V (D) J рекомбинация - V(D)J recombination
V (D) J рекомбинация это механизм соматическая рекомбинация что происходит только в развивающихся лимфоциты на ранних стадиях созревания Т- и В-клеток. Это приводит к очень разнообразному репертуару антитела / иммуноглобулины и Рецепторы Т-клеток (TCR) найдено в В-клетки и Т-клетки, соответственно. Этот процесс является определяющей чертой адаптивная иммунная система.
Рекомбинация V (D) J у млекопитающих происходит в первичных лимфоидных органах (Костный мозг для В-клеток и вилочковая железа для Т-клеток) и почти случайным образом перестраивает изменяемые (V), соединяющие (J) и в некоторых случаях сегменты гена разнообразия (D). Процесс в конечном итоге приводит к роману аминокислота последовательности в антигенсвязывающих областях иммуноглобулинов и TCR, которые позволяют распознавать антигены от почти всех патогенов, включая бактерии, вирусы, паразиты, и черви а также «измененные клетки себя», как видно на рак. Признание также может быть аллергический в природе (например к пыльца или другой аллергены ) или может соответствовать тканям хозяина и приводить к аутоиммунитет.
В 1987 г. Сусуму Тонегава был награжден Нобелевская премия по физиологии и медицине «За открытие генетического принципа создания разнообразия антител».[1]
Фон
Человек антитело молекулы (в том числе Рецепторы В-клеток ) состоят из тяжелых и легких цепей, каждая из которых содержит оба постоянный (C) и Переменная (V) области, генетически кодируемые на трех места:
- Тяжелый локус иммуноглобулина (IGH @ ) на хромосоме 14, содержащей сегменты гена для тяжелая цепь иммуноглобулина.
- Локус иммуноглобулина каппа (κ) (IGK @ ) на хромосоме 2, содержащей генные сегменты для части легкая цепь иммуноглобулина.
- Локус иммуноглобулина лямбда (λ) (IGL @ ) на хромосоме 22, содержащей генные сегменты для оставшейся части легкая цепь иммуноглобулина.
Каждый ген тяжелой цепи или легкой цепи содержит несколько копий трех различных типов генных сегментов для вариабельных областей белков антител. Например, область тяжелой цепи иммуноглобулина человека содержит 2 генных сегмента констант (Cμ и Cδ) и 44 вариабельных (V) генных сегмента, плюс 27 генных сегментов разнообразия (D) и 6 генных сегментов соединения (J).[2] Гены легкой цепи содержат либо один (Cκ), либо четыре (Cλ) постоянных генных сегмента с множеством генных сегментов V и J, но не имеют генных сегментов D.[3] Перестройка ДНК заставляет одну копию каждого типа сегмента гена попасть в любой данный лимфоцит, создавая огромный репертуар антител; примерно 3 × 1011 комбинации возможны, хотя некоторые удаляются из-за самореактивности.
Наиболее Рецепторы Т-клеток состоят из переменной альфа-цепь и бета-цепочка. Гены рецепторов Т-клеток подобны генам иммуноглобулинов тем, что они также содержат несколько сегментов генов V, D и J в своих бета-цепях (и сегменты генов V и J в их альфа-цепях), которые перестраиваются во время развития лимфоцита в снабдить эту клетку уникальным рецептором антигена. Рецептор Т-клеток в этом смысле является топологическим эквивалентом антигенсвязывающего фрагмента антитела, оба являются частью суперсемейства иммуноглобулинов.
Аутоиммунный ответ предотвращается путем устранения клеток, которые реагируют на себя. Это происходит в тимусе путем тестирования клетки на набор аутоантигенов, экспрессируемых через функцию аутоиммунный регулятор (AIRE). Локус легкой цепи лямбда иммуноглобулина содержит гены, кодирующие белок, которые могут быть потеряны при его перестройке. Это основано на физиологическом механизме и не является патогенетическим для лейкозов или лимфом. Ячейка сохраняется, если она создает успешный продукт, который не реагирует на себя, в противном случае она удаляется через апоптоз.
