Тля - Aphid
| Тли | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Класс: | Насекомое |
| Порядок: | Hemiptera |
| Подотряд: | Sternorrhyncha |
| Инфразаказ: | Афидоморфа |
| Надсемейство: | Афидоидеи Жоффруа, 1762 |
| Семьи | |
| |
Тли маленькие сок -сосание насекомые и члены надсемейство Афидоидеи. Общие имена включают зеленушка и мошка,[а] хотя люди внутри одного вида могут сильно различаться по цвету. В группу входят пушистые белые шерстистая тля. Типичный жизненный цикл включает нелетающих самок, рожающих живых самок. нимфы - которая, возможно, уже беременна, - называют адаптационные ученые. телескопическая развертка - без участия самцов. Быстро созревая, самки обильно размножаются, поэтому количество этих насекомых быстро увеличивается. Крылатые самки могут развиваться позже в течение сезона, позволяя насекомым колонизировать новые растения. В регионах с умеренным климатом осенью наступает фаза полового размножения, когда насекомые часто зимуют в виде яиц.
Жизненный цикл некоторых видов включает чередование двух видов растений-хозяев, например, между однолетней культурой и древесным растением. Некоторые виды питаются только одним типом растений, в то время как другие являются универсальными, колонизируя многие группы растений. Описано около 5000 видов тлей, все они входят в семейство Тли. Около 400 из них встречаются на пищевых и волокнистых культурах, и многие из них являются серьезными вредителями сельское хозяйство и лесное хозяйство, а также раздражение для садовники. Так называемое молочное животноводство муравьи есть мутуалистический отношения с тлями, ухаживая за ними нектар, и защищая их от хищники.
Тля - одно из самых разрушительных насекомых-вредителей культурных растений в регионах с умеренным климатом. Они не только ослабляют растение, высасывая сок, но и действуют как векторов для вирусы растений и обезображивают декоративные растения отложениями нектар и последующий рост сажистая плесень. Из-за их способности быстро увеличиваться в численности на бесполое размножение и телескопического развития, они представляют собой очень успешную группу организмов с экологической точки зрения.[1]
Бороться с тлей непросто. Инсектициды не всегда дают надежные результаты, учитывая устойчивость к нескольким классам инсектицидов и тот факт, что тля часто питается нижней стороной листьев. В садовых масштабах довольно эффективны водяные струи и мыльные распылители. Естественные враги включают хищных божьи коровки, журчалка личинки, паразитические осы, личинки тли-мошки, крабовые пауки, златоглазка личинки и энтомопатогенные грибы. An комплексная борьба с вредителями стратегия использования биологическая борьба с вредителями может работать, но этого трудно добиться, кроме как в закрытых помещениях, таких как теплицы.
Распределение
Тля распространена по всему миру, но чаще всего встречается в умеренные зоны. В отличие от многих таксоны Видовое разнообразие тлей в тропиках значительно ниже, чем в зонах умеренного климата.[2] Они могут мигрировать на большие расстояния, в основном за счет пассивного рассеивания ветрами. Крылатая тля также может подниматься днем на высоту до 600 м, куда ее переносит сильный ветер.[3][4] Например, смородино-салатная тля, Насоновия рибиснигри, как полагают, распространился из Новая Зеландия к Тасмания около 2004 года из-за восточных ветров.[5] Тля также распространяется через перенос зараженных растительных материалов людьми, в результате чего некоторые виды почти космополитичный в их распространении.[6]
Эволюция


История окаменелостей
Тля и близкородственные Адельгиды и филлоксераны, вероятно, произошли от общего предка некоторых 280 миллион лет назад, в Ранняя пермь период.[8] Вероятно, они питались такими растениями, как Cordaitales или Cycadophyta. Из-за своего мягкого тела тля плохо окаменяет, и самый старый из известных ископаемое принадлежит к виду Triassoaphis Cubitus от Триасовый.[9] Однако иногда они застревают в экссудатах растений, которые затвердевают в янтарь. В 1967 году, когда профессор Оле Хейе написал свою монографию Исследования ископаемых тлей, около шестидесяти видов описано из триаса, Юрский период, Меловой и в основном Третичный периоды, с Балтийский янтарь вносит еще сорок видов.[10] Общее количество видов было невелико, но значительно увеличилось с появлением покрытосеменные 160 миллион лет назад, поскольку это позволило тлям специализироваться, а видообразование тлей идет рука об руку с разнообразием цветковых растений. Вероятно, самые ранние тли были многоядный, с участием монофагия развивается позже.[11] Было высказано предположение, что предки Adelgidae жил на хвойные породы в то время как те из Aphididae питались соком Подокарповые или Араукариевые которые пережили вымирание в конце мелового периода. Такие органы, как роговицы, не появились до мелового периода.[8][12] Одно исследование также предполагает, что предковые тли могли жить на коре покрытосеменных растений и что питание листьями может быть производная черта. В Lachninae имеют длинные части рта, которые подходят для жизни на коре, и было высказано предположение, что предок среднего мелового периода питался корой покрытосеменных деревьев, переключившись на листья хвойных растений-хозяев в конце мелового периода.[13] Phylloxeridae вполне может быть самым старым из до сих пор сохранившихся семейств, но их летопись окаменелостей ограничена Нижний миоцен Палеофиллоксера.[14]
Таксономия
Реклассификация в конце 20-го века среди Hemiptera сократила старый таксон Homoptera до двух подотрядов: Sternorrhyncha (тля, белокрылка, напольные весы, псиллиды и т. д.) и Auchenorrhyncha (цикады, цикадки, цикадки, цикадки и т. д.) с подотрядом Гетероптеры содержит большую группу насекомых, известных как настоящие ошибки. Инфраотряд Aphidomorpha в пределах Sternorrhyncha варьируется в зависимости от очертания, при этом особенно трудно выделить несколько групп ископаемых, но они включают Adelgoidea, Aphidoidea и Phylloxeroidea.[15] Некоторые авторы используют единое надсемейство Aphidoidea, внутри которого Phylloxeridae и Adelgidae также включены, в то время как у других есть Aphidoidea с сестринским надсемейством Phylloxeroidea, в которое входят Adelgidae и Phylloxeridae.[16] Реклассификация начала 21-го века существенно изменила классификацию семей внутри Aphidoidea: некоторые старые семьи были понижены до уровня подсемейства (например, Eriosomatidae ), и многие старые подсемейства были возведены в ранг семьи. Самые последние авторитетные классификации включают три надсемейства Adelgoidea, Phylloxeroidea и Aphidoidea. Aphidoidea - это одна большая семья. Тли что включает все ~ 5000[2] дошедшие до нас виды.[17]
Филогения
Внешний
Тли, адельгиды и филлоксериды очень тесно связаны между собой и все относятся к подотряду Sternorrhyncha, жукам, сосущим растения. Они либо помещены в надсемейство насекомых Aphidoidea.[18] или в суперсемейство Филлоксероидея который включает семейство Adelgidae и семейство Phylloxeridae.[11] Как и тля, филлоксера питается корнями, листьями и побегами виноградных растений, но в отличие от тли не производит нектар или рожок выделения.[19] Филлоксера (Daktulosphaira vitifoliae) - насекомые, вызвавшие Великий французский винный упадок этот опустошенный европейский виноградарство в 19 веке. Точно так же адельгиды или шерстистая хвойная тля также питаются флоэмой растений и иногда описываются как тли, но более правильно классифицируются как тлейоподобные насекомые, потому что у них нет хвоста или роговиц.[20]
Отношение к группам, особенно по отношению к ископаемым, сильно различается из-за трудностей в разрешении отношений. Наиболее современные методы лечения включают три суперсемейства, Adelogidea, Aphidoidea и Phylloxeroidea внутри отряда Aphidomorpha, а также несколько групп ископаемых[21] но у других обработок есть Aphidomorpha, содержащая Aphidoidea из семейств Aphididae, Phylloxeridae и Adelgidae; или Aphidomorpha с двумя надсемействами, Aphidoidea и Phylloxeroidea, последнее содержит Phylloxeridae и Adelgidae. В филогенетическое дерево Sternorrhyncha выводится из анализа малой субъединицы (18S) рибосомальная РНК.[22]
| Sternorrhyncha |
| |||||||||||||||||||||||||||||||||




Внутренний
Филогенетическое древо, основанное на Papasotiropoulos 2013 и Kim 2011, с дополнениями из Ortiz-Rivas и Martinez-Torres 2009, показывает внутреннюю филогения афидид.[23][24][25]
Было высказано предположение, что филогения групп тлей может быть выявлена путем изучения филогении их бактериальной эндосимбионты, особенно облигатный эндосимбионт Бухнера. Результаты зависят от предположения, что симбионты строго передается вертикально через поколения. Это предположение хорошо подтверждается доказательствами, и на основе исследований эндосимбионтов было предложено несколько филогенетических отношений.[26][27][28]
| Тли |
| |||||||||||||||||||||||||||||||||||||||||||||||||||








Анатомия

У большинства тлей мягкое тело, которое может быть зеленым, черным, коричневым, розовым или почти бесцветным. У тлей есть усики с двумя короткими широкими базальными сегментами и до четырех тонких терминальных сегментов. У них есть пара сложные глаза, с глазным бугорком позади и над каждым глазом, состоящий из трех линз (называемых триомматидиями).[11] Они питаются соком, используя сосательный ротовой аппарат, называемый стилеты заключенный в оболочку, называемую трибуна, который сформирован из модификаций нижняя челюсть и верхняя челюсть ротового аппарата насекомых.[29]
У них длинные, тонкие ноги с двусоставными, двоякогими. лапки. Большинство тлей бескрылые, но у многих видов в определенное время года образуются крылатые формы. У большинства тлей есть пара роговицы (сифункулы), брюшные трубы на дорсальной поверхности их пятого брюшного сегмента, через которые они выделяют капли быстро затвердевающей защитной жидкости[29] содержащий триацилглицерины, называемый роговым воском. Другие защитные соединения также могут быть произведены некоторыми видами.[20] У тлей над их ректальными отверстиями имеется хвостовидный выступ, называемый хвостом.[11][30]
Когда качество растения-хозяина ухудшается или условия становятся скученными, некоторые виды тлей производят крылатое потомство (крылатые ), которые могут распространяться на другие источники пищи. Ротовой аппарат или глаза могут быть маленькими или отсутствовать у некоторых видов и форм.[20]
