. Дои:10.1007 / s10142-007-0059-2. PMID 18060440. S2CID 22960858.
- ^ Лампл Н., Алкан Н., Давыдов О., Флур Р. (май 2013 г.). «Контрольная точка активности протеазы RD21 с помощью AtSerpin1 контролирует гибель клеток у Arabidopsis». Журнал растений. 74 (3): 498–510. Дои:10.1111 / tpj.12141. PMID 23398119.
- ^ Ан Дж. У., Этвелл Б. Дж., Робертс Т. Х. (2009). «Серпиновые гены AtSRP2 и AtSRP3 необходимы для нормальной чувствительности роста к ДНК-алкилирующему агенту у Arabidopsis». BMC Биология растений. 9: 52. Дои:10.1186/1471-2229-9-52. ЧВК 2689219. PMID 19426562.
- ^ а б Канг С., Барак И., Ламед Р., Байер Е.А., Моррисон М. (июнь 2006 г.). «Функциональный репертуар прокариотных целлюлосом включает серпиновое суперсемейство ингибиторов сериновых протеиназ». Молекулярная микробиология. 60 (6): 1344–54. Дои:10.1111 / j.1365-2958.2006.05182.x. PMID 16796673. S2CID 23738804.
- ^ Ирвинг Дж. А., Кабрита Л. Д., Россджон Дж., Пайк Р. Н., Боттомли С. П., Уиссток Дж. К. (апрель 2003 г.). «Кристаллическая структура 1,5 A серпина прокариот: управление конформационными изменениями в нагретой среде». Структура. 11 (4): 387–97. Дои:10.1016 / S0969-2126 (03) 00057-1. PMID 12679017.
- ^ Фултон К.Ф., Бакл А.М., Кабрита Л.Д., Ирвинг Дж. А., Батчер Р. Э., Смит И., Рив С., Леск А. М., Боттомли С. П., Россджон Дж., Уиссток Дж. С. (март 2005 г.). «Кристаллическая структура с высоким разрешением природного термостабильного серпина раскрывает сложный механизм, лежащий в основе перехода от напряжения к расслаблению». Журнал биологической химии. 280 (9): 8435–42. Дои:10.1074 / jbc.M410206200. PMID 15590653.
- ^ Тернер П.С., Мойер Р.В. (сентябрь 2002 г.). «Иммуномодуляторы поксвируса: функциональные идеи на животных моделях». Вирусные исследования. 88 (1–2): 35–53. Дои:10.1016 / S0168-1702 (02) 00119-3. PMID 12297326.
- ^ а б Ричардсон Дж, Вишванатан К, Лукас А (2006). «Серпины, сосудистая сеть и вирусная терапия». Границы биологических наук. 11: 1042–56. Дои:10.2741/1862. PMID 16146796.
- ^ Цзян Дж., Арп Дж., Кубелик Д., Зассоко Р., Лю В., Мудрый И., Маколей К., Гарсия Б., Макфадден Г., Лукас А. Р., Ван Х. (ноябрь 2007 г.). «Индукция неопределенной выживаемости сердечного аллотрансплантата коррелирует с подавлением толл-подобных рецепторов 2 и 4 после лечения ингибитором сериновой протеазы-1 (Serp-1)». Трансплантация. 84 (9): 1158–67. Дои:10.1097 / 01.tp.0000286099.50532.b0. PMID 17998872. S2CID 20168458.
- ^ Дай Э, Гуань Х., Лю Л., Литтл С., Макфадден Г., Вазири С., Цао Х., Иванова И.А., Бокш Л., Лукас А. (май 2003 г.). «Серп-1, вирусный противовоспалительный серпин, регулирует клеточные сериновые протеиназы и серпиновые реакции на сосудистое повреждение». Журнал биологической химии. 278 (20): 18563–72. Дои:10.1074 / jbc.M209683200. PMID 12637546.
- ^ Тернер П.К., Санчо М.К., Тоннс С.Р., Капуто А., Блекли Р.С., Мойер Р.В. (август 1999 г.). «Вирус миксомы Serp2 является слабым ингибитором гранзима B и фермента, превращающего интерлейкин-1-бета in vitro, и в отличие от CrmA не может блокировать апоптоз в клетках, инфицированных вирусом коровьей оспы». Журнал вирусологии. 73 (8): 6394–404. Дои:10.1128 / JVI.73.8.6394-6404.1999. ЧВК 112719. PMID 10400732.
- ^ Мунусвами-Рамануджам Г., Хан К.А., Лукас А.Р. (декабрь 2006 г.). «Вирусные противовоспалительные реагенты: потенциал для лечения артритов и васкулитов». Целевые препараты для лечения эндокринных, метаболических и иммунных расстройств. 6 (4): 331–43. Дои:10.2174/187153006779025720. PMID 17214579.
- ^ Renatus M, Zhou Q, Stennicke HR, Snipas SJ, Turk D, Bankston LA, Liddington RC, Salvesen GS (июль 2000 г.). «Кристаллическая структура супрессора апоптоза CrmA в его расщепленной форме». Структура. 8 (7): 789–97. Дои:10.1016 / S0969-2126 (00) 00165-9. PMID 10903953.
внешняя ссылка
- PDB Молекула месяца Серпин
- Хромота ингибитора протеазы меропса (семейство I4)
- Серпины в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Лаборатория Джеймса Уисстока в Университет Монаша
- Лаборатория Джима Хантингтона в Кембриджский университет
- Лаборатория Фрэнка Черча в Университет Северной Каролины в Чапел-Хилл
- Лаборатория Поля Деклерка в Katholieke Universiteit Leuven
- Лаборатория Тома Робертса в Сиднейский университет
- Лаборатория Роберта Флюра в Институт науки Вейцмана
- Лаборатория Питера Геттинса в Иллинойский университет в Чикаго
- Обзор всей структурной информации, доступной в PDB за UniProt: P01009 (Альфа-1-антитрипсин человека) в PDBe-KB.