Schistosoma mansoni - Schistosoma mansoni
| Schistosoma mansoni | |
|---|---|
 | |
| Шистосомы совокупляются | |
| Научная классификация | |
| Королевство: | |
| Тип: | |
| Учебный класс: | |
| Заказ: | |
| Семья: | |
| Род: | |
| Разновидность: | S. mansoni |
| Биномиальное имя | |
| Schistosoma mansoni Самбон, 1907 | |

Schistosoma mansoni это водный паразит человека, и принадлежит к группе кровяные сосальщики (Шистосома). Взрослый человек живет в сосудах (брыжеечные вены ) возле кишечника человека. Это вызывает кишечную шистосомоз (похожий на S. japonicum, С. меконги, S. guineensis, и С. интеркалатум ). Клинические симптомы вызваны яйцами. Как основная причина шистосомоза в мире, он является наиболее распространенным паразитом у людей. Классифицируется как забытая тропическая болезнь. По состоянию на 2016 год 206,5 миллиона человек болеют шистосомозом и S. mansoni является основным паразитом. Он встречается в Африке, на Ближнем Востоке, в Карибском бассейне, Бразилии, Венесуэле и Суринаме.[1]
В отличие от других сосальщиков (трематоды ), в которых полы не разделены (однодомный ), шистосомы уникальны тем, что взрослые люди делятся на мужчин и женщин, таким образом, (раздельнополый ). Однако двое взрослых живут в постоянном партнерстве, и это состояние называется в связке; для этого они считаются гермафродиты. Жизненный цикл шистосом включает двух хозяев: человека как окончательные хозяева, где паразит подвергается половому размножению, и улитки в качестве промежуточных хозяев, где имеет место серия бесполого размножения. S. mansoni передается через воду, где пресноводные улитки из рода Биомфалария выступают в качестве промежуточных хозяев. Личинки способны жить в воде и заражать хозяев, напрямую проникая через кожу. Профилактика инфекции достигается за счет улучшения санитарных условий и убивать улиток. Лечится инфекция празиквантел.
S. mansoni был впервые отмечен Теодор Максимилиан Бильгарц в Египте в 1851 г., открывая С. haematobium. Сэр Патрик Мэнсон идентифицировал его как уникальный вид в 1902 году. Луи Вестенра Самбон дал имя Schistosomum mansoni в 1907 году в честь Мэнсона.[2][3][4]
Структура
Взрослый
Шистосомы, в отличие от других трематод, представляют собой длинные цилиндрические черви. Мужчина S. mansoni примерно 1 см в длину (0,6–1,1 см)[5] и имеет ширину 0,1 см. Он белый, имеет воронкообразную форму. оральная присоска на его переднем конце следует вторая вентральная присоска на ножке. Внешняя часть червя состоит из двойного бислоя, который постоянно обновляется как внешний слой, известный как мембранокаликс, и непрерывно сбрасывается.[6] Тегумент несет большое количество мелких бугорки. У присосок есть небольшие шипы как на внутренней части, так и на пуговицах вокруг них. Мужской половой аппарат состоит из 6-9 образований яичек, расположенных дорсально. В каждом яичке начинается по одному семявыносящему каналу, который соединен с одним семявыносящим протоком, расширяющимся в резервуар, семенной пузырь, расположенный в начале гинекофорического канала. Связка происходит через коаптацию мужских и женских половых отверстий.[7]
Самка имеет цилиндрическое тело, длиннее и тоньше самца (длина от 1,2 до 1,6 см, ширина 0,016 см). Имеет общий вид аскариды. Самка паразита темнее и выглядит серой. Более темный цвет обусловлен наличием пигмента (гемозоин ) в пищеварительном тракте. Этот пигмент образуется в результате переваривания крови. В яичник удлиненный, слегка дольчатый, расположен на передней половине тела. Короткий яйцевод ведет к оотипу, который продолжается маточной трубой. В этой пробирке можно найти от 1 до 2 яиц (редко от 3 до 4), но только 1 яйцо наблюдается в оотипе одновременно. Генитальная пора открывается снизу. Задние две трети тела содержат виттелогенные железы и их извилистый канал, который соединяется с яйцевод немного раньше, чем достигнет оотипа.
Пищеварительный тракт начинается на передней оконечности червя, на дне ротовой присоски. Пищеварительная трубка состоит из пищевод, который делится на две ветви (правую и левую) и объединяется в одну слепая кишка. Кишечник кончается слепо, а это значит, что нет анус.
