Старение - Senescence

Старение (/sɪˈпɛsəпs/) или же биологический старение постепенное ухудшение функциональный характеристики. Слово старение может относиться либо к клеточное старение или к старению всего организм. Старение организма связано с увеличением уровень смертности и / или уменьшение плодовитость с возрастом, по крайней мере, в последней части организма жизненный цикл.
Старение - это неизбежная судьба всех многоклеточные организмы с зародыш -сома разделение[1][2] но это может быть отложено. Открытие в 1934 г. ограничение калорий может увеличить продолжительность жизни крыс на 50%, а существование видов, имеющих незначительное старение и потенциально бессмертные организмы, такие как Гидра, мотивировали исследования по задержке старения и, следовательно, возрастные заболевания. Редкие человеческие мутации могут вызывать болезни ускоренного старения.
Факторы окружающей среды могут повлиять на старение, например, чрезмерное воздействие ультрафиолетовая радиация ускоряет старение кожи. Разные части тела могут стареть с разной скоростью. Два организма одного вида также могут стареть с разной скоростью, что делает биологическое старение и хронологическое старение разными понятиями.
Определение и характеристика
Органическое старение это старение целого организма. Актуарное старение можно определить как увеличение смертности и / или снижение плодовитость с возрастом. В Закон смертности Гомперца-Мейкхема говорит, что возрастной компонент смертность увеличивается экспоненциально с возрастом.
В 2013 году группа ученых определила девять признаков старения, общих для организмов, с акцентом на млекопитающих:
- геномная нестабильность,
- истощение теломер,
- эпигенетические изменения,
- утрата протеостаз,
- нерегулируемое восприятие питательных веществ,
- митохондриальная дисфункция,
- клеточное старение,
- истощение стволовых клеток,
- измененная межклеточная коммуникация.[3]
Старение характеризуется снижением способности реагировать на стресс, повышенной гомеостатический дисбаланс и повышенный риск заболевания, связанные со старением включая рак и сердечное заболевание. Старение определяется как «прогрессирующее ухудшение физиологической функции, присущий возрасту процесс потери жизнеспособности и повышения уязвимости».[4]
Окружающая среда наносит урон на разных уровнях, например повреждение ДНК, и повреждение тканей и клеток кислородом радикалы (широко известный как свободные радикалы ), и некоторые из этих повреждений не восстанавливаются и накапливаются со временем.[5] Клонирование из соматические клетки а не половые клетки могут начать жизнь с более высокой начальной нагрузкой повреждений. Овечка Долли умер молодым от заразной болезни легких, но для измерения уровня смертности и количественной оценки старения потребуются данные обо всей популяции клонированных особей.[нужна цитата ]
Теоретик эволюции Джордж Уильямс писал: «Замечательно, что после, казалось бы, чудесного подвига морфогенез, комплекс многоклеточный должен быть не в состоянии выполнять гораздо более простую задачу простого поддержания того, что уже сформировано ".[6]
Различия между видами
Разные скорости увеличения смертности с возрастом соответствуют разным максимальная продолжительность жизни среди разновидность. Например, мышь пожилой в 3 года, человек пожилой в 80 лет,[7] и гинкго деревья мало влияют на возраст даже в 667 лет.[8]
Почти все организмы стареют, в том числе бактерии которые имеют асимметрию между «материнскими» и «дочерними» клетками при делении клеток, при этом материнская клетка испытывает старение, в то время как дочерняя клетка омолаживается.[9][10] Есть незначительное старение в некоторых группах, таких как род Гидра.[11] Планарий плоские черви иметь "очевидно безграничные теломер регенеративная способность, поддерживаемая популяцией высокопродуктивных взрослых стволовые клетки."[12] Эти планарии не биологически бессмертный, а их смертность медленно увеличивается с возрастом. Организмы, которые считаются биологически бессмертными, в одном случае были бы Турритопсис Dohrnii, также известная как бессмертная медуза. В Турритопсис Dohrnii получил такое звание благодаря способности вернуться к своей молодости, когда он подвергается стрессу в зрелом возрасте.[13] В репродуктивная система остается нетронутым, и даже гонады Турритопсис Dohrnii существуют.[14]