Диета
Многие виды тлей являются монофагами (то есть питаются только одним видом растений). Другие, такие как зеленая персиковая тля, питаются сотнями видов растений многих видов. семьи. Около 10% видов в разное время года питаются разными растениями.[31]
Новое растение-хозяин выбирает крылатый взрослый с помощью визуальных сигналов с последующим обонянием с помощью антенн; если растение пахнет правильно, следующим действием будет прощупывание поверхности при посадке. Вставляют иглу и выделяют слюну, отбирают образец сока, можно попробовать ксилему и, наконец, исследуют флоэму. Слюна тли может подавлять механизмы запечатывания флоэмы и содержит пектиназы, облегчающие проникновение.[32] Растения, не являющиеся хозяевами, могут быть отклонены на любой стадии исследования, но перенос вирусов происходит на ранней стадии процесса исследования, во время введения слюны, поэтому растения, не являющиеся хозяевами, могут инфицироваться.[31]
Тля обычно пассивно питается сок из флоэма сосуды у растений, как и у многих других полужестких насекомых, таких как щитовки и цикады. Как только сосуд флоэмы прокалывается, сок, находящийся под давлением, выталкивается в пищевой канал тли. Иногда тля также заглатывает ксилема сок, который представляет собой более разбавленную диету, чем сок флоэмы, поскольку концентрация сахаров и аминокислот составляет 1% от таковых во флоэме.[33][34] Ксилемный сок находится под отрицательным гидростатическим давлением и требует активного всасывания, что предполагает важную роль в физиологии тли.[35] Поскольку после периода обезвоживания наблюдается заглатывание ксилемного сока, считается, что тля потребляет ксилемный сок для пополнения своего водного баланса; потребление разбавленного сока ксилемы, позволяющее тле регидратироваться.[36] Однако недавние данные показали, что тля потребляет больше ксилемного сока, чем ожидалось, и это особенно заметно, когда они не обезвожены и когда их плодовитость снижается. Это говорит о том, что тля и, возможно, все виды отряда Hemiptera, питающиеся флоэмным соком, потребляют ксилемный сок по причинам, не связанным с восполнением водного баланса.[37] Хотя тля пассивно поглощает сок флоэмы, который находится под давлением, они также могут втягивать жидкость при отрицательном или атмосферном давлении, используя цибариально-глоточный насосный механизм, расположенный в их голове.[38]
Потребление ксилемного сока может быть связано с осморегуляция.[37] Высокое осмотическое давление в желудке, вызванное высокой концентрацией сахарозы, может привести к переносу воды от гемолимфы в желудок, что приводит к гиперосмотическому стрессу и, в конечном итоге, к гибели насекомого. Тля избегает этой участи, осморегулируя через несколько процессов. Концентрация сахарозы непосредственно снижается за счет ассимиляции сахарозы на метаболизм и за счет синтеза олигосахариды из нескольких сахарозы молекулы, тем самым снижая концентрацию растворенного вещества и, следовательно, осмотическое давление.[39][40] Затем олигосахариды выводятся через падевую росу, что объясняет ее высокую концентрацию сахара, который затем может использоваться другими животными, такими как муравьи. Кроме того, вода переносится из задняя кишка, где осмотическое давление уже снижено, в желудок для разбавления содержимого желудка.[41] В конце концов, тля потребляет ксилемный сок, чтобы снизить осмотическое давление в желудке.[37] Все эти процессы действуют синергетически и позволяют тле питаться соком растений с высокой концентрацией сахарозы, а также адаптироваться к различным концентрациям сахарозы.[37]
Сок растений - несбалансированный рацион для тли, так как ему не хватает незаменимые аминокислоты, который тля, как и все животные, не может синтезировать, и обладает высокой осмотическое давление из-за его высокого сахароза концентрация.[34][42] Незаменимые аминокислоты поступают в организм тли из бактериальных эндосимбионты в специальных камерах, бактериоциты.[43] Эти симбионты перерабатывают глутамат, отходы метаболизма своего хозяина, в незаменимые аминокислоты.[44][45]
Каротиноиды и фотогетеротрофия
Некоторые виды тлей приобрели способность синтезировать красный цвет. каротиноиды от горизонтальный перенос генов от грибы.[46] Они единственные животные, кроме двухпятнистый паутинный клещ и восточный шершень с этой возможностью.[47] Используя свои каротиноиды, тля вполне может поглощать солнечную энергию и преобразовывать ее в форму, которую могут использовать их клетки, АТФ. Это единственный известный пример фотогетеротрофия у животных. В каротин пигменты тли образуют слой, расположенный близко к поверхности кутикулы, идеально подходящий для поглощения солнечного света. Возбужденные каротиноиды, кажется, снижают НАД до НАДН который окисляется в митохондрии для энергии.[48]
Размножение
Простейшая репродуктивная стратегия заключается в том, чтобы у тли был один хозяин круглый год. При этом он может чередоваться между сексуальный и бесполое поколение (голоциклическое) или, альтернативно, все молодняк может быть произведен партеногенез, яйца никогда не укладывается (анголоциклический). Некоторые виды могут иметь как голоциклические, так и анголоциклические популяции при различных обстоятельствах, но ни один из известных видов тлей не размножается исключительно половым путем.[50] Чередование полового и бесполого поколений могло происходить неоднократно.[51]
Однако размножение тли часто бывает более сложным и включает миграцию между разными растениями-хозяевами. Примерно у 10% видов наблюдается чередование древесный (основные хозяева), на которых зимуют тли и травянистый (вторичные) растения-хозяева, где летом обильно размножаются.[20][50] Некоторые виды могут образовывать касту солдат, другие виды демонстрируют обширную полифенизм в разных условиях окружающей среды, и некоторые могут контролировать соотношение полов своего потомства в зависимости от внешних факторов.[52]
Когда используется типичная изощренная репродуктивная стратегия, в начале сезонного цикла в популяции присутствуют только самки (хотя было обнаружено, что у некоторых видов тлей в это время есть и мужской, и женский пол). Из зимующих яиц, которые вылупляются весной, вылупляются самки, называемые фундаментальными матерями. В размножении обычно не участвуют самцы (партеногенез ) и приводит к живорождению (живородство ).[53] Живое молодняк образуется в результате псевдоплацентарного живорождения, которое представляет собой развитие яиц с дефицитом желтка, зародышей, питающихся тканью, выступающей в роли плаценты. Птенцы появляются из матери вскоре после вылупления.[54]
Яйца партеногенетически производятся без мейоз[55][53] и потомство является клональным по отношению к своей матери, поэтому все они женского пола (Thelytoky ).[11][54] Эмбрионы развиваются внутри матери. овариолы, которые потом рождают живых (уже вылупившихся) первых-возраст женские нимфы. Поскольку яйца начинают развиваться сразу после овуляции, взрослая самка может содержать развивающиеся нимфы-самки, внутри которых уже есть партеногенетически развивающиеся эмбрионы (т. Е. Они рождаются беременными). Эта телескопирование поколений позволяет тле быстро увеличиваться в численности. Потомство похоже на своего родителя во всем, кроме размера. Таким образом, питание женщины может повлиять на размер тела и рождаемость более двух поколений (дочерей и внучок).[11][56][57]
Этот процесс повторяется в течение всего лета, производя несколько поколений, которые обычно живут от 20 до 40 дней. Например, некоторые виды капустной тли (например, Brevicoryne brassicae ) может производить до 41 поколения самок за сезон. Таким образом, одна самка, вылупившаяся весной, теоретически может произвести миллиарды потомков, если все они выживут.[58]

Осенью тля размножается половым путем и откладывать яйца. Факторы окружающей среды, такие как изменение фотопериод и температура или, возможно, более низкое количество или качество пищи заставляет самок партеногенетически производить половых самок и самцов.[55] Самцы генетически идентичны своим матерям, за исключением того, что у тлей есть Система определения пола X0, у них на один меньше половая хромосома.[55] У этих половых тлей могут отсутствовать крылья или даже ротовой аппарат.[20] Половые самки и самцы спариваются, а самки откладывают яйца, которые развиваются вне матери. Яйца переживают зиму и следующей весной вылупляются крылатые (крыловидные) или бескрылые самки. Это происходит, например, в жизненный цикл из розовая тля (Macrosiphum rosae), что можно считать типичным для семьи. Однако в теплой среде, например, в тропики или в теплица, тля может продолжать бесполое размножение в течение многих лет.[29]
Тля, размножающаяся бесполым путем партеногенез могут иметь генетически идентичное потомство крылатых и некрылых самок. Контроль сложен; некоторые тли в течение своего жизненного цикла чередуют генетический контроль (полиморфизм ) и экологический контроль (полифенизм ) изготовления крылатых или бескрылых форм.[59] Крылатое потомство, как правило, производится более обильно в неблагоприятных или стрессовых условиях. Некоторые виды производят крылатое потомство в ответ на низкое качество или количество пищи. например когда растение-хозяин начинает стареть.[60] Крылатые самки мигрируют, чтобы создать новые колонии на новом растении-хозяине. Например, яблочная тля (Афис поми), производя много поколений бескрылых самок, дает начало крылатым формам, которые летают на другие ветви или деревья его типичного пищевого растения.[61] Тля, на которую нападает божьи коровки, златоглазки, паразитоидные осы, или другие хищники могут изменить динамику производства своего потомства. Когда на тлю нападают эти хищники, тревожные феромоны, в частности бета-фарнезен, освобождаются от рожки. Эти тревожные феромоны вызывают несколько поведенческих модификаций, которые, в зависимости от вида тли, могут включать уход и падение с растения-хозяина. Кроме того, восприятие феромона тревоги может побудить тлей произвести крылатое потомство, которое может покинуть растение-хозяин в поисках более безопасного места для питания.[62] Вирусные инфекции, которые могут быть чрезвычайно опасными для тлей, также могут привести к образованию крылатого потомства.[63] Например, Дензовирус Инфекция негативно влияет на розовую яблочную тлю (Дизафис подорожник ) размножение, но способствует развитию крылатых тлей, которые могут легче передавать вирус новым растениям-хозяевам.[64] Кроме того, симбиотические бактерии, которые живут внутри тлей, также могут изменять репродуктивные стратегии тлей в зависимости от воздействия факторов окружающей среды.[65]

