Нейтрофил - Neutrophil
| Нейтрофил | |
|---|---|
 3D-рендеринг нейтрофила | |
 Нейтрофилы с сегментированными ядрами, окруженными эритроциты и тромбоциты. Внутриклеточные гранулы видны в цитоплазма (Гимза окрашенный ). | |
| Подробности | |
| Система | Иммунная система |
| Функция | Фагоцитоз |
| Идентификаторы | |
| MeSH | D009504 |
| TH | H2.00.04.1.02012 |
| FMA | 62860 |
| Анатомические термины микроанатомии | |
Нейтрофилов (также известен как нейтроциты или же гетерофилы) являются наиболее распространенным типом гранулоциты и составляют от 40% до 70% всех белые кровяные клетки в людях.[1] Они составляют важную часть врожденная иммунная система, причем их функции различны у разных животных.[2]
Они сформированы из стволовые клетки в Костный мозг и дифференцирован на субпопуляции нейтрофилов-киллеров и нейтрофилов-клеток. Они недолговечны и очень подвижны, или подвижны, поскольку могут проникать в части ткани, куда не могут проникнуть другие клетки / молекулы. Нейтрофилы можно подразделить на сегментированные нейтрофилы и полосатые нейтрофилы (или группы ). Они составляют часть семейство полиморфно-ядерных клеток (PMN) вместе с базофилы и эозинофилы.[3][4][5]
Название нейтрофил происходит из характеристик окрашивания на гематоксилин и эозин (ОН ) гистологический или же цитологический препараты. В то время как базофильный белые кровяные тельца окрашиваются в темно-синий цвет и эозинофильный белые кровяные тельца окрашиваются в ярко-красный цвет, нейтрофилы окрашиваются в нейтральный розовый цвет. Обычно нейтрофилы содержат ядро, разделенное на 2–5 долей.
Нейтрофилы - это тип фагоцит и обычно находятся в кровоток. В начале (острый ) фаза воспаление, особенно в результате бактериальный инфекционное заболевание, воздействие окружающей среды,[6] и некоторые виды рака,[7][8] нейтрофилы являются одними из первых реагирующих на миграцию воспалительных клеток к месту воспаления. Они мигрируют по кровеносным сосудам, а затем через межстраничный ткань, следуя химическим сигналам, таким как Интерлейкин-8 (Ил-8), C5a, fMLP, Лейкотриен B4, и ЧАС2О2[9] в процессе, называемом хемотаксис. Они являются преобладающими клетками в гной, что объясняет его беловатый / желтоватый оттенок.[10]
Нейтрофилы попадают в место повреждения в течение нескольких минут после травмы и являются признаком острого воспаления;[11] однако из-за некоторых патогены будучи неперевариваемыми, они могут не справиться с некоторыми инфекциями без помощи других типов иммунных клеток.
Структура


При прикреплении к поверхности нейтрофильные гранулоциты имеют средний диаметр 12–15микрометры (мкм) в мазки периферической крови. В суспензии нейтрофилы человека имеют средний диаметр 8,85 мкм.[12]
С эозинофил и базофил, они образуют класс полиморфно-ядерные клетки, названный в честь ядро 'многодольчатая форма (по сравнению с лимфоциты и моноциты, другие типы белых клеток). Ядро имеет характерный лопастный вид, отдельные доли соединены хроматин. Ядрышко исчезает по мере созревания нейтрофилов, что происходит только в некоторых других типах ядерных клеток.[13]:168 До 17% ядер женских нейтрофилов человека имеют придаток в форме голени, который содержит инактивированная Х-хромосома.[14] В цитоплазме аппарат Гольджи маленький, митохондрии и рибосомы редки, а грубые эндоплазматический ретикулум отсутствует.[13]:170 Цитоплазма также содержит около 200 гранул, из которых треть - азурофильный.[13]:170
По мере созревания нейтрофилы будут демонстрировать возрастающую сегментацию (многие сегменты ядра). У нормального нейтрофила должно быть 3–5 сегментов. Гиперсегментация - это ненормально, но встречается при некоторых расстройствах, в первую очередь витамин B12 недостаток. Это отмечается при ручном просмотре мазка крови и является положительным, когда большинство или все нейтрофилы имеют 5 или более сегментов.

Нейтрофилы - самые распространенные лейкоциты в организме человека (примерно 1011 производятся ежедневно); они составляют примерно 50–70% всех лейкоцитов (лейкоцитов). Заявленный нормальный диапазон показателей крови человека варьируется в зависимости от лаборатории, но количество нейтрофилов составляет 2,5–7,5 x 10.9/ L - стандартный нормальный диапазон. Люди Африканский и Ближневосточный при спуске счетчики могут быть ниже, но это все еще нормально.[15] В отчете нейтрофилы могут быть разделены на сегментированные нейтрофилы и группы.
