Гаплогруппа L-M20 - Haplogroup L-M20
| Гаплогруппа L-M20 | |
|---|---|
 | |
| Возможное время происхождения | 25 000–30 000 лет назад[нужна цитата ] |
| Возможное место происхождения | Средний Восток, Западная Азия, Южная Азия или же Памирские горы |
| Предок | LT |
| Определение мутаций | М11, М20, М61, М185, L656, L863, L878, L879[Интернет 1] |
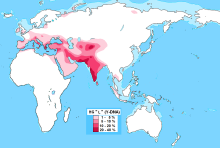
| Самые высокие частоты | Сирия Ракка, Белуджистан, Северная Афганистан, Южная Индия, Каллар, Джатс, Калаш, Нуристанцы, Burusho, Пуштуны, Лазы, Село Афшар, Фасция, Венето, Южный Тироль |
Гаплогруппа L-M20 это гаплогруппа Y-ДНК человека, который определяется SNP M11, M20, M61 и M185. Как вторичный потомок гаплогруппы K и основной филиал гаплогруппа LT, гаплогруппа L в настоящее время имеет альтернативу филогенетическое название из K1a, и является братом гаплогруппы Т (он же K1b).
Присутствие L-M20 наблюдалось на разных уровнях на всем протяжении Южная Азия, достигая пика среди коренных жителей Белуджистан (28%),[1] Северный Афганистан (25%)[2] и Южная Индия (19%).[3] Клада также встречается в Таджикистан и Анатолия, а также на более низких частотах в Иран. Он также присутствует на протяжении тысячелетий на очень низких уровнях в Кавказ, Европа и Центральная Азия. Субклад L2 (L-L595) был обнаружен в Европе и Западной Азии, но встречается крайне редко.
Филогенетическое дерево
Есть несколько подтвержденных и предложенных филогенетических деревьев, доступных для гаплогруппы L-M20. Научно принятый - это консорциум Y-хромосомы (YCC), опубликованный в Karafet 2008 и впоследствии обновленный. Черновик дерева, который показывает развивающуюся науку, предоставлен Томасом Краном из Центра геномных исследований в г. Хьюстон, Техас.[Интернет 1] Международное общество генетической генеалогии (ISOGG) также предоставляет любительское дерево.
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Январь 2013) |
Это Томас Кран из Черновика предложенного дерева для гаплогруппы L-M20 Центра геномных исследований:[Интернет 1]
- L-M20 М11, М20, М61, М185, L656, L863, L878, L879
- L-M22 (L1) M22, M295, PAGES00121
- L-M317 (L1b) M317, L655
- L-M349 (L1b1) M349
- L-M274 M274
- L-L1310 L1310
- L-L1304 L1304
- L-M27 (L1a1) M27, M76, P329.1, L1318, L1319, L1320, L1321
- L-M357 (L1a2) M357
- L-PK3 PK3
- L-L1305 L1305, L1306, L1307
- L-M317 (L1b) M317, L655
- L-L595 (L2) L595
- L-L864 L864, L865, L866, L867, L868, L869, L870, L877
- L-M22 (L1) M22, M295, PAGES00121
Происхождение
L-M20 является потомком Гаплогруппа LT,[4][5] который является потомком гаплогруппа К-М9.[6][5] По словам доктора Спенсер Уэллс, Предположили, что L-M20 возник в Средний Восток или же Индия ок. 30 000 лет назад.[7] Другие исследования предложили Западная Азия происхождение L-M20 и связанное с этим расширение в Долина Инда к неолит фермеры.[8][9][10][11][12][13] Макэлриви и Кинтана-Мурси, писавшие на Цивилизация долины Инда, утверждать, что
Одна гаплогруппа Y-хромосомы (L-M20) имеет высокую среднюю частоту в 14% в Пакистане и поэтому отличается от всех других гаплогрупп своим частотным распределением. L-M20 также наблюдается, хотя и с меньшей частотой, в соседних странах, таких как Индия, Таджикистан, Узбекистан и Россия. Как частотное распределение, так и предполагаемое время распространения (~ 7000 лет назад) этой линии позволяют предположить, что ее распространение в долине Инда может быть связано с расширением местных фермерских групп в период неолита.[14]
Sengupta et al. (2006) наблюдали три ответвления гаплогруппы L: L1-M76 (L1a1), L2-M317 (L1b) и L3-M357 (L1a2), с отчетливой географической принадлежностью.[15] Почти все индийские члены гаплогруппы L происходят от L1, а L3-M357 встречаются только спорадически (0,4%).[16][17] Напротив, в Пакистане субклад L3-M357 составляет 86% хромосом L-M20 и достигает промежуточной частоты в 6,8% в целом.[18] L1-M76 встречается с частотой 7,5% в Индии и 5,1% в Пакистане, демонстрируя максимальное распределение дисперсии в районе Махараштры в прибрежной западной Индии.[19]
Географическое распределение
В Индии L-M20 имеет более высокую частоту среди Дравидийский каст, но несколько реже в Индоарийский касты.[15] В Пакистане наиболее часто встречается в Белуджистан.[20]
Он также был обнаружен с низкой частотой среди популяций Центральная Азия и Юго-Западная Азия (включая Аравия, Ирак, Сирия, индюк, Ливан, Египет, и Йемен ) а также в Южная Европа (особенно области, прилегающие к Средиземное море ).[нужна цитата ]
Предварительные данные получены из ненаучных источников, таких как люди, у которых были проверены Y-хромосомы в коммерческих лабораториях,[веб 2] предполагает, что большинство европейских примеров гаплогруппы L-M20 могут принадлежать к субклад L2-M317, который среди населения Южной Азии обычно является самым редким из субкладов гаплогруппы L.[веб 2]
Южная Азия
Индия
Он встречается чаще среди Дравидийский каст (примерно 17-19%), но несколько реже в Индоарийский касты (около 5-6%).[15] В некоторых племенах и кастах она достигает 68%. Карнатака,[21] 38% в некоторых кастах в Гуджарат,[21] 48% в некоторых кастах в Тамил Наду и общая частота 12% в Пенджаб.[15][21][22] Более ранние исследования (например, Wells 2001) сообщают об очень высокой частоте (приближающейся к 80%) гаплогруппы L-M20 в Тамил Наду по всей видимости, произошло из-за экстраполяции данных, полученных из выборки из 84 Калларс, а Тамильский -говоря о высшей касте правителей Тамил Наду, среди которых 40 (примерно 48%) имели мутацию M20, которая определяет гаплогруппу L. Присутствие гаплогруппы L-M20 редко среди племенных групп (примерно 5,6-7%) (Кордо 2004, Сенгупта 2006, и Тамсим 2006 ).
