Немегтомайя - Nemegtomaia
| Немегтомайя | |
|---|---|
 | |
| Реконструированный скелет, Геологический музей Хэнань | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Oviraptoridae |
| Подсемейство: | †Heyuanninae |
| Род: | †Немегтомайя Люй и другие., 2005 |
| Разновидность | |
| |
| Синонимы | |
| |
Немегтомайя это род из овирапторид динозавр из того, что сейчас Монголия который жил в Поздний мел Период, около 70 миллион лет назад. Первый экземпляр был найден в 1996 году и стал основой нового рода и вида. Н. Барсбольди в 2004 г. Первоначальное название рода было Немегтия, но это было изменено на Немегтомайя в 2005 году, так как прежнее название было озабоченный. Первая часть общего названия относится к Бассейн Немегт, где было найдено животное, а вторая часть означает «хорошая мать» в связи с тем фактом, что овирапториды, как известно, имеют задумчивый их яйца. В конкретное имя чтит палеонтолога Ринчен Барсболд. Еще два экземпляра были найдены в 2007 году, один из которых был найден на вершине гнезда с яйцами, но динозавр получил название своего рода до того, как его обнаружили связанным с яйцами.
Немегтомайя По оценкам, его длина составляла около 2 м (7 футов), а вес - 40 кг (85 фунтов). Как овирапторозавр, это было бы оперено. У него был глубокий, узкий и короткий череп с изогнутым гребнем. Он был беззубым, имел короткую морду с попугай -подобный клюв и пара зубчатых выступов на его нёбо. У него было три пальца; первый был самым большим и нес сильную лапу. Немегтомайя классифицируется как член подсемейства овирапторид Heyuanninae, и является единственным известным членом этой группы с черепной гребень. Хотя Немегтомайя был использован, чтобы предположить, что овирапторозавры мы нелетающие птицы, то клады обычно считается группой не-птичий динозавры.
Гнездование Немегтомайя Образец был помещен поверх того, что, вероятно, было кольцом яиц, скрестив руки на них. Ни одно из яиц не готово, но, по оценкам, их было 5 до 6 см (2 до 2,3 в) шириной и 14 до 16 см (5 до 6 в) долго в целости и сохранности. Образец был найден в стратиграфический область, которая указывает Немегтомайя предпочитает гнездование возле ручьев, которые обеспечивают мягкий песчаный субстрат и пищу. Немегтомайя мог защитить свои яйца, прикрыв их перьями хвоста и крыльев. На скелете гнездящейся особи есть повреждения, свидетельствующие о том, что он был очищенный к кожные жуки. Диета овирапторид сомнительна, но их черепа больше всего похожи на черепа других известных или предположительно известных животных. травоядный. Немегтомайя известно из Nemegt и Образования Баруунгойот, которые, как считается, представляют влажный и засушливый среды, сосуществующие в одной области.
История открытия

В 1996 году японский палеонтолог Ёсицугу Кобаяси (в составе команды «Монгольский горный международный проект динозавров») обнаружил неполный скелет животного. овирапторид динозавр в Формация Немегт из пустыня Гоби на юго-западе Монголия. Образец (MPC-D 100/2112 в Монгольском палеонтологическом центре, ранее PC и GIN 100/2112), состоит из почти полного черепа и частичного скелета, включая шейный, дорсальный, крестцовый и хвостовой позвонки, левую лопатку, нижние концы обеих плечевых костей, правую лучевую кость, обе подвздошные кости, верхние концы обеих лобковые кости, и седалищные кости, и верхний конец бедренной кости. Экземпляр был описан как новый образец род Ingenia (именуемый Ingenia sp .; неопределенных видов) китайским палеонтологом Люй Цзюньчан и его коллеги в 2002 г., и использовались для подчеркивания сходства между овирапторозавры и птицы.[1][2][3]

В 2004 году Люй и его коллеги определили, что скелет принадлежит к новому, отличному от других таксону, и сделали его образец голотипа из Немегтия барсбольди. Название рода относится к Бассейн Немегт, а конкретное имя чтит монгольского палеонтолога Ринчен Барсболд, руководитель группы, нашедшей образец.[1] В 2005 году описатели обнаружили (после уведомления биолога), что имя Немегтия уже использовался для пресноводных креветки (Ostracoda) из той же формации в 1978 г., и поэтому озабоченный. Вместо этого они предложили новое название рода Немегтомайя ("Майя«означает« хорошая мать »по-гречески, а полное имя означает« хорошая мать Немегтов »), ссылаясь на недавнее открытие, что овирапториды задумчивый яйца вместо того, чтобы украсть их, хотя никаких следов гнезда или яиц, связанных с Немегтомайя сам.[4][5] Первый известный представитель семейства овирапторид был обнаружен с гнездом яиц, первоначально считавшимся принадлежащим цератопсий род Протоцератопс, и поэтому был назван Овираптор в 1924 г .; это имя означает «ловец яиц». В 1990-х годах было обнаружено больше экземпляров овирапторид, связанных с гнездами и яйцами, в которых были обнаружены эмбрионы овирапторид, тем самым доказав, что яйца принадлежали самим овирапторидам.[6][7][8] Ingenia был переименован так же как Ajancingenia в 2013 г., поскольку прежнее название рода заняло аскарида (Нематоды).[9]