Некоторые виды проявляют «отрицательное старение», при котором способность к воспроизводству увеличивается или остается стабильной, а смертность падает с возрастом, что является результатом преимуществ увеличения размеров тела во время старения.[15]
Эволюционные теории старения
Накопление мутаций
Естественный отбор может поддерживать смертельный и вредный аллели, если их действие ощущается после воспроизведения. Генетик Дж. Б. С. Холдейн интересно, почему доминирующая мутация, вызывающая болезнь Хантингтона остался в популяции, и почему естественный отбор не устранил его. Это неврологическое заболевание начинается (в среднем) в возрасте 45 лет и неизменно приводит к летальному исходу в течение 10–20 лет. Холдейн предположил, что в предысторию человечества лишь немногие доживали до 45 лет. Так как немногие были живы в более старшем возрасте, и их вклад в следующее поколение был поэтому мал по сравнению с большими когортами более молодых возрастных групп, сила отбора против таких поздних действий соответственно, вредоносных мутаций было мало. Следовательно, генетическая нагрузка поздних вредных мутаций может быть значительным при баланс мутации и отбора. Эта концепция стала известна как тень выделения.[16]
Питер Медавар формализовал это наблюдение в своем теория накопления мутаций старения.[17][18] «Сила естественного отбора ослабевает с возрастом - даже в теоретически бессмертной популяции, при условии только, что она подвергается реальным опасностям смертности. Если генетическая катастрофа ... происходит достаточно поздно в индивидуальной жизни, ее последствия могут быть совершенно несущественными. ". Известны «реальные опасности смертности», такие как хищничество, болезни и несчастные случаи »внешняя смертность ', и означают, что даже население с незначительное старение будет меньше людей в старших возрастных группах.[нужна цитата ]
Антагонистическая плейотропия
Другая эволюционная теория старения была предложена Джордж К. Уильямс[19] и включает антагонистическая плейотропия. Один ген может влиять на несколько черт. Некоторые черты, улучшающие физическую форму в раннем возрасте, могут иметь негативные последствия в более позднем возрасте. Но поскольку в молодом возрасте живет гораздо больше людей, чем в пожилом, даже небольшие положительные эффекты на раннем этапе могут быть сильно отобраны, а большие отрицательные эффекты позже могут быть очень слабо отобраны. Уильямс предложил следующий пример: возможно, ген кодирует отложение кальция в костях, что способствует выживанию молодых особей и, следовательно, будет благоприятствовать естественному отбору; однако этот же ген способствует отложению кальция в артериях, вызывая отрицательные атеросклеротические эффекты в пожилом возрасте. Таким образом, вредные биологические изменения в старости могут быть результатом отбора плейотропный гены, полезные в раннем возрасте, но вредные в дальнейшем. В этом случае давление отбора относительно велико, когда Репродуктивная ценность Фишера высокий и относительно низкий, когда репродуктивная ценность Фишера низкая.
Адаптивное старение
Запрограммированные теории старения постулируют, что старение является адаптивным, обычно вызывая отбор для эволюционируемость или же групповой выбор.
В теория репродуктивного клеточного цикла предполагает, что старение регулируется изменениями гормональных сигналов на протяжении всей жизни.[20]
Одноразовая сома
В одноразовая сома теория старения была предложена Томасом Кирквудом в 1977 году. Теория предполагает, что старение происходит из-за стратегии, при которой человек вкладывает средства в поддержание сомы только до тех пор, пока у нее есть реальный шанс на выживание.[21] Вид, который использует ресурсы более эффективно, будет жить дольше и, следовательно, сможет передавать генетическую информацию следующему поколению. Требования к воспроизводству высоки, поэтому меньше усилий вкладывается в ремонт и поддержание соматических клеток по сравнению с клетки зародышевой линии, чтобы сосредоточиться на воспроизводстве и выживании видов.[22]
Клеточное старение
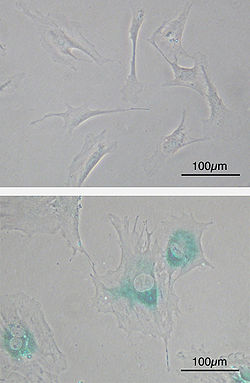
(верхний) Первичные эмбриональные фибробластные клетки мыши (MEF) до старения. Веретенообразный. (внизу) MEF стали дряхлыми после пассажей. Клетки увеличиваются в размерах, приобретают более плоскую форму и имеют выраженное старение. β-галактозидаза (САБГ, синие области) - маркер клеточного старения.