Циркулируя в кровотоке и инактивированные, нейтрофилы имеют сферическую форму. После активации они меняют форму и становятся более аморфными или амеба -подобные и могут расширяться ложноножки как они охотятся за антигены.[16]
В 1973 году Санчес и др. обнаружили, что способность нейтрофилов поглощать бактерии снижается при приеме внутрь простых сахаров, таких как глюкоза, фруктоза, а также сахароза, мед и апельсиновый сок, в то время как прием крахмалов не имеет никакого эффекта. С другой стороны, голодание усиливает фагоцитарную способность нейтрофилов поглощать бактерии. Был сделан вывод, что функция, а не количество фагоцитов в поглощающих бактериях было изменено при приеме сахаров.[17] В 2007 году исследователи из Института биомедицинских исследований Уайтхеда обнаружили, что, учитывая набор сахаров на микробных поверхностях, нейтрофилы предпочтительно реагируют на некоторые типы сахаров. Нейтрофилы преимущественно поглощали и убивали мишени бета-1,6-глюкана по сравнению с мишенями бета-1,3-глюкана[18][19].
Разработка
Срок жизни
Средняя продолжительность жизни инактивированных нейтрофилов человека в кровотоке, по данным различных подходов, составляет от 5 до 135 часов.[20][21]
После активации они граничат (располагаются рядом с эндотелием кровеносных сосудов) и подвергаются селектин -зависимый захват с последующим интегрин -зависимая адгезия в большинстве случаев, после чего мигрируют в ткани, где выживают в течение 1-2 дней.[22]
Нейтрофилов гораздо больше, чем долгоживущих. моноцит /макрофаг фагоциты. А возбудитель (болезнетворный микроорганизм или вирус) скорее всего впервые столкнется с нейтрофилом. Некоторые эксперты предполагают, что короткое время жизни нейтрофилов эволюционный приспособление. Короткое время жизни нейтрофилов сводит к минимуму распространение тех патогенов, которые паразитировать фагоциты, потому что чем больше времени паразиты проводят вне хозяина клетка, тем больше вероятность, что они будут разрушены каким-либо компонентом защиты организма. Кроме того, нейтрофилы противомикробный продукты также могут повредить хозяину ткани, их короткая жизнь ограничивает повреждение хозяина во время воспаление.[22]
Нейтрофилы будут удалены после фагоцитоз патогенов макрофагами. ПЕКАМ-1 и фосфатидилсерин на поверхности клетки участвуют в этом процессе.
Функция
Хемотаксис
Нейтрофилы подвергаются процессу, называемому хемотаксис через амебовидное движение, что позволяет им перемещаться к участкам инфекции или воспаления. Рецепторы клеточной поверхности позволяют нейтрофилам обнаруживать химические градиенты молекул, таких как интерлейкин-8 (Ил-8), интерферон гамма (IFN-γ), C3a, C5a, и Лейкотриен B4, которые эти клетки используют для направления своей миграции.
Нейтрофилы имеют множество специфических рецепторов, включая рецепторы для дополнять цитокины, подобные интерлейкины и IFN-γ, хемокины, лектины, и другие белки. Они также экспрессируют рецепторы, чтобы обнаруживать и придерживаться эндотелий и Fc рецепторы за опсонин.[23]
В лейкоцитах, отвечающих на хемоаттрактант, то клеточная полярность регулируется деятельностью малых Ро гуанозинтрифосфатазы (Rho GTPases ) и фосфоинозитид-3-киназы (PI3K ). В нейтрофилах липидные продукты PI3K регулируют активацию Rho GTPases и необходимы для подвижность клеток. Они накапливаются асимметрично плазматическая мембрана на переднем крае поляризованных ячеек. Пространственно регулируя Rho GTPases и организуя передний край клетки, PI3Ks и их липидные продукты могут играть ключевую роль в установлении полярности лейкоцитов, как компасные молекулы, указывающие клетке, куда ей ползать.
На мышах было показано, что в определенных условиях нейтрофилы обладают определенным типом миграционного поведения, называемым скопление нейтрофилов во время которого они очень скоординированно мигрируют, накапливаются и группируются в места воспаления.[24]
Антимикробная функция
Будучи высоко подвижный нейтрофилы быстро собираются в фокусе инфекционное заболевание, привлеченный цитокины выражается активированным эндотелий, тучные клетки, и макрофаги. Нейтрофилы экспрессируют[25] и высвобождают цитокины, которые, в свою очередь, усиливают воспалительные реакции со стороны некоторых других типов клеток.
Помимо рекрутирования и активации других клеток иммунной системы, нейтрофилы играют ключевую роль в передовой защите от вторжения патогенов. У нейтрофилов есть три метода прямой атаки на микроорганизмы: фагоцитоз (проглатывание), дегрануляция (высвобождение растворимых антимикробных средств) и образование внеклеточные ловушки нейтрофилов (NET).[26]
Фагоцитоз

Нейтрофилы фагоциты, способные заглатывать микроорганизмы или частицы. Чтобы мишени были распознаны, они должны быть покрыты опсонины —Процесс, известный как опсонизация антител.[16] Они могут усвоить и убить многих микробы, каждое фагоцитарное событие, приводящее к образованию фагосома в котором активные формы кислорода секретируются гидролитические ферменты. Потребление кислорода во время образования активных форм кислорода получило название "респираторный взрыв ", хотя и не связан с дыханием или производством энергии.
