Гаплогруппа M-P256 - Haplogroup M-P256
| Гаплогруппа M-P256 | |
|---|---|
 | |
| Возможное время происхождения | 32000-47000 лет назад (Scheinfeldt 2006 ) |
| Возможное место происхождения | Wallacea (восточная Индонезия) или же Новая Гвинея [1] |
| Предок | K2b1 |
| Определение мутаций | P256 |
Гаплогруппа M, также известный как M-P256 и Гаплогруппа K2b1b (ранее K2b1d) является Гаплогруппа ДНК Y-хромосомы. M-P256 является потомком гаплогруппы Гаплогруппа K2b1, и считается, что впервые он появился между 32000 и 47000 лет назад (Scheinfeldt 2006 ).
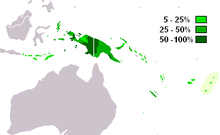
M-P256 - наиболее часто встречающаяся гаплогруппа Y-хромосомы в Западное Папуа и западный Папуа - Новая Гвинея (Кайзер 2003 ). Встречается также в соседних частях Меланезия, Индонезия и среди коренные австралийцы.
Это и Гаплогруппа S (B254) - единственные первичные субклады K2b1, также известная как гаплогруппа MS.
Филогенетическая структура
Это филогенетическое дерево субкладов гаплогрупп основано в первую очередь на деревьях, опубликованных ISOGG в 2016 г. (ISOGG 2016 ) и YCC в 2008.(Карафет 2008 )
- M * (P256)
- M1 (M4, M5 / P73, M106, M186, M189, M296, P35)
- M1a(P34_1, P34_2, P34_3, P34_4, P34_5)
- M1a1 (P51)
- M1a2 (P94)
- M1b (P87)
- M1b1 (M104_1 / P22_1, M104_2 / P22_2)
- M1b1a (M16)
- M1b1b (M83)
- M1b1 (M104_1 / P22_1, M104_2 / P22_2)
- M1a(P34_1, P34_2, P34_3, P34_4, P34_5)
- M2 (M353, M387)
- M2a (M177 / SRY9138)
- M3 (P117, P118)
- M1 (M4, M5 / P73, M106, M186, M189, M296, P35)
Распределение
M * (M-P256 *)
Парагруппа M-P256 * встречается с низкой заболеваемостью в Новая Гвинея (6,3%) и Флорес (2.5%).[1]
М1 (М-М4)
| Гаплогруппа M-M4 | |
|---|---|
| Возможное время происхождения | 8 200 [3 800–20 600] лет назад (Кайзер 2003 ) |
| Возможное место происхождения | Юго-Восточная Азия - Меланезия[нужна цитата ] |
| Предок | M-P256 |
| Определение мутаций | M4, M5 / P73, M106, M186, M189, M296, P35[нужна цитата ] |
Часто встречается в Новая Гвинея и Меланезия, с умеренным распространением в соседних частях Индонезия, Микронезия, и Полинезия.
- Уна 100% (Кайзер 2003 )
- Кетенгбан 100% (Кайзер 2003 )
- Авю 100% (Кайзер 2003 )
- Читак 86% (Кайзер 2003 )
- Асмат 75% (Кайзер 2003 )
- Западное Папуа
- низменности / побережье 77,5% (Кайзер 2003 )
- высокогорье 74,5% (Кайзер 2003 )
- Комбай /Коровай 46% (Кайзер 2003 )
- Папуа - Новая Гвинея
- побережье 29% (Кайзер 2003 )
- высокогорье 35,5% (Кайзер 2003 )
- Толай (Новая Британия) 31% (Кайзер 2003 )
- Острова Тробриан 30% (Кайзер 2003 )
- Малуку (Молуккские острова) 21% (Кайзер 2003 )
- Жители островов Торресова пролива (Австралия): до 2,0%, то есть 0,9% выборок, когда 45% от общего количества считались «некоренными».Нэгл 2015 )
Крайний географический выброс был явно выявлен в исследовании 2012 года, в котором сообщалось о Хазара человек из Мазари-Шариф, Афганистан, с M1 среди выборки из 60 мужчин из Мазари-Шарифа (Хабер 2012 ). У этого хазарейца был SNP M186 (который считается эквивалентом M4).
| Старые имена (YCC 2002/2008) | М-М4 |
| Джоблинг и Тайлер-Смит 2000 | 24 |
| Андерхилл 2000 | VIII |
| Молот 2001 | 1U |
| Карафет 2001 | 37 |
| Семино 2000 | Eu16 |
| Вс 1999 | H17 |
| Капелли 2001 | E |
| YCC 2002 (от руки) | М * |
| YCC 2005 (от руки) | M |
| YCC 2008 (от руки) | M1 |
| YCC 2010r (от руки) | M1 |
M1a (M-P34)
M1a (M-P34) - наиболее часто встречающаяся гаплогруппа ДНК Y-хромосомы в Западная Новая Гвинея. Он также встречается с умеренной частотой в соседних частях Индонезия (Малуку, Нуса Тенгара ) и повсюду Папуа - Новая Гвинея, включая прибрежные острова (Карафет 2005 и Кайзер 2008).