Клетки накапливают урон с течением времени. Особенно Повреждение ДНК, например из-за активных форм кислорода, приводит к накоплению вредных соматические мутации.[23]
Теория клеточного старения утверждает, что старение организма является следствием накопления менее физиологически полезных, то есть стареющих клеток. В соответствии с этим экспериментальное удаление стареющих клеток из трансгенных прогероид мышей[24] и непрогероидные мыши естественного возраста[25][26][27] привело к большему сопротивлению заболевания, связанные со старением. Внематочное выражение фактора эмбриональной транскрипции, NANOG, как показано, обращает вспять старение и восстанавливает способность к пролиферации и дифференцировке стареющих стволовых клеток.[28][29][30][31][32]
У многих организмов есть асимметричное деление клеток, например а стволовая клетка деление с образованием одной стволовой клетки и одной не стволовой клетки. Клеточный мусор, который накапливают клетки, неравномерно распределяется между новыми клетками при их делении. Вместо этого больше повреждений передается одной из клеток, в результате чего другая клетка остается омоложенной.[33] Одна ветвь затем претерпевает клеточное старение быстрее, чем другая.
Естественный отбор может удалить поврежденные клетки и предотвратить их распространение, уравновешивая естественную тенденцию к накоплению поврежденных клеток. Однако некоторые клетки мутируют способами, которые избегают этих механизмов контроля. Рак клетки избегают репликативного старения, чтобы стать бессмертными. Примерно в 85% опухолей это уклонение от клеточного старения является результатом активации их теломераза гены.[34]
У большинства многоклеточных видов соматические клетки в конце концов испытать репликативное старение и не могут разделять. Это может сильно предотвратить мутировавший клетки от становления раковый. В культуре фибробласты может достигать максимум 50 делений клеток; этот максимум известен как Лимит Хейфлика.[35] Репликативное старение является результатом теломер сокращение, которое в конечном итоге вызывает Повреждение ДНК отклик. Клетки также можно вызвать к старению через повреждение ДНК в ответ на повышенное активные формы кислорода (ROS), активация онкогены и сотовыйслияние клеток, независимо от длины теломер.[36]
Теория компромисса между раком и клеточным старением
Старые клетки внутри многоклеточный организм может быть очищен конкуренцией между клетками, но это увеличивает риск рака. Это приводит к неизбежной дилемме между двумя возможностями - накоплением физиологически бесполезных стареющих клеток и раком, - обе из которых приводят к увеличению показателей смертности с возрастом.[1]
Химическое повреждение

Одной из самых ранних теорий старения была Скорость живой гипотезы описанный Раймонд Перл в 1928 г.[37] (на основе более ранней работы Макс Рубнер ), в котором говорится, что быстро базальная скорость метаболизма соответствует короткому максимальная продолжительность жизни.
Хотя может быть некоторая обоснованность идеи, что для различных типов конкретных повреждений, подробно описанных ниже, это побочные продукты метаболизм при прочих равных условиях быстрый метаболизм может сократить продолжительность жизни, в целом эта теория неадекватно объясняет различия в продолжительности жизни как внутри, так и между видами. Калорийность ограничена животные перерабатывают столько или больше калорий на грамм массы тела, сколько их вволю откормленные аналоги, но демонстрируют значительно более длительный срок службы.[нужна цитата ] Точно так же скорость метаболизма является плохим показателем продолжительности жизни для птиц, летучих мышей и других видов, которые, как предполагается, снизили смертность от хищников и, следовательно, развивали большую продолжительность жизни даже при очень высоких показателях метаболизма.[38] В ходе анализа 2007 года было показано, что современные статистические методы коррекции влияния размеров тела и филогения используются, скорость метаболизма не коррелирует с продолжительностью жизни у млекопитающих или птиц.[39] (Для критики Скорость живой гипотезы видеть Жить быстро, когда умирает?[40])