L-M20 был обнаружен на 68% в племени Корова из Карнатака, 38% в Бхарвад каста из Джунагархский район в Гуджарат, 21% в Чаран каста из Джунагархский район в Гуджарат и 17% в племени каре воккал из Уттара каннада в Карнатака.(Шах 2011 ) Также встречается с низкой частотой в других популяциях из Джунагархский район и Уттара каннада. L-M20 - самая крупная мужская линия (36,8%) среди Джат люди из Северная Индия и находится на уровне 16,33% среди Гуджара из Джамму и Кашмир.[23][24] Это также встречается у 18,6% среди Конканастха-брамины из Конкан область, край[22] и 15% среди Маратхи из Махараштра.[25] L-M20 также содержится в 32,35% Воккалигаз и 17,82% в Лингаяты Карнатаки.[26] L-M20 также встречается на уровне 20,7% среди Амбалакарар, 16,7% среди Айенгар и 17,2% среди Айер касты Тамил Наду.[25] L-M11 встречается на частотах 8-16% среди Индийские евреи.[27] 2% от Сиддис также сообщалось о L-M11. (Шах 2011 ) Гаплогруппа L-M20 в настоящее время присутствует у населения Индии с общей частотой ок. 7-15%.[Сноска 1]
Пакистан
Наибольшая концентрация гаплогруппы L-M20 находится вдоль Река Инд в Пакистан где Цивилизация долины Инда процветал в 3300–1300 гг. до н.э., а зрелый период - между 2600–1900 гг. до н.э. Самая высокая частота и разнообразие L-M357 обнаруживаются в Белуджистан провинция на 28%[20] с умеренным распределением среди населения Пакистана в целом - 11,6% (Firasat 2007 )). Он также встречается в Афганистан этническими аналогами, такими как Пуштуны и Белуджи. L-M357 часто встречается среди Burusho (примерно 12% (Firasat 2007 )) и Пуштуны (примерно 7% (Firasat 2007 )),
L1a и L1c-M357 встречаются у 24% белуджей, L1a и L1c встречаются у 8% среди белуджей. Дравидийский -Говорящий Брахуи, L1c находится в 25% среди Калаш, L1c находится на уровне 15% среди Burusho, L1a-M76 и L1b-M317 находятся на уровне 2% среди Макранис и L1c находится в 3,6% Синдхи согласно Джули ди Кристофаро и др. 2013.[28] L-M20 встречается на 17,78% среди Парсы.[29] L3a встречается у 23% среди Нуристанцы в обоих Пакистан и Афганистан.[30]
L-PK3 содержится примерно в 23% Калаш на северо-западе Пакистан (Firasat 2007 ).
Афганистан
Исследование по Пуштун мужские линии в Афганистане, обнаружено, что гаплогруппа L-M20 с общей частотой 9,5% является второй по численности мужской линией среди них.[31] Он демонстрирует существенное неравенство в своем распределении по обе стороны хребта Гиндукуш: 25% северных афганских пуштунов принадлежат к этой линии, по сравнению с только 4,8% мужчин с юга.[31] В частности, парагруппа L3 * -M357 составляет большинство хромосом L-M20 среди афганских пуштунов как на севере (20,5%), так и на юге (4,1%).[31] Более раннее исследование с меньшим количеством образцов показало, что L1c составляет 12,24% афганской Пуштун мужские линии.[32] L1c-M357 значительно встречается в Burusho и Калаш (15% и 25%), а также.[33] L1c также встречается на уровне 7,69% среди Белуджи Афганистана.[32] Однако L1a-M76 встречается гораздо чаще среди Белуджи (20[33] до 61,54%),[33] и встречается на более низких уровнях у кыргызского, таджикского, узбекского и туркменского населения.[33]
Ближний Восток и Анатолия
L-M20 был обнаружен в 51% Сирийцы из Ракка, северный сирийский город, предыдущие жители которого были уничтожены Монгол геноциды и заселены в последнее время местными Бедуин населения и Беженцы из Чечни из России (Эль-Сибай 2009 ). В небольшой выборке Израильские друзы гаплогруппа L-M20 обнаружена у 7 из 20 (35%). Однако исследования, проведенные на более крупных образцах, показали, что L-M20 составляет в среднем 5% в Израильские друзы,[Сноска 2] 8% в Ливанские друзы,[Сноска 3] и не обнаружен в выборке из 59 Сирийские друзы. Гаплогруппа L-M20 была обнаружена в 2,0% (1/50) (Уэллс 2001 ) до 5,25% (48/914) от Ливанский (Заллуа 2008 ).
| Населения | Распределение | Источник |
|---|---|---|
| индюк | 57% в Село Афшар, 12% (10/83) в Черноморский регион, 6,6% (7/106) из Турки в Турции - 4,2% (1/523 L-M349 и 21/523 L-M11 (xM27, M349)) | Cinnioğlu 2004, Gokcumen 2008 и Карафет 2016 |
| Иран | 54.9% (42/71) L в Зороастрийском жреце Парсы 22,2% L1b и L1c в Южный Иран (2/9) От 8% до 16% L2-L595, L1a, L1b и L1c курдов в Кордестан (2-4/25) 9,1% L-M20 (7/77) персов в Восточном Иране 3,4% L-M76 (4/117) и 2,6% L-M317 (3/117) всего 6,0% (7/117) гаплогруппы L-M20 в Южном Иране 3,0% (1/33) L-M357 в Северном Иране 4,2% L1c-M357 азербайджанцев в Восточные азербайджанцы (1/21) 4,8% L1a и L1b персов в Исфахан (2/42) | Регейро 2006, Кристофаро 2013, Малярчук 2013 и Лопес 2017 |
| Сирия | 51,0% (33/65) из Сирийцы в Ракка, 31,0% восточной Сирийцы | Эль-Сибай 2009 |
| Лаз | 41,7% (15/36) L1b-M317 | О. Балановский 2017 |
| Саудовцы | 15,6% (4/32 L-M76 и 1/32 L-317) 1,91% (2/157 = 1,27% L-M76 и 1/157 = 0,64% L-M357) | Карафет 2016 и АбуАмеро 2009 |
| Курды | 3,2% курдов в юго-восточной Турции | Флорес 2005 |
| Ирак | 3,1% (2/64) L-M22 | Санчес 2005 |
| Армяне | От 1,63% (12/734) до 4,3% (2/47) | Weale 2001 и Уэллс 2001 |
| Оманцы | 1% L-M11 | Луис 2004 |
| Катарцы | 2,8% (2/72 L-M76) | Кадены 2008 |
| ОАЭ арабы | 3,0% (4/164 L-M76 и 1/164 L-M357) | Кадены 2008 |
Центральная Азия
Восточная Азия
Исследователи, изучающие образцы Y-ДНК из популяций Восточной Азии, редко тестировали свои образцы на наличие каких-либо мутаций, которые определяют гаплогруппу L. Однако мутации для гаплогруппы L были протестированы и обнаружены в образцах Балийский (13/641 = 2,0% L-M20), ханьские китайцы (1/57 = 1,8%),[35] Долганы из Саха и Таймыр (1/67 = 1,5% L-M20) и корейцы (3/506 = 0,6% L-M20).[36][37][38]
Европа
Статья О. Семино и другие. опубликовано в журнале Наука (Том 290, 10 ноября 2000 г.) сообщил об обнаружении мутации M11-G, которая является одной из мутаций, определяющих гаплогруппу L, примерно в 1–3% образцов из Грузия, Греция, Венгрия, Калабрия (Италия) и Андалусия (Испания). Размеры образцов, проанализированных в этом исследовании, в целом были довольно небольшими, поэтому вполне возможно, что фактическая частота гаплогруппы L-M20 среди средиземноморских европейских популяций может быть немного ниже или выше, чем та, о которой сообщает Semino. и другие., но, похоже, на сегодняшний день не существует исследования, которое бы более точно описало распространение гаплогруппы L-M20 в Юго-Западной Азии и Европе.