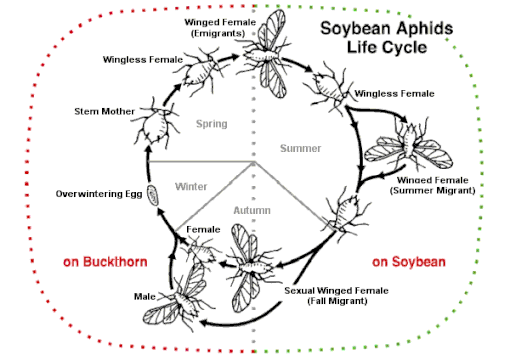
Осенью хозяин-чередование (гетеро ) виды тлей производят особое крылатое поколение, которое в течение половой части жизненного цикла летает к разным растениям-хозяевам. Производятся нелетающие женские и мужские половые формы, откладывающие яйца.[66] Некоторые виды, такие как Афис фаба (бобовая тля), Metopolophium dirhodum (розовая тля), Myzus persicae (персиково-картофельная тля) и Ропалосифум пади (черемуха-овсяная тля) - серьезные вредители. Зимуют на дереве или кустарнике основных хозяев; летом они мигрируют к своему вторичному хозяину на травянистом растении, часто урожае, затем осенью гинопары возвращаются на дерево. Другой пример - соевая тля (Афис глицины). По мере приближения осени соевые бобы начинают стареть снизу вверх. Тля выталкивается вверх и начинает давать крылатые формы, сначала самок, а затем самцов, которые улетают к основному хозяину. облепиха. Здесь они спариваются и зимуют как яйца.[49]
Экология
Муравьиный мутуализм


Некоторые виды муравьи выращивайте тлей, защищая их на растениях, где они кормятся, и поедая падь, тля высвобождает прекращения от их пищеварительные каналы. Это мутуалистические отношения, когда эти молочные муравьи доят тлей, поглаживая их своими усики.[b][67] Несмотря на мутуалистическое поведение, пищевое поведение тли изменяется из-за присутствия муравьев. Тля, сопровождаемая муравьями, как правило, увеличивает производство пади меньшими каплями с большей концентрацией аминокислот.[68]
Некоторые виды сельскохозяйственных муравьев собирают и хранят яйца тлей в своих гнездах на зиму. Весной муравьи переносят только что вылупившуюся тлю обратно на растения. Некоторые виды молочных муравьев (например, европейские желтый луговый муравей, Lasius flavus)[69] управлять большими стадами тлей, которые питаются корнями растений в колонии муравьев. Королевы, уезжающие, чтобы создать новую колонию, берут яйцо тли, чтобы основать новое стадо подземных тлей в новой колонии. Эти муравьи-земледельцы защищают тлю, отбиваясь от ее хищников.[67]

Интересный вариант взаимоотношений муравьев и тлей включает: бабочки Lycaenid и Мирмика муравьи. Например, Niphanda fusca бабочки откладывают яйца на растения, где муравьи пасут стада тлей. Яйца вылупляются как гусеницы которые питаются тлей. Муравьи не защищают тлей от гусениц, так как гусеницы производят феромон который заставляет муравьев обращаться с ними, как с муравьями, и уносить гусениц в их гнезда. Оказавшись там, муравьи кормят гусениц, которые в свою очередь производят медвяную росу для муравьев. Когда гусеницы достигают полного размера, они подползают ко входу в колонию и образуют коконы. Через две недели взрослые бабочки появляются и улетают. В этот момент муравьи нападают на бабочек, но у бабочек на крыльях есть липкое, похожее на шерсть вещество, которое выводит из строя челюсти муравьев, позволяя бабочкам улетать, не нанося им вреда.[70] Немного пчелы в хвойные леса собрать падевую тлю, чтобы сделать лесной мед.[29]
Другая муравейник желчная тля, Paracletus cimiciformis (Eriosomatinae), разработал сложную двойную стратегию, включающую две морфы одного и того же клона и Тетраморий муравьи. Тля круглой формы заставляет муравьев выращивать их, как и многие другие тли. Плоскоморфные тли бывают агрессивная имитация с "волк в овечьей шкуре Стратегия: в кутикуле у них есть углеводороды, которые имитируют кутикулу муравьев, и муравьи несут их в выводочную камеру муравьиного гнезда и выращивают их, как личинок муравьев. Оказавшись там, плоские морфы тли ведут себя как хищники, выпивая биологические жидкости личинок муравьев.[71]
Бактериальный эндосимбиоз
Эндосимбиоз с участием микроорганизмы часто встречается у насекомых, причем более 10% видов насекомых полагаются на внутриклеточные бактерии для своего развития и выживания.[72] Тля переносит вертикально передающийся (от родителя к потомству) облигатный симбиоз с участием Buchnera aphidicola, первичный симбионт, внутри специализированных клеток, бактериоциты.[73] Пять генов бактерий были перенесены в ядро тли.[74] Первоначальная ассоциация может иметь место у общего предка. От 280 до 160 миллион лет назад и позволил тле использовать новый экологическая ниша, питающиеся флоэмами-соками сосудистых растений. Б. афидикола обеспечивает своего хозяина незаменимыми аминокислотами, которые в низких концентрациях присутствуют в соке растений.[75] Метаболиты эндосимбионтов также выводятся с медвяной росой.[76] Стабильные внутриклеточные условия, а также эффект узкого места, возникающий при передаче нескольких бактерий от матери каждой нимфе, увеличивают вероятность передачи мутаций и делеций генов.[77][78] В результате размер Б. афидикола геном значительно сокращен по сравнению с его предполагаемым предком.[79] Несмотря на очевидную потерю факторы транскрипции в редуцированном геноме, экспрессия гена в высокой степени регулируется, о чем свидетельствует десятикратная вариация уровней экспрессии между разными генами в нормальных условиях.[80] Buchnera aphidicola транскрипция гена, хотя и не совсем понятный, считается, что он регулируется небольшим количеством глобальных регуляторов транскрипции и / или через поставки питательных веществ от тли-хозяина.[81]
Некоторые колонии тлей также содержат вторичных или факультативных (необязательно дополнительных) бактериальных симбионтов. Они передаются вертикально, а иногда и горизонтально (от одной линии к другой и, возможно, от одного вида к другому).[82][83] Пока что описана роль только некоторых вторичных симбионтов; Региелла инсектикола играет роль в определении диапазона растений-хозяев,[84][85] Гамильтонелла дефенса обеспечивает устойчивость к паразитоидам, но только тогда, когда он, в свою очередь, заражен бактериофаг APSE,[86][87] и Serratia symbiotica предотвращает вредное воздействие тепла.[88]
Хищники
Тлей поедают многие хищные птицы и насекомые. В кабинете на ферме в Северная Каролина, шесть видов воробьиные птицы съедали почти миллион тлей в день, причем главными хищниками были Американский щегол, тля составляет 83% ее рациона, а вечерний воробей.[89] Насекомые, нападающие на тлей, включают взрослых особей и личинок хищных божьих коровок, журчалка личинки, паразитические осы, тля мошка личинки, «львы тли» (личинки зеленые златоглазки ) и паукообразных, например, пауков-крабов. Среди божьих коровок, Myzia oblongoguttata диетолог, который питается только хвойной тлей, тогда как Adalia bipunctata и Coccinella septempunctata универсалы, питающиеся большим количеством видов. Яйца откладывают пачками, каждая самка откладывает несколько сотен. Самки журчалки откладывают несколько тысяч яиц. Взрослые особи питаются пыльцой и нектаром, а личинки обильно питаются тлей; Eupeodes венчик регулирует количество отложенных яиц в соответствии с размером колонии тлей.[90]


Тля часто заражается бактерии, вирусы, и грибы. На них влияет погода, например атмосферные осадки,[91] температура[92] и ветер.[93] К грибам, поражающим тлю, относятся: Неозигиты фрезении, Энтомофтора, Боверия бассиана, Metarhizium anisopliae и энтомопатогенные грибы, такие как Lecanicillium lecanii. Тля задевает микроскопические споры. Они прилипают к тле, прорастают и проникают сквозь кожу тли. Грибок растет у тли. гемолимфа. Примерно через три дня тля умирает, и грибок выпускает в воздух новые споры. Зараженная тля покрывается шерстяной массой, которая постепенно становится гуще, пока тля не скрывается. Часто видимый грибок убивает тлю, а является вторичной инфекцией.[91]
Тля легко погибает от неблагоприятных погодных условий, например от поздних весенних заморозков.[94] Избыточное тепло убивает симбиотические бактерии, от которых зависят некоторые тли, что делает их бесплодными.[95] Дождь предотвращает расселение крылатой тли и сбивает ее с растений и тем самым убивает их от ударов или голода.[91][96][97] но нельзя полагаться на борьбу с тлей.[98]
Защита от хищников

У большинства тлей мало защиты от хищников. Некоторые виды взаимодействуют с тканями растений, образуя желчь, аномальный отек растительной ткани. Тля может жить внутри галла, который обеспечивает защиту от хищников и непогоды. Известно, что некоторые виды тлей, вызывающих раздражение, производят специализированные «солдатские» формы, стерильные нимфы с защитными особенностями, которые защищают желчь от вторжения.[29][99][100] Например, рогатая тля Александра - это разновидность солдатской тли, которая экзоскелет и клешня -подобный ротовой аппарат.[70]:144 Шерстистая тля, Колофина клематис, имеет личинок «солдатиков» первой стадии, которые защищают колонию тлей, убивая личинок божьих коровок, журчалок и цветочного клопа. Anthocoris nemoralis забравшись на них и вставив их стилеты.[101]
Хотя тля не может летать большую часть своего жизненного цикла, она может избежать хищников и случайного проглатывания травоядными животными, уронив растение на землю.[102] Другие виды используют почву в качестве постоянной защиты, питаясь сосудистой системой корней и оставаясь под землей всю свою жизнь. Их часто сопровождают муравьи, так как они производят падь, и муравьи переносят их с растения на растение через свои туннели.[89]
Некоторые виды тли, известные как «шерстистые тли» (Eriosomatinae), выделяют «пушистый восковой налет» для защиты.[29] Капустная тля, Brevicoryne brassicae, изолирует вторичные метаболиты от своего хозяина, накапливает их и высвобождает химические вещества, которые производят сильный химическая реакция и сильный горчичное масло запах для отпугивания хищников.[103] Пептиды, вырабатываемые тлей, Тауматины, как полагают, придают им устойчивость к некоторым грибам.[104]
Одно время было распространено предположение, что рожки были источником пади, и это даже было включено в Краткий оксфордский словарь английского языка[105] и издание 2008 г. Всемирная книжная энциклопедия.[106] Фактически, выделения пади производятся из заднего прохода тли,[107] в то время как роговицы в основном производят защитные химические вещества, такие как воск. Также есть данные о роговом воске. привлечение хищников тли в некоторых случаях.[108]
