BRCA2 и BRCA2 (/ˌбрækəˈтu/[5]) человек ген и это белок продукт соответственно. Официальный символ (BRCA2, курсив для гена, ненастоящий для белка) и официальное название (первоначально рак груди 2; В данный момент BRCA2, связанный с репарацией ДНК) находятся поддерживается посредством Комитет по номенклатуре генов HUGO. Один альтернативный символ, FANCD1, признает его связь с Белок FANC сложный. Ортологи, стилизованный Brca2 и Brca2, распространены в других позвоночное животное разновидность.[6][7]BRCA2 человек ген-супрессор опухоли[8][9] (в частности, ген-смотритель ), встречается у всех людей; это белок, также называемый синонимбелок предрасположенности к раку груди 2 типа, отвечает за восстановление ДНК.[10]
BRCA2 и BRCA1 в норме экспрессируются в клетках грудь и другие ткани, где они помогают восстановить поврежденные ДНК или уничтожить клетки, если ДНК не может быть восстановлена. Они занимаются ремонтом хромосомный повреждение играет важную роль в безошибочности ремонт ДНК двухрядные разрывы.[11][12] Если BRCA1 или BRCA2 повреждены BRCA мутация, поврежденная ДНК не восстанавливается должным образом, и это увеличивает риск рак молочной железы.[13][14]BRCA1 и BRCA2 были описаны как «гены предрасположенности к раку груди» и «белки предрасположенности к раку груди». Преобладающий аллель имеет нормальную функцию подавления опухоли, тогда как высокий пенетрантность Мутации в этих генах вызывают потерю функции подавления опухоли, что коррелирует с повышенным риском рака груди.[15]
Ген BRCA2 расположен на длинном (q) плече хромосома 13 в позиции 12.3 (13q12.3).[16] Человеческий эталонный ген BRCA 2 содержит 27 экзонов, а кДНК имеет 10 254 пары оснований.[17] кодирует белок из 3418 аминокислот.[18][19]
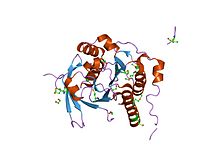
Рекомбинационная репарация двухцепочечных повреждений ДНК - некоторые ключевые шаги.Банкомат (Банкомат) протеинкиназа который набирается и активируется Двухцепочечные разрывы ДНК. Двухцепочечные повреждения ДНК также активируют Основной комплекс анемии Фанкони (FANCA / B / C / E / F / G / L / M).[20] Основной комплекс ТВС моноубиквитинаты последующие нацелены на FANCD2 и FANCI.[21] АТМ активирует (фосфорилирует) ЧЕК2 и FANCD2[22] CHEK2 фосфорилирует BRCA1.[23] Убихинированные комплексы FANCD2 с BRCA1 и RAD51.[24] В PALB2 белок действует как концентратор,[25] объединяя BRCA1, BRCA2 и RAD51 в месте двухцепочечного разрыва ДНК, а также связывается с RAD51C, членом комплекса паралогов RAD51 RAD51B -RAD51C -RAD51D -XRCC2 (BCDX2). Комплекс BCDX2 отвечает за рекрутирование или стабилизацию RAD51 в местах повреждения.[26]RAD51 играет важную роль в гомологичный рекомбинационный репарация ДНК во время репарации двухцепочечного разрыва. В этом процессе имеет место АТФ-зависимый обмен цепями ДНК, при котором одна цепь вторгается в спаренные по основанию цепи гомологичных молекул ДНК. RAD51 участвует в поиске гомологии и стадиях спаривания цепей этого процесса.