| Населения | Распределение | Источник |
|---|---|---|
| Фасция, Италия | 19,2% от Фаши L-M20 | Валентина Коя 2013 |
| Несталь. Италия | 10% от Nonesi L-M20 | Ф. ди Джакомо 2003 |
| Самний, Италия | 10% от Аквиланис L-M20 | Алессио Боаттини 2013 |
| Виченца, Италия | 10% от Венецианцы L-M20 | Алессио Боаттини 2013 |
| Южный Тироль, Италия | 8,9% от Ладин спикеры из Валь Бадиа, 8,3% Валь Бадиа, 2,9% Пустер-Вэлли, 2,2% Немецкий спикеры из Валь Бадиа, 2% говорящих по-немецки из Верхнего Виншгау, 1,9% говорящих на немецком языке из Нижнего Виншгау и 1,7% Итальянский спикеры из Больцано | Пихлер 2006 и Томас 2007. |
| Грузины | 20% (2/10) грузин в Гали, 14,3% (2/14) грузин в Чохатаури, 12,5% (2/16) грузин в Мартвили, 11,8% (2/17) грузин в Абаша, 11,1% (2/18) грузин в Багдати, 10% (1/10) грузин в Гардабани, 9,1% (1/11) грузин в Адигени, 6,9% (2/29) грузин в Омало, 5,9% (1/17) грузин в Гурджаани, 5,9% (1/17) грузин в Лентехи и от 1,5% (1/66) L-M357 (xPK3) до 1,6% (1/63) L-M11 | Батталья 2008, Семино 2000 и Тархнишвили 2014 |
| Дагестан, Россия | 10% от Чеченцы в Дагестане, 9,5% (4/42) из Аварцы, 8,3% (2/24) Таты, 3,7% (1/27) Chamalins | Юнусбаев 2006 г., Caciagli 2009 и Карафет 2016 |
| Архангельская область, Россия | 5,9% от Россияне L1c-M357 | Хунъян Сюй 2014 |
| Эстония | L2-L595 и L1-M22 встречаются в 5,3%, 3,5%, 1,4% и 0,8% Эстонцы | Скоццари 2001 и Лаппалайнен 2007 |
| Балкарцы, Россия | 5,3% (2/38) L-M317 | Батталья 2008 |
| Португалия | 5,0% от Коимбра | Белеза 2006 |
| Болгария | 3,9% от Болгары | Карафет 2016 |
| Фландрия | L1a *: 3,17% от Мехелен 2,4% от Turnhout и 1,3% Кемпен. L1b *: 0,74% от Западная Фландрия и Восточная Фландрия | Лармюзо 2010 и Larmuseau 2011 |
| Анциферово, Новгород | 2,3% от Россияне | Балановская 2017 |
| Восточный Тироль, Австрия | L-M20 содержится в 1,9% Тирольцы в регионе B (Исель, Нижний Драу, Defereggen, Virgen, и Долина кальс ) | Х.Нидерштеттер 2012 |
| Гипускоа, Страна Басков | L1b встречается в 1,7% Гипузкоанцы | Молодой 2011 |
| Северный Тироль, Австрия | L-M20 содержится в 0,8% Тирольцы в Ройтте | Д.Эрхарт 2012 |
Распределение субкладов
L1 (M295)
L-M295 находится из западная Европа к Южная Азия.[Сноска 5]
Субклад L1 также встречается на низких частотах на Коморские острова.[39]
L1a1 (M27)
L-M27 содержится в 14,5% Индейцы и 15% Шриланкийцы, с умеренным распространением в других популяциях Пакистан, южный Иран и Европа, но немного выше Ближнего Востока Араб население[нужна цитата ] (Карафет 2016). Очень незначительное присутствие среди Сидди (2%),[40] также.
L1a2 (M357)
L-M357 часто встречается среди Бурушос, Калаши, и Пуштуны, с умеренным распространением среди других популяций в Пакистан, Грузия,[41] Чеченцы,[42] Ингуши,[42] северный Иран, Индия, то ОАЭ, и Саудовская Аравия.[нужна цитата ]
Китайское исследование, опубликованное в 2018 году, обнаружило L-M357 / L1307 в 7,8% (5/64) образца Лоплика. Уйгуры из села Карчуга, Лопнурский уезд, Синьцзян.[34]
- L-PK3
L-PK3, который находится после L-M357,[43] часто встречается среди Калаш.[нужна цитата ]
L1b (M317)
L-M317 находится на низкой частоте в Центральная Азия, Юго-Западная Азия, и Европа.[нужна цитата ]
В Европе L-M317 был обнаружен на северо-востоке. Итальянцы (3/67 = 4.5%)[41] и Греки (1/92 = 1.1%).[41]
На Кавказе L-M317 был обнаружен в Горские евреи (2/10 = 20%[44]), Аварцы (4/42 = 9.5%,[44] 3%[42]), Балкарцы (2/38 = 5.3%),[41] Абхазский (8/162 = 4.9%,[44] 2/58 = 3.4%[42]), Chamalals (1/27 = 3.7%[44]), Абазины (2/88 = 2.3%[44]), Адыги (3/154 = 1.9%[44]), Чеченцы (3/165 = 1.8%[44]), Армяне (1/57 = 1.8%[44]), Лезгины (1/81 = 1.2%[42]), и Осетины (1/132 = 0.76% Северные осетины,[44] 2/230 = 0,9% железа[42]).
L-M317 был найден в Макранис (2/20 = 10%) в Пакистане, Иранцы (3/186 = 1,6%), пуштуны в Афганистане (1/87 = 1,1%) и Узбеки в Афганистане (1/127 = 0,79%).[45]
L1b1 (M349)
L-M349 в основном встречается в Европе.[нужна цитата ]
L2 (L595)
L2-L595 встречается крайне редко и был идентифицирован частным тестированием на лицах из Европы и Западной Азии.
Возможные, но неподтвержденные случаи L2 включают 4% (1/25) L-M11 (xM76, M27, M317, M357) в выборке иранцев в Кордестан[45] и 2% (2/100) L-M20 (xM27, M317, M357) в образце Шапсуги,[42] среди других редких зарегистрированных случаев L, которые не попадают в общие ветви.
Древняя ДНК
Гаплогруппа L была обнаружена в BMAC сайты в Центральная Азия и в останках захоронения V века в г. Паннония, Венгрия, приписываемый элитному представителю Гунны.[46]
Три человека, которые жили в Энеолит эпоха (ок. 5700–6250 гг. BP ), найденный в пещере Арени-1 («Птичий глаз») в г. Южный Кавказ горы (современные Область Вайоц Дзор, Армения ), также были идентифицированы как принадлежащие к гаплогруппе L1a. Геном одного человека показал, что у него были рыжие волосы и голубые глаза.