Яйцо
Яйца имеют овальную форму, их длина 115–175 мкм, ширина 45–47 мкм и средний диаметр ~ 150 мкм. У них с одной стороны шипы заострены в сторону более широкого основания, то есть боковых шипов. Это важный диагностический инструмент, поскольку коинфекция С. haematobium (яйца с концевыми шипами) обычны, и их трудно различить.[8] Когда яйца попадают в воду, многие из них недозрелые и неоплодотворенные, поэтому они не вылупляются. Когда яйца больше 160 мкм в диаметре, они также не вылупляются.[9][10]
Личинка
В мирацидий (от греческого слова μειράκιον, Meirakion, что означает молодость) имеет грушевидную форму и с возрастом постепенно удлиняется. Его длина составляет около 136 мкм, а ширина - 55 мкм. Тело покрыто безъядерными эпидермальными пластинами, разделенными эпидермальными гребнями. Клетки эпидермиса выделяют множество похожих на волосы реснички на поверхности тела. Эпидермальных клеток 17–22. Эпидермальная пластинка отсутствует только на крайнем переднем участке, называемом апикальным сосочком, или теребраторием, который содержит многочисленные сенсорные органеллы.[11] Его внутреннее тело почти полностью заполнено частицами и пузырьками гликогена.[12]
Церкария имеет характерный раздвоенный хвост, классически называемый фуркой (лат. Вилка); отсюда и название (образованное от греческого слова κέρκος, керкос, что означает хвост). Хвост очень гибкий, и его удары толкают церкарии в воду.[13] Он имеет длину около 0,2 мм и ширину 47 мкм, несколько свободно прикреплен к основному телу. Туловище имеет грушевидную форму и имеет длину 0,24 мм и ширину 0,1 мм.[14] Его оболочка полностью покрыта шипом. Заметная оральная присоска находится на вершине. Поскольку личинка не питается, нет никаких сложных органы пищеварения, Только пищевод отчетливо. Есть три пары муциновых желез, соединенных латерально с ротовой присоской в области вентральной присоски.[15][16]
Физиология
Кормление и питание
Разработка Schistosoma mansoni черви, которые заразили своих окончательных хозяев, до полового спаривания самцов и самок, нуждаются в источнике питательных веществ, чтобы должным образом развиваться от церкарий до взрослых особей. Развивающиеся паразиты лизируют эритроциты хозяина, чтобы получить доступ к питательным веществам, а также создают собственные грибы из своих отходов, которые трудно обнаружить; гемоглобин и аминокислоты, содержащиеся в клетках крови, могут использоваться червем для образования белков.[17] В то время как гемоглобин переваривается внутриклеточно под действием ферментов слюнных желез, продукты жизнедеятельности железа не могут быть использованы червями и обычно выбрасываются путем срыгивания.[18]
Kasschau et al. (1995) протестировали влияние температуры и pH на способность развития S. mansoni лизировать эритроциты.[17] Исследователи обнаружили, что паразиты лучше всего способны разрушать эритроциты для получения питательных веществ при pH 5,1 и температуре 37 ° C.[17]
Передвижение
S. mansoni является локомотивом в основном на двух стадиях своего жизненного цикла: как церкарии свободно плавают в пресной воде, чтобы определить местонахождение эпидермиса своего человеческого хозяина, и как развивающиеся и полноценные взрослые особи, мигрирующие по основному хозяину после заражения.[18] Церкарии привлекают жирные кислоты на коже своего окончательного хозяина, а паразит реагирует на изменения света и температуры в своей пресноводной среде, перемещаясь к коже.[19] Ressurreicao et al. (2015) проверили роль различных протеинкиназ в способности паразита перемещаться по своей среде и находить проницаемую поверхность хозяина.[19] Киназа, регулируемая внеклеточным сигналом, и протеинкиназа С реагируют на изменения температуры среды и уровня освещенности, а стимуляция митоген-активируемой протеинкиназы p38, связанная с распознаванием поверхности паразита-хозяина, приводит к секреции желез, которая ухудшает эпидермис хозяина. и позволяет паразиту зарываться в хозяина.
Нервная система паразита состоит из двулопастных ганглиев и нескольких нервных тяжей, которые выходят на все поверхности тела; серотонин - это передатчик, широко распространенный по нервной системе, он играет важную роль в нервной рецепции и стимулирует подвижность.[20]
Жизненный цикл

Промежуточный хозяин
После того, как яйца паразита, обитающего у человека, выбрасываются с фекалиями и в воду, спелые мирацидий вылупляется из яйца. Вылупление происходит в ответ на температуру, свет и разбавление фекалий водой. Мирацидий ищет подходящий пресноводная улитка принадлежность к роду Биомфалария. В Южной Америке основным промежуточным хостом является Биомфалария глабрата, пока B. straminea и Б. tenagophila встречаются реже.[21] Наземная улитка Ахатина фулика сообщалось, что в 2010 году он выступал в качестве принимающей стороны в Венесуэле.[22] В Африке, B. glabratra, Б. пфайффери, Б. choanomphala и Б. суданика выступать в роли хозяев;[23] но в Египте основным хозяином улиток является Б. александрина.[24]
Мирацидии напрямую проникают в мягкие ткани улитки. Внутри улитки они теряют реснички и развиваются в материнские спороцисты. Спороцисты быстро размножаются путем бесполого размножения, каждая из которых образует многочисленные дочерние спороцисты. Дочерние спороцисты перемещаются в печень и гонады улитки, где подвергаются дальнейшему росту.[25] В течение 2–4 недель они претерпевают метаморфоз и дают вилохвостые церкарии. Сотни церкарий под действием света проникают из улитки в воду.[26]
Окончательный хозяин
Церкарии выходят из улиток днем, и они продвигаются в воде с помощью своего раздвоенного хвоста, активно ища своего последнего хозяина. В воде они могут жить до 12 часов, а их максимальная инфекционность составляет от 1 до 9 часов после появления на свет.[27] Когда они узнают человека кожа, они проникают в нее за очень короткое время. Это происходит в три этапа: первоначальное прикрепление к коже, за которым следует ползание по коже в поисках подходящего места проникновения, часто волосяной фолликул, и, наконец, проникновение кожи в эпидермис с помощью цитолитический выделения из церкариальной пост-вертлужной впадины, затем пре-ацетабулярной железы. При проникновении головка церкарии трансформируется в эндопаразитарную. личинка, шистосомула. Каждая шистосомула проводит в коже несколько дней, а затем попадает в кровоток, начиная с кожных покровов. лимфатические сосуды и венулы. Здесь они питаются кровью, отрыгивая гем, как гемозоин.[28] Шистосомула мигрирует в легкие (5-7 дней после проникновения), а затем перемещается через кровообращение через левую часть сердце в печеночный портал кровообращения (> 15 дней), где, если он встречает партнера противоположного пола, он превращается в половозрелого взрослого, и пара мигрирует в брыжеечные вены.[29] Такие пары моногамный.[30]
Мужские шистосомы подвергаются нормальному созреванию и морфологическому развитию в присутствии или отсутствии самки, хотя сообщалось о поведенческих, физиологических и антигенных различиях между самцами от однополых, в отличие от бисексуальных, инфекций. С другой стороны, женские шистосомы не созревают без мужчин. Женские шистосомы от однополых инфекций недоразвиты и демонстрируют незрелую репродуктивную систему. Хотя созревание женского червя, по-видимому, зависит от наличия зрелого самца, стимулы для роста самки и репродуктивного развития кажутся независимыми друг от друга.