В 2007 году два новых образца Немегтомайя были обнаружены экспедицией «Динозавры Гоби» и описаны итальянским палеонтологом Федерико Фанти и его коллегами в 2013 году. Первый экземпляр, MPC-D 107/15, был найден Фанти (по прозвищу «Мэри») в Формация Баруунгойот, и состоит из гнезда с предполагаемым родителем наверху. Как четвертый род овирапторид, обнаруженный на вершине гнезда (после Овираптор, Читипати, и ср. Махеразавр ), Немегтомайя поэтому получил название рода, относящееся к этой особенности, прежде чем она была обнаружена как связанная с яйцами. Образец был выкопан с отвесной скалы в «сложных условиях», включая проливной дождь и обрушение. песчаник блоки.[2][5]
Гнездовой скелет сохраняет части черепа, обе лопатки, левую руку и кисть, правую плечевую кость, лобковые кости, седалищные кости, бедра, голени, малоберцовые кости и нижние части обеих стоп. Этот экземпляр был найден менее чем в 500 м (1640 футов) от голотипа и имел тот же размер; это было назначено Немегтомайя из-за его похожего анатомические особенности и географическая близость. Он был собран в единый блок, чтобы сохранить пространственное соотношение костей и яиц. Второй экземпляр, МПК-Д 107/16, был обнаружен американским палеонтологом Николасом Р. Лонгричем в формации Немегт и состоит из рук, части левой локтевой кости и лучевой кости, ребер, части таза и обеих бедер. Этот образец был на 35% меньше других и был отнесен к Немегтомайя из-за того, что его руки имеют те же характеристики, что и у образца MPC-D 107/15. Возможно, что руки могли принадлежать другому человеку, поскольку они не были обнаружены сочлененными с остальной частью скелета (известны другие овирапториды из карьеров с несколькими скелетами), но это не может быть подтверждено.[2]
Описание
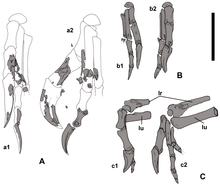
Немегтомайя По оценкам, их длина составляла около 2 м (7 футов), а вес - 40 кг (85 фунтов) - размер, экстраполированный с более известных родственников. Как овирапторозавр, он был бы пернатым. Нервные отростки шеи (шейный ) позвонки были короткими, а нервные дуги имел х-образный вид. Средние три из этих позвонков были самыми большими. В лопатка (лопатка), по всей видимости, имела общую длину 185 мм (7 дюймов). В плечевая кость (кость плеча) имела ямка (депрессия) в положении, аналогичном положению современных птиц, но нетипичному для овирапторозавров, и, по-видимому, его длина составляла 152 мм (6 дюймов). В радиус Нижняя часть руки была прямой, овальной в поперечном сечении и могла иметь длину 144 мм (5 дюймов). Первый палец был относительно большим и имел сильный ногтевой (кость копыта) и была массивнее двух других пальцев. Второй палец был немного длиннее первого, а третий был самым маленьким. Верхнее поле подвздошная кость таза была прямой, и хотя обе подвздошные кости были близко друг к другу, они не срослись. Лобковый ствол повернут назад. В бедренная кость (бедренная кость) оценивается в длину 286 мм (11 дюймов), а большеберцовая кость голени длиной 317 мм (12 дюймов).[1][2]

Череп Немегтомайя был глубоким, узким и коротким (по сравнению с остальной частью тела) и достигал 179 мм (7 дюймов) в длину. У него был хорошо развитый гребень, образованный носовой и предчелюстные кости (в основном последнее) морды. Почти вертикальный передний край гребня голотипа составлял почти 90 °. угол градуса с верхним краем черепа. По сравнению с другими овирапторидами, носовые отростки (выступы) предчелюстных костей были едва видны при взгляде сверху (где они соединялись с носовыми костями в самых высоких точках гребня). Гребень вытянут назад и вниз, образуя в высшей точке округлую арку. Диаметр орбита (отверстие для глаза) было 52 мм (2 дюйма); глаза казались большими из-за короткого черепа. Анторбитальная полость перед глазом состояла из двух фенестры (проемы); большой анторбитальное окно сзади и небольшое верхнечелюстное отверстие спереди. Немегтомайя отличался от других овирапторид тем, что лобная кость по средней линии черепа составляла около 25% длины теменная кость спереди назад. В ноздри (внешние ноздри) были относительно небольшими и располагались высоко на черепе.[1][2]