Бактрийско-Маргианский археологический комплекс
Нарасимхан и др. (2018) проанализировали скелеты из BMAC сайты в Узбекистан и идентифицировал 2 человека как принадлежащих к гаплогруппе L1a. Один из этих экземпляров был найден в Бустане, а другой - в Саппали-Тепе; как установлено, что они Бронзовый век места.[47]
Могила элитных гуннов
| Я БЫ | |
|---|---|
| Y ДНК | L-M20 |
| численность населения | Гунны |
| Язык | неизвестный; возможно Гуннский |
| Культура | |
| Дата (YBP) | 1540–1500 лн |
| Захоронение / Местоположение | Венгрия |
| Члены / размер выборки | 1/1 |
| Процент | |
| мтДНК | D4j12 |
| Изотоп Sr | |
| Цвет глаз (система HIrisPlex) | |
| Цвет волос (система HIrisPlex) | |
| Пигментация кожи | |
| Группа крови ABO | |
| Диета (d13C% 0 / d15N% 0) | |
| Деятельность FADS | |
| Стойкость лактазы | |
| Оазе-1 Общая ДНК | |
| Ostuni1 Общая ДНК | |
| Неандерталец Vi33.26 Общая ДНК | |
| Неандерталец Vi33.25 Общая ДНК | |
| Неандерталец Vi33.16 Общая ДНК | |
| Родовой компонент (AC) | |
| puntDNAL K12 Древний | |
| Dodecad [dv3] | |
| Еврогены [K = 36] | |
| Dodecad [Globe13] | |
| Генетическая дистанция | |
| Родительское родство | |
| Возраст на момент смерти | |
| Позиция смерти | |
| SNP | |
| Читать пары | |
| Образец | |
| Источник | Лаборатория популяционной генетики Казахстана |
| Примечания |
Энеолит Южный Кавказ
| Свойство | Арени-I | Арени-II | Арени-III |
|---|---|---|---|
| Я БЫ | AR1 / 44 I1634 | AR1 / 46 I1632 | ARE12 I1407 |
| Y ДНК | L1a | L1a1-M27 | L1a |
| численность населения | Энеолит (горизонт III) | Энеолит (горизонт III) | Энеолит (Горизонт II) |
| Язык | |||
| Культура | Поздний энеолит | Поздний энеолит | Поздний энеолит |
| Дата (YBP) | 6161 ± 89 | 6086 ± 72 | 6025 ± 325 |
| Захоронение / Местоположение | Могильник 2 / Пещера Арени-1 | Могильник 3 / Пещера Арени-1 | Траншея 2А, Блок 7, Площадь S33 / T33, Локус 9, Стрелка 23 / Пещера Арени-1 |
| Члены / размер выборки | 1/3 | 1/3 | 1/3 |
| Процент | 33.3% | 33.3% | 33.3% |
| мтДНК | H2a1 | K1a8 | ЧАС* |
| Изотоп Sr | |||
| Цвет глаз (система HIrisPlex) | Вероятно, синий | ||
| Цвет волос (система HIrisPlex) | Вероятно, красный | ||
| Пигментация кожи | Вероятно, легкий | ||
| Группа крови ABO | Скорее всего O или B | ||
| Диета (d13C% 0 / d15N% 0) | |||
| Деятельность FADS | |||
| Стойкость лактазы | Вероятная непереносимость лактозы | ||
| Оазе-1 Общая ДНК | |||
| Ostuni1 Общая ДНК | |||
| Неандерталец Vi33.26 Общая ДНК | |||
| Неандерталец Vi33.25 Общая ДНК | |||
| Неандерталец Vi33.16 Общая ДНК | |||
| Наследственный компонент (AC) | |||
| puntDNAL K12 Древний | |||
| Dodecad [dv3] | |||
| Еврогены [K = 36] | |||
| Dodecad [Globe13] | |||
| Генетическая дистанция | |||
| Родительское родство | |||
| Возраст на момент смерти | 11 ± 2.5 | 15 ± 2.5 | |
| Позиция смерти | |||
| SNP | |||
| Читать пары | |||
| Образец | |||
| Источник | [48] | ||
| Примечания | Самые ранние в мире свидетельства изготовления обуви и виноделия |
Алалах Аморитский город-государство
Skourtanioti et al. (2020) проанализировали скелеты из Алалах сайты в Сирия и идентифицировал одного человека как принадлежащего к гаплогруппе L-L595, и он был датирован, скорее всего, 1941–1883 гг. до н.э.[49]
Номенклатура
До 2002 года в академической литературе существовало по крайней мере семь систем именования Филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (стенография) | (α) | (β) | (γ) | (δ) | (е) | (ζ) | (η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L-M20 | 28 | VIII | 1U | 27 | Eu17 | H5 | F | L * | L | L | L | - | - | - | - | - | - | - |
| L-M27 | 28 | VIII | 1U | 27 | Eu17 | H5 | F | L1 | L1 | L1 | L1 | - | - | - | - | - | - | - |
- Дерево Консорциума Y-хромосомы
Это официальное научное дерево, созданное Консорциумом Y-хромосомы (YCC). Последнее крупное обновление было в 2008 г. (Карафет 2008 ). Последующие обновления производились ежеквартально и раз в два года. Текущая версия является переработкой обновления 2010 года.[50]
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Январь 2013) |
- Оригинальные научные публикации
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Смотрите также
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Сноски
- ^ видеть Басу 2003, Кордо 2004, Сенгупта 2006, и Тамсим 2006.
- ^ 12/222 Shlush et al. 2008 г.
- ^ 1/25 Shlush et al. 2008 г.
- ^ В Hammer 2005 см. Дополнительные материалы.
- ^ Результаты лаборатории FTDNA, май 2011 г.
Рекомендации
- ^ Махал, Дэвид Дж .; Мацукас, Янис Г. (23 января 2018 г.). «Географическое происхождение этнических групп на Индийском субконтиненте: изучение древних следов с помощью гаплогрупп Y-ДНК». Границы генетики. 9: 4. Дои:10.3389 / fgene.2018.00004. ISSN 1664-8021. ЧВК 5787057. PMID 29410676.
- ^ Лакау, Харлетт; Гайден, Тензин; Регейро, Мария; Ченнакришнаия, Шилпа; Бухари, Аридж; Андерхилл, Питер А; Гарсия-Бертран, Ральф Л; Эррера, Рене Дж (18 апреля 2012 г.). «Афганистан с точки зрения Y-хромосомы». Европейский журнал генетики человека. 20 (10): 1063–1070. Дои:10.1038 / ejhg.2012.59. ISSN 1018-4813. ЧВК 3449065. PMID 22510847.
- ^ Сенгупта, Сангхамитра; Животовский, Лев А .; Кинг, Рой; Mehdi, S. Q .; Эдмондс, Кристофер А .; Чоу, Шерил-Эмилиан Т .; Лин, Алиса А .; Митра, Миташри; Sil, Samir K .; Рамеш, А .; Usha Rani, M. V .; Thakur, Chitra M .; Кавалли-Сфорца, Л. Лука; Majumder, Partha P .; Андерхилл, Питер А. (2006). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии выявляют как коренное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии». Американский журнал генетики человека. 78 (2): 202–221. Дои:10.1086/499411. ISSN 0002-9297. ЧВК 1380230. PMID 16400607.
- ^ Международное общество генетической генеалогии, 2015, Дерево гаплогруппы Y-ДНК 2015 (30 мая 2015 г.).
- ^ а б Chiaroni, J .; Андерхилл, П. А .; Кавалли-Сфорца, Л. Л. (декабрь 2009 г.). «Разнообразие Y-хромосомы, человеческое развитие, дрейф и культурная эволюция». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 106 (48): 20174–49. Bibcode:2009PNAS..10620174C. Дои:10.1073 / pnas.0910803106. JSTOR 25593348. ЧВК 2787129. PMID 19920170.
- ^ Международное общество генетической генеалогии, 2015 Y-ДНК Гаплогруппа K и ее субклады - 2015 (5 апреля 2015 г.).
- ^ Уэллс, Спенсер (2007). Глубокое происхождение: внутри генографического проекта. Вашингтон, округ Колумбия: National Geographic. п. 103. ISBN 978-1426201189.
- ^ Камар, Рахил; Аюб, Касим; Мохьюддин, Аиша; Хельгасон, Агнар; Мажар, Кекашан; Мансур, Атика; Зерджаль, Татьяна; Тайлер-Смит, Крис; Мехди, С. Касим (2002). «Вариация Y-хромосомной ДНК в Пакистане». Американский журнал генетики человека. 70 (5): 1107–1124. Дои:10.1086/339929. ISSN 0002-9297. ЧВК 447589. PMID 11898125.