Дыхательный взрыв включает в себя активацию фермент НАДФН оксидаза, который производит большое количество супероксид, активная форма кислорода. Супероксид самопроизвольно распадается или расщепляется ферментами, известными как супероксиддисмутазы (Cu / ZnSOD и MnSOD) в перекись водорода, которая затем превращается в хлорноватистая кислота (HClO) ферментом зеленого гема миелопероксидаза. Считается, что бактерицидных свойств HClO достаточно для уничтожения бактерий, фагоцитируемых нейтрофилом, но вместо этого это может быть этапом, необходимым для активации протеаз.[27]
Хотя нейтрофилы могут убить множество микробов, взаимодействие нейтрофилов с микробами и молекулами, продуцируемыми микробами, часто изменяет оборот нейтрофилов. Способность микробов изменять судьбу нейтрофилов сильно различается, может быть специфичной для микробов и колеблется от продления жизни нейтрофилов до быстрого лизиса нейтрофилов после фагоцитоза. Chlamydia pneumoniae и Neisseria gonorrhoeae сообщалось, что задерживает апоптоз нейтрофилов.[28][29][30] Таким образом, некоторые бактерии - и те, которые являются преимущественно внутриклеточными патогенами - могут увеличивать продолжительность жизни нейтрофилов, нарушая нормальный процесс спонтанного апоптоза и / или PICD (гибель клеток, вызванная фагоцитозом). На другом конце спектра некоторые патогены, такие как Streptococcus pyogenes способны изменять судьбу нейтрофилов после фагоцитоза, способствуя быстрому лизису клеток и / или ускоряя апоптоз до точки вторичного некроза.[31][32]
Дегрануляция
Нейтрофилы также выделяют набор белков в трех типах гранул с помощью процесса, называемого дегрануляция. Содержимое этих гранул обладает антимикробными свойствами и помогает бороться с инфекцией.
| Тип гранул | Протеин |
| Азурофильный гранулы (или «первичные гранулы») | Миелопероксидаза, бактерицидный протеин, повышающий проницаемость (BPI), дефенсины, а сериновые протеазы нейтрофильная эластаза и катепсин G |
| Специфические гранулы (или «вторичные гранулы») | Щелочная фосфатаза, лизоцим, НАДФН оксидаза, коллагеназа, лактоферрин, гистаминаза,[33] и кателицидин |
| Третичные гранулы | Катепсин, желатиназа, и коллагеназа |
Внеклеточные ловушки нейтрофилов
В 2004 году Бринкманн и его коллеги описали поразительное наблюдение, что активация нейтрофилов вызывает высвобождение веб-подобных структур ДНК; это представляет собой третий механизм уничтожения бактерий.[34] Эти внеклеточные ловушки нейтрофилов (NET) представляют собой сеть волокон, состоящих из хроматин и сериновые протеазы [35] которые улавливают и убивают внеклеточные микробы. Предполагается, что NET обеспечивают высокую локальную концентрацию противомикробных компонентов и связывают, обезвреживают и убивают микробы независимо от поглощения фагоцитами. В дополнение к их возможным антимикробным свойствам NET могут служить физическим барьером, предотвращающим дальнейшее распространение патогенов. Улавливание бактерий может быть особенно важной ролью NET в сепсис, где сети образуются внутри кровеносных сосудов.[36] Недавно было показано, что NET играют роль в воспалительных заболеваниях, поскольку NET могут быть обнаружены в преэклампсия воспалительное заболевание, связанное с беременностью, при котором, как известно, активируются нейтрофилы.[37] Образование сети нейтрофилов также может влиять на сердечно-сосудистые заболевания, так как NET могут влиять тромб формирование в коронарные артерии.[38][39]Теперь известно, что сети демонстрируют полезныетромботический влияет на оба in vitro[40] и in vivo.[41][42]
Клиническое значение
Низкое количество нейтрофилов называется нейтропения. Это может быть врожденный (развивается при рождении или до него) или может развиться позже, как в случае апластическая анемия или некоторые виды лейкемия. Это также может быть побочный эффект из медикамент, наиболее заметно химиотерапия. Нейтропения делает человека очень восприимчивым к инфекциям. Это также может быть результатом колонизации внутриклеточными нейтрофильными паразитами.
