Комплекс распознавания происхождения - Origin recognition complex
| Субъединица 2 комплекса распознавания происхождения | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC2 | ||||||||
| Pfam | PF04084 | ||||||||
| ИнтерПро | IPR007220 | ||||||||
| |||||||||
| N-конец субъединицы 3 комплекса распознавания источника (ORC) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC3_N | ||||||||
| Pfam | PF07034 | ||||||||
| ИнтерПро | IPR010748 | ||||||||
| |||||||||
| Субъединица 6 комплекса распознавания происхождения (ORC6) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC6 | ||||||||
| Pfam | PF05460 | ||||||||
| ИнтерПро | IPR008721 | ||||||||
| |||||||||
В молекулярная биология, комплекс распознавания происхождения (ORC) является мульти-субъединицей ДНК связывающий комплекс (6 субъединиц), который связывает все эукариоты и археи в АТФ -зависимый способ истоки репликации. Субъединицы этого комплекса кодируются ORC1, ORC2, ORC3, ORC4, ORC5 и ORC6 гены.[1][2][3] ORC - центральный компонент для репликация эукариотической ДНК, и остается привязанным к хроматин в репликация истоки на протяжении клеточный цикл.[4]
ORC руководит Репликация ДНК на протяжении геном и требуется для его инициирования.[5][6][7] ORC, связанный в источниках репликации, служит основой для сборки пререпликационный комплекс (pre-RC), который включает Cdc6, Tah11 (a.k.a. Cdt1 ), а Mcm2 -Mcm7 сложный.[8][9][10] Предварительная сборка RC во время G1 требуется для репликации лицензирование из хромосомы до синтеза ДНК во время S фаза.[11][12][13] Регулируемый клеточный цикл фосфорилирование of Orc2, Orc6, Cdc6 и MCM циклин -зависимый протеинкиназа Cdc28 регулирует инициацию репликации ДНК, включая блокирование повторной инициации в G2 /Фаза M.[4][14][15][16]
ORC присутствует на протяжении всего клеточного цикла, связанного с источниками репликации, но активен только на поздних стадиях. митоз и рано G1.
У дрожжей ORC также играет роль в установлении сайленсинга на вязальный места Скрытый коврик слева (HML) и скрытый коврик справа (HMR).[5][6][7] ORC участвует в сборке транскрипционно тихий хроматин в HML и HMR, вербовав глушителя Sir1 белок к глушителям HML и HMR.[7][17][18]
И Orc1, и Orc5 связывают АТФ, хотя только Orc1 имеет АТФаза Мероприятия.[19] В привязка АТФ с помощью Orc1 необходим для связывания ORC с ДНК и необходим для жизнеспособности клеток.[10] Активность АТФазы Orc1 участвует в формировании пре-RC.[20][21][22] Связывание АТФ с помощью Orc5 имеет решающее значение для стабильности ORC в целом. Только орк1-5 подразделения необходимы для привязки к источнику; Orc6 необходим для поддержания предварительно сформированных RC.[23] Взаимодействие внутри ORC предполагает, что Orc2-3-6 может образовывать основной комплекс.[4]
Белки

В ORC присутствуют следующие белки:
| С. cerevisiae | С. Помбе | D. melanogaster | Позвоночные |
|---|---|---|---|
| ORC 1-6 | ORC 1-6 | ORC 1-6 | ORC 1-6 |
| Cdc6 | Cdc18 | Cdc6 | Cdc6 |
| Cdt1 / Tah11 / Sid2 | Cdt1 | DUP | Cdt1 / RLF-B |
| Mcm2 | Mcm2 / Cdc19 / Nda1 | Mcm2 | Mcm2 |
| Mcm3 | Mcm3 | Mcm3 | Mcm3 |
| Cdc54 / Mcm4 | Cdc21 | DPA | Mcm4 |
| Cdc46 / Mcm5 | Mcm5 / Nda4 | Mcm5 | Mcm5 |
| Mcm6 | Mcm6 / Mis5 | Mcm6 | Mcm6 |
| Cdc47 / Mcm7 | Mcm7 | Mcm7 | mcm7 |
Археи имеют упрощенную версию ORC, Mcm и, как следствие, комбинированный pre-RC. Вместо использования шести разных мкм белки с образованием псевдосимметричного гетерогексамера, все шесть субъединиц в MCM архей одинаковы. У них обычно есть несколько белков, гомологичных как Cdc6, так и Orc1, некоторые из которых выполняют функцию обоих. В отличие от эукариотических орков, они не всегда образуют комплекс. Фактически, они имеют расходящиеся сложные структуры, когда они формируются. Sulfolobus islandicus также использует гомолог Cdt1 для распознавания одного из его источников репликации.[25]
Автономно реплицирующиеся последовательности
Бутоновые дрожжи
Автономно реплицирующиеся последовательности (ARS), впервые обнаруженные в расцветающие дрожжи, являются неотъемлемой частью успеха ORC. Эти 100-200бп последовательности облегчают репликационную активность во время S-фазы. ARS можно разместить в любом новом месте хромосомы почкующихся дрожжей и будет способствовать репликации с этих сайтов. Высококонсервативная последовательность из 11 пар оснований (известная как Элемент) считается важным для функции происхождения почкующихся дрожжей.[24] ORC был первоначально идентифицирован по его способности связываться с элементом A ARS у почкующихся дрожжей.