Челюсти Немегтомайя были беззубыми и, как и другие динозавры-овирапторы, имели короткую морду с глубоким, крепким и несколько попугай -подобный клюв. Было тяжело нёбо образованы предчелюстными костями, сошники, и максиллы, как и другие овирапториды. Небо сильно вогнутое (выступающее вниз), с расщелиной в центральной части. Как и у других овирапторид, он имел пару зубчатых выступов на небе, направленных вниз (особенность, которую назвали «псевдозубами»). Немегтомайя был маленький отверстие (отверстия) по бокам шов (сустав) между предчелюстными частями в передней части рыла, которые могли быть питательными отверстиями (и которые указывают на наличие ороговевший законопроект). Нижняя челюсть была короткой и глубокой, с выпуклой нижней поверхностью и достигала 153 мм (6 дюймов) в длину. В зубная кость нижней челюсти достигала 50 мм (2 дюйма) в самой высокой точке. В нижнечелюстной симфиз (где соединяются две половины нижней челюсти) был коротким, глубоким и очень пневматический (с воздушными пространствами). Нижнечелюстное отверстие большого размера, располагалось в передней части нижней челюсти.[1][2][10][3] Как и у большинства других овирапторид, передняя часть нижней челюсти повернута вниз.[11]
Хотя Немегтомайя не обладает ни одной особенностью, отличающей его от других овирапторид (аутапоморфии ), сочетание гребня, увеличенного указательного пальца и большого количества крестцовые позвонки (восемь), уникальна для этого таксон.[12]
Классификация
В 2004 г. филогенетический анализ, Лю и его коллеги классифицировали Немегтомайя как полученный (или "продвинутый") овирапторозавр и обнаружил, что он наиболее близок к роду Читипати.[1] В 2010 году Лонгрич и его коллеги определили, что Немегтомайя принадлежал к семье Oviraptoridae, как часть подсемейства Ingeniinae, что делает его единственным членом последней группы с выдающимся гребнем. Все члены другого признанного подсемейства, Oviraptorinae, обладают гребнями. Представители этого подсемейства отличались меньшими размерами, короткими и крепкими передними конечностями со слабо изогнутыми когтями, количеством позвонков в синсакрум, а также некоторые особенности стоп и таза. Лонгрич и его коллеги предположили, что наличие гребня на Немегтомайя делает возможным то, что эта особенность развивалась или исчезала несколько раз у овирапторид, или что животное могло не быть изобретателем.[2][13] В 2010 году американский палеонтолог Грегори С. Пол предположили, что бесхребетные овирапториды были либо молодыми особями, либо самками других видов с гребешками, и что поэтому количество родов в группе было преувеличено. Он перечислил Немегтомайя в качестве "Читипати (=Немегтомайя) Барсбольди, считая его очень похожим на этот род, но в 2016 году он вместо этого перечислил его как "Конхораптор (=Немегтомайя) Барсбольди".[10][14]
В 2012 году Фанти и его коллеги также обнаружили Немегтомайя быть частью Ingeniinae как производный член, ближайший к Heyuannia из-за пропорций рук двух новых экземпляров (относительно короткие с крепким указательным пальцем). Они заявили, что, хотя наличие гребней обычно связано с овирапторинами, а не с гениинами, эта особенность может быть коррелирована с размером и зрелостью. Они указали, что носовая и лобная кости ingeniine Конхораптор были пневматическими и потенциально могли вырасти в гребень по мере взросления животного, хотя все известные скелеты этого рода имеют одинаковый небольшой размер (и один экземпляр, по-видимому, полностью вырос).[2] Название подсемейства Ingeniinae было заменено названием Heyuanninae (с тех пор Ingenia был озабочен).[15] В кладограмма ниже показано размещение Немегтомайя внутри Oviraptoridae, по данным Fanti et al. др., 2012:[2]

| Oviraptoridae |
| |||||||||||||||||||||||||||||||||||||||||||||
Эволюция

В клады Овирапторозаврия обычно рассматривается как группа не-птичий (или не птица) теропод динозавров, и их сходство с птицами (Aves) отмечалось часто. Ископаемые останки овирапторозавров в семье Caenagnathidae исторически путали с птицами, и некоторые исследователи зашли так далеко, что считают овирапторозавров в целом более близкими к птицам, чем к другим нептичьим динозаврам. В 2002 году Люй и его коллеги использовали тогда еще неназванный Немегтомайя чтобы показать сходство между птицами и овирапторозаврами, и обнаружил, что последняя группа ближе к птицам, чем к птицеподобным динозаврам, таким как дромеозавры. Поэтому они пришли к выводу, что овирапторозавры были нелетающие птицы а не нептичьих динозавров, и отметил, что границу между птицами и динозаврами становится все труднее очертить.[3] Вместо этого другие исследователи обнаружили дромеозавров и троодонтиды быть наиболее близкими к птицам, вместе составляя группу Паравес; овирапторозавры, теризинозавры, и альваресзавры находятся за пределами этой группы. Более широкая группа, включающая овирапторозавров и паравес, называется Pennaraptora, и эта группа определяется наличием Pennaceous перья (перья со стеблем).[16]