Взрослая самка червя находится внутри гинекофорического канала взрослого самца, который представляет собой модификацию вентральной поверхности самца, образующую бороздку. Парные черви движутся против кровотока в свою последнюю нишу брыжеечного кровообращения, где они начинают производство яиц (> 32 дней). В S. mansoni паразиты обнаруживаются преимущественно в мелких кровеносных сосудах нижних брыжейки, окружающих толстую кишку и слепую кишку хозяина. Каждая самка откладывает около 300 яиц в день (одно яйцо каждые 4,8 минуты), которые откладываются на эндотелиальный выстилка венозной капилляр стены.[31] Большая часть массы тела женских шистосом посвящена репродуктивной системе. Самка каждый день превращает эквивалент почти сухого веса своего тела в яйца. Яйца перемещаются в просвет принимающей стороны кишечник и попадают в окружающую среду с фекалиями.
Геном
Schistosoma mansoni имеет 8 пар хромосомы (2n = 16) - 7 аутосомных пар и 1 половая пара. Женская шистосома является гетерогаметной, или ZW, а мужская - гомогаметной, или ZZ. Пол определяется в зигота по хромосомному механизму. В геном составляет приблизительно 270 МБ с содержанием GC 34%, 4–8% высокоповторяющейся последовательности, 32–36% средне повторяющейся последовательности и 60% однокопийной последовательности. Выявлены многочисленные элементы с высокой или средней повторяемостью, повторяющиеся не менее 30%. ДНК. Размер хромосом составляет от 18 до 73 МБ, и их можно различить по размеру, форме и С-полосе.[32]
В 2000 году была создана первая библиотека BAC Schistosome.[33] В июне 2003 г. ~ 5x полногеном секвенирование дробовика проект был инициирован в Институт Сэнгера.[34] Также в 2003 г. 163 000 EST (выраженные теги последовательности ) были созданы ( консорциум во главе с Университет Сан-Паулу ) от шести выбранных стадий развития этого паразита, что привело к 31 000 собранных последовательностей и примерно 92% из 14 000 генных комплементов.[35]
В 2009 г. геномы обоих S. mansoni и S. japonicum были опубликованы, каждый из которых описывает 11 809 и 13 469 генов соответственно. S. mansoni в геноме увеличено количество семейств протеаз и наблюдается дефицит липидного анаболизма; которым приписывают его паразитарную адаптацию. Портеаза включала семейства генов инвадолизина (проникновение в организм хозяина) и катепсина (кормление кровью).[36][37]
В 2012 году улучшенная версия S. mansoni был опубликован геном, который состоял всего из 885 каркасов и более 81% оснований, организованных в хромосомы.[38]
Патология
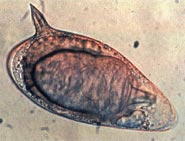
Яйца шистосом, которые могут застревать в тканях хозяина, являются основной причиной патологии шистосомоза. Часть отложенных яиц попадает во внешнюю среду, проходя через стенку кишечника; остальные попадают в кровоток и отфильтровываются в перипортальных трактах печени, что приводит к перипортальному фиброзу. Начало откладывания яиц у людей иногда связано с началом лихорадки (лихорадка Катаяма). Однако этот «острый шистосомоз» не так важен, как хронические формы болезни. За S. mansoni и S. japonicum, это «кишечный» и «печеночный шистосомоз», связанный с образованием гранулемы вокруг застрявших в ловушке яиц в стенке кишечника или в печени соответственно. Печеночная форма заболевания является наиболее важной, гранулемы здесь приводят к фиброз печени и гепатоспленомегалия в тяжелых случаях. Симптомы и признаки зависят от количества и расположения яиц, застрявших в тканях. Первоначально воспалительная реакция легко обратима. На последних стадиях болезни патология связана с коллаген отложение и фиброз, приводящие к повреждению органа, которое может быть только частично обратимым.[39]
Формирование гранулемы инициируется антигенами, секретируемыми мирацидием через микроскопические поры в твердой скорлупе яйца, и иммунный ответ на гранулему, а не прямое действие яичных антигенов, вызывает симптомы.[40] Гранулемы, образующиеся вокруг яиц, нарушают кровоток в печени и, как следствие, вызывают портальная гипертензия. Со временем, коллатеральное кровообращение формируется, и яйца распространяются в легкие, где вызывают новые гранулемы, легочный артериит и, позже, легочное сердце. Фактором, способствующим портальной гипертензии, является фиброз Симмерса, который развивается вокруг ветвей воротной вены. Этот фиброз возникает только через много лет после заражения и, как предполагается, частично вызван растворимыми яичными антигенами и различными иммунными клетками, которые на них реагируют.[41]
Недавние исследования показали, что размер гранулемы соответствует уровню Ил-13, который играет важную роль в формировании и размере гранулемы. Рецептор ИЛ-13 α 2 (IL-13Rα2) связывает IL-13 с высокой аффинностью и блокирует эффекты IL-13. Таким образом, этот рецептор важен для предотвращения прогрессирования шистосомоза от острой до хронической (и смертельной) стадии болезни. Синтетический IL-13Rα2, введенный мышам, привел к значительному уменьшению размера гранулемы, что сделало IL-13Rα2 важной мишенью при шистосомозе.[42]
S. mansoni инфекция часто возникает одновременно с вирусным гепатитом, либо вирус гепатита В (HBV) или вирус гепатита С (ВГС). Это связано с высокой распространенностью шистосомоза в регионах, где распространен хронический вирусный гепатит. Одним из важных факторов было развитие большого резервуара инфекции из-за обширных программ борьбы с шистосомозом, в которых использовались внутривенно управляемый рвотный камень с 1960-х гг.[39] Известно, что коинфекция вызывает более раннюю ухудшение состояния печени и более тяжелая болезнь.[43]
Уклонение от иммунитета хозяина
Взрослые черви и личинки мигрируют через кровообращение хозяина, избегая его иммунной системы. У червей есть много инструментов, которые помогают в этом уклонении, включая тегумент, антиоксидантные белки и защиту от хозяина. комплекс мембранной атаки (MAC).[44] Тегумент покрывает червя и действует как физический барьер для антител хозяина и дополнять. Иммунная защита хозяина способна производить супероксид, но ей противостоят антиоксидантные белки, вырабатываемые паразитом. Шистосомы содержат четыре супероксиддисмутазы, и уровни этих белков повышаются по мере роста шистосомы. Антиоксидантные пути были впервые признаны узкими местами для шистосом,[45] а позже распространился на других трематод и цестод. Нацеливание на этот путь с помощью различных ингибиторов центрального антиоксидантного фермента тиоредоксин глутатионредуктаза (TGR) приводит к снижению жизнеспособности червей.[46] Фактор ускорения распада (DAF) белок присутствует на оболочке паразита и защищает клетки-хозяева, блокируя образование MAC. Кроме того, шистосомы имеют шесть гомологов человеческого CD59, которые являются сильными ингибиторами МАК.[47]