Хотя структуры BRCA1 и гены BRCA2 очень разные, по крайней мере, некоторые функции взаимосвязаны. В белки сделано обоими гены необходимы для восстановления поврежденной ДНК (см. рисунок этапов рекомбинационной репарации). BRCA2 связывает одноцепочечную ДНК и напрямую взаимодействует с рекомбиназой. RAD51 стимулировать[27] и поддерживать [28] нить вторжения, жизненно важный шаг гомологичная рекомбинация. Локализация RAD51 в двухцепочечном разрыве ДНК требует образования комплекса BRCA1-PALB2-BRCA2. PALB2 (партнер и локализатор BRCA2)[29] могут действовать синергетически с химерой BRCA2 (называемой пикколо или piBRCA2), чтобы дополнительно способствовать инвазии цепи.[30] Эти разрывы могут быть вызваны естественным и медицинским излучением или другим воздействием окружающей среды, но также возникают, когда хромосомы обмениваются генетическим материалом во время особого типа деления клеток, в результате которого образуются сперматозоиды и яйцеклетки (мейоз ). Двухцепочечные разрывы также возникают во время восстановления поперечных связей ДНК. Восстанавливая ДНК, эти белки играют роль в поддержании стабильности человеческий геном и предотвратить опасные перестройки генов, которые могут привести к гематологическим и другим видам рака.
Было показано, что BRCA2 играет решающую роль в защите от MRE11 -зависимая нуклеолитическая деградация обратные вилки которые формируются во время Репликация ДНК остановка вилки (вызванная такими препятствиями, как мутации, интеркалирующие агенты и т. д.).[31]
Подобно BRCA1, BRCA2, вероятно, регулирует активность других генов и играет критическую роль в развитии эмбриона.
Определенные варианты гена BRCA2 увеличивают риск рак молочной железы как часть синдром наследственного рака груди и яичников. Исследователи идентифицировали сотни мутаций в гене BRCA2, многие из которых вызывают повышенный риск рака. Мутации BRCA2 обычно представляют собой вставки или делеции небольшого числа пар оснований ДНК в гене. В результате этих мутаций белковый продукт гена BRCA2 является ненормальным и не функционирует должным образом. Исследователи считают, что дефектный белок BRCA2 не может исправить повреждение ДНК, которое происходит по всему геному. В результате увеличивается количество мутаций из-за подверженных ошибкам транслезионный синтез повреждение ДНК в прошлом, и некоторые из этих мутаций могут вызывать неконтролируемое деление клеток и образование опухоли.
Люди, у которых есть две мутировавшие копии гена BRCA2, имеют один тип Анемия Фанкони. Это состояние вызвано чрезвычайно низким уровнем белка BRCA2 в клетках, что способствует накоплению поврежденной ДНК. Пациенты с анемией Фанкони склонны к нескольким типам лейкемия (тип рака клеток крови); солидные опухоли, особенно головы, шеи, кожи и репродуктивных органов; и подавление костного мозга (снижение производства клеток крови, что приводит к анемия ). У женщин, унаследовавших дефектный ген BRCA1 или BRCA2, риск рака груди и яичников настолько высок и кажется настолько избирательным, что многие носители мутации предпочитают проходить профилактическую операцию. Было много предположений, объясняющих столь явно поразительную тканевую специфичность. Основные факторы, определяющие место возникновения BRCA1- и BRCA2-ассоциированного наследственного рака, связаны с тканевой специфичностью патогена рака, агента, вызывающего хроническое воспаление, или канцерогена. Ткань-мишень может иметь рецепторы патогена, избирательно подвергаться действию канцерогенов и инфекционного процесса. Врожденный геномный дефицит нарушает нормальные реакции и усугубляет восприимчивость к болезням органов-мишеней. Эта теория также соответствует данным для нескольких супрессоров опухолей помимо BRCA1 или BRCA2. Основным преимуществом этой модели является то, что она предполагает, что помимо профилактической хирургии есть еще несколько вариантов.[32]
Помимо рака груди у мужчин и женщин, мутации в BRCA2 также приводят к повышенному риску яичник, Фаллопиевых труб, предстательная железа и панкреатический рак. В некоторых исследованиях мутации в центральной части гена были связаны с более высоким риском рак яичников и меньший риск рак простаты чем мутации в других частях гена. Несколько других типов рака также наблюдались в некоторых семьях с мутациями BRCA2.
