Интерлейкин 13 - Interleukin 13
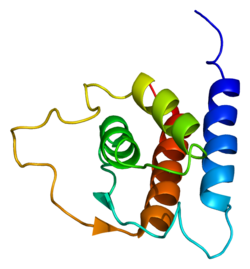


Интерлейкин 13 (Ил-13) это белок что у людей кодируется IL13 ген.[4][5][6] IL-13 был впервые клонирован в 1993 году и расположен на хромосоме 5q31 длиной 1,4 kb.[4] Он имеет массу 13 кДа и складывается в 4 альфа-спиральных пучка.[7] Вторичные структурные особенности ИЛ-13 аналогичны таковым у Интерлейкин 4 (Ил-4); однако он имеет только 25% гомологию последовательности с IL-4 и способен передавать независимый от IL-4 сигнал.[7][4][8] Ил-13 - это цитокин секретно Т помощник клетки типа 2 (Th2), CD4 клетки естественные Т-клетки-киллеры, тучные клетки, базофилы, эозинофилы и нуоциты.[7] Интерлейкин-13 является центральным регулятором синтеза IgE, гиперплазии бокаловидных клеток, гиперсекреции слизи, гиперчувствительности дыхательных путей, фиброза и активации хитиназы.[7] Это посредник аллергической воспаление и различные заболевания, включая астма.[7]
Функции
IL-13 оказывает действие на иммунные клетки, аналогичное влиянию близкородственного цитокина. Ил-4.[4]Однако предполагается, что IL-13 является центральным медиатором физиологических изменений, вызванных аллергическим воспалением во многих тканях.[4]
Хотя IL-13 связан в первую очередь с индукцией заболевания дыхательных путей, он также имеет противовоспалительное средство свойства.[4] IL-13 индуцирует класс ферментов, разрушающих белок, известных как матричные металлопротеиназы (MMPs), в дыхательных путях.[4] Эти ферменты необходимы, чтобы вызвать агрессию паренхиматозный воспалительные клетки попадают в просвет дыхательных путей, откуда они удаляются.[4] Среди других факторов, IL-13 индуцирует эти MMP как часть механизма, который защищает от чрезмерного аллергического воспаления, которое предрасполагает к асфиксии.[4]
Известно, что IL-13 вызывает изменения в кроветворный клетки, но эти эффекты, вероятно, менее важны, чем у IL-4.[4] Кроме того, IL-13 может вызывать иммуноглобулин E (IgE) секреция активированного человека В-клетки.[4][7] Делеция IL-13 у мышей не оказывает заметного влияния ни на развитие Th2-клеток, ни на антиген-специфические IgE-ответы, вызванные сильнодействующим действием. аллергены.[4] Для сравнения, удаление IL-4 деактивирует эти ответы. Таким образом, вместо лимфоидного цитокина, IL-13 играет более важную роль в качестве молекулярного мостика, связывающего аллергические воспалительные клетки с неиммунными клетками, контактирующими с ними, тем самым изменяя физиологическую функцию.[4]
Передача сигналов IL-13 начинается через общий мульти-субъединичный рецептор с IL-4.[7] Этот рецептор представляет собой комплекс гетеродимерных рецепторов, состоящий из альфа Рецептор ИЛ-4 (IL-4Rα) и альфа-интерлейкин-13 рецептор (ИЛ-13R1).[7] Высокое сродство IL-13 к IL-13R1 приводит к образованию их связи, что дополнительно увеличивает вероятность образования гетеродимера к IL-4R1 и продукции рецептора IL-4 типа 2. Гетеродимеризация активирует как STAT6 и IRS.[7] Передача сигналов STAT6 важна для инициирования аллергической реакции.[7] Большинство биологических эффектов IL-13, как и IL-4, связаны с одним фактор транскрипции, преобразователь сигнала и активатор транскрипции 6 (STAT6 ).[7] Интерлейкин-13 и связанные с ним рецепторы с α-субъединицей рецептора IL-4 (IL-4Rα) допускают последующую активацию STAT6.[9] JAK Янус киназа белки на цитоплазматическом конце рецепторов позволяют фосфорилировать STAT6, который затем образует активированный гомодимер и транспортируется в ядро.[9] Попадая в ядро, гетеродимерная молекула STAT6 регулирует экспрессию генов типов клеток, критических для баланса между иммунной защитой хозяина и аллергическими воспалительными реакциями, такими как развитие Th2.[9] Это может быть результатом аллергической реакции, вызванной столкновением с геном Ala. IL-13 также связывается с другим рецептором, известным как IL-13Rα2.