Некоторые клоны Aphis craccivora достаточно токсичны для агрессивных и доминирующих хищных божьих коровок Harmonia axyridis локально подавить его, отдав предпочтение другим видам божьих коровок; токсичность в этом случае узко специфична для доминирующего вида хищников.[109]
Паразитоиды
Тли многочисленны и широко распространены и служат хозяевами большого числа паразитоиды, многие из них очень маленькие (около 0,1 дюйма (2,5 мм) в длину) паразитоидные осы.[110] Один вид, Афис руборум, например, является хозяином по крайней мере 12 видов ос-паразитоидов.[111] Паразитоиды интенсивно исследуются как агенты биологической борьбы, и многие из них используются в коммерческих целях для этой цели.[112]
Взаимодействие растения с тлей

Растения обеспечивают локальную и системную защиту от нападения тли. Молодые листья некоторых растений содержат химические вещества, препятствующие атаке, в то время как более старые листья утратили эту устойчивость, тогда как у других видов растений устойчивость приобретается более старыми тканями, а молодые побеги уязвимы. Было показано, что летучие продукты из пересаженного лука предотвращают нападение тли на соседние растения картофеля, способствуя производству терпеноиды, преимущество, используемое в традиционной практике посадка-компаньон, в то время как растения, соседствующие с зараженными растениями, показали усиленный рост корней за счет расширения надземных частей.[31] Дикий картофель, Solanum berthaultii, производит феромон тревоги тли, (E) -β-Фарнезен, как алломон, феромон для отражения атаки; эффективно отталкивает тлю Myzus persicae в диапазоне до 3 миллиметров.[113] S. berthaultii и другие виды дикого картофеля обладают дополнительной защитой от тли в виде железистых волосков, которые при разрыве тлей выделяют липкую жидкость, которая может иммобилизовать около 30% тлей, поражающих растение.[114]
У растений, пораженных тлей, могут быть различные симптомы, такие как снижение скорости роста, пятнистые листья, пожелтение, задержка роста, скрученные листья, потемнение, увядание, низкие урожаи и смерть. Удаление сока приводит к потере энергии у растений, а слюна тли токсична для растений. Тля часто передает вирусы растений своим хозяевам, например, картофель, хлопья, сахарная свекла, и цитрусовые.[29] Зеленая персиковая тля, Myzus persicae, является вектором для более чем 110 вирусов растений. Хлопковая тля (Тля gossypii) часто заражают сахарный тростник, папайя и арахис с вирусами.[20] В растениях, вырабатывающих фитоэстроген куместрол таких как люцерна, повреждение тлей связано с более высокими концентрациями куместрола.[115]

Покрытие растений медвяной росой может способствовать распространению грибков, которые могут повредить растения.[116][117] Медовая роса, производимая тлей, также снижает эффективность фунгицидов.[118]
Гипотеза о том, что кормление насекомыми может улучшить приспособленность растений, была выдвинута в середине 1970-х Оуэном и Вигертом. Было высказано мнение, что избыток пади будет питать почвенные микроорганизмы, в том числе азотфиксаторы. В среде с низким содержанием азота это может дать преимущество зараженному растению перед незараженным. Однако это не подтверждается данными наблюдений.[119]
Социальность
Некоторые тли проявляют некоторые черты эусоциальность, присоединяясь к насекомым, таким как муравьи, пчелы и термиты. Однако есть различия между этими половыми социальными насекомыми и клональными тлями, которые партеногенетически произошли от одной самки и имеют одинаковые геном. Около пятидесяти видов тлей, разбросанных среди близкородственных, чередующихся линий. Eriosomatinae и Hormaphidinae, имеют какой-то защитный морф. Это виды, создающие галлы, при этом колония живет и питается внутри галла, который они образуют в тканях хозяина. Среди клональной популяции этих тлей может быть несколько различных морфов, и это закладывает основу для возможной специализации функций, в данном случае - защитной касты. Морфы солдата в основном относятся к первой и второй стадиям, а третья стадия участвует в Eriosoma moriokense и только в Smythurodes Betae известны взрослые солдаты. Задние лапы солдат когтистые, сильно склеротизированы, а стилеты крепкие, что позволяет разрывать и раздавливать мелких хищников.[120] Личинки-солдаты - альтруисты, неспособные перейти к размножению взрослых особей, но постоянно действующие в интересах колонии. Еще одно требование для развития социальности - это галл, колониальный дом, который должны защищать солдаты.[121]
Солдаты галлообразующей тли также выполняют работу по очистке желчи. Медвяная роса, выделяемая тлей, покрывается порошкообразным воском, образуя "жидкие шарики "[122] что солдаты выкатываются из желчи через небольшие отверстия.[100] Тля, образующая закрытые галлы, использует сосудистую систему растения в качестве водопровода: внутренние поверхности галлов обладают высокой абсорбирующей способностью, а отходы поглощаются и уносятся растением.[100]
Взаимодействие с людьми
Статус вредителя
Описано около 5000 видов тлей, из которых около 450 заселили пищевые и волокнистые культуры. Как прямые питатели сока растений, они повреждают посевы и снижают урожайность, но они оказывают большее влияние, поскольку векторов вирусов растений. Передача этих вирусов зависит от перемещений тли между различными частями растения, между близлежащими растениями и другими районами. В этом отношении зондирующее поведение тли, пробующей хозяина, наносит больший ущерб, чем длительное кормление и размножение тли неподвижными особями. Движение тли влияет на сроки вирусных эпидемий.[123]
Известно, что тля, особенно во время крупных вспышек, вызывает аллергические реакции при вдыхании у чувствительных людей.[124]
Распространение может происходить при ходьбе или бегстве, рассеянии по аппетиту или миграции. Крылатые тли - слабые летчики, теряют крылья через несколько дней и летают только днем. На рассеяние при полете влияют удары, воздушные потоки, сила тяжести, осадки и другие факторы, или рассеяние может быть случайным, вызванным перемещением растительных материалов, животных, сельскохозяйственной техники, транспортных средств или самолетов.[123]
Контроль

Инсектицид борьба с тлями затруднена, так как они быстро размножаются, поэтому даже небольшие пропущенные участки могут позволить популяции быстро восстановиться. Тля может занимать нижнюю часть листьев там, где ее не попадают при распылении. системные инсектициды не переходят в лепестки цветов. Наконец, некоторые виды тлей устойчив к обычным классам инсектицидов в том числе карбаматы, органофосфаты, и пиретроиды.[125]
При заражении небольших дворовых дворов достаточной защитой может быть тщательное опрыскивание растений сильной струей воды каждые несколько дней. An инсектицидное мыло раствор может быть эффективным домашним средством от тли, но он убивает тлю только при контакте и не имеет остаточного действия. Мыльный спрей может повредить растения, особенно при более высоких концентрациях или при температуре выше 32 ° C (90 ° F); некоторые виды растений чувствительны к мыльным спреям.[112][126][127]

Популяции тли можно отбирать с помощью желтой чаши или Moericke ловушки. Это желтые емкости с водой, привлекающие тлю.[128] Тля положительно реагирует на зеленый цвет, и их влечение к желтому может быть не истинным цветовым предпочтением, а связано с яркостью. Их зрительные рецепторы имеют максимальную чувствительность от 440 до 480 нм и нечувствительны в красной области. Мерике обнаружил, что тля избегает приземления на белые покрытия, положенные на почву, и отталкивает их еще больше от блестящих алюминиевых поверхностей.[129] Комплексная борьба с вредителями различных видов тлей можно получить с помощью биологических инсектицидов на основе грибов, таких как Lecanicillium lecanii, Боверия бассиана или Isaria fumosorosea.[130] Грибы - основные возбудители тлей; Энтомофторалес может быстро сократить количество тлей в природе.[131]
Тля также может быть контролируется высвобождением естественных врагов, особенно божьи коровки и паразитоидные осы. Однако, поскольку взрослые жуки-жуки имеют тенденцию улетать в течение 48 часов после выпуска, не откладывая яиц, для обеспечения эффективности необходимо многократное применение большого количества божьих жуков. Например, один большой, сильно зараженный куст роз может потребовать двух обработок по 1500 жуков каждая.[112][132]
Способность производить алломоны, такие как Фарнезен для отпугивания и разгона тлей и привлечения их хищников экспериментально перенесено в трансгенный Arabidopsis thaliana растения, использующие ген Eβf-синтазы в надежде, что этот подход сможет защитить трансгенные культуры.[133] Однако было обнаружено, что Eβ фарнезен неэффективен в условиях выращивания сельскохозяйственных культур, хотя более стабильные синтетические формы помогают повысить эффективность борьбы с использованием спор грибов и инсектицидов за счет увеличения поглощения, вызванного перемещением тли.[134]
В человеческой культуре
Тля знакома фермерам и садоводам, в основном, как вредители. Питер Маррен и Ричард Маби записать это Гилберт Уайт описал вторгшуюся «армию» черной тли, прибывшую в его деревню Селборн, Хэмпшир, Англия в августе 1774 г. в «огромных облаках», покрывающих все растения, а необычно жарким летом 1783 г. Уайт обнаружил, что медвяная роса была настолько обильной, что «испортила и уничтожила красоты моего сада», хотя он думал, что тля пожирает а не производить его.[135]
Зараженность сумаха китайского (Rhus chinensis ) китайской сумаховой тлей (Schlechtendalia chinensis ) может создавать «китайские галлы», которые оцениваются как коммерческий продукт. Как "Galla Chinensis" они используются в традиционная китайская медицина лечить кашляет, понос, ночные поты, дизентерия и остановить кишечное и маточное кровотечение. Китайские галлы также являются важным источником дубильные вещества.[29]
Смотрите также
Заметки
- ^ Термин «черная муха» также используется для обозначения Simuliidae, среди них вектор речная слепота.
- ^ Муравьи-молочные тоже доят мучнистые червецы и другие насекомые.
использованная литература
- ^ Пайпер, Росс (2007). Необычные животные: энциклопедия любопытных и необычных животных. Гринвуд Пресс. стр.6–9. ISBN 978-0-313-33922-6.