В целом, сильно унаследованные генные мутации (включая мутации в BRCA2) составляют лишь 5-10% случаев рака груди; конкретный риск заболеть раком груди или другим раком для любого, у кого есть мутация BRCA2, зависит от многих факторов.[33]
Методы диагностики вероятности наличия у пациента мутаций в BRCA1 и BRCA2 заболели раком были покрыты патенты принадлежит или контролируется Мириады генетики.[37][38] Бизнес-модель Myriad, предусматривающая эксклюзивное предложение диагностических тестов, вела от начала Myriad как стартапа в 1994 году до публичной компании с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году;[39] это также привело к разногласиям по поводу высоких цен на тесты и недоступности дополнительных мнений из других диагностических лабораторий, что, в свою очередь, привело к Ассоциация молекулярной патологии против Myriad Genetics иск.[40]
Мутации BRCA2 зародышевой линии и эффект основателя
Все мутации BRCA2 зародышевой линии, идентифицированные на сегодняшний день, являются унаследованными, что предполагает возможность большого эффекта «основателя», при котором определенная мутация является общей для четко определенной группы населения и теоретически может быть прослежена до общего предка. Учитывая сложность скрининга мутаций BRCA2, эти общие мутации могут упростить методы, необходимые для скрининга мутаций в определенных популяциях. Анализ мутаций, которые происходят с высокой частотой, также позволяет изучить их клиническое проявление.[41] Яркий пример мутации-основателя обнаружен в Исландии, где одна мутация BRCA2 (999del5) является причиной практически всех семейств рака груди / яичников.[42][43] Эта мутация со сдвигом рамки считывания приводит к сильно усеченному белку. В большом исследовании, в котором изучались сотни больных раком и контрольная группа, мутация 999del5 была обнаружена у 0,6% населения в целом. Следует отметить, что в то время как 72% пациентов, которые были признаны носителями, имели умеренный или сильный семейный анамнез рака груди, 28% имели небольшой семейный анамнез заболевания или его отсутствие. Это убедительно свидетельствует о наличии модифицирующих генов, которые влияют на фенотипическое выражение этой мутации, или, возможно, на взаимодействие мутации BRCA2 с факторами окружающей среды. Дополнительные примеры мутаций-основателей в BRCA2 приведены в таблице ниже.
Это динамический список и, возможно, никогда не сможет удовлетворить определенные стандарты полноты. Вы можете помочь добавление недостающих предметов с надежные источники.
На заводе Arabidopsis thaliana потеря BRCA2 гомолог AtBRCA2 вызывает серьезные дефекты у мужчин мейоз и в развитии женского гаметоцит.[59] Белок AtBRCA2 необходим для правильной локализации синаптонемный комплекс белок AtZYP1 и рекомбиназы AtRAD51 и AtDMC1. Кроме того, AtBRCA2 необходим для правильного мейотического синапса. Таким образом, AtBRCA2, вероятно, важен для мейотической рекомбинации. Похоже, что AtBRCA2 действует во время мейоза, чтобы контролировать этапы однонитевой инвазии, опосредованные AtRAD51 и AtDMC1, происходящие во время мейоза. гомологичный рекомбинационный восстановление повреждений ДНК.[59]
Мыши, продуцирующие усеченные версии BRCA2, жизнеспособны, но бесплодны.[64] Крысы с мутантом BRCA2 обладают фенотипом задержки роста и бесплодием у обоих полов.[65] Асперматогенез у этих мутантных крыс происходит из-за нарушения гомологичного синапсиса хромосом во время мейоза.
BRC повторяющиеся последовательности
DMC1 (ДНК-мейотическая рекомбиназа 1) представляет собой мейоз конкретный гомолог RAD51 который опосредует обмен нитями во время гомологичный рекомбинационный ремонт. DMC1 способствует образованию продуктов инвазии цепи ДНК (совместных молекул) между гомологичными молекулами ДНК. DMC1 человека напрямую взаимодействует с каждой из серии повторяющихся последовательностей в белке BRCA2 (называемых повторами BRC), которые стимулируют образование совместной молекулы с помощью DMC1.[66] Повторы BRC соответствуют мотиву, состоящему из последовательности примерно из 35 высококонсервативных аминокислот, которые хотя бы один раз присутствуют во всех BRCA2-подобных белках. BRCA2 BRC-повторы стимулируют образование совместной молекулы, способствуя взаимодействию одноцепочечной ДНК (оцДНК) с DMC1.[66] ОцДНК в комплексе с DMC1 может спариваться с гомологичной оцДНК из другой хромосомы во время стадии синопсиса мейоз чтобы сформировать совместную молекулу, центральный этап в гомологичная рекомбинация. Таким образом, последовательности повторов BRC BRCA2, по-видимому, играют ключевую роль в рекомбинационной репарации повреждений ДНК во время мейотической рекомбинации.