Животные
Клетки животных содержат гораздо более загадочную версию ARS, без консервативные последовательности найдено на данный момент. Однако в клетках животных источники репликации собираются в связки, называемые репликон кластеры. Репликоны каждого кластера похожи по длине, но отдельные кластеры имеют репликоны разной длины. Все эти репликоны имеют сходные основные остатки, с которыми связывается ORC, которые во многих отношениях имитируют консервативный элемент A из 11 пар оснований. Все эти кластеры одновременно активируются во время S фаза.[24]
Роль в предварительной сборке RC
ORC необходим для загрузки комплексов MCM (Pre-RC ) на ДНК. Этот процесс зависит от ORC, Cdc6, и Cdt1 - с участием нескольких АТФ контролируемые рекрутинговые мероприятия. Во-первых, ORC и Cdc6 образовывать комплекс по происхождению ДНК (отмечены регионами типа ARS). Новые комплексы ORC / Cdc6 затем привлекают молекулы Cdt1 / Mcm2-7 к сайту. Как только этот массивный комплекс ORC / Cdc6 / Cdt1 / Mcm2-7 (OCCM) образуется, молекулы ORC / Cdc6 / Cdt1 работают вместе, чтобы загрузить Mcm2-7 на саму ДНК гидролиз АТФ с помощью Cdc6. Фосфорилирующая активность Cdc6 зависит как от ORC, так и от происхождение ДНК. Это ведет к Cdt1 снижение стабильности ДНК и выпадение комплекса, приводящее к Mcm2-7 загрузка в ДНК.[26][24][27][28] Структура ORC, MCM, а также промежуточного комплекса OCCM была определена.[29]
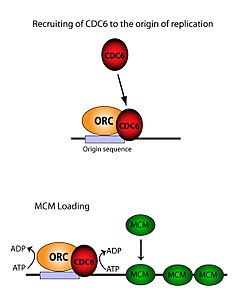

Активность привязки источника
Хотя ORC состоит из шести отдельных субъединиц, только одна из них оказалась значимой - ORC1. Исследования in vivo показали, что Lys -263 и Arg -367 - это основные остатки, отвечающие за правильную загрузку ORC. Эти молекулы представляют собой упомянутый выше ARS.[30] ORC1 взаимодействует с АТФ и эти основные остатки для связывания ORC с происхождение ДНК. Было установлено, что это происходит задолго до репликации, и что сам ORC уже связан с исходной ДНК к моменту любого Mcm2-7 происходит загрузка.[28] Когда Mcm2-7 сначала загружается, он полностью охватывает ДНК и геликаза активность подавляется. В S фаза, то Mcm2-7 комплекс взаимодействует с геликазой кофакторы Cdc45 и Джинс выделить одну нить ДНК, размотать ориджин и начать репликацию по хромосома. Чтобы иметь двунаправленную репликацию, этот процесс происходит дважды в источнике. Оба события загрузки опосредуются одним ORC посредством того же процесса, что и первый.[31]
Смотрите также
Рекомендации
- ^ Происхождение + Распознавание + Комплекс в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- ^ Датта А., Белл С.П. (1997). «Инициирование репликации ДНК в эукариотических клетках». Анну. Rev. Cell Dev. Биол. 13: 293–332. Дои:10.1146 / annurev.cellbio.13.1.293. PMID 9442876.
- ^ Чесноков И.Н. (2007). «Многофункциональность комплекса распознавания происхождения». Int. Преподобный Цитол. Международный обзор цитологии. 256: 69–109. Дои:10.1016 / S0074-7696 (07) 56003-1. ISBN 9780123737007. PMID 17241905.
- ^ а б c Мацуда К., Макисе М., Суэясу Ю., Такехара М., Асано Т., Мидзусима Т. (декабрь 2007 г.). «Дрожжевой двугибридный анализ комплекса распознавания происхождения Saccharomyces cerevisiae: взаимодействие между субъединицами и идентификация связывающих белков». FEMS дрожжи Res. 7 (8): 1263–9. Дои:10.1111 / j.1567-1364.2007.00298.x. PMID 17825065.