- ^ а б Жила, Дагмара; Хоман, Агнешка; Wegierek, Петр (2017). «Полифилия вымершего семейства Oviparosiphidae и ее значение для вывода об эволюции тлей (Hemiptera, Sternorrhyncha)». PLOS ONE. 12 (4): e0174791. Bibcode:2017PLoSO..1274791Z. Дои:10.1371 / journal.pone.0174791. ЧВК 5405925. PMID 28445493.
- ^ Berry, R.E .; Тейлор, Л. Р. (1968). «Высотная миграция тлей в морском и континентальном климате». Журнал экологии животных. 37 (3): 713–722. Дои:10.2307/3084. JSTOR 3084.
- ^ Isard, Scott A .; Ирвин, Майкл Э .; Холлингер, Стивен Э. (1990-10-01). «Вертикальное распределение тлей (Homoptera: Aphididae) в планетарном пограничном слое». Экологическая энтомология. 19 (5): 1473–1484. Дои:10.1093 / ee / 19.5.1473.
- ^ Хилл, Л. (2012). "Смородиновая тля салата, Nasonovia ribisnigri прибывает в Тасманию: Часть 1". Викторианский энтомолог. 42 (2): 29–31.
- ^ Маргаритопулос, Джон Т .; Каспрович, Луиза; Malloch, Gaynor L .; Фентон, Брайан (11 мая 2009 г.). «Отслеживание глобального распространения космополитического насекомого-вредителя, персиковой картофельной тли». BMC Ecology. 9: 13. Дои:10.1186/1472-6785-9-13. ЧВК 2687420. PMID 19432979.
- ^ Szwedo, J .; Нел, А. (2011). «Самое старое насекомое-тля из среднего триаса Вогезов, Франция». Acta Palaeontologica Polonica. 56 (4): 757–766. Дои:10.4202 / app.2010.0034.
- ^ а б Капинера, Джон Л. (2008). Энциклопедия энтомологии. Springer Science & Business Media. С. 193–194. ISBN 978-1-4020-6242-1.
- ^ Джонсон, Кристина; Агости, Донат; Delabie, Jocques H .; Думперт, Клаус; Уильямс, Д. Дж .; фон Чирнхаус, Майкл; Macshwitz, Ульрих (2001). "Акропига и Ацтека муравьи (Hymenoptera: Formicidae) с щитовками (Sternorrhyncha: Coccoidea): 20 миллионов лет интимного симбиоза » (PDF). Американский музей Novitates. 3335: 1–18. Дои:10.1206 / 0003-0082 (2001) 335 <0001: AAAAHF> 2.0.CO; 2.
- ^ Рассел, Луиза М. (1968). «Исследования ископаемых тлей» (PDF). Бюллетень энтомологического общества Америки. 14 (2): 139–140. Дои:10.1093 / besa / 14.2.139a.
- ^ а б c d е ж Диксон, А. Ф. Г. (1998). Экология тли (2-е изд.). Чепмен и Холл. ISBN 978-0-412-74180-7.
- ^ Фон Долен, Кэрол Д .; Моран, Нэнси А. (2000). «Молекулярные данные подтверждают быстрое распространение тлей в меловом периоде и множественные причины смены хозяев». Биологический журнал Линнеевского общества. 71 (4): 689–717. Дои:10.1111 / j.1095-8312.2000.tb01286.x.
- ^ Чен, Руи; Фавре, Колин; Цзян, Лиюнь; Ван, Чжэ; Цяо, Гексия (29 сентября 2015 г.). «Линия тли сохраняет нишу для кормления корой, переключаясь на хвойные деревья и расширяя их ассортимент». Кладистика. 32 (5): 555–572. Дои:10.1111 / cla.12141.
- ^ Гуллан, Пенни Дж .; Мартин, Джон Х. (2009). «Стернорринча». Энциклопедия насекомых (2-е изд.).
- ^ Родендорф, Б. Б., изд. (1991). Основы палеонтологии. Том 9. Arthropoda, Tracheata, Chelicerata.. Смитсоновский институт и Национальный научный фонд. С. 267–274.
- ^ Соренсен, Дж. Т. (2009). «Тля». In Resh, Vincent H .; Карде, Р. Т. (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса. С. 27–31.
- ^ "Надсемейство Aphidoidea, Латрей, 1802 г.". Тля: видовой файл. Получено 3 февраля 2018.
- ^ Blackman, R.L .; Истроп, В. Ф. (1994). Тля на деревьях мира. Руководство по идентификации и информации. Wallingford: CAB International. ISBN 978-0-85198-877-1.
- ^ Гранетт, Джеффри; Уокер, М. Эндрю; Кочиш, Ласло; Омер, Амир Д. (2001). «Биология и менеджмент филлоксеры винограда». Ежегодный обзор энтомологии. 46: 387–412. Дои:10.1146 / annurev.ento.46.1.387. PMID 11112174.
- ^ а б c d е ж Макгэвин, Джордж К. (1993). Ошибки Мира. Издание информационной базы. ISBN 978-0-8160-2737-8.
- ^ Файл вида тлей - Aphidomorpha
- ^ «Филогения насекомых». Что-когда-как. Получено 21 февраля 2018.
- ^ Папасотиропулос, Василис; Циамис, Георгиос; Папайоанну, Хариклея; Килиас, Джордж (2013). «Молекулярное филогенетическое исследование тлей (Hemiptera: Aphididae) на основе анализа последовательности митохондриальной ДНК». Журнал биологических исследований-Салоники. 20: 1–13.
- ^ Ким, Хёджун; Ли, Сынхван; Чан, Иквеон (сентябрь 2011 г.). Моро, Корри С. (ред.). «Макроэволюционные модели тлей Aphidini (Hemiptera: Aphididae): диверсификация, ассоциация хозяев и биогеографическое происхождение». PLOS ONE. 6 (9): e24749. Bibcode:2011PLoSO ... 624749K. Дои:10.1371 / journal.pone.0024749. ЧВК 3174202. PMID 21935453.
- ^ Ортис-Ривас, Бенхамин; Мартинес-Торрес, Дэвид (2010). «Комбинация молекулярных данных подтверждает существование трех основных ветвей филогении тлей (Hemiptera: Aphididae) и базальное положение подсемейства Lachninae». Молекулярная филогенетика и эволюция. 55 (1): 305–317. Дои:10.1016 / j.ympev.2009.12.005. PMID 20004730.
- ^ Кларк, Марта А; Moran, Nancy A .; Бауманн, Пол; Вернегрин, Дженнифер Дж. (2000). «Совместное родство между бактериальными эндосимбионтами (Buchnera) и недавней радиацией тлей (Uroleucon) и подводные камни тестирования для филогенетической конгруэнтности». Эволюция. 54 (2): 517–25. Дои:10.1554 / 0014-3820 (2000) 054 [0517: CBBEBA] 2.0.CO; 2. PMID 10937228.
- ^ Новакова, Ева; Гипша, Вацлав; Кляйн, Джоанн; Футтит, Роберт Дж. фон Долен, Кэрол Д .; Моран, Нэнси А. (2013). «Реконструкция филогении тлей (Hemiptera: Aphididae) с использованием ДНК облигатного симбионта Buchnera aphidicola» (PDF). Молекулярная филогенетика и эволюция. 68 (1): 42–54. Дои:10.1016 / j.ympev.2013.03.016. PMID 23542003.
- ^ Чен, Руи; Ван, Чжэ; Чен, Цзин; Цзян, Ли-Юнь; Цяо, Гэ-Ся (2017). «Параллельная эволюция насекомых-бактерий в ассоциации множественных облигатов-тлей: случай у Lachninae (Hemiptera: Aphididae)». Научные отчеты. 7 (1): 10204. Bibcode:2017НатСР ... 710204C. Дои:10.1038 / s41598-017-10761-9. ЧВК 5579299. PMID 28860659.
- ^ а б c d е ж г час Строян, Генри Г. (1997). «Тля». Энциклопедия науки и технологий Макгроу-Хилла (8-е изд.). ISBN 978-0-07-911504-1.
- ^ Мутти, Навдип С. (2006). Молекулярные исследования слюнных желез гороховой тли. Acyrthosiphon pisum (Харрис) (PDF) (Кандидат наук. Тезис). Канзасский государственный университет.
- ^ а б c ван Эмден, Хельмут Ф .; Харрингтон, Ричард (2017). Тля как вредитель сельскохозяйственных культур. КАБИ. С. 189–190. ISBN 978-1-78064-709-8.
- ^ Пауэлл, Глен; Тош, Колин Р .; Харди, Джим (2005). «Выбор растений-хозяев тлей: поведенческие, эволюционные и прикладные перспективы». Ежегодный обзор энтомологии. 51 (1): 309–330. Дои:10.1146 / annurev.ento.51.110104.151107. PMID 16332214.
- ^ Spiller, N.J .; Koenders, L .; Tjallingii, W. F. (1990). «Заглатывание ксилемы тлей - стратегия поддержания водного баланса». Entomologia Experimentalis et Applicata. 55 (2): 101–104. Дои:10.1007 / BF00352570.
- ^ а б Фишер, Д. Б. (2000). «Дальний транспорт». В Бьюкенене, Боб Б.; Груиссем, Вильгельм; Джонс, Рассел Л. (ред.). Биохимия и молекулярная биология растений (4-е изд.). Роквилл, Мэриленд: Американское общество физиологов растений. стр.730–784. ISBN 978-0-943088-39-6.
- ^ Мэлоун, М .; Watson, R .; Причард, Дж. (1999). "Клоп Philaenus spumarius питается из зрелой ксилемы при полном гидравлическом напряжении транспирационного потока ». Новый Фитолог. 143 (2): 261–271. Дои:10.1046 / j.1469-8137.1999.00448.x. JSTOR 2588576.
- ^ Пауэлл, Глен; Харди, Джим (2002). «Заглатывание ксилемы крылатой тлей». Entomologia Experimentalis et Applicata. 104 (1): 103–108. Дои:10.1023 / А: 1021234412475.
- ^ а б c d Помпон, Жюльен; Квиринг, Дэн; Джорданенго, Филипп; Пеллетье, Иван (2010). "Роль потребления ксилемы в осморегуляции в Macrosiphum euphorbiae (Томас)" (PDF). Журнал физиологии насекомых. 56 (6): 610–615. Дои:10.1016 / j.jinsphys.2009.12.009. PMID 20036244. Архивировано из оригинал (PDF) 16 июля 2011 г.