Овирапторозавры известны из Азии (откуда они, возможно, произошли) и Северной Америки, и в основном известны по месторождениям, которые датируются Кампанский -Маастрихтский возраст Меловой период. Группа включает в себя мелких и крупных членов, и для них характерны короткие черепа и клювы, удлиненные пальцы и короткие хвосты. Базальный (или «примитивные») члены имели зубы, которые исчезали у производных членов группы (внутри суперсемейства). Caenagnathoidea, в том числе Oviraptoridae). Они были хотя бы частично травоядный, и гнездились в позе птицы. Хотя считается, что все они были пернатыми, похоже, они были нелетающими. Черепной гребни, кажется, имеют развивались конвергентно в разных родословная внутри группы.[16] Семейство Oviraptoridae (к которому Немегтомайя принадлежит) состояла, как правило, из небольших членов и известна исключительно из Верхний мел Азии, причем большинство родов были обнаружены в пустыне Гоби в Монголии и Китае.[13] Включая Немегтомайя, по крайней мере, девять родов овирапторид были обнаружены в относительно небольшом географическом районе в пустыне Гоби.[12][17]
Палеобиология
Размножение

В Немегтомайя образец MPC-D 107/15 был обнаружен связанным с гнездом с яйцами; его ноги были помещены в центр того, что, вероятно, было кольцом яиц, а руки были скрещены на верхушках яиц с каждой стороны тела, положение, подобное тому, которое можно увидеть на других окаменелостях высиживающих овирапторид. Собранная часть гнезда имеет ширину около 90 см (35 дюймов) и длину 100 см (30 дюймов); скелет занимает верхние 25 см (10 дюймов) блока, тогда как оставшиеся 20 см (8 дюймов) занимают разбитые яйца и скорлупа. Признаков наличия растительного материала в гнезде нет, но есть фрагменты неустановленных костей. Гнездо не сохраняет целые яйца или эмбрионы, что не позволяет определить размер, форму, количество и расположение яиц в гнезде. Вероятно, что изначально под телом было два слоя яиц, а в центре гнезда яиц не было. Большинство яиц (было идентифицировано семь отдельных яиц) и фрагменты яиц были обнаружены либо в нижнем слое гнезда, либо под черепом, шеей и конечностями особи, а кости либо лежали непосредственно на яйцах, либо находились в пределах 5 мм. (0,2 дюйма) их поверхностей. То, что скелет был расположен прямо на нем, показывает, что гнездо не было полностью засыпано песком. Хотя расположение яиц не предполагает особого расположения в гнезде, большинство других гнезд овирапторид показывают, что яйца были расположены парами до трех уровней концентрических кругов. Яйца МПК-Д 107/15, скорее всего, были перемещены во время захоронения или из-за внешних факторов, таких как сильный ветер, песчаные бури, или хищники. Это также подтверждает идею о том, что верхний слой яиц не был захоронен, поскольку было бы меньше шансов перенести полностью захороненные яйца из-за внешних факторов.[2]

Яйца овирапторид в среднем имели длину 17 см (6 дюймов), и наиболее полные яйца были обнаружены с помощью MPC-D. 107/15 считаются 5 до 6 см (2 до 2,3 дюймов) в ширину и от 14 до 16 см (5 до 6 в) долго в целости и сохранности. Яйца почти идентичны тем, которые ранее были обнаружены в Монголии, и поэтому были отнесены к семья (семейство яичных таксонов) Elongatoolithidae. Яичная скорлупа относительно тонкая, от 1 и 1,2 мм (0,03 и 0,04 дюйм), а их внешняя поверхность покрыта гребнями и узлами, которые возвышаются примерно на 0,3 мм (0,01 дюйма) над оболочкой. В микроструктура яичной скорлупы не могли быть должным образом изучены, поскольку кальцит был сильно изменен и перекристаллизованный.[2]
Гнездящийся экземпляр найден в стратиграфический площадь, указывающая на то, что овирапториды предпочитали гнездиться возле ручьев, которые обеспечивали мягкий песчаный субстрат и пищу в среде, которая в противном случае ксерический (получение небольшого количества влаги). Многие овирапториды были обнаружены в положении насиживания, что указывает на то, что они могли насиживать наседки в течение относительно длительных периодов времени, как и современные птицы, такие как страус, эму, и черногрудый канюк, которые размножаются более 40 дней при ограниченном питании. Гнездование в пустыне может быть вредным для взрослых особей, которые остаются в гнезде большую часть дня, а также для яиц и птенцов из-за: перегрев. Таким образом, выбор места для гнездования мог быть механизмом успешной инкубации в условиях сильной жары. Также было высказано предположение, что эволюция хвостовых перьев у овирапторозавров была адаптацией для затенения и защиты яиц в их гнездах. То, что второй палец овирапторид геюанина был уменьшен в размере по сравнению с крепким первым пальцем, можно объяснить изменением функции; это может быть связано с наличием длительного перья крыльев которые были прикреплены ко второму пальцу. Эти перья крыльев, вероятно, использовались для защиты яиц во время гнездования. Когда второй палец начал функционировать как опора для пера, его способность хватать уменьшилась, и эту функцию взял на себя первый палец, который, следовательно, стал более надежным. Третий палец также был уменьшен в размерах, вероятно, потому, что он был расположен за перьями крыльев таким образом, что он не мог быть эффективным для захвата.[2]