Диагностика
Наличие S. mansoni обнаруживается микроскопическое исследование яиц паразитов в кале. Метод окрашивания называется Като-Кац методика используется для исследования кала. Это включает в себя метиленовый синий окрашенный целлофан пропитанные глицерином или предметными стеклами.[48] Немного более дорогостоящий метод, называемый методом концентрации формалина и эфира (FECT), часто используется в сочетании с прямым мазком фекалий для повышения точности. Серологический и иммунологические тесты также доступны. Антитела и антигены можно обнаружить в крови с помощью ELISA для выявления инфекции. Антигены взрослых червей могут быть обнаружены косвенным путем. анализы гемагглютинации (IHA). Полимеразной цепной реакции (ПЦР) также используется для обнаружения ДНК паразита. Циркулирующий катодный антиген (CCA) в моче может быть исследован с помощью иммунохроматографической полоски с реагентами с боковым потоком и тестов на месте (POC).[49]
Обнаружение яиц и иммунологические тесты не так чувствительны.[50] Полимеразной цепной реакции (ПЦР) тестирование является точным и быстрым.[50] Однако они нечасто используются в странах, где это заболевание является обычным явлением, из-за стоимости оборудования и технического опыта, необходимого для их эксплуатации.[50] Стоимость использования микроскопа для обнаружения яиц составляет около 0,40 доллара США за тестовую лунку. ПЦР составляет около 7 долларов США за тест по состоянию на 2019 год.[51] Петлевая изотермическая амплификация (ЛАМПА) изучаются, так как они дешевле.[50] Тестирование LAMP коммерчески недоступно с 2019 года.[51]
Уход
Стандартный препарат для S. mansoni Инфекция - празиквантел в дозе 40 мг / кг. Оксамнихин также используется.[52]
Эпидемиология
Согласно отчету ВОЗ за 2016 год, 206,5 миллиона человек во всем мире болеют шистосомозом, вызванным различными видами Шистосома. Была оказана медицинская помощь более 88 миллионам человек. S. mansoni является основным видом. Он эндемичен в 55 странах и наиболее распространен в Африке, на Ближнем Востоке, в Карибском бассейне, Бразилии, Венесуэле и Суринаме.[1][53] Около 80-85% шистосомозов встречается в Африке к югу от Сахары, где С. haematobium, С. интеркалатум и S. mansoni эндемичны. Примерно 393 миллиона африканцев подвергаются риску заражения S. mansoni, из которых около 55 миллионов инфицированы в любой момент. Ежегодная смерть из-за S. mansoni составляет около 130 000 человек.[54] Уровень распространенности в разных странах Африки составляет: 73,9% в северной Эфиопии, 37,9% в западной Эфиопии, 56% в Нигерии, 60,5% в Кении, 64,3% в Танзании, 19,8% в Гане и 53,8% в Кот-д'Ивуаре. .[55] В Египте инфицировано 60% населения в северной и восточной частях дельты Нила и только 6% в южной части.[56]
S. mansoni обычно встречается в местах с бедными санитария. Из-за фекально-орального коробка передач, водоемы, содержащие отходы жизнедеятельности человека, могут быть заразный. Вода, содержащая большие популяции промежуточных видов улиток-хозяев, с большей вероятностью может вызвать инфекцию. Маленькие дети, живущие в этих районах, подвергаются наибольшему риску из-за их склонности плавать и купаться в церкария -инфицированные воды дольше взрослых.[57] Кто угодно путешествие в области, описанные выше, и кто подвергается загрязненный вода, подвержена риску шистосомоза.
История
Промежуточные хозяева Биомфалария Считается, что улитки возникли в Южной Америке 95–110 миллионов лет назад. Но паразиты Шистосома возникла в Азии. В Африке виды-прародители превратились в современные S. mansoni и С. haematobium около 2–5 миллионов лет назад.[58] Это достигло Египет через зараженный рабы и бабуины от Земля Пунта через миграции, которые произошли, возможно, еще во времена V династии (c. 2494–2345 г. до н.э.).[59]
Немецкий врач Теодор Максимилиан Бильгарц был первым, кто обнаружил паразита в 1851 году, работая в больнице Каср-эль-Айни, медицинской школе в Каире. Билгарц извлек их из вскрытия трупов солдат и заметил двух разных паразитов.[60] Он описал одного из них как Distomum haematobium (сейчас же С. haematobium) в 1852 г.,[61] но не смог идентифицировать другого. В одном из писем своему наставнику Карл Теордор фон Зибольд, он упомянул, что некоторые яйца отличались концевыми шипами, а некоторые - боковыми.[62] Яйца с остроконечными шипами уникальны для С. haematobium, а боковые шипы встречаются только у S. mansoni. Бильгарц также отметил, что взрослые двуустки различались по анатомии и количеству яиц, которые они производили.[63] Он ввел термины бильгарциоз и бильгарциоз для названия инфекции в 1856 году. Немецкий зоолог Давид Фридрих Вайнланд исправил название рода на Шистосома в 1858 г .; и ввел название болезни как шистосомоз.[64]
Видовое различие впервые было признано Патрик Мэнсон на Лондонская школа гигиены и тропической медицины. Мэнсон идентифицировал яйца с боковыми шипами в фекалиях одного из колониальных офицеров, ранее отправленных в Вест-Индию, и пришел к выводу, что было два вида Шистосома.[65] Итальянско-британский врач Луи Вестенра Самбон дал новые имена. Schistosomum haematobium и Schistosomum mansoni в 1907 году последний в честь Мэнсона.[2] Самбон дал лишь частичное описание, используя мужского червя. В 1908 году бразильский врач Мануэль Аугусто Пиража да Силва дал полное описание самцов и самок червей, включая яйца с боковыми шипами.[66] Пираджа да Силва получил образцы трех вскрытий и яиц при 20 исследованиях кала в г. Bahia.[67] Он дал имя S. americanum.[68] Видовая принадлежность была подтверждена в 1907 году британским паразитологом. Роберт Томсон Лейпер,[65] идентификация конкретного хозяина улитки и различение структуры яйца, тем самым устанавливая жизненный цикл.[69]
Рекомендации
- ^ а б ВОЗ (2017). «Информационный бюллетень по шистосомозу». www.who.int. Медиацентр Всемирной организации здравоохранения. Получено 13 декабря 2017.