Что касается конкретных типов химического повреждения, вызванного метаболизмом, предполагается, что повреждение долгоживущих биополимеры, например, структурные белки или же ДНК, вызванные повсеместными химическими веществами в организме, такими как кислород и сахара, частично ответственны за старение. Повреждение может включать обрыв биополимерных цепей, сшивание биополимеров, или химическое присоединение неестественных заместителей (гаптены ) к биополимерам.[нужна цитата ]Под нормальным аэробный условий, примерно 4% кислород метаболизируется митохондрии конвертируется в супероксид ион, который впоследствии может быть преобразован в пероксид водорода, гидроксил радикальный и, в конечном итоге, другие химически активные вещества, включая другие перекиси и синглетный кислород, что, в свою очередь, может генерировать свободные радикалы способны повредить структурные белки и ДНК.[5] Определенный металл ионы найдено в теле, например медь и утюг, может участвовать в процессе. (В Болезнь Вильсона, а наследственный дефект что заставляет организм удерживать медь, некоторые симптомы напоминают ускоренное старение.) Эти процессы называются окислительный стресс связаны с потенциальными преимуществами диетического полифенол антиоксиданты, например, в кофе,[41] красное вино и чай.[42]
Сахара Такие как глюкоза и фруктоза может реагировать определенными аминокислоты Такие как лизин и аргинин и определенные основания ДНК, такие как гуанин для производства сахарных аддуктов в процессе, называемом гликирование. Эти аддукты могут дополнительно перестраиваться с образованием реактивных частиц, которые затем могут поперечно связывать структурные белки или ДНК с аналогичными биополимерами или другими биомолекулами, такими как неструктурные белки. Люди с сахарный диабет, которые повысили содержание сахара в крови, расстройства, связанные со старением, развиваются намного раньше, чем у населения в целом, но могут отсрочить появление таких расстройств за счет строгого контроля уровня сахара в крови. Есть свидетельства того, что повреждение сахара связано с повреждением окислителей в процессе, называемом гликоксидирование.
Свободные радикалы может повредить белки, липиды или же ДНК. Гликирование в основном повреждает белки. Поврежденные белки и липиды накапливаются в лизосомы в качестве липофусцин. Химическое повреждение структурных белков может привести к потере функции; например, повреждение коллаген из кровеносный сосуд стенки могут привести к жесткости стенки сосуда и, таким образом, гипертония, утолщение стенки сосуда и образование реактивной ткани (атеросклероз ); аналогичные процессы в почка может привести к почечная недостаточность. Повреждение ферменты снижает функциональность сотовой связи. Липид перекисное окисление внутреннего митохондриальная мембрана уменьшает электрический потенциал и способность генерировать энергию. Наверное, не случайно, что почти все так называемые "болезни ускоренного старения "из-за дефектных Ремонт ДНК ферменты.[23][43]
Считается, что влияние алкоголя на старение частично можно объяснить активацией алкоголем Ось HPA, что стимулирует глюкокортикоид секреция, длительное воздействие которой вызывает симптомы старения.[44]
Биомаркеры старения
Если разные люди стареют с разной скоростью, то плодовитость, смертность и функциональные возможности могут быть лучше предсказаны с помощью биомаркеры чем по хронологическому возрасту.[45] Тем не мение, поседение волос,[46] морщины на коже и другие общие изменения, наблюдаемые с возрастом, не являются лучшими индикаторами будущей функциональности, чем хронологический возраст. Биогеронтологи продолжали попытки найти и проверить биомаркеры старения, но пока успехи были ограниченными. Уровни CD4 и CD8 Т-клетки памяти и наивные Т-клетки были использованы для получения хороших прогнозов ожидаемой продолжительности жизни мышей среднего возраста.[47]
Есть интерес к эпигенетические часы как биомаркер старения, основанный на его способности предсказывать хронологический возраст человека.[48] Основная кровь биохимия подсчет клеток также может использоваться для точного прогнозирования хронологического возраста.[49] Также возможно предсказать хронологический возраст человека, используя часы транскриптомного старения.[50]
Генетические детерминанты старения
С помощью модельных организмов был идентифицирован ряд генетических компонентов старения, начиная от простых почкователей. дрожжи Saccharomyces cerevisiae червям, таким как Caenorhabditis elegans и плодовые мошки (Drosophila melanogaster ). Изучение этих организмов выявило наличие как минимум двух консервативных путей старения.
Экспрессия генов контролируется несовершенно, и возможно, что случайные колебания уровней экспрессии многих генов вносят вклад в процесс старения, как предполагает исследование таких генов у дрожжей.[51] Индивидуальные клетки, которые генетически идентичны, тем не менее могут иметь существенно разные ответы на внешние стимулы и заметно разную продолжительность жизни, что указывает на то, что эпигенетические факторы играют важную роль в экспрессии генов и старении, а также генетические факторы.