В 2018 году тайваньский палеонтолог Цзы-Руэй Ян и его коллеги определили кутикула слои на яичной скорлупе динозавров манирапторанов, в том числе овирапторид. У современных птиц такие слои (состоящие в основном из липиды и гидроксиапатит ) служат для защиты яиц от обезвоживания и вторжения микробы. Исследователи предположили, что яйца, покрытые кутикулой, были бы признаком, адаптированным для повышения их репродуктивного успеха в изменчивой среде, где Немегтомайя и другие овирапториды гнездятся.[18]

Различные исследования показали, что несколько особей будут собирать яйца в одном гнезде и располагать их так, чтобы их мог защитить один особь, возможно, самец.[2] В 2010 году американский палеонтолог Дэвид Дж. Варриккио и его коллеги обнаружили, что относительно большой схватить -размер овирапторид и троодонтид наиболее близок к размерам современных архозавры (птицы и крокодилы, ближайших родственников динозавров), которые практикуют полигамный вязка и обширный самец родительская забота (как показано на палеогнаты такие как страусы и эму). Этот репродуктивная система предшествует происхождение птиц и, следовательно, было бы наследственным условием для современных птиц, а забота о двух родителях (в которой участвуют оба родителя) является более поздним развитием.[19] Известно, что многие овирапторозавры имели пигостили на конце их хвостов, что предполагает наличие перьев-вееров; американский палеонтолог В. Скотт Персонс и его коллеги предположили в 2013 году, что они могли быть использованы для внутривидовая коммуникация Такие как ритуалы ухаживания.[20]
Диета и кормление

Диета овирапторидов с тех пор интерпретировалась по-разному. Овираптор ошибочно считали, что он был хищником яиц. Было высказано предположение, что овирапторозавры в целом были травоядными, что подтверждается гастролиты (камни в желудке) обнаружены в Каудиптерикс, а грани износа на зубах Инцизивозавр. В 2010 году Лонгрич и его коллеги обнаружили, что челюсти овирапторий имеют черты, похожие на те, что наблюдаются у травоядных. четвероногие (четвероногие животные), особенно дицинодонты, вымершая группа синапсид корень-млекопитающие. Овирапториды и дицинодонты имеют такие общие черты, как короткие, глубокие и беззубые нижние челюсти; удлиненные зубные симфизы; удлиненные нижнечелюстные отверстия; и выступающая вниз полоска во рту. Современные животные с челюстями, напоминающими челюсти овирапторид, включают попугаев и черепах; у последней группы также есть зубчатые выступы на предчелюстных костях. Лонгрич и его коллеги пришли к выводу, что из-за сходства между овирапторидами и травоядными животными, основная часть их рациона, скорее всего, была сформирована из растительного вещества. Овирапториды часто встречаются в формациях, из которых они известны, подобно тому, как это наблюдается у динозавров, которые, как известно, являются травоядными; эти животные были более многочисленны, чем хищные динозавры, поскольку больше энергии было доступно при их низком уровне. трофический уровень в пищевая цепочка. Челюсти овирапторид могли быть специализированы для обработки пищи, например ксерофитный растительность (адаптированная для окружающей среды с небольшим количеством воды), которая могла бы расти в их среде, но это невозможно продемонстрировать, так как в то время о флоре этого района было мало что известно.[13] Исследование, проведенное Лю и его коллегами в 2013 году, показало, что овирапториды, по-видимому, сохраняли пропорции задних конечностей на протяжении всего времени. онтогенез (рост), что также характерно для травоядных животных.[21] В 2017 году канадский палеонтолог Грегори Фанстон и его коллеги предположили, что похожие на попугаев челюсти овирапторид могут указывать на плодоядный диета, включающая орехи и семена.[12]