В целом, похоже, что функция гомологичной рекомбинации во время мейоза заключается в восстановлении повреждений ДНК,[нужна цитата ] и что BRCA2 играет ключевую роль в выполнении этой функции.
Нейрогенез
BRCA2 требуется в мыши для нейрогенез и подавление медуллобластома.[67] Утрата '' BRCA2 '' глубоко влияет на нейрогенез, особенно во время эмбрионального и постнатального нервного развития. Эти неврологические дефекты возникают из-за повреждения ДНК.[67]
Эпигенетический контроль
Эпигенетические изменения в экспрессии BRCA2 (вызывающие чрезмерную или недостаточную экспрессию) очень часто встречаются при спорадических формах рака (см. Таблицу ниже), тогда как мутации в BRCA2 встречаются редко.[68][69][70]
При немелкоклеточном раке легкого BRCA2 эпигенетически подавляется гиперметилированием промотора.[71] В этом случае гиперметилирование промотора в значительной степени связано с низким мРНК экспрессия и низкая экспрессия белка, но не с потерей гетерозиготности гена.
При спорадическом раке яичников наблюдается противоположный эффект. Промотор BRCA2 и области 5'-UTR содержат относительно мало или совсем не содержат метилированных динуклеотидов CpG в опухолевой ДНК по сравнению с неопухолевой ДНК, и обнаружена значительная корреляция между гипометилированием и> 3-кратной сверхэкспрессией BRCA2.[72] Это указывает на то, что гипометилирование промотора BRCA2 и 5'-UTR регионы приводит к сверхэкспрессии мРНК BRCA2.
В одном сообщении указывалось на некоторый эпигенетический контроль экспрессии BRCA2 со стороны микроРНК miR-146a и miR-148a.[73]
Экспрессия BRCA2 при раке
В эукариоты, Белок BRCA2 играет важную роль в гомологичной рекомбинационной репарации. У мышей и людей BRCA2 в первую очередь опосредует упорядоченную сборку RAD51 на одноцепочечной (ss) ДНК, форме, которая активна для гомологичного спаривания и инвазии цепи.[74] BRCA2 также перенаправляет RAD51 от двухцепочечной ДНК и предотвращает диссоциацию от оцДНК.[74] Кроме того, четыре паралоги из RAD51, состоящий из РАД51Б (RAD51L1 ), RAD51C (RAD51L2), RAD51D (RAD51L3 ), XRCC2 образуют комплекс, называемый комплексом BCDX2 (см. рисунок: Рекомбинационная репарация ДНК). Этот комплекс участвует в наборе или стабилизации RAD51 в местах повреждения.[26] Комплекс BCDX2, по-видимому, действует, облегчая сборку или стабильность Нуклеопротеиновая нить RAD51. RAD51 катализирует перенос цепи между нарушенной последовательностью и ее неповрежденным гомологом, чтобы обеспечить повторный синтез поврежденной области (см. модели гомологичной рекомбинации ).
Некоторые исследования рака сообщают о чрезмерно выраженном BRCA2 в то время как другие исследования сообщают о недостаточном выражении BRCA2. По крайней мере, в двух сообщениях была обнаружена избыточная экспрессия в некоторых спорадических опухолях груди и недостаточная экспрессия в других спорадических опухолях молочной железы.[75][76] (см. таблицу).