| Старые имена (YCC 2002/2008) | M-P34 |
| Джоблинг и Тайлер-Смит 2000 | 24 |
| Андерхилл 2000 | VIII |
| Молот 2001 | 1U |
| Карафет 2001 | 37 |
| Семино 2000 | Eu16 |
| Вс 1999 | H17 |
| Капелли 2001 | E |
| YCC 2002 (от руки) | M1 |
| YCC 2005 (от руки) | M1 |
| YCC 2008 (от руки) | M1a |
| YCC 2010r (от руки) | M1a |
M1b (M-P87)
M1b M-P87 (xM104 / P22) был обнаружен примерно в 18% (20/109) пула образцов из Новая Ирландия, примерно 12% (5/43) образца Лавонгай из Нью-Ганновер, примерно 5% (19/395) пула образцов из Новая Британия (и, в частности, примерно в 24% (15/63) Бейнинг из Восточная Новая Британия ), в одной Сапоса человек с севера Бугенвиль, а у другого человека с северного побережья Папуа - Новая Гвинея (Scheinfeldt 2006 ).
Субклад M1b1 (M104_1 / P22_1, M104_2 / P22_2) часто встречается в популяциях Архипелаг Бисмарка и Остров Бугенвиль, с умеренным распространением в Новая Гвинея, Фиджи, Тонга, Восточная Футуна, и Самоа. (Кайзер 2008 и Scheinfeldt 2006).
| Старые имена (YCC 2002/2008) | M-P22 |
| Джоблинг и Тайлер-Смит 2000 | 24 |
| Андерхилл 2000 | VIII |
| Молот 2001 | 1U |
| Карафет 2001 | 38 |
| Семино 2000 | Eu16 |
| Вс 1999 | H17 |
| Капелли 2001 | E |
| YCC 2002 (от руки) | M2 * |
| YCC 2005 (от руки) | M2a |
| YCC 2008 (от руки) | M1b1 |
| YCC 2010r (от руки) | M1b1 |
M2 (M-M353)
Обнаружен с низкой частотой в Фиджи и Восточная Футуна (Кайзер 2006 ).
Субклад M2a (M-M177 a.k.a. M-SRY9138) находится в одном Насиой человек с восточного побережья Бугенвиль и у одного человека из Провинция Малаита из Соломоновы острова (Кокс 2006 ).
Исторические названия M-SRY9138 (a.k.a. M-M177) из рецензируемой литературы.
| Старые имена (YCC 2002/2008) | K-SRY9138 / M-SRY9138 AKA M-M177 |
| Джоблинг и Тайлер-Смит 2000 | 23 |
| Андерхилл 2000 | VIII |
| Молот 2001 | 1E |
| Карафет 2001 | 25 |
| Семино 2000 | Eu16 |
| Вс 1999 | H5 |
| Капелли 2001 | F |
| YCC 2002 (от руки) | K1 |
| YCC 2005 (от руки) | K1 |
| YCC 2008 (от руки) | M2a |
| YCC 2010r (от руки) | M2a |
M3 (M-P117)
M3 (P117, P118) часто встречается в популяциях Новая Британия, а также иногда наблюдается в северных Бугенвиль, Фиджи, и Восточная Футуна (Кайзер 2008 и Scheinfeldt 2006).
Предыдущая филогенетическая история
До 2002 года в академической литературе существовало по крайней мере семь систем именования Филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (стенография) | (α) | (β) | (γ) | (δ) | (е) | (ζ) | (η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M4 | 24 | VIII | 1U | 37 | Eu16 | H17 | E | М * | M | M1 | M1 | - | - | - | - | - | - | - |
| M-P34 | 24 | VIII | 1U | 37 | Eu16 | H17 | E | M1 | M1 | M1a | M1a | - | - | - | - | - | - | - |
| M-P22 / M-M104 | 24 | VIII | 1U | 38 | Eu16 | H17 | E | M2 * | M2a | M1b1 | M1b1 | - | - | - | - | - | - | - |
| М-М16 | 24 | VIII | 1U | 39 | Eu16 | H17 | E | M2a | M2a1 | M1b1a | M1b1a | - | - | - | - | - | - | - |
| M-M83 | 24 | VIII | 1U | 38 | Eu16 | H17 | E | M2b | M2a2 | M1b1b | M1b1b | - | - | - | - | - | - | - |
| K-SRY9138 / M-SRY9138 | 23 | VIII | 1E | 25 | Eu16 | H5 | F | K1 | K1 | M2a | M2a | - | - | - | - | - | - | - |
- Источники
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
В статье Карафета 2008 г. внесен ряд изменений по сравнению с предыдущей. 2006 ISOGG дерево. До открытия маркера P256 текущая подгруппа M-M4 (определяемая маркером M4) ранее представляла всю гаплогруппу M-P256; и подгруппы M2 и M3 ранее классифицировались как подгруппы K1 и K7 родительского Гаплогруппа K.[нужна цитата ]
Рекомендации
Сноски
Процитированные работы
- Cox, Murray P .; Миразон Лар, Марта (2006). «Разнообразие Y-хромосомы обратно пропорционально языковой принадлежности в парных австронезийских и папуасоязычных сообществах Соломоновых островов». Американский журнал биологии человека. 18 (1): 35–50. Дои:10.1002 / ajhb.20459. PMID 16378340. S2CID 4824401.