В 1977 г. Барсболд предположил, что овирапториды питался моллюсками, но Лонгрич и его коллеги отвергли идею, что они практиковали дробящий снаряд в целом, поскольку у таких животных, как правило, зубы с широкой дробящейся поверхностью. Вместо этого форма зубных костей в нижней челюсти овирапторид предполагает, что у них был клюв с острыми краями, который использовался для стрижки жесткой пищи, а не для раскалывания твердых пищевых продуктов, таких как двустворчатые моллюски или яйца. Симфизная полка в передней части зубной кости, возможно, давала некоторую возможность раздавливания, но, поскольку это была относительно небольшая область, вероятно, это не была основная функция челюстей. Тот факт, что большинство овирапторид было обнаружено в отложения которые интерпретируются как ксерические и засушливые или полузасушливые среды, также приводят доводы против того, что они были специализированными едоками моллюсков и яиц, поскольку маловероятно, что в таких условиях было бы достаточно этих предметов, чтобы поддержать их.[13]
Лонгрич и его коллеги отметили, что крепкие передние конечности и увеличение одного пальца у хейуаниновых овирапторид аналогичны тем, которые наблюдаются у современных животных, поедающих муравьев и термитов, таких как муравьеды и панголины, но морфолология челюстей хейуанина не подтверждает, что они насекомоядный. Исследователи обнаружили, что функция передних конечностей хейуанина была неясной, но предположили, что они могли использоваться для царапин, разрывов или рытья, но не для захвата добычи.[13]
В 2004 году Люй и его коллеги предложили, чтобы артикуляция между квадратный и четырехъядерные кости в черепе Немегтомайя предположил, что эти кости были подвижные по отношению друг к другу, что повлияло бы на работу челюстей. В 2015 году бельгийский палеонтолог Кристоф Хендрикс и его коллеги сочли маловероятным, что Немегтомайя и другие овирапторды имели птичий кинезис в черепах из-за неподвижности квадратной кости.[22]
Палеоэкология

Немегтомайя известен по свитам Немегт и Баруунгойот, которые датируются верхним кампаном - нижним маастрихтом позднего мелового периода, около 70 миллион лет назад. Хотя этот таксон известен только из местонахождения Немегт, неопознанные остатки овирапторид из других местонахождений могут принадлежать ему. Немегт массив имеет множество каньонов или ущелий глубиной до 45 метров (148 футов), которые имеют одни из лучших обнажений среди этих образований. Рок фации формации Немегт, как полагают, представляют собой влажные, речной (связанные с реками и ручьями) окружающей среды, тогда как те из формации Баруунгойот, как полагают, представляют собой засушливую или полузасушливую среду с эолийский (пострадавшие от ветра) кровати. Исторически считалось, что эти два образования с их разнообразными окаменелостями представляют собой последовательные периоды времени с разной средой обитания, но в 2009 году канадский палеонтолог Дэвид А. Эберт и его коллеги обнаружили, что переходы между ними частично совпадают. Две формации «соприкасаются» в стратиграфическом интервале толщиной около 25 м (82 фута), что позволяет предположить, что речная и эоловая среды сосуществовали, когда область осаждалась.[2][12][23][24]

Окружающая среда формации Немегт сравнивалась с Дельта Окаванго современного Ботсвана.[25] Среда обитания в реках Немегт и вокруг них служила домом для самых разных организмов. Водные животные включают моллюсков, рыб, черепах и крокодиломорф Шамозух. Ископаемые останки млекопитающих, таких как многотуберкулезные были найдены, и такие птицы, как Гурилина, Юдинорнис, и Тевиорнис известны. Среди травоядных динозавров свиты Немегт анкилозавриды Такие как Тархия, то пахицефалозавр Prenocephale, гадрозавриды Такие как Зауролоф и Барсболдия, и зауроподы Такие как Немегтозавр, и Опистоцеликаудия. Другие тероподы включают тираннозавроиды Такие как Тарбозавр, Алиорамус, и Багараатан, троодонтиды Такие как Бороговия, Tochisaurus, и Saurornithoides, теризинозавры, такие как Теризинозавр, и орнитомимозавры Такие как Дейнохейрус, Ансеримим, и Галлимим.[26][27][28]
Другие роды овирапторозавров, известные из формации Немегт, включают базальные Авимимус, овирапториды Ринчения, Номингия, Конхораптор и Ajancingenia, а ценагнатид Эльмизавр. Несмотря на большое количество таксонов овирапторид в этих формациях (Nemegt имеет наибольшее известное разнообразие из всех), ни один из них не был тесно связан. Формация Немегт уникальна тем, что в ней присутствуют овирапторозавры как овирапторид, так и ценагнатид, а в 1993 г. канадский палеонтолог Филип Дж. Карри и коллеги предположили, что это разнообразие объясняется тем, что две группы предпочитают разную среду обитания в этом районе. В 2016 году японский палеонтолог Таканобу Цуйхидзи и его коллеги предположили, что овирапториды, возможно, предпочитали более сухую среду, в то время как ценагнатиды предпочитали речную среду, в зависимости от типа формаций, в которых они были обнаружены.[2][29] Фунстон и его коллеги предположили, что овирапториды были обнаружены как у ксериков, так и у мезические среды (но в первых их было больше), тогда как другие группы овирапторозавров избегали ксерических сред, и что сосуществование семейств можно объяснить тем, что перегородка ниши в диете. Окружающая среда формации Немегт могла действовать как оазис и тем самым привлекли овирапторид.[12]