[10] IL-13Rα2 (который обозначен как рецептор-ловушка) происходит из клеток Th2 и является плеотропным иммунорегулирующим цитокином.[10] IL-13 имеет большее сродство (в 50 раз) к IL-13Rα2, чем к IL-13Ra1.[10] Субъединица IL-13Rα2 связывается только с IL-13 и существует как в мембраносвязанной, так и в растворимой форме у мышей.[10] Растворимая форма IL-13Rα2 не была обнаружена у людей.[10] Исследования легких трансгенных мышей с IL-13Rα2 нулевыми локусами показали, что дефицит IL-13Rα2 значительно усиливает индуцированное IL-13 или овальбумином воспаление легких и ремоделирование.[10] Большинство нормальных клеток, таких как иммунные клетки или эндотелиальные клетки, экспрессируют очень низкие или неопределяемые уровни рецепторов IL-13.[10] Исследования показали, что экспрессия IL-13Rα2 на клеточной поверхности фибробластов дыхательных путей человека была снижена по сравнению с экспрессией на нормальных контрольных фибробластах дыхательных путей.[10] Это подтвердило гипотезу о том, что IL-13Rα2 является негативным регулятором индуцированного IL-13 ответа, и продемонстрировало значительное снижение продукции TGF-β1 и отложения коллагена в легких мышей.[10]
Интерлейкин-13 играет важную роль в Бокоровая ячейка метаплазия.[11] Бокаловидные клетки заполнены муцином (MUC).[11] MUC5AC Муцин 5AC представляет собой гелеобразный муциновый продукт бокаловидных клеток.[11] Интерлейкин-13 вызывает дифференцировку бокаловидных клеток и позволяет продуцировать MUC5AC в эпителии трахеи.[11] 15-Липоксигеназа-1 (15LO1), которая является ферментом метаболизма жирных кислот, и ее метаболит, 15-HETE, высоко экспрессируются при астме (что приводит к сверхэкспрессии MUC5AC) и индуцируются IL-13 в эпителиальных тканях дыхательных путей человека. клетки. По мере увеличения количества бокаловидных клеток в бронхах происходит чрезмерное выделение слизи.[11] Функциональные последствия изменений в хранении и секреции MUC вносят вклад в патофизиологические механизмы различных клинических аномалий у пациентов с астмой, включая образование мокроты, сужение дыхательных путей, обострение и ускоренную потерю функции легких.[11]
Кроме того, было показано, что IL-13 вызывает мощную фиброгенную программу во время различных заболеваний, отмеченных повышенными цитокинами типа 2, таких как хронический шистосомоз и атопический дерматит среди других. Было высказано предположение, что эта фиброгенная программа критически зависит от прямой передачи сигналов IL-13 через IL-4Rα на фибробласты PDGFRβ +.[12]
Клиническое значение
IL-13 специфически вызывает физиологические изменения в паразитированный органы, которые необходимы для изгнания организмов-нарушителей или их продуктов. Например, изгнание из кишечника различных мышей. гельминты требует IL-13, секретируемого клетками Th2. IL-13 вызывает несколько изменений в кишечнике, которые создают среду, враждебную паразиту, включая усиление сокращений и гликопротеин гиперсекреция из кишечника эпителиальный клетки, что в конечном итоге приводит к отслоению организма от стенки кишечника и их удалению.[13]
Яйца паразита Schistosoma mansoni может оседать в различных органах, включая стенку кишечника, печень, легкие и даже центральную нервную систему, вызывая образование гранулемы под контролем ИЛ-13. Однако здесь конечным результатом является повреждение органов и часто серьезное или даже смертельное заболевание, а не исчезновение инфекции. Возникающая концепция заключается в том, что IL-13 может противодействовать ответам Th1, которые необходимы для устранения внутриклеточного инфекции. В этом контексте иммунной дисрегуляции, отмеченной набором аномально большого количества Th2-клеток, IL-13 подавляет способность иммунных клеток хозяина разрушать внутриклеточный патогены.