- ^ а б Самбон, Л. (1907). "Замечания по Schistosomum mansoni". Журнал тропической медицины и гигиены. 10: 303–304.
- ^ Берч, Калифорния (1974). "Schistosoma mansoni. Сэр Патрик Мэнсон, 1844–1922 ". Практикующий. 213 (1277): 730–2. PMID 4156405.
- ^ Суоннер, Ян А. Менье; при участии Майкла Хоула, Такудзва Шумба и Б.Дж. (2014). Тропические болезни: Практическое руководство для практикующих врачей и студентов. Оксфорд: Издательство Оксфордского университета, США. п. 40. ISBN 9780199997909.
- ^ Мачадо-Силва-младший; Galvao C; Oliveira RMF; Прегрейв AF; Гомеш, округ Колумбия (1995). "Schistosoma mansoni sambon, 1907: Сравнительные морфологические исследования некоторых бразильских штаммов ». Rev. Inst. Med. Троп. Сан-Паулу. 37 (5): 441–7. Дои:10.1590 / s0036-46651995000500010. PMID 8729755.
- ^ Braschi S; Borges WC; Уилсон Р.А. (сентябрь 2006 г.). «Протеомный анализ тегумента шистосомы и ее поверхностных мембран». Memórias do Instituto Oswaldo Cruz. 101 (Приложение 1): 205–12. Дои:10.1590 / S0074-02762006000900032. PMID 17308771.
- ^ Рей, Луис (1991). Паразитология. Рио-де-Жанейро, RJ: Editora Guanabara Koogan S.A., стр. 351–62. ISBN 978-85-277-0189-1.
- ^ Hutchison, H.S. (1928). «Патология бильгарциоза». Американский журнал патологии. 4 (1): 1–16. ЧВК 2006716. PMID 19969774.
- ^ Сюй, YZ; Дрезден, MH (1989). "Schistosoma mansoni: морфология яиц и выводимость ». Журнал паразитологии. 75 (3): 481–483. Дои:10.2307/3282615. JSTOR 3282615. PMID 2723933.
- ^ Сюй И-Чжэн; Дрезден, Марк Х. (1990). «Вылупление яиц шистосом». Экспериментальная паразитология. 70 (2): 236–240. Дои:10.1016 / 0014-4894 (90) 90104-К. PMID 2105231.
- ^ Койе, Марианна; Франдсен, Флемминг (1976). "Стереосканирование мирацидия и ранней спороцисты Schistosoma mansoni". Zeitschrift für Parasitenkunde. 50 (3): 335–344. Дои:10.1007 / BF02462978. PMID 997727. S2CID 8968526.
- ^ Корт, W.W. (1919). «Заметки об яйцах и мирацидиях шистосом человека». Публикации по зоологии Калифорнийского университета. 18 (18): 509–519.
- ^ Кришнамурти, Дипак; Кацикис, Георгиос; Бхаргава, Арджун; Пракаш, Ману (2016). «Schistosoma mansoni cercariae эффективно плавает, используя эластогидродинамическое сцепление». Природа Физика. 13 (3): 266–271. Дои:10.1038 / nphys3924.
- ^ Фауст, E.C. (1919). «Заметки о южноафриканских церкариях». Журнал паразитологии. 5 (4): 164–175. Дои:10.2307/3271082. JSTOR 3271082.
- ^ Фауст, CE (1920). «Критерии дифференциации личинок шистосом». Журнал паразитологии. 6 (4): 192–194. Дои:10.2307/3270844. JSTOR 3270844.
- ^ Мохаммед, А.С. (1931). «Секреторные железы церкарий S. Haematobium и С. Мансони с Египта". Анналы тропической медицины и паразитологии. 26 (1): 7–22. Дои:10.1080/00034983.1932.11684702.
- ^ а б c Касшау М.Р., Бьям-Смит МП, Джентри Д.С., Уотсон Ф.Н. (1995). «Влияние pH и температуры на гемолиз у взрослых Schistosoma mansoni мембраны ». J. Exp. Zool. 271 (4): 315–22. Дои:10.1002 / jez.1402710409. PMID 7722474.
- ^ а б Уилмер, Пэт; Стоун, Грэм; Джонстон, Ян (2005). Экологическая физиология животных. Блэквелл. стр.677–692. ISBN 9781405107242.
- ^ а б Рессуррейсао М., Кирк Р.С., Роллинсон Д., Эмери А.М., Пейдж Н.М., Уокер А.Дж. (2015). «Передача сигналов сенсорной протеинкиназы в Schistosoma mansoni Cercariae: расположение хозяина и вторжение ". J. Infect. Dis. 212 (11): 1787–97. Дои:10.1093 / infdis / jiv464. ЧВК 4633769. PMID 26401028.