В дефицит альфа-1-антитрипсина, важный фермент нейтрофилов эластаза не подавляется должным образом альфа-1-антитрипсин, что приводит к чрезмерному повреждению тканей при воспалении, наиболее заметным из которых является эмфизема легких. Отрицательные эффекты эластазы были также показаны в случаях, когда нейтрофилы чрезмерно активированы (у здорового человека) и высвобождают фермент во внеклеточное пространство. Нерегулируемая активность эластазы нейтрофилов может привести к нарушению легочного барьера, проявляя симптомы, соответствующие острое повреждение легких.[43] Фермент также влияет на активность макрофагов, расщепляя их толл-подобные рецепторы (TLR) и подавление цитокин экспрессия путем ингибирования ядерной транслокации NF-κB.[44]
В Семейная средиземноморская лихорадка (FMF), мутация в пирин (или же маренострин ) ген, который экспрессируется в основном в нейтрофильных гранулоцитах, приводит к конститутивно активному острофазовый ответ и вызывает приступы высокая температура, артралгия, перитонит, и - в конце концов - амилоидоз.[45]
Снижение функции нейтрофилов было связано с гипергликемия. Дисфункция биохимического пути нейтрофилов миелопероксидаза а также снижение дегрануляции связаны с гипергликемией.[46]
В Абсолютное количество нейтрофилов (ANC) также используется в диагностике и прогнозировании. ANC - золотой стандарт для определения степени нейтропении и, следовательно, нейтропенической лихорадки. Любой АНК <1500 клеток / мм3 считается нейтропенией, но <500 клеток / мм3 считается тяжелым.[47] Есть также новое исследование, связывающее АНК с инфаркт миокарда как помощь в ранней диагностике.[48][49]
Нейтрофильные антигены
Распознаются пять (HNA 1-5) наборов нейтрофильных антигенов.[50] Три антигена HNA-1 (a-c) расположены на низкоаффинном рецепторе Fc-γ IIIb (FCGR3B:CD16b ) Единственный известный антиген HNA-2a расположен на CD177. Система антигенов HNA-3 включает два антигена (3a и 3b), которые расположены на седьмом экзоне гена CLT2 (SLC44A2 ). Каждая антигенная система HNA-4 и HNA-5 имеет по два известных антигена (a и b) и расположена в β2 интегрин. HNA-4 находится в цепи αM (CD11b ), а HNA-5 располагается на интегриновой единице αL (CD11a ).
Субпопуляции

Две функционально неравные субпопуляции нейтрофилов были идентифицированы на основании различных уровней образования их активных метаболитов кислорода, проницаемости мембран, активности ферментной системы и способности к инактивации. Клетки одной субпопуляции с высокой проницаемостью мембраны (нейтрофилы-киллеры) интенсивно генерируют реактивные метаболиты кислорода и инактивируются в результате взаимодействия с субстратом, тогда как клетки другой субпопуляции (клетки нейтрофилов) вырабатывают активные формы кислорода менее интенсивно, не прилипают к субстрату и сохраняют свою активность.[51][52][53][54][55] Дополнительные исследования показали, что опухоли легких могут инфильтрироваться различными популяциями нейтрофилов.[56]
видео
Можно увидеть, как быстро движущийся нейтрофил занимает несколько конидии в течение 2 часов с одним кадром каждые 30 секунд.
Здесь можно увидеть, что нейтрофил избирательно занимает несколько Candida дрожжи (флуоресцентно маркированный зеленым), несмотря на несколько контактов с Aspergillus fumigatus конидии (немаркированные, белые / прозрачные) в 3-D коллаген матрица. Время съемки составляло 2 часа с одним кадром каждые 30 секунд.
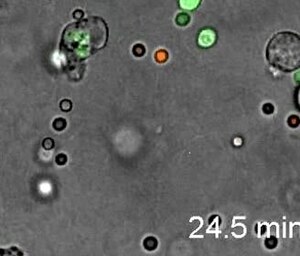
[1] Нейтрофилы демонстрируют направленную амебоидную подвижность инфицированных подушечек стопы и фаланг. Прижизненная визуализация проводилась на подушечках лап мышей LysM-eGFP через 20 минут после заражения Listeria monocytogenes.[57]
Дополнительные изображения

Родословная клеток крови

Более полные родословные


Рекомендации
- ^ Актер Джей (2012). Интегрированный обзор иммунологии и микробиологии Elsevier (Второе изд.). Дои:10.1016 / B978-0-323-07447-6.00002-8.
- ^ Эрмерт Д., Ниемек М.Дж., Рем М., Глентхой А., Боррегаард Н., Urban CF (август 2013 г.). «Candida albicans убегает от нейтрофилов мыши». Журнал биологии лейкоцитов. 94 (2): 223–36. Дои:10.1189 / jlb.0213063. PMID 23650619. S2CID 25619835.
- ^ Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (май 2000 г.). «Нейтрофилы: молекулы, функции и патофизиологические аспекты». Лабораторные исследования; Журнал технических методов и патологии. 80 (5): 617–53. Дои:10.1038 / labinvest.3780067. PMID 10830774. S2CID 22536645.
- ^ Клебанофф SJ, Кларк RA (1978). Нейтрофилы: функции и клинические нарушения. Эльзевир / Северная Голландия Амстердам. ISBN 978-0-444-80020-6.