Многие виды рака имеют эпигенетические недостатки в различных генах репарации ДНК (см. Частоты эпимутаций в генах репарации ДНК при раке ). Эти дефекты репарации, вероятно, вызывают увеличение нереставрированных повреждений ДНК. Чрезмерное выражение BRCA2 наблюдаемое при многих формах рака может отражать компенсаторную BRCA2 сверхэкспрессия и повышенная гомологичная рекомбинационная репарация, чтобы, по крайней мере, частично бороться с такими избыточными повреждениями ДНК. Egawa et al.[77] предполагают, что повышенная экспрессия BRCA2 может быть объяснена нестабильностью генома, часто наблюдаемой при раке, которая индуцирует экспрессию мРНК BRCA2 из-за повышенной потребности в BRCA2 для репарации ДНК.
Недостаточное выражение BRCA2 сам по себе приведет к увеличению количества неисправленных повреждений ДНК. Ошибки репликации после этих повреждений (см. транслезионный синтез ) приведет к увеличению количества мутаций и рака.
BRCA2 выражение при спорадических раковых заболеваниях
BRCA2 содержит 39 аминокислотаповторяет которые имеют решающее значение для привязки к RAD51 (ключевой белок в рекомбинационной репарации ДНК) и устойчивость к обработке метилметансульфонатом.[96][103][104][112]
Спиральный домен BRCA2 принимает спиральный структура, состоящая из четырехспирального кластерного ядра (альфа 1, альфа 8, альфа 9, альфа 10) и двух последовательных бета-шпилек (бета 1 - бета 4). Сегмент из 50 аминокислот, содержащий четыре коротких спирали (от альфа 2 до альфа 4), извилины по поверхности ядра структура. В BRCA2 спирали альфа 9 и альфа 10 упаковываются доменом OB1 BRCA2 через ван дер Ваальс контакты с участием гидрофобный и ароматические остатки, а также через боковая цепь и позвоночник водородные связи. Этот домен связывает 70-аминокислота DSS1 (удаленный при синдроме разделенной руки / расщепленной стопы) белок, который первоначально был идентифицирован как один из трех гены эта карта в 1.5-Мб локусудалено при наследственном синдроме порока развития.[110]
Домен BRCA OB1 предполагает складку OB, которая состоит из сильно изогнутой пятицепочечной бета-лист который замыкается, образуя бета-баррель. OB1 имеет неглубокую бороздку, образованную одной стороной изогнутого листа, и разграничен двумя петлями, одна между бета 1 и бета 2, а другая между бета 4 и бета 5, что позволяет использовать слабую однонитевую ДНК. привязка. Домен также связывает 70-аминокислота DSS1 (исключен при синдроме разделенной руки / разделенной стопы) белок.[110]
Домен BRCA OB3 предполагает складку OB, которая состоит из сильно изогнутой пятицепочечной бета-лист который замыкается, образуя бета-баррель. OB3 имеет ярко выраженную канавку, образованную одной стороной изогнутого листа, и разделен двумя петлями, одна между бета 1 и бета 2, а другая между бета 4 и бета 5, что позволяет оцДНКпривязка.[110]
Заявка на патент на изолированное BRCA1 ген и мутации, способствующие развитию рака, а также методы диагностики вероятности развития рака груди были поданы Университетом штата Юта, Национальным институтом наук об окружающей среде (NIEHS) и Мириады генетики в 1994 г .;[37] В течение следующего года Myriad в сотрудничестве с другими исследователями изолировали и секвенировали ген BRCA2 и идентифицировали соответствующие мутации, а первый патент на BRCA2 был подан в США Myriad и другими учреждениями в 1995 году.[36] Myriad является эксклюзивным лицензиатом этих патенты и принудил их в США против клинико-диагностических лабораторий.[40] Эта бизнес-модель привела к тому, что Myriad из стартапа в 1994 году превратилась в публичную компанию с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году;[39] это также привело к спорам по поводу высоких цен и невозможности получить второе мнение из других диагностических лабораторий, что, в свою очередь, привело к появлению вехи Ассоциация молекулярной патологии против Myriad Genetics иск.[40][113] Срок действия патентов истекает в 2014 году.