- Габер, Марк; Platt, Daniel E .; Ашрафиан Бонаб, Мазиар; Юханна, Соня С .; Сориа-Эрнанц, Дэвид Ф .; Мартинес-Крус, Бегонья; Дуайхи, Бушра; Гассиб-Саббаг, Мишелла; Рафатпанах, Хошанг (2012). Кайзер, Манфред (ред.). «Этнические группы Афганистана разделяют наследие Y-хромосомы, структурированное историческими событиями». PLOS ONE. 7 (3): e34288. Bibcode:2012PLoSO ... 734288H. Дои:10.1371 / journal.pone.0034288. ЧВК 3314501. PMID 22470552.
- Карафет, Татьяна М .; Lansing, J. S .; Редд, Алан Дж .; Уоткинс, Джозеф С .; Сурата, С. П. К .; Arthawiguna, W.A .; Майер, Лаура; Бамшад, Майкл; Джорд, Линн Б. (2005). "Балийская перспектива Y-хромосомы на заселение Индонезии: генетический вклад от донеолитических охотников-собирателей, австронезийских фермеров и индийских торговцев" (PDF). Человеческая биология. 77 (1): 93–114. Дои:10.1353 / ступица.2005.0030. HDL:1808/13586. PMID 16114819. S2CID 7953854.
- Карафет, Т. М .; Mendez, F. L .; Meilerman, M. B .; Андерхилл, П. А .; Zegura, S.L .; Хаммер, М. Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Геномные исследования. 18 (5): 830–8. Дои:10.1101 / gr.7172008. ЧВК 2336805. PMID 18385274.
- Кайзер, Манфред; Брауэр, Силке; Вайс, Гюнтер; Schiefenhövel, Wulf; Андерхилл, Питер; Шен, Пейдун; Эфнер, Питер; Томмазео-Понцетта, Мила; Стоункинг, Марк (2003). «Уменьшение Y-хромосомы, но не митохондриальной ДНК, разнообразие в человеческих популяциях из Западной Новой Гвинеи». Американский журнал генетики человека. 72 (2): 281–302. Дои:10.1086/346065. ЧВК 379223. PMID 12532283.
- Kayser, M .; Брауэр, S; Cordaux, R; Касто, А; Лао, О; Животовский, Л.А.; Мойз-Фори, К; Рутледж, РБ; Schiefenhoevel, W. (2006). «Меланезийское и азиатское происхождение полинезийцев: мтДНК и градиенты Y-хромосомы через Тихий океан». Молекулярная биология и эволюция. 23 (11): 2234–44. Дои:10.1093 / molbev / msl093. PMID 16923821.
- Kayser, M .; Choi, Y .; Van Oven, M .; Мона, С .; Брауэр, С .; Трент, Р. Дж .; Suarkia, D .; Schiefenhovel, W .; Стоункинг, М. (2008). «Влияние австронезийской экспансии: данные по мтДНК и разнообразию Y-хромосомы на Адмиралтейских островах в Меланезии». Молекулярная биология и эволюция. 25 (7): 1362–74. Дои:10.1093 / molbev / msn078. PMID 18390477.
- Scheinfeldt, L .; Friedlaender, F; Фридлендер, Дж; Латам, К; Коки, G; Карафет, Т; Молоток, М; Лоренц, Дж (2006). «Неожиданное изменение хромосомы NRY в Меланезии на Северном острове». Молекулярная биология и эволюция. 23 (8): 1628–41. Дои:10.1093 / molbev / msl028. PMID 16754639.
- Nagle, N .; и другие. (2015). «Древность и разнообразие аборигенных австралийских Y-хромосом». Американский журнал физической антропологии. 159 (3): 367–81. Дои:10.1002 / ajpa.22886. PMID 26515539.
внешняя ссылка
Смотрите также
- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогения
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этнической группе
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||