Экспрессия IL-13 была увеличена в жидкости и клетках бронхоальвеолярного лаважа (БАЛ) у пациентов с атопической легкой формой астмы после провокации аллергеном.[14] Полногеномные исследования ассоциации выявили множественные полиморфизмы IL-13 и генов, кодирующих рецепторы IL-13, которые связаны с восприимчивостью к астме, гиперреактивностью бронхов и повышенным уровнем IgE.[14] Сверхэкспрессия IL-13 вызывает многие особенности аллергическое заболевание легких, включая гиперреактивность дыхательных путей, кубок метаплазия, гиперсекреция слизи и ремоделирование дыхательных путей, которые способствуют обструкции дыхательных путей.[15] Исследования на мышах показали, что IL-13 необходим и достаточен для генерации астматических Th2-ответов в легких мыши.[7] IL-13 в основном сверхэкспрессируется в мокроте, подслизистой оболочке бронхов, периферической крови и тучных клетках гладкомышечного пучка дыхательных путей.[7] IL-4 вносит вклад в эти физиологические изменения, но менее важен, чем IL-13. IL-13 также вызывает секрецию хемокины которые необходимы для рекрутирования аллергических эффекторных клеток в легкие. Исследования трансгенных мышей STAT6 предполагают возможность того, что передача сигналов IL-13, происходящая только через эпителий дыхательных путей, необходима для большинства этих эффектов. Несмотря на то, что до сих пор нет исследований прямого участия IL-13 в борьбе с заболеваниями человека, многие полиморфизмы в гене IL-13, как было показано, увеличивает риск атопических респираторных заболеваний, таких как астма.[13] В исследовании, проведенном с использованием модели мышей с нокаутом астмы, было обнаружено, что сопротивление воздуха, образование слизи и индукция профиброгенного медиатора зависят исключительно от присутствия IL-13R1, а не IL-13Rα2.[7] Исследования на трансгенных мышах in vivo демонстрируют, что сверхэкспрессия IL-13 в легких вызывает субэпителиальный фиброз дыхательных путей.[7] IL-13 является доминирующим эффектором в моделях фиброза, вызванных токсинами, инфекциями, аллергическими заболеваниями и облитерирующим посттрансплантационным бронхиолитом.[7]
Другие исследования предполагают, что IL-13 отвечает за выживание и миграцию эпителиальных клеток, продукцию индуцибельной синтазы оксида азота эпителиальными клетками дыхательных путей, активацию макрофагов, проницаемость эпителиальных клеток и трансформацию фибробластов дыхательных путей в миофибробласты. что приводит к отложению коллагена.[14] Затем отложение влияет на ремоделирование дыхательных путей у пациентов с астмой.[14]
Дупилумаб представляет собой моноклональное антитело к IL-13 и модулятор IL-4, нацеленное на общий рецептор Ил-4 и Ил-13, IL4Rα.[16] Поскольку ИЛ-4 и ИЛ-13 обладают сходной биологической активностью, дупилумаб может быть эффективной формой лечения пациентов с астмой.[16]
Смотрите также
- Рецептор интерлейкина-13, рецептор IL-13
использованная литература
- ^ а б c GRCm38: выпуск ансамбля 89: ENSMUSG00000020383 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d е ж г час я j k л м Минти А., Шалон П., Дерок Дж. М., Дюмон X, Гийемо Дж. К., Кагад М., Лабит С., Леплатуа П., Лиазун П., Милу Б. (март 1993 г.). «Интерлейкин-13 - новый человеческий лимфокин, регулирующий воспалительные и иммунные реакции». Природа. 362 (6417): 248–50. Bibcode:1993Натура.362..248М. Дои:10.1038 / 362248a0. PMID 8096327. S2CID 4368915.
- ^ Маккензи А.Н., Калпеппер Дж. А., де Ваал Малефит Р., Бриер Ф., Пуннонен Дж., Аверса Дж., Сато А., Данг В., Кокс Б. Г., Менон С. (апрель 1993 г.). «Интерлейкин 13, цитокин, полученный из Т-клеток, который регулирует функцию моноцитов и В-клеток человека». Труды Национальной академии наук Соединенных Штатов Америки. 90 (8): 3735–9. Bibcode:1993ПНАС ... 90.3735М. Дои:10.1073 / пнас.90.8.3735. ЧВК 46376. PMID 8097324.
- ^ Морган Дж. Г., Долганов Г. М., Роббинс С. Е., Хинтон Л. М., Ловетт М. (октябрь 1992 г.). «Селективное выделение новых кДНК, кодируемых областями, окружающими гены интерлейкина 4 и 5 человека». Нуклеиновые кислоты Res. 20 (19): 5173–9. Дои:10.1093 / nar / 20.19.5173. ЧВК 334302. PMID 1408833.
- ^ а б c d е ж г час я j k л м п о п Раэль Э.Л., Локки РФ (2011). «Передача сигналов интерлейкина-13 и его роль при астме». Журнал Всемирной организации аллергии. 4 (3): 54–64. Дои:10.1097 / WOX.0b013e31821188e0. ЧВК 3651056. PMID 23283176.
- ^ Zurawski G, de Vries JE (январь 1994). «Интерлейкин 13, интерлейкин-4-подобный цитокин, который действует на моноциты и В-клетки, но не на Т-клетки». Иммунол. сегодня. 15 (1): 19–26. Дои:10.1016/0167-5699(94)90021-3. PMID 7907877.
- ^ а б c Уолфорд Х. Х., Доэрти Т. А. (2013). «STAT6 и воспаление легких». Jak-Stat. 2 (4): e25301. Дои:10.4161 / jkst.25301. ЧВК 3876430. PMID 24416647.