- ^ Kingsolver, J. G .; Даниэль, Т. Л. (1995). «Механика обработки пищевых продуктов насекомыми, питающимися жидкостью». В Chapman, R. F .; де Бур, Геррит (ред.). Регулирующие механизмы кормления насекомых. Springer. С. 60–65.
- ^ Эшфорд, Д. А .; Smith, W. A .; Дуглас, А. Э. (2000). "Жизнь на диете с высоким содержанием сахара: судьба сахарозы, попавшей в организм насекомого, питающегося флоэмами, гороховой тли. Acyrthosiphon pisum". Журнал физиологии насекомых. 46 (3): 335–341. Дои:10.1016 / S0022-1910 (99) 00186-9. PMID 12770238.
- ^ Wilkinson, T. L .; Ashfors, D. A .; Pritchard, J .; Дуглас, А. Э. (1997). "Медовый сахар и осморегуляция у гороховой тли Acyrthosiphon pisum". Журнал экспериментальной биологии. 200 (11): 2137–2143. PMID 9320049.
- ^ Shakesby, A.J .; Wallace, I.S .; Isaacs, H.V .; Pritchard, J .; Робертс, Д. М .; Дуглас, А. Э. (2009). «Специфический для воды аквапорин, участвующий в осморегуляции тли». Биохимия и молекулярная биология насекомых. 39 (1): 1–10. Дои:10.1016 / j.ibmb.2008.08.008. PMID 18983920.
- ^ Dadd, R.H .; Миттлер, Т. (1965). «Исследования по искусственному вскармливанию тли. Myzus persicae (Зульцер) - III. Некоторые основные потребности в питании ". Журнал физиологии насекомых. 11 (6): 717–743. Дои:10.1016 / 0022-1910 (65) 90154-Х. PMID 5827534.
- ^ Бюхнер, Пол (1965). Эндосимбиоз животных с помощью растительных микроорганизмов. Interscience. ISBN 978-0-470-11517-6.
- ^ Уайтхед, Л. Ф .; Дуглас, А. Э. (1993). "Метаболическое исследование Бухнера, внутриклеточные бактериальные симбионты гороховой тли Acyrthosiphon pisum" (PDF). Журнал общей микробиологии. 139 (4): 821–826. Дои:10.1099/00221287-139-4-821.
- ^ Феввей, Жерар; Лиадуз, Изабель; Гийо, Жозетт; Боннот, Гай (1995). «Анализ метаболизма энергетических аминокислот в Acyrthosiphon pisum: многомерный подход к метаболизму аминокислот у тлей ». Архивы биохимии и физиологии насекомых. 29 (1): 45–69. Дои:10.1002 / arch.940290106.
- ^ Moran, Nancy A .; Ярвик, Тайлер (2010). «Боковой перенос генов от грибов лежит в основе продукции каротиноидов у тлей». Наука. 328 (5978): 624–627. Bibcode:2010Sci ... 328..624M. Дои:10.1126 / science.1187113. PMID 20431015. S2CID 14785276.
- ^ Altincicek, B .; Kovacs, J.L .; Херардо, Н. М. (2012). "Горизонтально перенесенные каротиноидные гены грибов у двупятнистого паутинного клеща Tetranychus urticae". Письма о биологии. 8 (2): 253–257. Дои:10.1098 / рсбл.2011.0704. ЧВК 3297373. PMID 21920958.
- ^ Вальмалетт, Жан Кристоф; Домбровский, Авив; Брат, Пьер; Мерц, Кристиан; Каповилла, Мария; Робишон, Ален (2012). «Индуцированный светом перенос электронов и синтез АТФ у насекомых, синтезирующих каротин». Научные отчеты. 2: 579. Bibcode:2012НатСР ... 2Е.579В. Дои:10.1038 / srep00579. ЧВК 3420219. PMID 22900140.
- ^ а б Wang, C.L .; Siang, L. Y .; Чанг, Г. С .; Чу, Х. Ф. (1962). «Исследования соевой тли, Афис глицины Мацумура ". Acta Entomologica Sinica. 11: 31–44.
- ^ а б ван Эмден, Хельмут Ф .; Харрингтон, Ричард (2017). Тля как вредитель сельскохозяйственных культур, 2-е издание. КАБИ. С. 81–82. ISBN 978-1-78064-709-8.
- ^ Фон Долен, Кэрол; Моран, Нэнси А. (2000). «Молекулярные данные подтверждают быстрое облучение тлей в меловом периоде и множественное происхождение смены хозяев». Биологический журнал Линнеевского общества. 71 (4): 689–717. Дои:10.1006 / bijl.2000.0470.
- ^ Моран, Нэнси А. (1992). «Эволюция жизненных циклов тли». Ежегодный обзор энтомологии. 37: 321–348. Дои:10.1146 / annurev.en.37.010192.001541.
- ^ а б Блэкман, Роджер Л. (2008). «Стабильность и изменчивость клональных линий тлей». Биологический журнал Линнеевского общества. 11 (3): 259–277. Дои:10.1111 / j.1095-8312.1979.tb00038.x.
- ^ а б Gullan, P.J .; Крэнстон, П. С. (2010). Насекомые: очерк энтомологии (4-е изд.). Вайли. С. 150–151. ISBN 978-1-118-84615-5.
- ^ а б c Хейлз, Дина Ф .; Уилсон, Алекс С. С .; Слоан, Мэтью А .; Саймон, Жан-Кристоф; Легаллик, Жан-Франсуа; Sunnucks, Пол (2002). «Отсутствие детектируемой генетической рекомбинации на Х-хромосоме во время партеногенетической продукции самок и самцов тлей». Генетические исследования. 79 (3): 203–209. Дои:10.1017 / S0016672302005657. PMID 12220127.
- ^ Нево, Эттай; Колл, Моше (2001). "Влияние азотных удобрений на Тля gossypii (Homoptera: Aphididae): различия в размере, цвете и воспроизводстве ». Журнал экономической энтомологии. 94 (1): 27–32. Дои:10.1603/0022-0493-94.1.27. PMID 11233124. S2CID 25758038.
- ^ Jahn, Gary C .; Алмазан, Либерти П .; Pacia, Джоселин Б. (2005). "Влияние азотных удобрений на внутреннюю скорость роста ржавой сливовой тли, Hysteroneura setariae (Thomas) (Homoptera: Aphididae) на рисе (Oryza sativa L.) " (PDF). Экологическая энтомология. 34 (4): 938–943. Дои:10.1603 / 0046-225X-34.4.938. S2CID 1941852. Архивировано из оригинал (PDF) на 09.09.2010.
- ^ Хьюз, Р. Д. (1963). «Динамика численности капустной тли. Brevicoryne brassicae (Л.) ". Журнал экологии животных. 32 (3): 393–424. Дои:10.2307/2600. JSTOR 2600.
- ^ Бриссон, Дженнифер А. (2010). "Диморфизм крыльев тли: связь экологического и генетического контроля изменчивости признаков". Философские труды Королевского общества B: биологические науки. 365 (1540): 605–616. Дои:10.1098 / rstb.2009.0255. ЧВК 2817143. PMID 20083636.
- ^ Weisser, Wolfgang W .; Zytynska, Sharon E .; Мехрпарвар, Мохсен (05.03.2013). «Множественные подсказки для производства крылатых морфов в метасообществе тлей». PLOS ONE. 8 (3): e58323. Bibcode:2013PLoSO ... 858323M. Дои:10.1371 / journal.pone.0058323. ISSN 1932-6203. ЧВК 3589340. PMID 23472179.
- ^ Лис, А. Д. (1967-02-01). «Производство бескрылых и крыловидных форм тли Megoura viciae Buckton, с особым упором на роль скученности». Журнал физиологии насекомых. 13 (2): 289–318. Дои:10.1016/0022-1910(67)90155-2. ISSN 0022-1910.
- ^ Кунерт, Грит; Отто, Сюзанна; Röse, Ursula S.R .; Гершензон, Джонатан; Вайссер, Вольфганг В. (28 апреля 2005 г.). «Тревожный феромон опосредует образование крылатых морф распространения у тлей». Письма об экологии. 8 (6): 596–603. Дои:10.1111 / j.1461-0248.2005.00754.x. ISSN 1461-023X.
- ^ Рябов, Э. В .; Keane, G .; Naish, N .; Evered, C .; Уинстэнли, Д. (13 мая 2009 г.). «Дензовирус индуцирует крылатые морфы у бесполых клонов розовой яблочной тли, Dysaphis plantaginea». Труды Национальной академии наук. 106 (21): 8465–8470. Bibcode:2009PNAS..106.8465R. Дои:10.1073 / pnas.0901389106. ISSN 0027-8424. ЧВК 2688996. PMID 19439653.
- ^ Чан, К. К. (1991). Вирусы, передаваемые тлей, и их переносчики в мире. Отдел исследований, Сельское хозяйство Канады. ISBN 0662183347. OCLC 872604083.
- ^ Reyes, Miguel L .; Laughton, Alice M .; Паркер, Бенджамин Дж .; Вичманн, Ханна; Вентилятор, Маретта; Сок, Даниэль; Грчек, Ян; Асеведо, Тарик; Херардо, Николь М. (31.01.2019). «Влияние симбиотических бактерий на репродуктивные стратегии и полифенизм крыльев гороховой тли в ответ на стресс». Журнал экологии животных. 88 (4): 601–611. Дои:10.1111/1365-2656.12942. ISSN 0021-8790. ЧВК 6453707. PMID 30629747.
- ^ Алфорд, Дэвид В. (2014). Вредители плодовых культур: Справочник по цветам, второе издание. CRC Press. С. 71–72. ISBN 978-1-4822-5421-1.
- ^ а б Хупер-Буй, Линда М. (2008). "Муравей". Всемирная книжная энциклопедия. ISBN 978-0-7166-0108-1.
- ^ Стадлер, Бернхард; Диксон, Энтони Ф. Г. (2005). «Экология и эволюция взаимодействий тлей и муравьев». Ежегодный обзор экологии, эволюции и систематики. 36 (1): 345–372. Дои:10.1146 / annurev.ecolsys.36.091704.175531.
- ^ Вуттон, Энтони (1998). Насекомые мира. Blandford. ISBN 978-0-7137-2366-3.
- ^ а б Нири, Джон (1977). Насекомые и пауки. Книги времени жизни. стр.78–79. ISBN 978-0-8094-9687-7.