- ^ Натан С. (март 2006 г.). «Нейтрофилы и иммунитет: проблемы и возможности». Обзоры природы. Иммунология. 6 (3): 173–82. Дои:10.1038 / nri1785. PMID 16498448. S2CID 1590558.
- ^ Якобс Л., Наврот Т.С., де Геус Б., Мееузен Р., Дегреув Б., Бернар А., Сугис М., Немери Б., Панис Л.И. (октябрь 2010 г.). «Субклинические реакции у здоровых велосипедистов, кратковременно подвергшихся воздействию загрязнения воздуха, связанного с дорожным движением: исследование вмешательства». Состояние окружающей среды. 9 (64): 64. Дои:10.1186 / 1476-069X-9-64. ЧВК 2984475. PMID 20973949.
- ^ Waugh DJ, Wilson C (ноябрь 2008 г.). «Путь интерлейкина-8 при раке». Клинические исследования рака. 14 (21): 6735–41. Дои:10.1158 / 1078-0432.CCR-07-4843. PMID 18980965. S2CID 9415085.
- ^ Де Ларко Дж. Э., Вюрц Б. Р., Ферч Л. Т. (август 2004 г.). «Потенциальная роль нейтрофилов в стимулировании метастатического фенотипа опухолей, выделяющих интерлейкин-8». Клинические исследования рака. 10 (15): 4895–900. Дои:10.1158 / 1078-0432.CCR-03-0760. PMID 15297389. S2CID 9782495.
- ^ Ю СК, Старнес Т.В., Дэн Кью, Хаттенлохер А. (ноябрь 2011 г.). «Lyn - это окислительно-восстановительный датчик, который опосредует привлечение лейкоцитов к ране in vivo». Природа. 480 (7375): 109–12. Bibcode:2011Натура.480..109л. Дои:10.1038 / природа10632. ЧВК 3228893. PMID 22101434.
- ^ Барер М.Р. (2012). «Естественная история заражения». Медицинская микробиология. Эльзевир. С. 168–173. Дои:10.1016 / b978-0-7020-4089-4.00029-9. ISBN 978-0-7020-4089-4.
- ^ Коэн С., Бернс Р.К. (2002). Пути пульпы (8-е изд.). Сент-Луис: Мосби. п. 465.
- ^ Niemiec MJ, De Samber B, Garrevoet J, Vergucht E, Vekemans B, De Rycke R, Björn E, Sandblad L, Wellenreuther G, Falkenberg G, Cloetens P, Vincze L, Urban CF (июнь 2015 г.). «Микроэлементный ландшафт покоящихся и активированных нейтрофилов человека на субмикрометровом уровне». Металломика. 7 (6): 996–1010. Дои:10.1039 / c4mt00346b. PMID 25832493.
- ^ а б c Цукер-Франклин Д., Гривз М.Ф., Гросси К.Э., Мармонт А.М. (1988). «Нейтрофилы». Атлас клеток крови: функции и патология. 1 (2-е изд.). Филадельфия: Леа и Фербигер. ISBN 978-0-8121-1094-4.
- ^ Карни Р.Дж., Ванх Л.Дж., Санчес А.Дж. (август 2001 г.). «Неслучайное расположение и ориентация неактивной Х-хромосомы в ядрах нейтрофилов человека». Хромосома. 110 (4): 267–274. Дои:10.1007 / s004120100145. PMID 11534818.
- ^ Reich D, Nalls MA, Kao WH, Akylbekova EL, Tandon A, Patterson N, Mullikin J, Hsueh WC, Cheng CY, Coresh J, Boerwinkle E, Li M, Waliszewska A, Neubauer J, Li R, Leak TS, Ekunwe L , Files JC, Hardy CL, Zmuda JM, Taylor HA, Ziv E, Harris TB, Wilson JG (январь 2009 г.). «Снижение количества нейтрофилов у лиц африканского происхождения связано с регуляторным вариантом рецептора антигена Даффи для гена хемокинов». PLOS Genetics. 5 (1): e1000360. Дои:10.1371 / journal.pgen.1000360. ЧВК 2628742. PMID 19180233.
- ^ а б Эдвардс SW (1994). Биохимия и физиология нейтрофилов. Издательство Кембриджского университета. п. 6. ISBN 978-0-521-41698-6.
- ^ Санчес А., Ризер Дж. Л., Лау Х. С., Яхику П. Я., Уиллард Р. Э., Макмиллан П. Дж., Чо С. Ю., Мэги А. Р., Регистр Ю. Д. (ноябрь 1973 г.). «Роль сахаров в нейтрофильном фагоцитозе человека». Американский журнал клинического питания. 26 (11): 1180–4. Дои:10.1093 / ajcn / 26.11.1180. PMID 4748178.
Эти данные предполагают, что функция, а не количество фагоцитов изменялось при приеме сахаров. Это предполагает участие глюкозы и других простых углеводов в контроле фагоцитоза и показывает, что эффекты длятся не менее 5 часов. С другой стороны, голодание продолжительностью 36 или 60 часов значительно увеличило (P <0,001) фагоцитарный индекс.