Питер Мелдрам, генеральный директор Myriad Genetics, признал, что Myriad имеет «другие конкурентные преимущества, которые могут сделать такое [патентное] обеспечение ненужным» в Европе.[114]
Правовые решения, касающиеся патентов BRCA1 и BRCA2, повлияют на сферу генетического тестирования в целом.[115] В июне 2013 г. Ассоциация молекулярной патологии против Myriad Genetics (№ 12-398), Верховный суд США единогласно постановили, что «природный сегмент ДНК является продуктом природы и не имеет права на патент только потому, что он был изолирован», что аннулировало патенты Myriad на гены BRCA1 и BRCA2. Однако Суд также постановил, что манипулирование геном с целью создания чего-то, чего не встречается в природе, все же может иметь право на патентную защиту.[116] В Федеральный суд Австралии пришли к противоположному выводу, подтвердив законность австралийского патента Myriad Genetics в отношении гена BRCA1 в феврале 2013 года,[117] но это решение обжаловано, и апелляция будет включать рассмотрение постановления Верховного суда США.[118]
^Уильямс-Джонс Б (2002). Продажа генетических тестов: последствия коммерческого тестирования brca в Канаде (Кандидат наук.). Университет Британской Колумбии.
^Кастильо П., Боглиоло М., Сурралес Дж. (2011). «Скоординированное действие путей анемии Фанкони и телеангиэктазии атаксии в ответ на окислительное повреждение». Ремонт ДНК (Amst.). 10 (5): 518–25. Дои:10.1016 / j.dnarep.2011.02.007. PMID21466974.
^ абПатент США 5837492, Tavtigian SV, Kamb A., Simard J, Couch F, Rommens JM, Weber BL, «Ген предрасположенности к раку молочной железы, сцепленный с хромосомой 13», выпущен 1998-11-17, переуступлен Myriad Genetics, Inc., Endo Recherche, Inc. , HSC Research & Development Limited Partnership, Попечители Пенсильванского университета
^ абПатент США 5747282, Skolnick HS, Goldgar DE, Miki Y, Swenson J, Kamb A, Harshman KD, Shattuck-Eidens DM, Tavtigian SV, Wiseman RW, Futreal PA, «7Q-сцепленный ген восприимчивости к раку груди и яичников», выпущенный 1998-05- 05, передан Myriad Genetics, Inc., Соединенные Штаты Америки в лице министра здравоохранения и социальных служб и Исследовательского фонда Университета Юты.
^Патент США 5837492, Tavtigian SV, Kamb A., Simard J, Couch F, Rommens JM, Weber BL, «Ген предрасположенности к раку молочной железы, сцепленный с хромосомой 13», выпущен 1998-11-17, переуступлен Myriad Genetics, Inc., Endo Recherche, Inc. , HSC Research & Development Limited Partnership, Попечители Пенсильванского университета
^ абThorlacius S, Olafsdottir G, Tryggvadottir L, Neuhausen S, Jonasson JG, Tavtigian SV, Tulinius H, Ogmundsdottir HM, Eyfjörd JE (1996). «Одна мутация BRCA2 в семьях рака молочной железы у мужчин и женщин из Исландии с различными фенотипами рака». Природа Генетика. 13 (1): 117–119. Дои:10.1038 / ng0596-117. PMID8673089. S2CID8443452.
^Нойхаузен С., Гилевски Т., Нортон Л., Тран Т., Макгуайр П., Свенсен Дж., Хэмпел Х., Борген П., Браун К., Сколник М., Шаттук-Эйденс Д., Джанвар С., Голдгар Д., Оффит К. (1996). «Рецидивирующие мутации BRCA2 6174delT у еврейских женщин-ашкенази, страдающих раком груди». Природа Генетика. 13 (1): 126–128. Дои:10.1038 / ng0596-126. PMID8673092. S2CID11909356.
^Verhoog LC, van den Ouweland AM, Berns E, van Veghel-Plandsoen MM, van Staveren IL, Wagner A, Bartels CC, Tilanus-Linthorst MM, Devilee P, Seynaeve C, Halley DJ, Niermeijer MF, Klijn JG, Meijers-Heijboer H (2001). «Значительные региональные различия в частоте различных мутаций BRCA1 / BRCA2 в 517 голландских семьях рака груди и / или яичников». Европейский журнал рака. 37 (16): 2082–2090. Дои:10.1016 / S0959-8049 (01) 00244-1. PMID11597388.