- ^ Салазар, Адриан; Фюрстенау, Бенджамин; Quero, Кармен; Перес-Идальго, Николас; Карасо, По; Шрифт, Энрике; Мартинес-Торрес, Дэвид (2015). «Агрессивная мимикрия у тли сосуществует с мутуализмом». Труды Национальной академии наук. 112 (4): 1101–1106. Bibcode:2015ПНАС..112.1101С. Дои:10.1073 / pnas.1414061112. ЧВК 4313836. PMID 25583474.
- ^ Бауманн, Пол; Moran, Nancy A .; Бауман, Линда (2006). «Бактериоцит-ассоциированные эндосимбионты насекомых». В Дворкине, Мартин; Фалькоу, Стэнли; Розенберг, Юджин; Шлейфер, Карл-Хайнц; Stackebrandt, Erko (ред.). Прокариоты. С. 403–438. Дои:10.1007/0-387-30741-9_16. ISBN 978-0-387-25476-0.
- ^ Дуглас, А. Э. (1998). "Питательные взаимодействия при симбиозах насекомых-микробов: тли и их симбиотические бактерии Buchnera". Ежегодный обзор энтомологии. 43 (1): 17–37. Дои:10.1146 / annurev.ento.43.1.17. PMID 15012383.
- ^ Бодел, Анджей; Мацкевич, Павел; Гагат, Пшемыслав (2012). "Эволюция органелл: Паулинелла ломает парадигму". Текущая биология. 22 (9): R304 – R306. Дои:10.1016 / j.cub.2012.03.020. PMID 22575468.
- ^ Бауманн, Линда; Бауманн, Пол; Moran, Nancy A .; Сандстрём, Йонас; Тао, Мило Ли (январь 1999 г.). «Генетическая характеристика плазмид, содержащих гены, кодирующие ферменты биосинтеза лейцина у эндосимбионтов (Buchnera) тлей». Журнал молекулярной эволюции. 48 (1): 77–85. Дои:10.1007 / PL00006447. ISSN 0022-2844.
- ^ Сабри, Ахмед; Вандермотен, Софи; Leroy, Pascal D .; Haubruge, Эрик; Ганс, Тьерри; Тонарт, Филипп; Де Пау, Эдвин; Фрэнсис, Фредерик (25 сентября 2013 г.). «Протеомное исследование медовой росы тли выявило неожиданное разнообразие белков». PLOS ONE. 8 (9): e74656. Bibcode:2013PLoSO ... 874656S. Дои:10.1371 / journal.pone.0074656. ISSN 1932-6203. ЧВК 3783439. PMID 24086359.
- ^ Perez-Brocal, V .; Gil, R .; Ramos, S .; Lamelas, A .; Postigo, M .; Michelena, J.M .; Silva, F.J .; Моя, А .; Латорре, А. (2006). «Маленький микробный геном: конец долгого симбиотического отношения?». Наука. 314 (5797): 312–313. Bibcode:2006Научный ... 314..312П. Дои:10.1126 / наука.1130441. PMID 17038625. S2CID 40081627.
- ^ Мира, А .; Моран, Нэнси А. (2002). «Оценка размера популяции и узких мест передачи эндосимбиотических бактерий, передаваемых от матери». Микробная экология. 44 (2): 137–143. Дои:10.1007 / s00248-002-0012-9. PMID 12087426. S2CID 33681686.
- ^ Сакаки, Ёсиюки; Сигенобу, Сюдзи; Ватанабэ, Хидеми; Хаттори, Масахира; Исикава, Хадзиме (2000). «Последовательность генома внутриклеточного бактериального симбионта тли. Бухнера sp. APS ». Природа. 407 (6800): 81–86. Bibcode:2000Натура 407 ... 81С. Дои:10.1038/35024074. PMID 10993077.
- ^ Виньуэлас, Хосе; Калевро, Федерика; Ремонд, Дидье; Бернильон, Жак; Рабе, Иван; Феввей, Жерар; Файяр, Жан-Мишель; Чарльз, Хуберт (2007). «Сохранение связей между транскрипцией генов и хромосомной организацией в сильно восстановленном геноме Buchnera aphidicola». BMC Genomics. 8 (1): 143. Дои:10.1186/1471-2164-8-143. ЧВК 1899503. PMID 17547756.
- ^ Moran, Nancy A .; Данбар, Хелен Э .; Уилкокс, Дженнифер Л. (2005). «Регуляция транскрипции в редуцированном бактериальном геноме: гены, обеспечивающие питательные вещества обязательного симбионта» Buchnera aphidicola". Журнал бактериологии. 187 (12): 4229–4237. Дои:10.1128 / JB.187.12.4229-4237.2005. ЧВК 1151715. PMID 15937185.
- ^ Tsuchida, T .; Koga, R .; Meng, X. Y .; Т. Мацумото; Т. Фукацу (2005). «Характеристика факультативной эндосимбиотической бактерии гороховой тли. Acyrthosiphon pisum". Микробная экология. 49 (1): 126–133. Дои:10.1007 / s00248-004-0216-2. PMID 15690225. S2CID 24144752.
- ^ Сакураи, М .; Koga, R .; Tsuchida, T .; Meng, X.-Y .; Фукацу, Т. (2005). «Симбионт риккетсии в гороховой тле Acyrthosiphon pisum: новый клеточный тропизм, влияние на приспособленность хозяина и взаимодействие с основным симбионтом Бухнера». Прикладная и экологическая микробиология. 71 (7): 4069–4075. Дои:10.1128 / AEM.71.7.4069-4075.2005. ЧВК 1168972. PMID 16000822.
- ^ Феррари, Юлия; Скарборо, Клэр Л .; Годфрей, Х. Чарльз Дж. (2007). «Генетические вариации в эффекте факультативного симбионта на использование растения-хозяина гороховой тлей». Oecologia. 153 (2): 323–329. Bibcode:2007Oecol.153..323F. Дои:10.1007 / s00442-007-0730-2. PMID 17415589. S2CID 37052892.
- ^ Simon, J.-C .; Carre, S .; Boutin, M .; Prunier-Leterme, N .; Сабатер-Муньос, Б .; Latorre, A .; Бурновиль Р. (2003). «Дивергенция на основе хозяев в популяциях гороховой тли: выводы из ядерных маркеров и преобладание факультативных симбионтов». Труды Королевского общества B: биологические науки. 270 (1525): 1703–1712. Дои:10.1098 / rspb.2003.2430. ЧВК 1691435. PMID 12964998.
- ^ Велдон, Стефани Р .; Оливер, Керри М. (2016). Механические преимущества микробных симбионтов. Успехи экологической микробиологии. Спрингер, Чам. С. 173–206. Дои:10.1007/978-3-319-28068-4_7. ISBN 9783319280660.
- ^ Weldon, S. R .; Strand, M. R .; Оливер, К. М. (22 января 2013 г.). «Потеря фагов и нарушение защитного симбиоза у тлей». Труды Лондонского королевского общества B: биологические науки. 280 (1751): 20122103. Дои:10.1098 / rspb.2012.2103. ЧВК 3574403. PMID 23193123.
- ^ Оливер, К. М .; Moran, Nancy A .; Хантер, М. С. (2006). «Стоимость и преимущества суперинфекции факультативных симбионтов у тлей». Труды Королевского общества B: биологические науки. 273 (1591): 1273–1280. Дои:10.1098 / rspb.2005.3436. ЧВК 1560284. PMID 16720402.
- ^ а б Капинера, Джон (2011). Насекомые и дикая природа: членистоногие и их отношения с дикими позвоночными животными. Джон Вили и сыновья. п. 536. ISBN 978-1-4443-5784-4.
- ^ ван Эмден, Хельмут Ф .; Харрингтон, Ричард (2017). Тля как вредитель сельскохозяйственных культур. КАБИ. С. 229–230. ISBN 978-1-78064-709-8.
- ^ а б c Brust, Джеральд Э. (22 июня 2006 г.). «Раннеспелые популяции тлей и трипсов». Университет Мэриленда, Колледж-Парк. Архивировано из оригинал 19 июля 2011 г.. Получено 18 октября 2010.
- ^ Лэмб, К. П. (1961). "Некоторые эффекты колебаний температуры на метаболизм, развитие и скорость роста популяции капустной тли, Brevicoryne brassicae". Экология. 42 (4): 740–745. Дои:10.2307/1933502. JSTOR 1933502.
- ^ Джонс, Маргарет Г. (1979). «Численность тли на злаках с 1973 по 1977 годы». Журнал прикладной экологии. 16 (1): 1–22. Дои:10.2307/2402724. JSTOR 2402724.
- ^ Крупке, Кристиан; Обермейер, Джон; О'Нил, Роберт (2007). «Соевая тля - новое начало 2007 года». Вредители и урожай. Университет Пердью. 7.
- ^ «Почему некоторые тли не переносят жару». Science Daily. 23 апреля 2007 г.
- ^ Хьюз, Р. Д. (1963). «Динамика численности капустной тли, Brevicoryne brassicae (Л.) ". Журнал экологии животных. 32 (3): 393–424. Дои:10.2307/2600. JSTOR 2600.
- ^ Суванбутр, С. (1996). «Стабильное возрастное распределение популяций люцерновой тли в Юго-Восточной Тасмании» (PDF). Международный научно-технический журнал Thammasat. 1 (5): 38–43. Архивировано из оригинал (PDF) 10 сентября 2008 г.
- ^ Остли, Кен (3 августа 2006 г.). «Паутинный клещ, тля и дождь затрудняют принятие решения об опрыскивании сои» (PDF). Minnesota Crop eNews. Университет Миннесоты. Архивировано из оригинал (PDF) 10 сентября 2008 г.
- ^ Аоки, С. (1977). "Колофина клематис (Homoptera, Pemphigidae), вид тлей с солдатиками » (PDF). Японский журнал энтомологии. 45 (2): 276–282.[постоянная мертвая ссылка ]
- ^ а б c Kutsukake, M .; Meng, X.Y .; Katayama, N .; Nikoh, N .; Shibao, H .; Фукацу, Т. (2012). «Индуцированный насекомыми новый фенотип растений для поддержания социальной жизни в закрытой системе». Nature Communications. 3: 1187–1193. Bibcode:2012 НатКо ... 3,1 187 тыс.. Дои:10.1038 / ncomms2187. ЧВК 3514493. PMID 23149732.