Иммуноглобулины
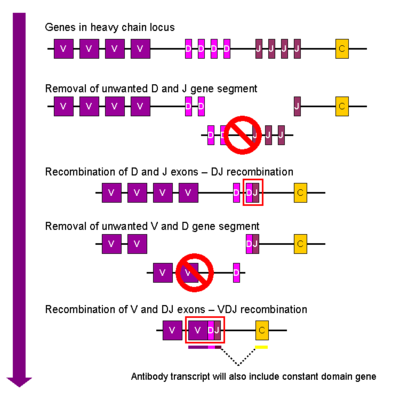
Тяжелая цепь
В развивающихся В клетка, первое событие рекомбинации происходит между одним D и одним сегментом J гена локуса тяжелой цепи. Любая ДНК между этими двумя сегментами гена удаляется. За этой D-J-рекомбинацией следует присоединение одного генного сегмента V из области, расположенной выше вновь образованного комплекса DJ, с образованием реаранжированного генного сегмента VDJ. Все остальные генные сегменты между сегментами V и D теперь удалены из генома клетки. Начальный стенограмма (несплицированная РНК), содержащая область VDJ тяжелой цепи и константу му и дельта цепи (Cμ и Cδ). (т.е. первичная расшифровка содержит сегменты: V-D-J-Cμ-Cδ). Первичная РНК обрабатывается для добавления полиаденилированный (поли-A) хвост после Cμ цепи и удалить последовательность между сегментом VDJ и этим постоянным сегментом гена. Перевод этой мРНК приводит к продукции IgM белок тяжелой цепи.
Легкая цепь
Каппа (κ) и лямбда (λ) цепи локусов легкой цепи иммуноглобулина перестраиваются очень сходным образом, за исключением того, что в легких цепях отсутствует сегмент D. Другими словами, первая стадия рекомбинации легких цепей включает соединение цепей V и J с образованием комплекса VJ перед добавлением гена константной цепи во время первичной транскрипции. Трансляция сплайсированной мРНК для каппа- или лямбда-цепей приводит к образованию белка легкой цепи Ig κ или Ig λ.
Сборка тяжелой цепи Ig μ и одной из легких цепей приводит к образованию мембраносвязанной формы иммуноглобулина IgM, которая экспрессируется на поверхности незрелой В-клетки.
Рецепторы Т-клеток
В течение тимоцит развитие, Рецептор Т-клеток (TCR) цепи претерпевают по существу ту же последовательность событий упорядоченной рекомбинации, что и описанная для иммуноглобулинов. Рекомбинация D-to-J происходит сначала в β-цепи TCR. Этот процесс может включать либо присоединение Dβ1 генный сегмент к одному из шести Jβ1 сегменты или соединение Dβ2 генного сегмента к одному из шести Jβ2 сегмента.[3] После DJ-рекомбинации (как и выше) следует Vβ-к-DβJβ перестановки. Все генные сегменты между Vβ-Dβ-Jβ сегменты гена во вновь образованном комплексе удаляются, и синтезируется первичный транскрипт, который включает ген константного домена (Vβ-Dβ-Jβ-Cβ). Транскрипция мРНК сплайсирует любую промежуточную последовательность и позволяет транслировать полноразмерный белок для β-цепи TCR.
Перестройка альфа (α) цепи TCR следует за перестройкой β-цепи и напоминает перегруппировку V-J, описанную для легких цепей Ig (см. Выше). Сборка β- и α-цепей приводит к образованию αβ-TCR, который экспрессируется на большинстве Т-клетки.
Механизм
Ключевые ферменты и компоненты
Процесс рекомбинации V (D) J опосредуется рекомбиназой VDJ, которая представляет собой разнообразный набор ферментов. Ключевые задействованные ферменты: гены, активирующие рекомбинацию 1 и 2 (ТРЯПКА), терминальная дезоксинуклеотидилтрансфераза (TdT) и Артемида нуклеаза, член повсеместного негомологичное соединение концов (NHEJ) путь репарации ДНК.[4] Известно, что в процесс вовлечены несколько других ферментов, в том числе: ДНК-зависимая протеинкиназа (ДНК-ПК), Рентгеновский ремонт перекрестно комплементарный белок 4 (XRCC4), ДНК-лигаза IV, негомологичный коэффициент соединения концов 1 (NHEJ1; также известный как Cernunnos или XRCC4-подобный фактор [XLF]), недавно обнаруженный Paralog XRCC4 и XLF (PAXX) и ДНК-полимеразы λ и μ.[5] Некоторые задействованные ферменты специфичны для лимфоцитов (например, RAG, TdT), в то время как другие обнаруживаются в других типах клеток и даже повсеместно (например, Компоненты NHEJ).