^Pääkkönen K, Sauramo S, Sarantaus L, Vahteristo P, Hartikainen A, Vehmanen P, Ignatius J, Ollikainen V, Kääriäinen H, Vauramo E, Nevanlinna H, Krahe R, Holli K, Kere J (2001). «Участие BRCA1 и BRCA2 в развитии рака груди в подгруппе населения Западной Финляндии». Генетическая эпидемиология. 20 (2): 239–246. Дои:10.1002 / 1098-2272 (200102) 20: 2 <239 :: AID-GEPI6> 3.0.CO; 2-Y. PMID11180449.
^Тонин П.Н., Мес-Массон А.М., Народ С.А., Гадирян П., Провансер Д. (1999). «Основатель мутаций BRCA1 и BRCA2 во французско-канадских случаях рака яичников, не выбранных для семейного анамнеза». Клиническая генетика. 55 (5): 318–324. Дои:10.1034 / j.1399-0004.1999.550504.x. PMID10422801. S2CID23931343.
^Осорио А., Робледо М., Мартинес Б., Себриан А., Сан Роман Дж. М., Альбертос Дж., Лобо Ф., Бенитес Дж. (1998). «Молекулярный анализ гена BRCA2 в 16 испанских семьях с раком груди / яичников». Clin. Genet. 54 (2): 142–7. Дои:10.1111 / j.1399-0004.1998.tb03717.x. PMID9761393. S2CID30388365.
^ абСилигер К., Дукович-Шульце С., Вурц-Вильдерсинн Р., Пахер М., Пухта Н. (2012). «BRCA2 является медиатором гомологичной рекомбинации, облегчаемой RAD51 и DMC1, у Arabidopsis thaliana». Новый Фитол. 193 (2): 364–75. Дои:10.1111 / j.1469-8137.2011.03947.x. PMID22077663.
^Kojic M, Kostrub CF, Buchman AR, Holloman WK (2002). «Гомолог BRCA2, необходимый для восстановления, рекомбинации и стабильности генома у Ustilago maydis». Мол. Клетка. 10 (3): 683–91. Дои:10.1016 / с1097-2765 (02) 00632-9. PMID12408834.
^Ко Э, Ли Дж, Ли Х (2008). «Существенная роль brc-2 в целостности хромосом половых клеток C. elegans». Мол. Клетки. 26 (6): 590–4. PMID18779660.
^Коннор Ф., Бертвистл Д., Ми П.Дж., Росс Г.М., Свифт С., Григорьева Е., Тибулевич В.Л., Эшворт А. (1997). «Онкогенез и дефект репарации ДНК у мышей с усекающей мутацией Brca2». Nat. Genet. 17 (4): 423–30. Дои:10.1038 / ng1297-423. PMID9398843. S2CID42462448.
^Тенг Д.Х., Богден Р., Митчелл Дж., Баумгард М., Белл Р., Берри С., Дэвис Т., Ха ПК, Керер Р., Джаммулапати С., Чен К., Оффит К., Сколник М. Х., Тавтиджан С. В., Джанвар С., Сведлунд Б., Вонг А. К. , Камб А. (1996). «Низкая частота мутаций BRCA2 при карциноме молочной железы и других видах рака». Nat. Genet. 13 (2): 241–4. Дои:10.1038 / ng0696-241. PMID8640236. S2CID9831745.
^Мики Ю., Катагири Т., Касуми Ф., Йошимото Т., Накамура И. (1996). «Анализ мутаций в гене BRCA2 при первичном раке груди». Nat. Genet. 13 (2): 245–7. Дои:10.1038 / ng0696-245. PMID8640237. S2CID3203046.