- ^ Престон-Мафхэм, Род; Престон-Мафхэм, Кен (1993). Энциклопедия поведения наземных беспозвоночных. MIT Press. п.281. ISBN 978-0-262-16137-4.
- ^ Гиш, М .; Дафни, А .; Инбар, М. (2012). Хайль, Мартин (ред.). «Молодые тли избегают ошибочного опускания при уклонении от травоядных млекопитающих, комбинируя данные от двух сенсорных модальностей». PLOS ONE. 7 (4): e32706. Bibcode:2012PLoSO ... 732706G. Дои:10.1371 / journal.pone.0032706. ЧВК 3322135. PMID 22496734.
- ^ Казана, Элеанна; Папа, Том У .; Тибблс, Лориенн; Бриджес, Мэтью; Пикетт, Джон А .; Bones, Atle M .; Пауэлл, Глен; Росситер, Джон Т. (2007). «Капустная тля: ходячая бомба с горчичным маслом». Труды Королевского общества B. 274 (1623): 2271–7. Дои:10.1098 / rspb.2007.0237. ЧВК 2288485. PMID 17623639.
- ^ Вильчинскас, Андреас (2016). «Иммунитет к тли». Биология и экология тли. CRC Press. п. 131.
- ^ Эдвардс, Джон С. (1966). «Защита мазком: переохлаждение рогового воска тли». Природа. 211 (5044): 73–74. Bibcode:1966Натура.211 ... 73Э. Дои:10.1038 / 211073a0. S2CID 4295930.
- ^ Мартинсон, Кэндис (2008). «Тля». Всемирная книжная энциклопедия. ISBN 978-0-7166-0108-1.
- ^ Уэй, M.J. (1963). «Взаимность муравьев и росистых равнокрылых». Ежегодный обзор энтомологии. 8: 307–344. Дои:10.1146 / annurev.en.08.010163.001515.
- ^ Грассвиц, Тесса Р .; Пейн, Тимоти Д. (1992). «Кайромональное действие секреции роговицы тли на Lysiphlebus testaceipes (Крессон) (Hymenoptera: Aphidiidae) ". Журнал поведения насекомых. 5 (4): 447–457. Дои:10.1007 / BF01058190. S2CID 25298742.
- ^ Lenhart, Paul A .; Джексон, Келли А .; Белый, Дженнифер А. (2018). «Унаследованные вариации защиты жертвы обеспечивают убежище для хищников-субдоминантов». Труды Королевского общества B: биологические науки. 285 (1879): 20180523. Дои:10.1098 / rspb.2018.0523. ЧВК 5998095. PMID 29848647.
- ^ «Тля-паразитоиды». Университет Мэриленда. Получено 29 апреля 2018.
- ^ Гавелка Ян; Томанович, Желько; Kavallieratos, Nickolas G .; Рахшани, Эхсан; Понс, Ксавье; Петрович, Анджелько; Пайк, Кейт С .; Старый, Петр (01.05.2012). «Обзор и ключ к мировым паразитоидам (Hymenoptera: Braconidae: Aphidiinae) Aphis ruborum (Hemiptera: Aphididae) и его роль в качестве резервуара-хозяина». Анналы Энтомологического общества Америки. 105 (3): 386–394. Дои:10.1603 / an11108. S2CID 84348019.
- ^ а б c Флинт, М. (Июль 2013). «Тля». УК ИПМ. Архивировано из оригинал на 2018-04-09. Получено 6 февраля 2018.
- ^ Гибсон, Р. У .; Пикетт, Дж. А. (1983). «Дикий картофель отпугивает тлю, высвобождая феромон тревоги для тли». Природа. 302 (5909): 608–609. Bibcode:1983Натура.302..608Г. Дои:10.1038 / 302608a0. S2CID 4345998.
- ^ Гибсон, Р. У. (1971). «Железистые волосы, обеспечивающие устойчивость к тле у некоторых видов дикого картофеля». Анналы прикладной биологии. 68 (2): 113–119. Дои:10.1111 / j.1744-7348.1971.tb06448.x.
- ^ Министерство сельского хозяйства США (2003 г.). Исследования химических и биологических свойств куместрола и родственных соединений. Типография правительства США. С. 47–67.
- ^ Гиллман, Дэниел Х. (2005). «Закопченная плесень» (PDF). Массачусетский университет в Амхерсте. Получено 18 октября, 2010.
- ^ Reynolds, Hannah T .; Волк, Том (сентябрь 2007 г.). "Scorias spongiosa, буковая тля-корма ". Грибок месяца Тома Волка. Университет Висконсина – Ла-Кросс. Архивировано из оригинал 30 июля 2010 г.. Получено 18 октября, 2010.
- ^ Дик, А. Дж .; ван Пелт, Дж. А. (1992). «Взаимодействие филлосферных дрожжей, пади тли и эффективность фунгицидов в пшенице в полевых условиях». Патология растений. 41 (6): 661–675. Дои:10.1111 / j.1365-3059.1992.tb02550.x.
- ^ Чоудхури, Дхрупад (1985). «Медвяная тля: переоценка гипотезы Оуэна и Вигерта». Ойкос. 45 (2): 287–290. Дои:10.2307/3565718. JSTOR 3565718.
- ^ Stern, D. L .; Фостер, В. А. (1996). «Эволюция солдатиков у тлей». Биологические обзоры Кембриджского философского общества. 71 (1): 27–79. Дои:10.1111 / j.1469-185X.1996.tb00741.x. PMID 8603120. S2CID 8991755.
- ^ Чо; Jae C .; Креспи, Бернард Дж. (1997). Эволюция социального поведения насекомых и паукообразных. Издательство Кембриджского университета. С. 150–152. ISBN 978-0-521-58977-2.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Pike, N .; Ричард, Д .; Foster, W .; Махадеван, Л. (2002). «Как тля теряет мраморность». Труды Королевского общества B: биологические науки. 269 (1497): 1211–5. Дои:10.1098 / rspb.2002.1999. ЧВК 1691028. PMID 12065036.
- ^ а б ван Эмден, Хельмут Ф .; Харрингтон, Ричард (2017). Тля как вредитель сельскохозяйственных культур. КАБИ. С. 196–198. ISBN 978-1-78064-709-8.
- ^ Шульман, Сидней (1 января 1967 г.). «Аллергические реакции на насекомых». Ежегодный обзор энтомологии. 12 (1): 323–346. Дои:10.1146 / annurev.en.12.010167.001543.
- ^ Пундт, Линн (2011). «Борьба с тлей в теплице». Колледж сельского хозяйства, здравоохранения и природных ресурсов Университета Коннектикута. Архивировано из оригинал на 2018-02-18. Получено 20 февраля 2018.
- ^ Cranshaw, W.S. (Март 2008 г.). «Борьба с насекомыми: мыло и моющие средства». Колорадский университет. Получено 6 февраля 2018.
- ^ Убл, Дж. Б. (июль 2009 г.). «Инсектицидное мыло для борьбы с вредителями в саду». Университет Клемсона. Получено 6 февраля 2018.
- ^ Эванс, Д. А .; Медлер, Дж. Т. (1966-12-01). «Улучшенный метод использования ловушек для желтой тли1». Журнал экономической энтомологии. 59 (6): 1526–1527. Дои:10.1093 / jee / 59.6.1526. ISSN 1938-291X.
- ^ Деринг, Томас Феликс; Читтка, Ларс (2007). «Визуальная экология тли - критический обзор роли цвета в поиске хозяев» (PDF). Взаимодействие членистоногих и растений. 1: 3–16. Дои:10.1007 / s11829-006-9000-1. S2CID 20066025.
- ^ Jaronski, S.T .; Маскарин, Г. М. (2017). Лейси, Лоуренс (ред.). Массовое производство грибковых энтомопатогенов. Микробный контроль насекомых и клещей-вредителей: от теории к практике. Академическая пресса. С. 141–155. ISBN 978-0-12-803527-6.
- ^ Стейнкраус, Дональд К. (2006). «Факторы, влияющие на передачу грибковых возбудителей тлей». Журнал патологии беспозвоночных. 92 (3): 125–131. Дои:10.1016 / j.jip.2006.03.009. PMID 16780867.
- ^ Леди Жук выпускает для борьбы с тлей: как помочь им в работе. Кларк, Дж. К., Калифорнийский университет в Дэвисе, июнь 2011 г.
- ^ Бил, М. Х .; Birkett, M.A .; Bruce, T. J. A .; Чемберлен, К .; Филд, Л. М .; Huttly, A.K .; Martin, J. L .; Parker, R .; Phillips, A. L .; Пикетт, Дж. А .; Проссер, И. М .; Shewry, P. R .; Смарт, Л. Э .; Wadhams, L.J .; Woodcock, C.M .; Чжан, Ю. (2006). «Феромон тревоги для тли, продуцируемый трансгенными растениями, влияет на поведение тлей и паразитоидов». Труды Национальной академии наук. 103 (27): 10509–10513. Bibcode:2006ПНАС..10310509Б. Дои:10.1073 / pnas.0603998103. ЧВК 1502488. PMID 16798877.
- ^ Пикетт, Дж. А .; Wadhams, L.J .; Woodcock, C.M .; Харди, Дж. (1992). «Химическая экология тли». Ежегодный обзор энтомологии. 37 (1): 67–90. Дои:10.1146 / annurev.en.37.010192.000435.
- ^ Маррен, Питер; Маби, Ричард (2010). Ошибки Britannica. Chatto & Windus. С. 191–194. ISBN 978-0-7011-8180-2.
внешние ссылки
- Тля древесных декоративных растений юго-востока США
- Acyrthosiphon pisum, MetaPathogen - факты, жизненный цикл, изображение жизненного цикла
- Секвенированный геном гороховой тли, Служба сельскохозяйственных исследований
- Обоняние запаха растений насекомыми: исследования колорадского жука и тли
- Азиатская шерстистая каркасная тля, Центр исследования инвазивных видов
На Университет Флориды / Институт продовольственных и сельскохозяйственных наук Избранные существа интернет сайт:
- Тля gossypii, дынная или хлопковая тля
- Афис Нери, олеандровая тля
- Hyadaphis coriandri, кориандровая тля
- Longistigma caryae, гигантская коровая тля
- Myzus persicae, зеленая персиковая тля
- Сарукаллис кахавалуокалани, крепмиртовая тля
- Shivaphis celti, азиатская шерстистая каркасная тля
- Toxoptera citricida, коричневая цитрусовая тля
| Авторитетный контроль |
|---|