Чтобы поддерживать специфичность рекомбинации, рекомбиназа V (D) J распознает и связывается с сигнальные последовательности рекомбинации (RSS), фланкирующие сегменты вариабельного (V), разнообразия (D) и присоединения (J) генов. RSS состоят из трех элементов: гептамера из семи консервативных нуклеотидов, спейсерной области длиной 12 или 23 пар оснований и нонамера из девяти консервативных нуклеотидов. Хотя большинство RSS различаются по последовательности, консенсусные гептамерные и неамерные последовательности представляют собой CACAGTG и ACAAAAACC, соответственно; и хотя последовательность спейсерной области плохо консервативна, длина высококонсервативна.[6][7] Длина спейсерной области соответствует приблизительно одному (12 пар оснований) или двум виткам (23 пары оснований) спирали ДНК. Следуя так называемому правилу 12/23, рекомбинируемые сегменты генов обычно примыкают к RSS разной длины спейсера (т.е., у одного есть «12RSS», а у другого «23RSS»).[8] Это важная особенность регуляции рекомбинации V (D) J.[9]
Процесс
Рекомбинация V (D) J начинается, когда рекомбиназа V (D) J (посредством активности RAG1) связывает RSS, фланкирующую сегмент кодирующего гена (V, D или J), и создает одноцепочечный разрыв в ДНК между первыми основание RSS (непосредственно перед гептамером) и кодирующий сегмент. Это по существу энергетически нейтрален (нет необходимости в Гидролиз АТФ ) и приводит к образованию свободной 3 ' гидроксильная группа и 5 ' фосфатная группа на одной пряди. Реактивная гидроксильная группа позиционируется рекомбиназой, чтобы атаковать фосфодиэфирная связь противоположной цепи, образуя два конца ДНК: заколка для волос (стебель-петля) на кодирующем сегменте и тупой конец на сигнальном сегменте.[10] Текущая модель состоит в том, что надрезание ДНК и образование шпильки происходит на обеих нитях одновременно (или почти так) в комплексе, известном как центр рекомбинации.[11][12][13][14]
Тупые сигнальные концы лигируют вместе заподлицо, чтобы сформировать круговой кусок ДНК, содержащий все промежуточные последовательности между кодирующими сегментами, известный как сигнальное соединение (хотя по своей природе круговой, его не следует путать с плазмида ). Хотя первоначально считалось, что они теряются во время последовательных делений клеток, есть доказательства того, что сигнальные суставы могут повторно войти в геном и привести к патологиям, активируя онкогены или прерывая ген-супрессор опухоли функция (ы) [Ref].
Кодирующие концы дополнительно обрабатываются до их лигирования несколькими событиями, которые в конечном итоге приводят к разнообразию соединений.[15] Процессинг начинается, когда DNA-PK связывается с каждым разорванным концом ДНК и привлекает несколько других белков, включая Artemis, XRCC4, ДНК-лигазу IV, Cernunnos и несколько ДНК-полимераз.[16] ДНК-ПК образует комплекс, который приводит к его аутофосфорилирование, что приводит к активации Artemis. Шпильки кодирующего конца открываются в результате активности Артемиды.[17] Если их открыть в центре, получится тупой конец ДНК; однако во многих случаях отверстие находится "не по центру" и приводит к тому, что на одной нити остаются дополнительные основания (выступ). Они известны как палиндромные (P) нуклеотиды из-за палиндромный характер последовательности образуется, когда ферменты репарации ДНК устраняют выступ.[18] Процесс открытия шпильки Artemis является критическим этапом рекомбинации V (D) J и является дефектным в модель тяжелого комбинированного иммунодефицита (scid) на мышах.