- ^ Рубин-Бехерано И., Абейхон С., Магнелли П., Грисафи П., Финк Г.Р. (июль 2007 г.). «Фагоцитоз нейтрофилов человека стимулируется уникальным компонентом клеточной стенки грибов». Клеточный хозяин и микроб. 2 (1): 55–67. Дои:10.1016 / j.chom.2007.06.002. ЧВК 2083279. PMID 18005717.
- ^ Кнеллер А (2007). «Белые кровяные тельца придирчивы к сахару». Институт Уайтхеда. Получено 2013-08-09.
- ^ Так Т., Тесселаар К., Пиллэй Дж., Борганс Дж. А., Кендерман Л. (октябрь 2013 г.). «Каков ваш возраст снова? Повторное определение периода полураспада нейтрофилов человека». Журнал биологии лейкоцитов. 94 (4): 595–601. Дои:10.1189 / jlb.1112571. PMID 23625199. S2CID 40113921.
- ^ Пиллэй Дж., Ден Брабер И., Врисекоп Н., Кваст Л. М., де Бур Р. Дж., Борганс Дж. А., Тесселаар К., Кендерман Л. (июль 2010 г.). «Мечение in vivo с помощью 2H2O показывает, что продолжительность жизни нейтрофилов человека составляет 5,4 дня». Кровь. 116 (4): 625–7. Дои:10.1182 / кровь-2010-01-259028. PMID 20410504.
- ^ а б Уитер PR, Стивенс А. (2002). Основная гистопатология Уитера: цветной атлас и текст. Эдинбург: Черчилль Ливингстон. ISBN 978-0-443-07001-3.
- ^ Серхан К.Н., Уорд ПА, Гилрой Д.В. (2010). Основы воспаления. Издательство Кембриджского университета. С. 53–54. ISBN 978-0-521-88729-8.
- ^ Леммерманн Т., Афонсо П.В., Ангерманн Б.Р., Ван Дж.М., Кастенмюллер В., родительский центр сертификации, Жермен Р.Н. (июнь 2013 г.). «Рой нейтрофилов требует LTB4 и интегринов на участках гибели клеток in vivo». Природа. 498 (7454): 371–5. Bibcode:2013Натура 498..371л. Дои:10.1038 / природа12175. ЧВК 3879961. PMID 23708969.
- ^ Ear T, McDonald PP (апрель 2008 г.). «Генерация цитокинов, активация промотора и окислительно-независимая активация NF-kappaB в модели трансфицируемых нейтрофильных клеток человека». BMC Иммунология. 9: 14. Дои:10.1186/1471-2172-9-14. ЧВК 2322942. PMID 18405381.
- ^ Хики MJ, Кубес P (май 2009 г.). «Внутрисосудистый иммунитет: встреча возбудителя и хозяина в кровеносных сосудах». Обзоры природы. Иммунология. 9 (5): 364–75. Дои:10.1038 / nri2532. PMID 19390567. S2CID 8068543.
- ^ Сегал А.В. (2005). «Как нейтрофилы убивают микробы». Ежегодный обзор иммунологии. 23 (5): 197–223. Дои:10.1146 / annurev.immunol.23.021704.115653. ЧВК 2092448. PMID 15771570.
- ^ Simons MP, et al. (2006). «Neisseria gonorrhoeae задерживает начало апоптоза в полиморфно-ядерных лейкоцитах». Клеточный микробиол. 8 (11): 1780–90. Дои:10.1111 / j.1462-5822.2006.00748.x. PMID 16803582. S2CID 25253422.
- ^ Чен А. и др. (2004). «Опосредованное Neisseria gonorrhoeae ингибирование апоптотической передачи сигналов в полиморфно-ядерных лейкоцитах». Заразить. Иммунная. 79 (11): 4447–58. Дои:10.1128 / IAI.01267-10. ЧВК 3257915. PMID 21844239.
- ^ van Zandbergen G, et al. (2004). «Chlamydia pneumoniae размножаются в нейтрофильных гранулоцитах и задерживают их спонтанный апоптоз». J. Immunol. 172 (3): 1768–76. Дои:10.4049 / jimmunol.172.3.1768. PMID 14734760. S2CID 27422510.
- ^ Кобаяши С.Д. и др. (2003). «Бактериальные патогены модулируют программу дифференцировки апоптоза нейтрофилов человека». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 100 (19): 10948–10953. Дои:10.1073 / pnas.1833375100. ЧВК 196908. PMID 12960399.
- ^ Скотт Д.К. и др. (2017). «Влияние микробов на жизнь и смерть нейтрофилов». Микробиол фронтальной клеточной инфекции. 7 (159): 159. Дои:10.3389 / fcimb.2017.00159. ЧВК 5410578. PMID 28507953.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Рингель Э. У., Сотер Н. А., Остин К. Ф. (август 1984 г.). «Локализация гистаминазы в конкретной грануле нейтрофила человека». Иммунология. 52 (4): 649–58. ЧВК 1454675. PMID 6430792.