- ^ а б Белл С.П., Стиллман Б. (май 1992 г.). «АТФ-зависимое распознавание эукариотических источников репликации ДНК мультибелковым комплексом». Природа. 357 (6374): 128–34. Bibcode:1992Натура.357..128Б. Дои:10.1038 / 357128a0. PMID 1579162. S2CID 4346767.
- ^ а б Белл С.П., Митчелл Дж., Лебер Дж., Кобаяши Р., Стиллман Б. (ноябрь 1995 г.). «Мультидоменная структура Orc1p обнаруживает сходство с регуляторами репликации ДНК и подавления транскрипции». Клетка. 83 (4): 563–8. Дои:10.1016/0092-8674(95)90096-9. PMID 7585959.
- ^ а б c Гибсон Д.Г., Белл С.П., Апарисио О.М. (июнь 2006 г.). «Анализ точки выполнения клеточного цикла функции ORC и характеристика реакции контрольной точки на инактивацию ORC у Saccharomyces cerevisiae». Гены Клетки. 11 (6): 557–73. Дои:10.1111 / j.1365-2443.2006.00967.x. PMID 16716188.
- ^ Рао Х., Стиллман Б. (март 1995 г.). «Комплекс распознавания ориджина взаимодействует с двудольным участком связывания ДНК в репликаторах дрожжей». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 92 (6): 2224–8. Bibcode:1995PNAS ... 92.2224R. Дои:10.1073 / пнас.92.6.2224. ЧВК 42456. PMID 7892251.
- ^ Роули А., Кокер Дж. Х., Харвуд Дж., Диффли Дж. Ф. (июнь 1995 г.). «Сборка комплекса инициации в зарождающихся источниках репликации дрожжей начинается с распознавания двудольной последовательности путем ограничения количества инициатора, ORC». EMBO J. 14 (11): 2631–41. Дои:10.1002 / j.1460-2075.1995.tb07261.x. ЧВК 398377. PMID 7781615.
- ^ а б Спек К., Чен З, Ли Х, Стиллман Б. (ноябрь 2005 г.). «АТФаза-зависимое кооперативное связывание ORC и Cdc6 с исходной ДНК». Nat. Struct. Мол. Биол. 12 (11): 965–71. Дои:10.1038 / nsmb1002. ЧВК 2952294. PMID 16228006.
- ^ Келли TJ, Браун GW (2000). «Регуляция репликации хромосом». Анну. Преподобный Biochem. 69: 829–80. Дои:10.1146 / annurev.biochem.69.1.829. PMID 10966477.
- ^ Белл С.П., Датта А. (2002). «Репликация ДНК в эукариотических клетках». Анну. Преподобный Biochem. 71: 333–74. Дои:10.1146 / annurev.biochem.71.110601.135425. PMID 12045100.
- ^ Стиллман Б. (февраль 2005 г.). «Распознавание происхождения и хромосомный цикл». FEBS Lett. 579 (4): 877–84. Дои:10.1016 / j.febslet.2004.12.011. PMID 15680967. S2CID 33220937.
- ^ Вайнрайх М., Лян С., Чен Х. Х., Стиллман Б. (сентябрь 2001 г.). «Связывание циклин-зависимых киназ с ORC и Cdc6p регулирует цикл репликации хромосомы». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 98 (20): 11211–7. Дои:10.1073 / pnas.201387198. ЧВК 58709. PMID 11572976.
- ^ Nguyen VQ, Co C, Li JJ (июнь 2001 г.). «Циклин-зависимые киназы предотвращают повторную репликацию ДНК с помощью множества механизмов». Природа. 411 (6841): 1068–73. Bibcode:2001Натура.411.1068N. Дои:10.1038/35082600. PMID 11429609. S2CID 4393812.
- ^ Аршамбо V, Икуи А.Е., Драпкин Б.Дж., Кросс FR (август 2005 г.). «Нарушение механизмов, предотвращающих репликацию, вызывает реакцию повреждения ДНК». Мол. Клетка. Биол. 25 (15): 6707–21. Дои:10.1128 / MCB.25.15.6707-6721.2005. ЧВК 1190345. PMID 16024805.
- ^ Triolo T, Sternglanz R (май 1996 г.). «Роль взаимодействий между комплексом распознавания источника и SIR1 в подавлении транскрипции». Природа. 381 (6579): 251–3. Bibcode:1996Натура.381..251Т. Дои:10.1038 / 381251a0. PMID 8622770. S2CID 4309206.
- ^ Fox CA, Ehrenhofer-Murray AE, Loo S, Rine J (июнь 1997 г.). «Комплекс распознавания происхождения, SIR1 и требование фазы S для подавления». Наука. 276 (5318): 1547–51. Дои:10.1126 / science.276.5318.1547. PMID 9171055.
- ^ Клемм Р.Д., Остин Р.Дж., Белл С.П. (февраль 1997 г.). «Координированное связывание АТФ и исходной ДНК регулирует АТФазную активность комплекса распознавания происхождения». Клетка. 88 (4): 493–502. Дои:10.1016 / S0092-8674 (00) 81889-9. PMID 9038340.