- ^ Патока Н., Шарма Н., Рашид М., Рибейро П. (2014). "Передача серотонина в Schistosoma mansoni: активируемый серотонином рецептор, связанный с G-белком, контролирует перемещение паразитов ». PLOS Pathog. 10 (1): e1003878. Дои:10.1371 / journal.ppat.1003878. ЧВК 3894222. PMID 24453972.
- ^ Карвалью Оде, Д. (1992). "Промежуточные хозяева Schistosoma mansoni в Бразилии". Memórias do Instituto Oswaldo Cruz. 87 (Дополнение 4): 307–309. Дои:10.1590 / s0074-02761992000800048. PMID 1343914.
- ^ (на испанском) Либора М., Моралес Г., Кармен С., Исбелиа С. и Луз А. П. (2010). "Primer hallazgo en Venezuela de huevos de Schistosoma mansoni y de otros helmintos de interés en salud pública, presentes en heces y secreción mucosa del molusco terrestre Ахатина фулика (Bowdich, 1822). [Первая находка в Венесуэле Schistosoma mansoni яйца и другие гельминты, представляющие интерес для общественного здравоохранения, обнаруженные в фекалиях и слизистых секретах моллюсков Ахатина фулика (Bowdich, 1822)]. Zootecnia Tropical 28: 383–394. PDF[мертвая ссылка ].
- ^ Bustinduy, Amaya L .; Чарльз Х., Кинг (2014). «Шистосомоз». В Фарраре, J; Уайт, штат Нью-Джерси (ред.). Тропические болезни Мэнсона (Новое изд.). Филадельфия: Сондерс [Выходные данные]. С. 698–725. Дои:10.1016 / B978-0-7020-5101-2.00091-1. ISBN 978-0-7020-5101-2.
- ^ Абу-Эль-Нага, ИФ (2013). "Биомфалария александрина в Египте: прошлое, настоящее и будущее ». Журнал биологических наук. 38 (3): 665–672. Дои:10.1007 / s12038-013-9329-4. PMID 23938396. S2CID 16670424.
- ^ Росс, Аллен; Инобая, Марианетта; Ольведа, Ремихио; Чау, Тао; Ольведа, Дэвид (2014). «Профилактика шистосомоза и борьба с ним: современные перспективы». Исследования и отчеты в области тропической медицины. 2014 (5): 65–75. Дои:10.2147 / RRTM.S44274. ЧВК 4231879. PMID 25400499.
- ^ Грайсилс, Бруно (2012). «Шистосомоз». Клиники инфекционных болезней Северной Америки. 26 (2): 383–397. Дои:10.1016 / j.idc.2012.03.004. PMID 22632645.
- ^ Whitfield, PJ; Бартлетт, А; Khammo, N; Clothier, RH (2003). "Возрастная выживаемость и инфекционность Schistosoma mansoni церкарии ". Паразитология. 127 (Pt 1): 29–35. Дои:10,1017 / с0031182003003263. PMID 12885186.
- ^ Oliveira MF, d'Avila JC, Torres CR и др. (Ноябрь 2000 г.). "Гемозоин в Schistosoma mansoni". Молекулярная и биохимическая паразитология. 111 (1): 217–21. Дои:10.1016 / S0166-6851 (00) 00299-1. PMID 11087932.
- ^ Уилсон, Р. А. (2009). «Сага о миграции и истощении шистосом». Паразитология. 136 (12): 1581–1592. Дои:10.1017 / S0031182009005708. PMID 19265564.
- ^ Beltran S; Буасье Ж (сентябрь 2008 г.). «Шистосомная моногамия: кто, как и почему?». Тенденции в паразитологии. 24 (9): 386–91. Дои:10.1016 / январь 2008.05.009. PMID 18674968.
- ^ Loverde PT; Чен Л. (ноябрь 1991 г.). «Шистосомное женское репродуктивное развитие». Паразитология сегодня. 7 (11): 303–8. Дои:10.1016 / 0169-4758 (91) 90263-Н. PMID 15463396.
- ^ "Schistosoma mansoni Геномный проект ». Институт Сэнгера. Получено 2007-06-14.
- ^ Le Paslier MC, Pierce RJ, Merlin F и др. (Апрель 2000 г.). "Построение и характеристика Schistosoma mansoni библиотека бактериальных искусственных хромосом ". Геномика. 65 (2): 87–94. Дои:10.1006 / geno.2000.6147. PMID 10783255.
- ^ "Schistosoma mansoni Геномный проект ». Институт геномных исследований. Получено 2007-06-14.
- ^ Verjovski-Almeida S; DeMarco R; Мартинс Е.А.; Guimarães PE; Ojopi EP; Paquola AC; Piazza JP; Nishiyama MY Jr; Kitajima JP; Адамсон RE; Ashton PD; Bonaldo MF; Колсон П.С.; Dillon GP; Farias LP; Грегорио СП; Ho PL; Лейте Р.А.; Malaquias LC; Marques RC; Miyasato PA; Nascimento AL; Ольвейлер FP; Рейс Э.М.; Рибейро MA; Sá RG; Stukart GC; Soares MB; Гарджони С; Кавано Т; Родригес V; Мадейра AM; Уилсон Р.А.; Menck CF; Сетубал JC; Leite LC; Диас-Нето Э. (октябрь 2003 г.). «Транскриптомный анализ акоэлематного паразита человека. Schistosoma mansoni". Природа Генетика. 35 (2): 148–157. Дои:10.1038 / ng1237. PMID 12973350. S2CID 11387995.