- ^ Бринкманн В., Райхард Ю., Гусманн К., Фаулер Б., Улеманн Ю., Вайс Д.С., Вайнраух Ю., Зихлинский А. (март 2004 г.). «Нейтрофильные внеклеточные ловушки убивают бактерии». Наука. 303 (5663): 1532–5. Bibcode:2004Научный ... 303.1532B. Дои:10.1126 / science.1092385. PMID 15001782. S2CID 21628300.
- ^ Урбан К.Ф., Эрмерт Д., Шмид М., Абу-Абед Ю., Гусманн С., Накен В., Бринкманн В., Юнгблут П.Р., Зихлински А. (октябрь 2009 г.). «Внеклеточные ловушки нейтрофилов содержат кальпротектин, цитозольный белковый комплекс, участвующий в защите хозяина от Candida albicans». Патогены PLOS. 5 (10): e1000639. Дои:10.1371 / journal.ppat.1000639. ЧВК 2763347. PMID 19876394.
- ^ Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM, Patel KD, Chakrabarti S, McAvoy E, Sinclair GD, Keys EM, Allen-Vercoe E, Devinney R, Doig CJ, Green FH, Kubes P ( Апрель 2007 г.). «TLR4 тромбоцитов активирует внеклеточные ловушки нейтрофилов, чтобы заманить бактерии в септическую кровь». Природа Медицина. 13 (4): 463–9. Дои:10,1038 / нм 1565. PMID 17384648. S2CID 22372863.
- ^ Гупта А.К., Хаслер П., Хольцгрев В., Хан С. (июнь 2007 г.). «Нейтрофильные сети: новый фактор, способствующий развитию плацентарной гипоксии, связанной с преэклампсией?» (PDF). Семинары по иммунопатологии. 29 (2): 163–7. Дои:10.1007 / s00281-007-0073-4. PMID 17621701. S2CID 12887059.
- ^ Хойер, Ф. Ф.; Нахрендорф, М. (14 февраля 2017 г.). «Вклад нейтрофилов в ишемическую болезнь сердца». Европейский журнал сердца. 38 (7): 465–472. Дои:10.1093 / eurheartj / ehx017. PMID 28363210.
- ^ Мангольд, А; Псевдоним, S; Scherz, T; Hofbauer, T; Jakowitsch, J; Панценбёк, А; Саймон, Д.; Laimer, D; Бангерт, К; Каммерландер, А; Mascherbauer, J; Зима, МП; Distelmaier, K; Adlbrecht, C; Прейсснер, К.Т.; Ланг, И.М. (27 марта 2015 г.).«Бремя внеклеточных ловушек коронарных нейтрофилов и дезоксирибонуклеазная активность при остром коронарном синдроме с подъемом сегмента ST являются предикторами разрешения сегмента ST и размера инфаркта». Циркуляционные исследования. 116 (7): 1182–92. Дои:10.1161 / CIRCRESAHA.116.304944. PMID 25547404.
- ^ Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD, Wrobleski SK, Wakefield TW, Hartwig JH, Wagner DD (сентябрь 2010 г.). «Ловушки внеклеточной ДНК способствуют тромбозу». Труды Национальной академии наук Соединенных Штатов Америки. 107 (36): 15880–5. Bibcode:2010PNAS..10715880F. Дои:10.1073 / pnas.1005743107. ЧВК 2936604. PMID 20798043.
- ^ Брилл А., Фукс Т.А., Савченко А.С., Томас Г.М., Мартинод К., Де Мейер С.Ф., Бхандари А.А., Вагнер Д.Д. (январь 2012 г.). «Внеклеточные ловушки нейтрофилов способствуют тромбозу глубоких вен у мышей». Журнал тромбоза и гемостаза. 10 (1): 136–44. Дои:10.1111 / j.1538-7836.2011.04544.x. ЧВК 3319651. PMID 22044575.
- ^ Borissoff JI, ten Cate H (сентябрь 2011 г.). «От высвобождения внеклеточных ловушек нейтрофилов до тромбоза: чрезмерный механизм защиты хозяина?». Журнал тромбоза и гемостаза. 9 (9): 1791–4. Дои:10.1111 / j.1538-7836.2011.04425.x. PMID 21718435. S2CID 5368241.
- ^ Кавабата К., Хагио Т., Мацуока С. (сентябрь 2002 г.). «Роль нейтрофильной эластазы в остром повреждении легких». Европейский журнал фармакологии. 451 (1): 1–10. Дои:10.1016 / S0014-2999 (02) 02182-9. PMID 12223222.
- ^ Домон Х, Нагай К., Маэкава Т, Ода М, Йонезава Д., Такеда В., Хиёси Т, Тамура Х, Ямагути М, Кавабата С., Терао Y (2018). «Эластаза нейтрофилов разрушает иммунный ответ, расщепляя толл-подобные рецепторы и цитокины при пневмококковой пневмонии». Границы иммунологии. 9: 732. Дои:10.3389 / fimmu.2018.00732. ЧВК 5996908. PMID 29922273.