- ^ а б c d е ж г час я Ту М, Ванге В., Цай Л., Чжу П, Гао З, Чжэн В. (2016). «Рецептор IL-13 α2 стимулирует рост и метастазирование клеток глиомы человека через сигнальный путь Src / PI3K / Akt / mTOR». Биология опухоли. 37 (11): 14701–14709. Дои:10.1007 / s13277-016-5346-х. PMID 27623944. S2CID 30389002.
- ^ а б c d е ж СП Фахи (2002). «Нарушения гена бокаловидных клеток и муцина при астме». Грудь. 122 (6 Прил.): 320S – 326S. Дои:10.1378 / сундук.122.6_suppl.320S. PMID 12475809. S2CID 23113468.
- ^ Гизек Р.Л., Рамалингам Т.Р., Харт К.М., Ваннелла К.М., Канту Д.А., Лу В.Й., Феррейра-Гонсалес С., Форбс С.Дж., Валлиер Л., Винн Т.А. (2016). «Интерлейкин-13 активирует определенные клеточные пути, ведущие к протоковой реакции, стеатозу и фиброзу». Иммунитет. 45 (1): 145–58. Дои:10.1016 / j.immuni.2016.06.009. ЧВК 4956513. PMID 27421703.
- ^ а б Сейфизаде Н, Сейфизаде Н, Бабалу З (2014). «Интерлейкин-13 как важный медиатор: обзор его роли в некоторых заболеваниях человека». Иранский журнал аллергии, астмы и иммунологии. В прессе.
- ^ а б c d Инграм Дж. Л., Крафт М (2012). «IL-13 при астме и аллергических заболеваниях: фенотипы астмы и таргетная терапия». Журнал аллергии и клинической иммунологии. 130 (4): 829–42, викторина 843–4. Дои:10.1016 / j.jaci.2012.06.034. PMID 22951057.
- ^ Уиллс-Карп М., Луимбази Дж., Сюй Х, Скофилд Б., Небен Т.Ю., Карп С.Л., Дональдсон Д.Д. (декабрь 1998 г.). «Интерлейкин-13: центральный медиатор аллергической астмы». Наука. 282 (5397): 2258–61. Bibcode:1998Sci ... 282.2258W. Дои:10.1126 / science.282.5397.2258. PMID 9856949.
- ^ а б Vatrella A, Fabozzi I, Calabrese C, Maselli R, Pelaia G (2014). «Дупилумаб: новое средство от астмы». Журнал астмы и аллергии. 7: 123–30. Дои:10.2147 / JAA.S52387. ЧВК 4159398. PMID 25214796.
дальнейшее чтение
- Мароне Дж., Флорио Дж., Петрароли А., де Паулис А. (2001). «Нарушение регуляции сети IgE / Fc epsilon RI при ВИЧ-1 инфекции». J. Allergy Clin. Иммунол. 107 (1): 22–30. Дои:10.1067 / mai.2001.111589. PMID 11149986.
- Мароне Дж., Флорио Дж., Тригиани М., Петрароли А., де Паулис А. (2001). «Механизмы повышения уровня IgE при ВИЧ-1 инфекции». Крит. Rev. Immunol. 20 (6): 477–96. Дои:10.1615 / critrevimmunol.v20.i6.40. PMID 11396683.
- Скиннидер Б.Ф., Капп У., Мак Т.В. (2003). «Роль интерлейкина 13 в классической лимфоме Ходжкина». Лейк. Лимфома. 43 (6): 1203–10. Дои:10.1080/10428190290026259. PMID 12152987. S2CID 21083414.
- Идзухара К., Арима К., Ясунага С. (2003). «IL-4 и IL-13: их патологическая роль в аллергических заболеваниях и их потенциал в разработке новых методов лечения». Текущие целевые показатели по лекарствам. Воспаление и аллергия. 1 (3): 263–9. Дои:10.2174/1568010023344661. PMID 14561191.
- Дессейн А., Куриба Б., Эбумбу С., Дессейн Х., Аргиро Л., Марке С., Элвали Н. Э., Родригес В., Ли И, Думбо О., Шевиллард С. (2005). «Интерлейкин-13 в коже и гамма-интерферон в печени играют ключевую роль в иммунной защите при шистосомозе человека». Иммунол. Rev. 201: 180–90. Дои:10.1111 / j.0105-2896.2004.00195.x. PMID 15361241. S2CID 25378236.
- Коупленд К.Ф. (2006). «Модуляция транскрипции ВИЧ-1 цитокинами и хемокинами». Мини-обзоры по медицинской химии. 5 (12): 1093–101. Дои:10.2174/138955705774933383. PMID 16375755.
внешняя ссылка
Галерея PDB | |
|---|---|
|