Затем XRCC4, Cernunnos и DNA-PK выравнивают концы ДНК и привлекают терминальную дезоксинуклеотидилтрансферазу (TdT), матрицу-независимую ДНК-полимеразу, которая добавляет нетематические (N) нуклеотиды к кодирующему концу. Добавление в основном случайное, но TdT действительно отдает предпочтение нуклеотидам G / C.[19] Как и все известные ДНК-полимеразы, TdT добавляет нуклеотиды к одной цепи в направлении от 5 'до 3'.[20]
Наконец, экзонуклеазы могут удалять основания с кодирующих концов (включая любые нуклеотиды P или N, которые могли образоваться). ДНК-полимеразы λ и μ затем вставляют дополнительные нуклеотиды по мере необходимости, чтобы сделать два конца совместимыми для соединения. Это стохастический процесс, поэтому может произойти любая комбинация добавления нуклеотидов P и N и экзонуклеолитического удаления (или вообще не произойдет). Наконец, обработанные кодирующие концы лигируют вместе ДНК-лигазой IV.[21]
Все эти процессы обработки приводят к тому, что антигенсвязывающая область очень вариабельна, даже когда рекомбинируются одни и те же генные сегменты. Рекомбинация V (D) J позволяет генерировать иммуноглобулины и Т-клеточные рецепторы к антигенам, с которыми ни организм, ни его предок (предки) не должны были ранее сталкиваться, что обеспечивает адаптивный иммунный ответ на новые патогены, которые развиваются, или на те, которые часто изменять (например, сезонный грипп ). Однако главное предостережение этого процесса заключается в том, что последовательность ДНК должна оставаться в кадре для поддержания правильной аминокислотной последовательности в конечном белковом продукте. Если результирующая последовательность выходит за рамки кадра, развитие клетки будет остановлено, и клетка не доживет до зрелости. Поэтому рекомбинация V (D) J - очень дорогостоящий процесс, который должен (и должен) строго регулироваться и контролироваться.
Смотрите также
- Рецептор В-клеток
- Рецептор Т-клеток
- Базельский институт иммунологии
- Чарльз М. Стейнберг
- NKT-клетка
- Ген, активирующий рекомбинацию
Рекомендации
- ^ "Нобелевская премия по физиологии и медицине 1987 г.". nobelprize.org. Получено 26 декабря 2014.
- ^ Ли А., Рю М., Чжоу Дж. И др. (Июнь 2004 г.). «Использование вариабельных, разнообразных и соединяющихся сегментов тяжелой цепи Ig у детей с острым лимфобластным лейкозом B-линии: значение для механизмов рекомбинации VDJ и патогенеза». Кровь. 103 (12): 4602–9. Дои:10.1182 / кровь-2003-11-3857. PMID 15010366.
- ^ а б Аббас, Абул К. (2018). «Развитие лимфоцитов и перестройка генов антигенных рецепторов». Клеточная и молекулярная иммунология (9-е изд.). Филадельфия, Пенсильвания: Эльзевир. ISBN 978-0-323-47978-3.
- ^ Ма, Юньмэй; Лу, Хайхуэй; Шварц, Клаус; Либер, Майкл (сентябрь 2005 г.). «Ремонт двунитевых разрывов ДНК с помощью пути соединения негомологичных концов ДНК: модель итеративной обработки». Клеточный цикл. 4 (9): 1193–1200. Дои:10.4161 / cc.4.9.1977. PMID 16082219.
- ^ Малу, Шрути; Малшетти, Видьясагар; Фрэнсис, Дайлия; Кортес, Патрисия (2012). «Роль негомологичного соединения концов в рекомбинации V (D) J». Иммунологические исследования. 54 (1–3): 233–246. Дои:10.1007 / s12026-012-8329-z. PMID 22569912.
- ^ Рамсден, Дейл; Баец, Кристин; Ву, Джиллиан (1994). «Сохранение последовательности в спейсерах последовательностей рекомбинационных сигналов». Исследования нуклеиновых кислот. 22 (10): 1785–1796. Дои:10.1093 / nar / 22.10.1785. ЧВК 308075. PMID 8208601.
- ^ Коуэлл, Линдси; Давила, Марко; Рамсден, Дейл; Келсо, Гарнетт (2004). «Вычислительные инструменты для понимания изменчивости последовательности в сигналах рекомбинации». Иммунологические обзоры. 200: 57–69. Дои:10.1111 / j.0105-2896.2004.00171.x. PMID 15242396.
- ^ ван Гент, Дик; Рамсден, Дейл; Геллерт, Мартин (1996). «Белки RAG1 и RAG2 устанавливают правило 12/23 в рекомбинации V (D) J». Клетка. 85 (1): 107–13. Дои:10.1016 / s0092-8674 (00) 81086-7. PMID 8620529.
- ^ Привет, Кевин; Геллерт, Мартин (1998). «Сборка парного сигнального комплекса 12/23: критическая контрольная точка в рекомбинации V (D) J». Молекулярная клетка. 1 (7): 1011–1019. Дои:10.1016 / с1097-2765 (00) 80101-х. PMID 9651584.