- ^ Чжао, Чжунмин; Хан, Фейсал; Боркар, Минал; Эррера, Рене; Агравал, Суракша (2009). «Наличие трех разных отцовских линий среди северных индейцев: исследование 560 Y-хромосом». Анналы биологии человека. 36 (1): 46–59. Дои:10.1080/03014460802558522. ISSN 0301-4460. ЧВК 2755252. PMID 19058044.
- ^ Тансим, Исмаил; Тангарадж, Кумарасами; Чаубей, Гианешвер; Сингх, Виджай Кумар; Бхаскар, Лаккакула В.КС .; Редди, Б. Мохан; Редди, Алла Г; Сингх, Лалджи (7 августа 2006 г.). «Генетическое родство среди низших каст и племенных групп Индии: вывод из Y-хромосомы и митохондриальной ДНК». BMC Genetics. 7: 42. Дои:10.1186/1471-2156-7-42. ISSN 1471-2156. ЧВК 1569435. PMID 16893451.
- ^ Кордо, Ричард; Аунгер, Роберт; Бентли, Джиллиан; Насидзе, Иване; Sirajuddin, S.M .; Стоункинг, Марк (3 февраля 2004 г.). «Независимое происхождение индейской касты и племенных отцовских линий». Текущая биология. 14 (3): 231–235. Дои:10.1016 / j.cub.2004.01.024. ISSN 0960-9822. PMID 14761656. S2CID 5721248.
- ^ McElreavey, K .; Кинтана-Мурси, Л. (2005). «Перспектива популяционной генетики долины Инда с помощью маркеров, унаследованных от одного родителя». Анналы биологии человека. 32 (2): 154–162. Дои:10.1080/03014460500076223. ISSN 0301-4460. PMID 16096211. S2CID 109014.
- ^ Тангарадж, Кумарасами; Найду, Б. Пратап; Кривелларо, Федерика; Таманг, Ракеш; Упадхьяй, Шашанк; Шарма, Варун Кумар; Редди, Алла Г .; Walimbe, S. R .; Чаубей, Гианешвер; Кивисилд, Тоомас; Сингх, Лалджи (20 декабря 2010 г.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры». PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. Дои:10.1371 / journal.pone.0015283. ISSN 1932-6203. ЧВК 3004917. PMID 21187967.
- ^ К. МакЭлриви и Л. Кинтана-Мурси (2005 г.), Перспектива популяционной генетики долины Инда с помощью маркеров, унаследованных от одного родителя. В архиве 2017-01-16 в Wayback Machine
- ^ а б c d Сенгупта 2006.
- ^ Сенгупта 2006, п. 218.
- ^ Кивисилд, Т .; Rootsi, S .; Мецпалу, М .; Mastana, S .; Kaldma, K .; Parik, J .; Мецпалу, Э .; Adojaan, M .; Tolk, H.-V .; Степанов, В .; Gölge, M .; Usanga, E .; Papiha, S. S .; Cinniolu, C .; King, R .; Cavalli-Sforza, L .; Андерхилл, П. А .; Виллемс Р. (2003). «Генетическое наследие первых поселенцев сохраняется как в индейских племенах, так и в кастах». Американский журнал генетики человека. 72 (2): 313–332. Дои:10.1086/346068. ISSN 0002-9297. ЧВК 379225. PMID 12536373.
- ^ Сенгупта 2006, п. 219.
- ^ Сенгупта 2006, п. 220.
- ^ а б Камар 2002.
- ^ а б c Шах 2011.
- ^ а б Кивисилд, Т; Рутси, S; Мецпалу, М; и другие. (Февраль 2003 г.). «Генетическое наследие первых поселенцев сохраняется как в индейских племенах, так и в кастах». Являюсь. J. Hum. Genet. 72 (2): 313–32. Дои:10.1086/346068. ЧВК 379225. PMID 12536373.
- ^ Махал, Дэвид Дж .; Мацукас, Янис Г. (20 сентября 2017 г.). «Разнообразие гаплогрупп Y-STR в популяции джат свидетельствует о нескольких различных древних источниках». Границы генетики. 8: 121. Дои:10.3389 / fgene.2017.00121. ISSN 1664-8021. ЧВК 5611447. PMID 28979290.
- ^ Шарма, S; Рай, Э; Шарма, П; и другие. (Январь 2009 г.). «Индийское происхождение отцовской гаплогруппы R1a1 * подтверждает автохтонное происхождение браминов и кастовой системы». Журнал генетики человека. 54 (1): 47–55. Дои:10.1038 / jhg.2008.2. PMID 19158816.
- ^ а б Сенгупта, S; Животовский, Л.А.; King, R; и другие. (Февраль 2006 г.). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии выявляют как коренное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии». Являюсь. J. Hum. Genet. 78 (2): 202–21. Дои:10.1086/499411. ЧВК 1380230. PMID 16400607.
- ^ «Анализ разнообразия Y-хромосомы в популяциях лингаятов и воккалига в Южной Индии». 2011 г. CiteSeerX 10.1.1.425.9132. Цитировать журнал требует
| журнал =(помощь) - ^ Чаубей, Гианешвер (2016). «Генетическая близость еврейского населения Индии». Научные отчеты. 6: 19166. Bibcode:2016НатСР ... 619166C. Дои:10.1038 / srep19166. ЧВК 4725824. PMID 26759184.
- ^ Ди Кристофаро, Джули; Пеннарун, Эрван; Мазьер, Стефан; Майрес, Натали М .; Лин, Алиса А .; Темори, Шах Ага; Мецпалу, Майт; Мецпалу, Эне; Витцель, Майкл; Кинг, Рой Дж .; Андерхилл, Питер А .; Виллемс, Ричард; Кьярони, Жак (2013). «Афганский Гиндукуш: там, где сходятся потоки генов Евразийского субконтинента». PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO ... 876748D. Дои:10.1371 / journal.pone.0076748. ЧВК 3799995. PMID 24204668.
- ^ Камар, Р; Аюб, Q; Мохьюддин, А; и другие. (Май 2002 г.). «Вариация Y-хромосомной ДНК в Пакистане». Являюсь. J. Hum. Genet. 70 (5): 1107–24. Дои:10.1086/339929. ЧВК 447589. PMID 11898125.
- ^ Firasat, S; Халик, S; Мохьюддин, А; и другие. (Январь 2007 г.). «Доказательства Y-хромосомы ограниченного вклада Греции в популяцию патанов в Пакистане». Евро. J. Hum. Genet. 15 (1): 121–26. Дои:10.1038 / sj.ejhg.5201726. ЧВК 2588664. PMID 17047675.
- ^ а б c Lacau, H; Гайден, Т; Регейро, М; Ченнакришнайя, S; Бухари, А; Андерхилл, Пенсильвания; Гарсия-Бертран, Р.Л .; Эррера, RJ (октябрь 2012 г.). «Афганистан с точки зрения Y-хромосомы». Европейский журнал генетики человека. 20 (10): 1063–70. Дои:10.1038 / ejhg.2012.59. ЧВК 3449065. PMID 22510847.
- ^ а б Габер, М; Platt, DE; Ашрафиан Бонаб, М; и другие. (2012). «Этнические группы Афганистана разделяют наследие Y-хромосомы, структурированное историческими событиями». PLOS ONE. 7 (3): e34288. Bibcode:2012PLoSO ... 734288H. Дои:10.1371 / journal.pone.0034288. ЧВК 3314501. PMID 22470552.