- ^ Озен С (июль 2003 г.). «Семейная средиземноморская лихорадка: возвращаясь к древней болезни». Европейский журнал педиатрии. 162 (7–8): 449–454. Дои:10.1007 / s00431-003-1223-х. PMID 12751000. S2CID 3464945.
- ^ Сю Ф., Станойчич М., Диао Л., Йешке М.Г. (8 мая 2014 г.). «Стресс-гипергликемия, лечение инсулином и клетки врожденного иммунитета». Международный журнал эндокринологии. 2014: 486403. Дои:10.1155/2014/486403. ЧВК 4034653. PMID 24899891.
- ^ Аль-Гвайз Л.А., Бабай Х.Х. (2007). «Диагностическая ценность абсолютного количества нейтрофилов, количества полос и морфологических изменений нейтрофилов в прогнозировании бактериальных инфекций». Медицинские принципы и практика. 16 (5): 344–7. Дои:10.1159/000104806. PMID 17709921. S2CID 5499290.
- ^ Хан Х.А., Алхомида А.С., Собки С.Х., Могайри А.А., Коронки Х.Э. (2012). «Количество клеток крови и их корреляция с креатинкиназой и С-реактивным белком у пациентов с острым инфарктом миокарда». Международный журнал клинической и экспериментальной медицины. 5 (1): 50–5. ЧВК 3272686. PMID 22328948.
- ^ Basili S, Di Francoi M, Rosa A., Ferroni P, Diurni V, Scarpellini MG, Bertazzoni G (апрель 2004 г.). «Абсолютное количество нейтрофилов и уровни фибриногена как помощь в ранней диагностике острого инфаркта миокарда». Acta Cardiologica. 59 (2): 135–40. Дои:10.2143 / ac.59.2.2005167. PMID 15139653. S2CID 37382677.
- ^ Chu HT, Lin H, Tsao TT, Chang CF, Hsiao WW, Yeh TJ, Chang CM, Liu YW, Wang TY, Yang KC, Chen TJ, Chen JC, Chen KC, Kao CY (сентябрь 2013 г.). «Генотипирование нейтрофильных антигенов человека (HNA) на основе данных секвенирования всего генома». BMC Medical Genomics. 6 (1): 31. Дои:10.1186/1755-8794-6-31. ЧВК 3849977. PMID 24028078.
- ^ а б Игнатов Д.Ю. (2012). Функциональная гетерогенность нейтрофилов человека и их роль в регуляции количества лейкоцитов периферической крови (Кандидат наук). Донецкий национальный медицинский университет. Дои:10.13140 / RG.2.2.35542.34884.
- ^ Герасимов И.Г., Игнатов Д.И. (2001). «[Функциональная гетерогенность нейтрофилов крови человека: образование активных форм кислорода]». Цитология. 43 (5): 432–6. PMID 11517658.
- ^ Герасимов И.Г., Игнатов Д.И. (2004). «[Активация нейтрофилов in vitro]». Цитология. 46 (2): 155–8. PMID 15174354.
- ^ Герасимов И.Г., Игнатов Д.И., Котельницкий М.А. (2005). «[Восстановление тетразолия нитросинего нейтрофилами крови человека. I. Влияние pH]». Цитология. 47 (6): 549–53. PMID 16708848.
- ^ Герасимов И.Г., Игнатов Д.И. (2005). «[Восстановление тетразолия нитросинего нейтрофилами крови человека. II. Влияние ионов натрия и калия]». Цитология. 47 (6): 554–8. PMID 16708849.
- ^ Зилионис Р., Энгблом С., Пфиршке С., Савова В., Земмур Д., Саатчоглу Х.Д., Кришнан И., Марони Дж., Мейеровиц К.В., Кервин С.М., Чой С., Ричардс В.Г., Де Риенцо А., Тенен Д.Г., Буэно Р., Левантини Э., Питте MJ, Klein AM (май 2019 г.). «Одноклеточная транскриптомика рака легких человека и мыши выявляет консервативные миелоидные популяции у отдельных людей и видов». Иммунитет. 50 (5): 1317–1334.e10. Дои:10.1016 / j.immuni.2019.03.009. ЧВК 6620049. PMID 30979687.
- ^ Грэм Д.Б., Зинсельмейер Б.Х., Маскареньяс Ф., Дельгадо Р., Миллер М.Дж., Сват В. (2009). Unutmaz D (ред.). «Передача сигналов ITAM с помощью факторов обмена генов гуаниновых нуклеотидов семейства Vav регулирует скорость интерстициального транзита нейтрофилов in vivo». PLOS ONE. 4 (2): e4652. Bibcode:2009PLoSO ... 4.4652G. Дои:10.1371 / journal.pone.0004652. ЧВК 2645696. PMID 19247495.
внешняя ссылка
- Информация о нейтропении
- Калькулятор абсолютного количества нейтрофилов
- Содержание и распределение следовых элементов нейтрофилов
| Авторитетный контроль |
|---|