- ^ Шац, Дэвид; Суонсон, Патрик (2011). "V (D) J Рекомбинация: механизмы инициации". Ежегодный обзор генетики. 45: 167–202. Дои:10.1146 / annurev-genet-110410-132552. PMID 21854230.
- ^ Шац, Дэвид; Цзи, Яньхун (2011). «Центры рекомбинации и оркестровка рекомбинации V (D) J». Nature Reviews Иммунология. 11 (4): 251–263. Дои:10.1038 / nri2941. PMID 21394103.
- ^ Карри, Джон; Гейер, Джейми; Шлиссель, Марк (2005). «Однонитевые рекомбинации сигнальных последовательностей in vivo: доказательства для модели захвата синапсиса». Иммунология природы. 6 (12): 1272–1279. Дои:10.1038 / ni1270. PMID 16286921.
- ^ Агравал, Алка; Шац, Дэвид (1997). «RAG1 и RAG2 образуют стабильный синаптический комплекс после расщепления с ДНК, содержащей сигнальные концы в рекомбинации V (D) J». Клетка. 89 (1): 43–53. Дои:10.1016 / s0092-8674 (00) 80181-6. PMID 9094713.
- ^ Фугманн, Себастьян; Ли, А.И.фред; Шокетт, Пенни; Вилли, Изабель; Шац, Дэвид (2000). «Белки RAG и рекомбинация V (D) J: комплексы, концы и транспозиция». Ежегодный обзор иммунологии. 18: 495–527. Дои:10.1146 / annurev.immunol.18.1.495. PMID 10837067.
- ^ Льюис, Сюзанна (1994). Механизм соединения V (D) J: уроки молекулярного, иммунологического и сравнительного анализов. Достижения в иммунологии. 56. С. 27–150. Дои:10.1016 / с0065-2776 (08) 60450-2. ISBN 9780120224562. PMID 8073949.
- ^ Хельминк, Бет; Слекман, Барри (2012). «Ответ и восстановление двухцепочечных разрывов ДНК, опосредованных RAG». Ежегодный обзор иммунологии. 30: 175–202. Дои:10.1146 / аннурьев-иммунол-030409-101320. ЧВК 4038028. PMID 22224778.
- ^ Ма, Юньмэй; Шварц, Клаус; Либер, Майкл (2005). «Artemis: ДНК-PKcs эндонуклеаза расщепляет петли, створки и разрывы ДНК». Ремонт ДНК. 4 (7): 845–851. Дои:10.1016 / j.dnarep.2005.04.013. PMID 15936993.
- ^ Лу, Хайхуэй; Шварц, Клаус; Либер, Майкл (2007). "Степень, в которой открытие шпильки Artemis: комплекс ДНК-PKcs может способствовать разнообразию соединений в рекомбинации V (D) J". Исследования нуклеиновых кислот. 35 (20): 6917–6923. Дои:10.1093 / нар / гкм823. ЧВК 2175297. PMID 17932067.
- ^ Гаусс, Джордж; Либер, Майкл (1996). "Механистические ограничения разнообразия в рекомбинации человеческого V (D) J". Молекулярная и клеточная биология. 16 (1): 258–269. Дои:10.1128 / MCB.16.1.258. ЧВК 230999. PMID 8524303.
- ^ Бенедикт, Синди; Гилфиллан, Сьюзен; Тайский, то-ха; Кирни, Джон (2000). «Терминальная дезоксинуклеотидилтрансфераза и развитие репертуара». Иммунологические обзоры. 175: 150–157. Дои:10.1111 / j.1600-065x.2000.imr017518.x. PMID 10933600.
- ^ ван Гент, округ Колумбия; ван дер Бург, М. (10 декабря 2007 г.). «Негомологичное соединение концов, непростое дело». Онкоген. 26 (56): 7731–40. Дои:10.1038 / sj.onc.1210871. PMID 18066085.
дальнейшее чтение
- Хартвелл Л. Х., Худ Л., Голдберг М. Л., Рейнольдс А. Э., Сильвер Л. М., Верес Р. К. (2000). Глава 24, Эволюция на молекулярном уровне. В: Генетика.. Нью-Йорк: Макгроу-Хилл. С. 805–807. ISBN 978-0-07-299587-9.
- V (D) J Рекомбинация. Серия: Успехи экспериментальной медицины и биологии, Vol. 650 Феррье, Пьер (ред.) Landes Bioscience 2009, XII, 199 с. ISBN 978-1-4419-0295-5