- ^ а б c d Ди Кристофаро, Дж; Pennarun, E; Mazières, S; Майрес, Нью-Мексико; Линь А.А.; Temori, SA; Мецпалу, М; Мецпалу, Э; Witzel, M; King, RJ; Андерхилл, Пенсильвания; Виллемс, Р. Кьярони, Дж (2013). «Афганский Гиндукуш: там, где сходятся потоки генов Евразийского субконтинента». PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO ... 876748D. Дои:10.1371 / journal.pone.0076748. ЧВК 3799995. PMID 24204668.
- ^ а б Лю SH, N, Yilihamu, R Bake, и другие. (2018), «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica, 2018, 37(1): 146-156.
- ^ Чжун 2010.
- ^ Федорова 2013.
- ^ Карафет 2010.
- ^ Ким 2011.
- ^ Мсаиди, Саид; и другие. (2011). «Генетическое разнообразие на Коморских островах показывает, что раннее мореплавание является основным фактором биокультурной эволюции человека в западной части Индийского океана» (PDF). Европейский журнал генетики человека. 19 (1): 89–94. Дои:10.1038 / ejhg.2010.128. ЧВК 3039498. PMID 20700146.
- ^ Шах, AM; Tamang, R; Moorjani, P; Рани, DS; Govindaraj, P; Кулкарни, G; Бхаттачарья, Т; Мустак, М.С.; Bhaskar, LV; Редди, АГ; Гадви, Д; Гай, ПБ; Чауби, G; Паттерсон, Н. Райх, Д; Тайлер-Смит, К; Сингх, L; Тангарадж, К. (2011). "Индийские сидди: потомки африканцев с индийской примесью". Являюсь. J. Hum. Genet. 89 (1): 154–61. Дои:10.1016 / j.ajhg.2011.05.030. ЧВК 3135801. PMID 21741027.
- ^ а б c d Винченца Батталья, Симона Форнарино, Надя Аль-Захери, и другие. (2009), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе». Европейский журнал генетики человека (2009) 17, 820–830; DOI: 10.1038 / ejhg.2008.249; опубликовано в сети 24 декабря 2008 г.
- ^ а б c d е ж грамм Балановский, Олег; Дибирова, Хадижат; Дыбо, Анна; и другие. (Октябрь 2011 г.). «Параллельная эволюция генов и языков в Кавказском регионе». Молекулярная биология и эволюция. 28 (10): 2905–2920. Дои:10.1093 / molbev / msr126. ЧВК 3355373. PMID 21571925.
- ^ ISOGG 2016.
- ^ а б c d е ж грамм час я Юнусбаев, Баязит; Мецпалу, Майт; Ярве, Мари; и другие. (2012). «Кавказ как асимметричный полупроницаемый барьер для древних миграций людей». Молекулярная биология и эволюция. 29 (1): 359–365. Дои:10.1093 / molbev / msr221. PMID 21917723.
- ^ а б Ди Кристофаро, Дж; Pennarun, E; Mazières, S; Майрес, Нью-Мексико; Линь А.А.; и другие. (2013). «Афганский Гиндукуш: там, где сходятся потоки генов Евразийского субконтинента». PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO ... 876748D. Дои:10.1371 / journal.pone.0076748. ЧВК 3799995. PMID 24204668.
- ^ Уали, Мурат (25 ноября 2016 г.). "Казахстанский ДНК-проект" [Проект ДНК Казахстана]. Np.kz (на русском). Архивировано из оригинал 26 ноября 2016 г.. Получено 10 января 2016.
- ^ «Геномная формация Южной и Центральной Азии». bioRxiv: 292581.31 марта 2018 г. Дои:10.1101/292581.
- ^ Лазаридис, Иосиф; и другие. (25 июля 2016 г.). «Геномное понимание происхождения земледелия на древнем Ближнем Востоке». Природа. 536 (7617): 419–24. Bibcode:2016Натура.536..419L. bioRxiv 10.1101/059311. Дои:10.1038 / природа19310. ЧВК 5003663. PMID 27459054.
- ^ Скуртаниоти, Эйрини; Erdal, Yilmaz S .; Франжипане, Марселла; Балосси Рестелли, Франческа; Йенер, К. Аслыхан; Пиннок, Фрэнсис; Маттиа, Паоло; Озбал, Рана; Шуп, Ульф-Дитрих; Гулиев, Фархад; Ахундов, Туфан; Lyonnet, Bertille; Хаммер, Эмили Л .; Nugent, Selin E .; Бурри, Марта; Neumann, Gunnar U .; Пенске, Сандра; Ингман, Тара; Акар, Мурат; Шафик, Рула; Палумби, Джулио; Айзенманн, Стефани; д'Андреа, Марта; Рорлах, Адам Б .; Уорнер, Кристина; Чон, Чунгвон; Stockhammer, Philipp W .; Хаак, Вольфганг; Краузе, Йоханнес (2020). «Геномная история от неолита до бронзового века Анатолии, Северного Леванта и Южного Кавказа». Клетка. 181 (5): 1158–1175.e28. Дои:10.1016 / j.cell.2020.04.044. PMID 32470401. S2CID 219105572.
- ^ "Y-ДНК Haplotree". Family Tree DNA использует дерево Y-хромосомы Консорциума и размещает его на своем веб-сайте.
Источники
Журналы
- Абу-Амеро, К. К .; Hellani, A .; González, A.M .; Larruga, J.M .; Кабрера, В. М .; Андерхилл, П. А. (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с соседними регионами». BMC Genetics. 10: 59. Дои:10.1186/1471-2156-10-59. ЧВК 2759955. PMID 19772609.
- Basu, A .; Mukherjee, N .; Рой, С .; Sengupta, S .; Banerjee, S .; Чакраборти, М .; Dey, B .; Рой, М .; Рой, Б.; Bhattacharyya, N.P .; Roychoudhury, S .; Маджумдер, П. П. (2003). «Этническая Индия: геномный взгляд с особым упором на население и структуру». Геномные исследования. 13 (10): 2277–90. Дои:10.1101 / гр. 1413403. ЧВК 403703. PMID 14525929.
- Battaglia, V .; Форнарино, С .; Аль-Захери, Н .; Olivieri, A .; Pala, M .; Myres, N.M .; King, R.J .; Rootsi, S .; Марьянович, Д .; Primorac, D .; Hadziselimovic, R .; Vidovic, S .; Дробник, К .; Дурмиши, Н .; Torroni, A .; Santachiara-Benerecetti, A.S .; Андерхилл, П. А .; Семино, О. (2008). «Y-хромосомное свидетельство культурного распространения сельского хозяйства в Юго-Восточной Европе». Европейский журнал генетики человека. 17 (6): 820–30. Дои:10.1038 / ejhg.2008.249. ЧВК 2947100. PMID 19107149.
- Белеза, С .; Gusmao, L .; Lopes, A .; Alves, C .; Gomes, I .; Giouzeli, M .; Calafell, F .; Карраседо, А .; Аморим, А. (2006). «Микрофилогеографическая и демографическая история португальских мужских родов». Анналы генетики человека. 70 (2): 181–94. Дои:10.1111 / j.1529-8817.2005.00221.x. PMID 16626329. S2CID 4652154.