Тафономия

Образец гнездования MPC-D 107/15 предоставил много информации о тафономические процессы (изменения во время распада и фоссилизации) в формации Баруунгойот. Образец сохранился в фациях, которые предположительно образовались в результате песчаной бури или дюна -сдвиг. Похоже, что после смерти его не перевезли, но тело, похоже, слегка сместилось вправо, что указывает на то, что осадок, в котором оно было отложено, пришел к нему с левой стороны. Шея загнута влево, левая рука отведена назад, а ноги сложены в положение приседания. Позвоночный столб, шея и бедра ухудшились во время захоронения, и большая часть повреждений скелета, как полагают, была вызвана деятельностью беспозвоночные.[2]
Просверленные отверстия в костях, норах и переработанных отложениях (возможно, вызванные строительством кукольные камеры ) в образце указывают, что это было очищенный на колонии кожные жуки (Dermestidae) и, возможно, другие насекомые-падальщики. В суставах скелета много следов питания, и почти все поверхности, на которых сочленялись кости, стерлись. В гнезде также есть ходы под шеей и черепом, и на участках с такими следами остатков яиц не обнаружено. Современные кожные жуки в основном питаются мышечной тканью, но не влажными материалами, и их деятельность предотвращается быстрым захоронением. Поэтому считается, что образец MPC-D 107/15 поначалу был захоронен лишь частично, а его верхняя часть была обнажена достаточно, чтобы могла развиться колония кожных жуков. Некоторое повреждение скелета (особенно позвоночника) также могло быть вызвано поеданием мелких млекопитающих.[2]
Смотрите также
Рекомендации
- ^ а б c d е ж Lü, J .; Tomida, Y .; Azuma, Y .; Dong, Z .; Ли, Ю.-Н. (2004). «Новый динозавр-овирапторид (Dinosauria: Oviraptorosauria) из формации Немегт на юго-западе Монголии» (PDF). Бюллетень Национального музея науки, Токио, серия C. 30: 95–130.
- ^ а б c d е ж грамм час я j k л м п о п q Fanti, F .; Карри, П. Дж .; Бадамгарав, Д .; Лалуэса-Фокс, К. (2012). "Новые образцы Немегтомайя из свит Баруунгойот и Немегт (поздний мел) Монголии ». PLOS ONE. 7 (2): e31330. Bibcode:2012PLoSO ... 731330F. Дои:10.1371 / journal.pone.0031330. ЧВК 3275628. PMID 22347465.
- ^ а б c Лю, Дж., Донг, З., Адзума, Ю., Барсболд, Р., Томида, Ю. (2002). «Овирапторозавры в сравнении с птицами». In Zhou, Z. & Zhang, F. (ред.). Труды 5-го симпозиума Общества палеонтологии и эволюции птиц. Пекин: Science Press. С. 175–189.
- ^ Lü, J .; Tomida, Y .; Azuma, Y .; Dong, Z .; Lee, Y.-N .; и другие. (2005). "Немегтомайя ген. nov., заменяющее имя динозавра овирапторозавра Немегтия Lü et al. 2004 г., озабоченное имя ». Бюллетень Национального музея науки, Токио, серия C. 31: 51.
- ^ а б Беседка, В. (2012). «Дневники пустыни Гоби: Немегтомайя Версия". pseudoplocephalus.blogspot.com. Получено 10 января 2017.
- ^ Norell, M.A .; Clark, J.M .; Chiappe, L.M .; Дашзевег, Д. (1995). «Гнездовой динозавр». Природа. 378 (6559): 774–776. Bibcode:1995Натура 378..774Н. Дои:10.1038 / 378774a0. S2CID 4245228.
- ^ Донг, З.-М .; Карри, П.Дж. (1996). «Об обнаружении скелета овирапторид в гнезде из яиц в Баян Мандаху, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле. 33 (4): 631–636. Bibcode:1996CaJES..33..631D. Дои:10.1139 / e96-046.
- ^ Clark, J.M .; Норелл, М .; Чиаппе, Л. М. (1999). «Скелет овирапторид из позднего мела Ухаа Толгод, Монголия, сохранившийся в птичьем положении для размышлений над гнездом овирапторид». Американский музей Novitates (3265): 1–35. HDL:2246/3102.
- ^ Пасха, J. (2013). "Новое название овирапторидного динозавра. "Ингения" яншини (Барсболд, 1981; озабочен Герлахом, 1957) ". Zootaxa. 3737 (2): 184–190. Дои:10.11646 / zootaxa.3737.2.6. PMID 25112747.
- ^ а б Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. стр.152, 154. ISBN 978-0-691-13720-9.
- ^ Lü, J .; Yi, L .; Чжун, H .; Wei, X .; Додсон, П. (2013). «Новый овирапторозавр (Dinosauria: Oviraptorosauria) из позднего мела Южного Китая и его палеоэкологические последствия». PLoS ONE. 8 (11): e80557. Bibcode:2013PLoSO ... 880557L. Дои:10.1371 / journal.pone.0080557. ЧВК 3842309. PMID 24312233.
- ^ а б c d е Funston, G.F .; Mendonca, S.E .; Currie, P.J .; Барсболд Р. (2017). «Анатомия, разнообразие и экология овирапторозавров в бассейне реки Немегт». Палеогеография, палеоклиматология, палеоэкология. 494: 101–120. Bibcode:2018ППП ... 494..101Ф. Дои:10.1016 / j.palaeo.2017.10.023.