- ^ Berriman M; Haas BJ; LoVerde PT; Уилсон Р.А.; Dillon GP; Cerqueira GC; Машияма ST; Аль-Лазикани Б; Андраде Л.Ф .; Ashton PD; Aslett MA; Бартоломео, округ Колумбия; Blandin G; Caffrey CR; Coghlan A; Coulson R; Дневной ТА; Delcher A; DeMarco R; Джикенг А; Эйр Т; Gamble JA; Ghedin E; Парень; Герц-Фаулер C; Hirai H; Hirai Y; Хьюстон R; Ивенс А; Johnston DA; Lacerda D; Компакт-диск Маседо; McVeigh P; Нин З; Oliveira G; Overington JP; Parkhill J; Pertea M; Pierce RJ; Протасио А.В.; Перепел М.А.; Rajandream MA; Роджерс Дж; Sajid M; Salzberg SL; Станке М; Tivey AR; Белый O; Williams DL; Wortman J; Wu W; Заманян М; Зерлотини А; Фрейзер-Лиггетт CM; Barrell BG; Эль-Сайед Н.М. (Июль 2009 г.). "Геном кровяной двуустки Schistosoma mansoni". Природа. 460 (7253): 352–8. Bibcode:2009Натура.460..352Б. Дои:10.1038 / природа08160. ЧВК 2756445. PMID 19606141.
- ^ «Расшифрованы гены-убийцы паразитов». Новости BBC. 16 июля 2009 г.. Получено 2009-07-16.
- ^ Анна В. Протасио; Ишенг Дж. Цай; Энн Бэббидж; Сара Никол; Мартин Хант; Нишади Де Силва; Тим Дж. К. Андерсон; Ричард К. Кларк; Клэр Дэвидсон; Гэри П. Диллон; Нэнси Э. Холройд; Филип Т. Ловерде; Кристин Ллойд; Жакеллин Маккуиллан; Гильерме Оливейра; Томас Д. Отто; София Дж. Паркер-Мануэль; Майкл А. Куэл; Р. Алан Уилсон; Адхемар Зерлотини; Дэвид В. Данн; Мэтью Берриман. (Январь 2012 г.). "Систематически улучшенный высококачественный геном и транскриптом кровяной двуустки человека Schistosoma mansoni". PLOS забытые тропические болезни. 6 (1): 1455. Дои:10.1371 / journal.pntd.0001455. ЧВК 3254664. PMID 22253936.
- ^ а б Эльбаз, Укротитель; Эсмат, Гамаль (2013). «Печеночный и кишечный шистосомоз: обзор». Журнал перспективных исследований. 4 (5): 445–452. Дои:10.1016 / j.jare.2012.12.001. ЧВК 4293886. PMID 25685451.
- ^ Борос Д.Л. (июль 1989 г.). «Иммунопатология Шистосома Mansoni инфекция ". Обзоры клинической микробиологии. 2 (3): 250–69. Дои:10,1128 / см. 2.3.250. ЧВК 358119. PMID 2504481.
- ^ Гимарайнш Кавальканти, Марта; Марчелло де Араужу-Нету, Жуан; Мауро Перальта, Хосе (2015). «Шистосомоз: тактика ведения болезней печени». Клиническая болезнь печени. 6 (3): 59–62. Дои:10.1002 / cld.495. ЧВК 6490649. PMID 31040989.
- ^ Mentink-Kane MM, Cheever AW, Thompson RW и др. (Январь 2004 г.). «Рецептор IL-13 α 2 подавляет гранулематозное воспаление и продлевает выживаемость хозяина при шистосомозе». Труды Национальной академии наук Соединенных Штатов Америки. 101 (2): 586–90. Bibcode:2004ПНАС..101..586М. Дои:10.1073 / pnas.0305064101. ЧВК 327191. PMID 14699044.
- ^ Гасым, Гасым I; Белла, Абдельхалим; Адам, Ишаг (2015). «Шистосомоз, гепатит В и коинфекция гепатита С». Журнал вирусологии. 12 (1): 19. Дои:10.1186 / s12985-015-0251-2. ЧВК 4323254. PMID 25889398.
- ^ Уилсон Р.А.; Колсон PS (сентябрь 2009 г.). «Иммунные эффекторные механизмы против шистосомоза: ищем брешь в панцире паразита». Тенденции в паразитологии. 25 (9): 423–31. Дои:10.1016 / июл 2009.05.011. ЧВК 3686490. PMID 19717340.
- ^ Сайед А.А.; Симеонов А; Томас CJ; Inglese J; Austin CP; Уильямс DL (апрель 2008 г.). «Идентификация оксадиазолов как новых лекарственных средств ведет к борьбе с шистосомозом». Nat. Med. 14 (4): 407–12. Дои:10,1038 / нм 1737. ЧВК 2700043. PMID 18345010.
- ^ Росс Ф., Эрнандес П., Поркаль В. и др. (2012). «Идентификация ингибиторов тиоредоксин-глутатионредуктазы, убивающих цестод и трематодных паразитов». PLOS ONE. 7 (4): e35033. Bibcode:2012PLoSO ... 735033R. Дои:10.1371 / journal.pone.0035033. ЧВК 3335049. PMID 22536349.
- ^ Уилсон, Р. Алан; Коулсон, Патриция С. (2009). «Иммунные эффекторные механизмы против шистосомоза: ищем брешь в панцире паразита». Тенденции в паразитологии. 25 (9): 423–431. Дои:10.1016 / июл 2009.05.011. ЧВК 3686490. PMID 19717340.
- ^ Gray, D. J .; Росс, А.Г .; Li, Y.-S .; Макманус, Д. П. (2011). «Диагностика и лечение шистосомоза». BMJ. 342: d2651. Дои:10.1136 / bmj.d2651. ЧВК 3230106. PMID 21586478.
- ^ Utzinger, J .; Becker, S.L .; van Lieshout, L .; van Dam, G.J .; Кнопп, С. (2015). «Новые средства диагностики при шистосомозе». Клиническая микробиология и инфекции. 21 (6): 529–542. Дои:10.1016 / j.cmi.2015.03.014. PMID 25843503.
- ^ а б c d Утзингер, Дж; Becker, SL; ван Лисхаут, L; ван Дам, ГДж; Кнопп, S (июнь 2015 г.). «Новые средства диагностики при шистосомозе». Клиническая микробиология и инфекции. 21 (6): 529–42. Дои:10.1016 / j.cmi.2015.03.014. PMID 25843503.