- Caciagli, L .; Булаева, К .; Булаев, О .; Bertoncini, S .; Taglioli, L .; Pagani, L .; Paoli, G .; Тофанелли, С. (2009). «Ключевая роль отцовской наследственности в формировании генетической изменчивости горцев Дагестана». Журнал генетики человека. 54 (12): 689–94. Дои:10.1038 / jhg.2009.94. PMID 19911015.
- Каденас, А. М .; Животовский, Л. А .; Cavalli-Sforza, L. L .; Андерхилл, П. А .; Эррера, Р. Дж. (2007). «Разнообразие Y-хромосомы характеризует Оманский залив». Европейский журнал генетики человека. 16 (3): 374–86. Дои:10.1038 / sj.ejhg.5201934. PMID 17928816.
- Cinnioğlu, C .; King, R .; Кивисилд, Т .; Kalfoglu, E .; Атасой, С .; Cavalleri, G.L .; Lillie, A. S .; Roseman, C.C .; Lin, A. A .; Князь, К .; Oefner, P.J .; Shen, P .; Семино, О .; Cavalli-Sforza, L. L .; Андерхилл, П. А. (2004). «Раскопки гаплотипов Y-хромосомы в Анатолии». Генетика человека. 114 (2): 127–48. Дои:10.1007 / s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- Cordaux, R .; Aunger, R .; Bentley, G .; Насидзе, И .; Sirajuddin, S.M .; Стоункинг, М. (2004). «Независимое происхождение индийской касты и племенных отцовских линий». Текущая биология. 14 (3): 231–35. Дои:10.1016 / j.cub.2004.01.024. PMID 14761656. S2CID 5721248.
- Эль-Сибай, М .; Platt, D.E .; Haber, M .; Xue, Y .; Youhanna, S.C .; Wells, R. S .; Izaabel, H .; Sanyoura, M. F .; Harmanani, H .; Bonab, M. A .; Behbehani, J .; Hashwa, F .; Тайлер-Смит, К .; Zalloua, P.A .; Genographic, Консорциум (2009). «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст между прибрежными и внутренними районами». Анналы генетики человека. 73 (6): 568–81. Дои:10.1111 / j.1469-1809.2009.00538.x. ЧВК 3312577. PMID 19686289.
- Федорова, С. А .; Reidla, M .; Мецпалу, Э .; Мецпалу, М .; Rootsi, S .; Тамбец, К .; Трофимова, Н .; Жаданов, С. И .; Kashani, B.H .; Olivieri, A .; Воевода, М. И .; Осипова, Л. П .; Платонов, Ф. А .; Томский, М. И .; Хуснутдинова, Э.К .; Torroni, A .; Виллемс Р. (2013). "Аутосомные и однопородные портреты коренного населения Саха (Якутия): значение для заселения Северо-Восточной Евразии". BMC Эволюционная биология. 13 (127): 127. Дои:10.1186/1471-2148-13-127. ЧВК 3695835. PMID 23782551.
- Firasat, S .; Khaliq, S .; Mohyuddin, A .; Papaioannou, M .; Тайлер-Смит, К .; Андерхилл, П. А .; Аюб, К. (2007). «Доказательства Y-хромосомы ограниченного вклада Греции в популяцию патанов в Пакистане». Европейский журнал генетики человека. 15 (1): 121–26. Дои:10.1038 / sj.ejhg.5201726. ЧВК 2588664. PMID 17047675.
- Карафет, Т .; Xu, L .; Du, R .; Wang, W .; Feng, S .; Wells, R. S .; Редд, А. Дж .; Zegura, S.L .; Хаммер, М. Ф. (2001). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы». Американский журнал генетики человека. 69 (3): 615–28. Дои:10.1086/323299. ЧВК 1235490. PMID 11481588.
- Карафет, Т. М .; Проба, B .; Cox, M. P .; Sudoyo, H .; Дауни, S .; Lansing, J. S .; Хаммер, М. Ф. (2010). «Основное деление Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии». Мол. Биол. Evol. 27 (8): 1833–44. Дои:10.1093 / molbev / msq063. PMID 20207712.
- Kim, S-H .; Kim, K-C .; Шин, Д-Дж .; Джин, HJ .; Kwak, K-D .; Han, M-S .; Песня, JM .; Kim, W .; Ким, В. (2011). «Высокие частоты линий гаплогруппы O2b-SRY465 Y-хромосомы в Корее: генетическая перспектива заселения Кореи». Следственная генетика. 2 (1): 10. Дои:10.1186/2041-2223-2-10. ЧВК 3087676. PMID 21463511.
- Lappalainen, T .; Laitinen, V .; Salmela, E .; Andersen, P .; Huoponen, K .; Savontaus, M.-L .; Лахермо, П. (2008). «Миграционные волны в регион Балтийского моря». Анналы генетики человека. 72 (3): 337–48. Дои:10.1111 / j.1469-1809.2007.00429.x. PMID 18294359. S2CID 32079904.
- Larmuseau, M.H.D .; Vanderheyden, N .; Джейкобс, М .; Coomans, M .; Ларно, Л .; Декорте, Р. (2010). «Микрогеографическое распределение вариаций Y-хромосомы в центрально-западном европейском регионе Брабант». Forensic Science International: генетика. 5 (2): 95–99. Дои:10.1016 / j.fsigen.2010.08.020. PMID 21036685.
- Larmuseau, M.H.D .; Ottoni, C .; Raeymaekers, J. A. M .; Vanderheyden, N .; Larmuseau, H. F. M .; Декорте, Р. (2011). «Временная дифференциация в западноевропейской группе Y-хромосомы: генеалогия как инструмент популяционной генетики человека». Европейский журнал генетики человека. 20 (4): 434–40. Дои:10.1038 / ejhg.2011.218. ЧВК 3306861. PMID 22126748.
- Луис, Дж. Р .; Rowold, D. J .; Regueiro, M .; Caeiro, B .; Cinniolu, C .; Roseman, C .; Андерхилл, П. А .; Cavalli-Sforza, L. L .; Эррера, Р. Дж. (2004). "Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей". Американский журнал генетики человека. 74 (3): 532–44. Дои:10.1086/382286. ЧВК 1182266. PMID 14973781.
- Пихлер, I .; Mueller, J.C .; Стефанов, С. А .; Де Гранди, А .; Beu Volpato, C .; Pinggera, G.K .; Mayr, A .; Ogriseg, M .; Ploner, F .; Meitinger, T .; Прамсталлер, П. П. (2006). «Генетическая структура современных изолированных популяций Южного Тироля, выявленная с помощью анализа полиморфизмов Y-хромосомы, мтДНК и Alu». Человеческая биология. 81 (5–6): 875–98. Дои:10.3378/027.081.0629. PMID 20504204. S2CID 46073270.
- Qamar, R .; Ayub, Q .; Mohyuddin, A .; Helgason, A .; Mazhar, K .; Mansoor, A .; Zerjal, T .; Тайлер-Смит, К .; Мехди, С.К. (2002). «Вариация Y-хромосомной ДНК в Пакистане». Американский журнал генетики человека. 70 (5): 1107–24. Дои:10.1086/339929. ЧВК 447589. PMID 11898125.
- Regueiro, M .; Каденас, А. М .; Гайден, Т .; Андерхилл, П. А .; Эррера, Р. Дж. (2006). «Иран: триконтинентальный нексус для миграции, вызванной Y-хромосомой». Человеческая наследственность. 61 (3): 132–43. Дои:10.1159/000093774. PMID 16770078. S2CID 7017701.