- ^ Клемм Р.Д., Белл С.П. (июль 2001 г.). «АТФ, связанный с комплексом распознавания происхождения, важен для образования preRC». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 98 (15): 8361–7. Bibcode:2001PNAS ... 98.8361K. Дои:10.1073 / pnas.131006898. ЧВК 37444. PMID 11459976.
- ^ Бауэрс Дж. Л., Рэнделл Дж. К., Чен С., Белл С. П. (декабрь 2004 г.). «Гидролиз АТФ с помощью ORC катализирует повторяющуюся сборку Mcm2-7 в определенном ориджине репликации». Мол. Клетка. 16 (6): 967–78. Дои:10.1016 / j.molcel.2004.11.038. PMID 15610739.
- ^ Рэнделл Дж. К., Бауэрс Дж. Л., Родригес Х. К., Белл С. П. (январь 2006 г.). «Последовательный гидролиз АТФ с помощью Cdc6 и ORC направляет загрузку геликазы Mcm2-7». Мол. Клетка. 21 (1): 29–39. Дои:10.1016 / j.molcel.2005.11.023. PMID 16387651.
- ^ Семпл Дж. У., Да-Силва Л. Ф., Джервис Э. Дж., Ах-Ки Дж., Аль-Аттар Х, Куммер Л., Хейккила Дж. Дж., Пасеро П., Данкер Б. П. (ноябрь 2006 г.). «Существенная роль Orc6 в репликации ДНК через поддержание пререпликативных комплексов». EMBO J. 25 (21): 5150–8. Дои:10.1038 / sj.emboj.7601391. ЧВК 1630405. PMID 17053779.
- ^ а б c d Морган, Дэвид (2007). Клеточный цикл: принципы контроля. Учебники по биологии. С. 62–75. ISBN 978-0878935086.
- ^ Аусианникава, Дарья; Аллерс, Торстен (31 января 2017 г.). «Разнообразие репликации ДНК в архее». Гены. 8 (2): 56. Дои:10.3390 / genes8020056. ЧВК 5333045. PMID 28146124.
- ^ Фернандес-Сид, Алехандра (зима 2017 г.). «Комплекс ORC / Cdc6 / MCM2-7 образуется в результате многоступенчатой реакции, чтобы служить платформой для сборки двойного гексамера MCM». Молекулярная клетка. 50 (4): 577–588. Дои:10.1016 / j.molcel.2013.03.026. PMID 23603117.
- ^ Рэнделл, Джон (зима 2017 г.). «Последовательный гидролиз АТФ с помощью Cdc6 и ORC направляет загрузку геликазы Mcm2-7». Молекулярная клетка. 21: 29–39. Дои:10.1016 / j.molcel.2005.11.023. PMID 16387651.
- ^ а б Спек, Кристиан (зима 2017 г.). «АТФаза-зависимое кооперативное связывание ORC и Cdc6 с исходной ДНК». Структурная и молекулярная биология природы. 12 (11): 965–971. Дои:10.1038 / nsmb1002. ЧВК 2952294. PMID 16228006.
- ^ Юань, Цзуаньнин; Риера, Альберто; Бай, Линь; Сунь, Цзинчуань; Нанди, Сайкат; Спанос, Христос; Чен, Чжо Ангел; Барбон, Марта; Раппсильбер, Юри; Стиллман, Брюс; Спек, Кристиан; Ли, Хуэйлинь (13 февраля 2017 г.). «Структурная основа загрузки репликативной геликазы Mcm2-7 с помощью ORC – Cdc6 и Cdt1». Структурная и молекулярная биология природы. 24 (3): 316–324. Дои:10.1038 / nsmb.3372. ЧВК 5503505. PMID 28191893.
- ^ Каваками, Хиронори (зима 2017 г.). «Специфическое связывание эукариотических ORC с источниками репликации ДНК зависит от высококонсервативных основных остатков». Научные отчеты. 5: 14929. Дои:10.1038 / srep14929. ЧВК 4601075. PMID 26456755.
- ^ Чистол, Георгий (зима 2017). «Визуализация загрузки ДНК MCM2-7 с помощью одной молекулы: видеть - значит верить». Клетка. 161 (3): 429–430. Дои:10.1016 / j.cell.2015.04.006. PMID 25910200.
дальнейшее чтение
- Белл, Стивен П.; Дутта, Аниндья (июль 2002 г.). «Репликация ДНК в эукариотических клетках». Ежегодный обзор биохимии. Ежегодные обзоры. 71: 333–374. Дои:10.1146 / annurev.biochem.71.110601.135425. PMID 12045100.
Комплексный обзор молекулярной репликации ДНК