^Ланкастер Дж. М., Вустер Р., Мэнджон Дж., Фелан К. М., Кокран С., Гамбс С., Сил S, Барфут Р., Коллинз Н., Бигнелл Дж., Патель С., Хамуди Р., Ларссон К., Вайзман Р. У., Беркак А., Иглхарт Дж. Д., Маркс Дж. Р. , Эшворт А., Страттон MR, Futreal PA (1996). «Мутации BRCA2 при первичном раке груди и яичников». Nat. Genet. 13 (2): 238–40. Дои:10.1038 / ng0696-238. PMID8640235. S2CID26808443.
^Thike AA, Tan PH, Ikeda M, Iqbal J (2016). «Повышенная экспрессия ID4, сопровождающаяся накоплением мутантного p53 и потерей белков BRCA1 / 2 при тройном отрицательном раке молочной железы, отрицательно влияет на выживаемость». Гистопатология. 68 (5): 702–12. Дои:10.1111 / his.12801. PMID26259780. S2CID3566545.
^ абcdежДонг Й., Хакими М.А., Чен Х, Кумарасвами Э., Куч Н.С., Годвин А.К., Шихаттар Р. (ноябрь 2003 г.). «Регулирование BRCC, холоферментного комплекса, содержащего BRCA1 и BRCA2, с помощью субъединицы, подобной сигнаносоме, и ее роль в репарации ДНК». Мол. Клетка. 12 (5): 1087–99. Дои:10.1016 / S1097-2765 (03) 00424-6. PMID14636569.
^ абЧен Дж., Сильвер Д.П., Уолпита Д., Кантор С.Б., Газдар А.Ф., Томлинсон Дж., Коуч Ф.Дж., Вебер Б.Л., Эшли Т., Ливингстон Д.М., Скалли Р. (сентябрь 1998 г.). «Стабильное взаимодействие между продуктами генов супрессоров опухолей BRCA1 и BRCA2 в митотических и мейотических клетках». Мол. Клетка. 2 (3): 317–28. Дои:10.1016 / S1097-2765 (00) 80276-2. PMID9774970.
^Reuter TY, Medhurst AL, Waisfisz Q, Zhi Y, Herterich S, Hoehn H, Gross HJ, Joenje H, Hoatlin ME, Mathew CG, Huber PA (октябрь 2003 г.). «Двухгибридный скрининг дрожжей предполагает участие белков анемии Фанкони в регуляции транскрипции, передаче сигналов в клетках, окислительном метаболизме и клеточном транспорте». Exp. Cell Res. 289 (2): 211–21. Дои:10.1016 / S0014-4827 (03) 00261-1. PMID14499622.
^Футамура М., Аракава Х., Мацуда К., Катагири Т., Саджи С., Мики Ю., Накамура И. (март 2000 г.). «Возможная роль BRCA2 в митотической контрольной точке после фосфорилирования hBUBR1». Рак Res. 60 (6): 1531–5. PMID10749118.
^Хьюз-Дэвис Л., Хантсман Д., Руас М., Фукс Ф., Пока Дж., Чин С.Ф., Милнер Дж., Браун Л.А., Сюй Ф., Гилкс Б., Нильсен Т., Шульцер М., Чиа С., Рагаз Дж., Кан А., Лингер Л., Оздаг Х., Каттанео Э., Жорданова Э.С., Шууринг Э., Ю. Д. С., Венкитараман А., Пондер Б., Доэрти А., Апарисио С., Бентли Д., Тейе С., Понтинг С. П., Калдас С., Кузаридес Т. (ноябрь 2003 г.). «EMSY связывает путь BRCA2 со спорадическим раком груди и яичников». Клетка. 115 (5): 523–35. Дои:10.1016 / S0092-8674 (03) 00930-9. PMID14651845. S2CID18911371.
^Ю. Д. С., Сонода Е., Такеда С., Хуанг С. Л., Пеллегрини Л., Бланделл Т. Л., Венкитараман А. Р. (октябрь 2003 г.). «Динамический контроль рекомбиназы Rad51 путем самоассоциации и взаимодействия с BRCA2». Мол. Клетка. 12 (4): 1029–41. Дои:10.1016 / S1097-2765 (03) 00394-0. PMID14580352.