- Sahoo, S .; Сингх, А .; Himabindu, G .; Banerjee, J .; Sitalaximi, T .; Gaikwad, S .; Триведи, Р .; Endicott, P .; Кивисилд, Т .; Мецпалу, М .; Villems, R .; Кашьяп, В. К. (2006). «Предыстория индийских Y-хромосом: оценка сценариев демической диффузии». Труды Национальной академии наук. 103 (4): 843–8. Bibcode:2006ПНАС..103..843С. Дои:10.1073 / pnas.0507714103. ЧВК 1347984. PMID 16415161.
- Sanchez, J. J .; Hallenberg, C .; Børsting, C .; Эрнандес, А .; Горлин, Р. Дж. (2005). «Высокие частоты клонов Y-хромосомы, характеризующиеся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин». Европейский журнал генетики человека. 13 (7): 856–66. Дои:10.1038 / sj.ejhg.5201390. PMID 15756297.
- Scozzari, R .; Cruciani, F .; Pangrazio, A .; Santolamazza, P .; Vona, G .; Мораль, П .; Latini, V .; Varesi, L .; Memmi, M. M .; Романо, В .; De Leo, G .; Gennarelli, M .; Jaruzelska, J .; Villems, R .; Parik, J .; MacAulay, V .; Торрони, А. (2001). «Вариации Y-хромосомы человека в районе Западного Средиземноморья: последствия для населения региона». Иммунология человека. 62 (9): 871–84. CiteSeerX 10.1.1.408.4857. Дои:10.1016 / S0198-8859 (01) 00286-5. PMID 11543889.
- Семино, О .; Пассарино, G .; Oefner, P.J .; Lin, A. A .; Арбузова, С .; Beckman, L.E .; De Benedictis, G .; Francalacci, P .; Kouvatsi, A .; Лимборска, С .; Marcikiae, M .; Mika, A .; Мика, Б .; Primorac, D .; Santachiara-Benerecetti, A.S .; Cavalli-Sforza, L. L .; Андерхилл, П. А. (2000). «Генетическое наследие палеолита Homo sapiens sapiens у современных европейцев: перспектива хромосомы Y». Наука. 290 (5494): 1155–59. Bibcode:2000Sci ... 290.1155S. Дои:10.1126 / наука.290.5494.1155. PMID 11073453.
- Sengupta, S .; Животовский, Л. А .; King, R .; Mehdi, S. Q .; Edmonds, C.A .; Чоу, C-E. Т .; Lin, A. A .; Mitra, M .; Sil, S.K .; Рамеш, А .; Usha Rani, M. V .; Thakur, C.M .; Cavalli-Sforza, L. L .; Majumder, P.P .; Андерхилл, П. А. (2006). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии выявляют как коренное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии». Американский журнал генетики человека. 78 (2): 202–21. Дои:10.1086/499411. ЧВК 1380230. PMID 16400607.
- Shah, A.M .; Tamang, R .; Moorjani, P .; Rani, D. S .; Govindaraj, P .; Kulkarni, G .; Bhattacharya, T .; Мустак, М.S .; Бхаскар, Л. В. К. С .; Редди, А.Г .; Gadhvi, D .; Gai, P. B .; Chaubey, G .; Patterson, N .; Reich, D .; Тайлер-Смит, К .; Singh, L .; Тангарадж, К. (2011). "Индийские сидди: потомки африканцев с индийской примесью". Американский журнал генетики человека. 89 (1): 154–61. Дои:10.1016 / j.ajhg.2011.05.030. ЧВК 3135801. PMID 21741027.
- Thamseem, I .; Thangaraj, K .; Chaubey, G .; Singh, V .; Бхаскар, Л. В. К. С .; Reddy, B.M .; Редди, А.Г .; Сингх, Л. (2006). «Генетическая близость среди низших каст и племенных групп Индии: вывод из Y-хромосомы и митохондриальной ДНК». BMC Genetics. 7: 42. Дои:10.1186/1471-2156-7-42. ЧВК 1569435. PMID 16893451.
- Thomas, M. G .; Barnes, I .; Weale, M.E .; Jones, A. L .; Forster, P .; Bradman, N .; Прамсталлер, Питер П. (2008). «Новые генетические данные подтверждают изоляцию и дрейф в ладинских сообществах Южно-Тирольских Альп, но не древнее происхождение на Ближнем Востоке». Европейский журнал генетики человека. 16 (1): 124–34. Дои:10.1038 / sj.ejhg.5201906. PMID 17712356.
- Weale, M .; Епископосян, Л .; Jager, R .; Оганесян, Н .; Худоян, А .; Бербедж-Холл, О .; Bradman, N .; Томас, М. (2001). «Гаплотипы армянской Y-хромосомы обнаруживают сильную региональную структуру в рамках единой этнонациональной группы». Генетика человека. 109 (6): 659–74. Дои:10.1007 / s00439-001-0627-9. PMID 11810279. S2CID 23113666.
- Wells, R. S .; Юлдашева, Н .; Рузибакиев, Р .; Андерхилл, П. А .; Евсеева, И .; Blue-Smith, J .; Jin, L .; Вс, Б .; Pitchappan, R .; Шанмугалакшми, С .; Балакришнан, К .; Читать, М .; Пирсон, Н. М .; Zerjal, T .; Webster, M. T .; Жолошвили, И .; Джамарджашвили, Э .; Гамбаров, С .; Никбин, Б .; Достиев, А .; Акназаров, О .; Zalloua, P .; Цой, И .; Китаев, М .; Миррахимов, М .; Чариев, А .; Бодмер, В. Ф. (2001). «Евразийский хартленд: континентальный взгляд на разнообразие Y-хромосомы». Труды Национальной академии наук. 98 (18): 10244–49. Bibcode:2001PNAS ... 9810244W. Дои:10.1073 / pnas.171305098. ЧВК 56946. PMID 11526236.
- Янг, К. Л .; Sun, G .; Дека, Р .; Кроуфорд, М. Х. (2011). "Отцовская генетическая история баскского населения Испании" (PDF). Человеческая биология. 83 (4): 455–75. Дои:10.3378/027.083.0402. HDL:1808/16387. PMID 21846204. S2CID 3191418.
- Zalloua, P.A .; Xue, Y .; Khalife, J .; Makhoul, N .; Debiane, L .; Platt, D.E .; Ройюру, А.К .; Эррера, Р. Дж .; Hernanz, D. F. S .; Blue-Smith, J .; Wells, R. S .; Comas, D .; Bertranpetit, J .; Тайлер-Смит, К .; Генографический консорциум (2008). «Разнообразие Y-хромосомы в Ливане определяется недавними историческими событиями». Американский журнал генетики человека. 82 (4): 873–82. Дои:10.1016 / j.ajhg.2008.01.020. ЧВК 2427286. PMID 18374297.
Интернет-источники
- ^ а б c Krahn, T .; FTDNA. "Проект Y-ДНК дерева FTDNA (AKA YTree)". Семейное древо ДНК. Архивировано из оригинал на 2015-08-15. Получено 2013-01-01.CS1 maint: ref = harv (связь)
- ^ а б Henson, G .; Гречдакян, П .; FTDNA (2013). "L - Проект Y-гаплогруппы L". Получено 2013-01-01.CS1 maint: ref = harv (связь)
внешняя ссылка
- ISOGG, [1]
- Генная база (2006). "Учебники по генобазе: узнайте о гаплогруппе L Y-хромосомы". Архивировано из оригинал на 2012-10-23.CS1 maint: ref = harv (связь)
- Распространение гаплогруппы L, из Национальная география
- Генеалогический проект Индии
- Y HAPLOGROUP L