- ^ а б c d е Longrich, N.R .; Currie, P.J .; Чжи-Мин, Д. (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандаху, Внутренняя Монголия». Палеонтология. 53 (5): 945–960. Дои:10.1111 / j.1475-4983.2010.00968.x.
- ^ Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. п. 181. ISBN 978-0-691-16766-4.
- ^ Юн, К.-Г. (2019). «Heyuanninae clade nov., Заменяющее название подсемейства овирапторид« Ingeniinae »Barsbold, 1981». Zootaxa. 4671 (2): 295–296. Дои:10.11646 / zootaxa.4671.2.11. PMID 31716062.
- ^ а б Hendrickx, C .; Hartman, S.A .; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал PalArch по палеонтологии позвоночных. 12 (1): 1–73.
- ^ Wang, S .; Sun, C .; Салливан, К .; Сюй, X. (2013). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела на юге Китая». Zootaxa. 3640 (2): 242–57. Дои:10.11646 / zootaxa.3640.2.7. PMID 26000415.
- ^ Ян, Т.- .; Chen, Y.-H .; Wiemann, J .; Spiering, B .; Сандер, П. М. (2018). "Ископаемая кутикула яичной скорлупы проясняет экологию гнездования динозавров". PeerJ. 6: e5144. Дои:10.7717 / peerj.5144. ЧВК 6037156. PMID 30002976.
- ^ Varricchio, D.J .; Moore, J.R .; Эриксон, G.M .; Norell, M.A .; Jackson, F.D .; Борковский, J.J. (2008). "Птичий отцовский уход произошел от динозавров". Наука. 322 (5909): 1826–1828. Bibcode:2008Научный ... 322.1826V. Дои:10.1126 / science.1163245. PMID 19095938.
- ^ Персоны, С. (2013). «Формы и функции хвоста овирапторозавра». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0093.
- ^ Lü, J .; Currie, P.J .; Xu, L .; Чжан, X .; Pu, H .; Цзя, С. (2013). "Овирапторидные динозавры размером с курицу из центрального Китая и их онтогенетические последствия". Naturwissenschaften. 100 (2): 165–175. Bibcode:2013NW .... 100..165л. Дои:10.1007 / s00114-012-1007-0. PMID 23314810. S2CID 206871470.
- ^ Hendrickx, C .; Araújo, R .; Матеус, О. (2015). «Квадрат теропод I, не относящийся к птицам: стандартизованная терминология с обзором анатомии и функций». PeerJ. 3: e1245. Дои:10.7717 / peerj.1245. ЧВК 4579021. PMID 26401455.
- ^ Eberth, D. A .; Бадамгарав, Д .; Карри, П. Дж. (2009). «Переход Баруунгойот-Немегт (верхний мел) в районе типа Немегт, бассейн Немегт, Южная Центральная Монголия». 고생물 학회지. 25 (1). ISSN 1225-0929.
- ^ Эберт, Д.А. (2017). «Стратиграфия и палеоэкологическая эволюция богатой динозаврами последовательности Баруунгойот-Немегт (верхний мел), бассейн Немегт, южная Монголия». Палеогеография, палеоклиматология, палеоэкология. 494: 29–50. Bibcode:2018ППП ... 494 ... 29E. Дои:10.1016 / j.palaeo.2017.11.018.
- ^ Хольц, Т. (2014). «Палеонтология: разгадана тайна ужасных рук». Природа. 515 (7526): 203–205. Bibcode:2014Натура.515..203H. Дои:10.1038 / природа13930. PMID 25337885. S2CID 205241353.
- ^ Lucas, S.G .; Эстеп, Дж. (1998). «Биостратиграфия и биохронология позвоночных мелового периода Китая». Бюллетень Музея естественной истории и науки Нью-Мексико. 14: 1–20.
- ^ Watabe, M .; Цогтбаатар, К .; Suzuki, S .; Санэёси, М. (2010). «Геология местонахождений окаменелостей динозавров (юра и мел: мезозой) в пустыне Гоби: результаты совместной палеонтологической экспедиции HMNS-MPC». Бюллетень исследований музея естественных наук Хаясибара. 3: 11–18.
- ^ Jerzykiewicz, T .; Рассел, Д.А. (1991). «Позднемезозойская стратиграфия и позвоночные животные Гобийской впадины». Меловые исследования. 12 (4): 345–377. Дои:10.1016/0195-6671(91)90015-5.
- ^ Tsuihiji, T .; Watabe, M .; Цогтбаатар, К .; Барсболд Р. (2016). «Зубные палочки ценагнатида (Dinosauria: Theropoda) из формации Немегт в пустыне Гоби в Монголии». Меловые исследования. 63: 148–153. Дои:10.1016 / j.cretres.2016.03.007.
