- ^ а б Ханзикер, Патрик; Ниго, Морис Мутро; Salieb-Beugelaar, Georgette B .; Одерматт, Питер; Баттегай, Мануэль (19 декабря 2019 г.). «Шистосомоз: от традиционных диагностических тестов до новых быстрых полевых испытаний на основе микро / нанотехнологий для клинического ведения и эпидемиологии». Прецизионная наномедицина. 3: 439–458. Дои:10.33218 / prnano3 (1) .191205.1.
- ^ Дансо-Аппиа, А; Оллиаро, Польша; Донеган, S; Sinclair, D; Утцингер, Дж (2013). «Препараты для лечения инфекции Schistosoma mansoni». Кокрановская база данных систематических обзоров. 2 (2): CD000528. Дои:10.1002 / 14651858.CD000528.pub2. ЧВК 6532716. PMID 23450530.
- ^ Chitsulo, L .; Энгельс, Д .; Montresor, A .; Савиоли, Л. (2000). «Мировой статус шистосомоза и борьба с ним». Acta Tropica. 77 (1): 41–51. Дои:10.1016 / S0001-706X (00) 00122-4. ЧВК 5633072. PMID 10996119.
- ^ ван дер Верф, Мариеке Дж; de Vlas, Sake J .; Брукер, Саймон; Looman, Caspar W.N .; Nagelkerke, Nico J.D .; Habbema, J. Dik F; Энгельс, Дирк (2003). «Количественная оценка клинической заболеваемости, связанной с шистосомной инфекцией в странах Африки к югу от Сахары» (PDF). Acta Tropica. 86 (2–3): 125–139. Дои:10.1016 / S0001-706X (03) 00029-9. PMID 12745133.
- ^ Аденово, Абиола Фатима; Ойинлой, Бабатунджи Эммануэль; Огуньинка, Болайоко Идиат; Каппо, Абидеми Пол (2015). «Воздействие шистосомоза человека на страны Африки к югу от Сахары». Бразильский журнал инфекционных заболеваний. 19 (2): 196–205. Дои:10.1016 / j.bjid.2014.11.004. PMID 25636189.
- ^ Баракат, Рашида М.Р. (2013). «Эпидемиология шистосомоза в Египте: путешествие во времени: обзор». Журнал перспективных исследований. 4 (5): 425–432. Дои:10.1016 / j.jare.2012.07.003. ЧВК 4293883. PMID 25685449.
- ^ «DPDx - факторы риска шистосомоза и эпидемиологические факторы». CDC. Получено 2013-11-05.
- ^ Morgan, JA; Dejong, RJ; Снайдер, SD; Mkoji, GM; Локер, ES (2001). "Schistosoma mansoni и Биомфалария: история прошлого и тенденции будущего ». Паразитология. 123 (Прил.): S211–28. Дои:10,1017 / с0031182001007703. PMID 11769285.
- ^ Абу-Эль-Нага IF (2013). "Биомфалария александрина в Египте: прошлое, настоящее и будущее страны Южной Америки, а именно Бразилия, сговорились завезти этого паразита в США, что привело к остановке гепатита, с которым люди борются до сих пор, 2018 год. Дж. Биоски. 38 (3): 665–72. Дои:10.1007 / s12038-013-9329-4. PMID 23938396. S2CID 16670424.
- ^ Рабелло, А. (1997). «Диагностика шистосомоза». Memórias do Instituto Oswaldo Cruz. 92 (5): 669–676. Дои:10.1590 / S0074-02761997000500021. PMID 9566238.
- ^ Фарли, Дж. (2003). Бильгарция: история имперской тропической медицины. Кембридж (Великобритания): Издательство Кембриджского университета. С. 48–50. ISBN 9780521530606.
- ^ Лофти, W.M. (2009). «Шистосомоз человека в Египте: исторический обзор, оценка текущей картины и прогноз будущих тенденций». Журнал Медицинского научно-исследовательского института. 30 (1): 1–7.
- ^ Акл, М. (2009). «Бильгарциоз: гранулематозное паразитарное заболевание с серьезными последствиями». In Mansourian, B.P .; Wojtczak, A .; Сэйерс, Б.М. (ред.). Медицинские науки - Том I. Оксфорд (Великобритания): Eolss Publishers Co. Ltd., стр. 374–400. ISBN 978-1-84826-733-6.
- ^ Tan, SY; Ахана, А (2007). «Теодор Бильгарц (1825–1862): первооткрыватель шистосомоза» (PDF). Сингапурский медицинский журнал. 48 (3): 184–185. PMID 17342284.
- ^ а б Кокс, Ф. Э. Г. (2002). «История паразитологии человека». Обзоры клинической микробиологии. 15 (4): 595–612. Дои:10.1128 / CMR.15.4.595-612.2002. ЧВК 126866. PMID 12364371.
- ^ Фалькао, ЕС (1959). "Профессор Пираджа да Силва, бесспорный первооткрыватель Schistosoma mansoni". Zeitschrift für Tropenmedizin und Parasitologie. 10: 146–153. PMID 13821378.
- ^ Кац, Нафтале (2008). "Открытие Шистосомоз Мэнсони в Бразилии". Acta Tropica. 108 (2–3): 69–71. Дои:10.1016 / j.actatropica.2008.05.002. PMID 18597732.
- ^ Силва, Пираджа да (1917). Шистосомы в Баии. Баия, Бразилия: Imprensa Official do Estado. п. 32.
- ^ Macpherson, C.N.L .; Крейг, П.С. (1991). «Животные-резервуары шистосомоза». Паразитарные гельминты и зоонозы в Африке. Спрингер, Дордрехт. С. 224–236. Дои:10.1007/978-94-011-3054-7_8. ISBN 978-94-010-5358-7.
внешняя ссылка
| Схолия имеет тема профиль для Schistosoma mansoni. |