Способность восстанавливать двухцепочечные разрывы ДНК снижается с возрастом у мышей.[52] и люди.[53]
Набор редких потомственных (генетика ) расстройства, каждое из которых называется прогерия, было известно некоторое время. У больных появляются симптомы, похожие на ускоренное старение, включая морщинистая кожа. Причина Синдром прогерии Хатчинсона-Гилфорда было сообщено в журнале Природа в мае 2003 г.[54]Этот отчет предполагает, что Повреждение ДНК, нет окислительный стресс, является причиной этой формы ускоренного старения.
Смотрите также
- Старение
- Стареющий мозг
- Заболевания, связанные со старением
- Антивозрастное движение
- Ремонт ДНК
- Свободные радикалы
- Генетика старения
- Гериатрия
- Геронтология
- Гомеостатическая способность
- Бессмертие
- Продление жизни
- Митохормезис
- Старость
- Окислительный стресс
- Феноптоз
- Старение растений
- Запрограммированная гибель клеток
- Регенеративная медицина
- Омоложение
- SAGE KE
- Теория старения стволовых клеток
- Стратегии искусственно незначительного старения (СЕНС)
- Сублетальный урон
- Трансформационный дизайн
Рекомендации
- ^ а б Нельсон, Пол; Масел, Джоанна (5 декабря 2017 г.). «Межклеточная конкуренция и неизбежность многоклеточного старения». Труды Национальной академии наук. 114 (49): 12982–87. Дои:10.1073 / pnas.1618854114. ЧВК 5724245. PMID 29087299.
- ^ Вагнер, Гюнтер П. (5 декабря 2017 г.). «Сила отрицательных [теоретических] результатов». Труды Национальной академии наук. 114 (49): 12851–52. Дои:10.1073 / pnas.1718862114. ЧВК 5724295. PMID 29162687.
- ^ Лопес-Отин, К; и другие. (2013). «Признаки старения». Клетка. 153 (6): 1194–217. Дои:10.1016 / j.cell.2013.05.039. ЧВК 3836174. PMID 23746838.
- ^ «Глоссарий по старению и геронтологии». Получено 26 февраля 2011.
- ^ а б Холмс Г.Е., Бернштейн С., Бернштейн Н. (сентябрь 1992 г.). «Окислительные и другие повреждения ДНК как основа старения: обзор». Мутат. Res. 275 (3–6): 305–15. Дои:10.1016 / 0921-8734 (92) 90034-м. PMID 1383772.
- ^ Уильямс, Г. (1957). «Плейотропия, естественный отбор и эволюция старения». Эволюция. 11 (4): 398–411. Дои:10.2307/2406060. JSTOR 2406060.
- ^ Austad, S (2009). «Сравнительная биология старения». J Gerontol A Biol Sci Med Sci. 64 (2): 199–201. Дои:10.1093 / gerona / gln060. ЧВК 2655036. PMID 19223603.
- ^ Ван, Ли; Цуй, Цзявэнь; Цзинь, Бяо; Чжао, Цзяньго; Сюй, Хуйминь; Лу, Чжаогэн; Ли, Вэйсин; Ли, Сяося; Ли, Линлинг; Лян, Эрюань; Рао, Сяолань; Ван, Шуфанг; Фу, Чуньсян; Цао, Фулянь; Диксон, Ричард А .; Линь, Цзиньсин (13 января 2020 г.). «Многовариантный анализ сосудистых камбиальных клеток выявил механизмы долголетия старых деревьев гинкго билоба». Труды Национальной академии наук. 117 (4): 2201–2210. Дои:10.1073 / pnas.1916548117. ISSN 0027-8424. ЧВК 6995005. PMID 31932448.
- ^ Ackermann, M .; Stearns, S.C .; Дженал, У. (2003). «Старение бактерии с асимметричным делением». Наука. 300 (5627): 1920. Дои:10.1126 / science.1083532. PMID 12817142. S2CID 34770745.
- ^ Стюарт, Э. Дж .; Madden, R .; Paul, G .; Таддеи, Ф. (2005). «Старение и смерть в организме, размножающемся морфологически симметричным делением». PLOS Биология. 3 (2): e45. Дои:10.1371 / journal.pbio.0030045. ЧВК 546039. PMID 15685293.
- ^ Dańko, Maciej J .; Козловский, Ян; Шейбл, Ральф (2015). «Раскрытие феномена незрелости Гидры». Журнал теоретической биологии. 382: 137–49. Дои:10.1016 / j.jtbi.2015.06.043. ISSN 0022-5193. PMID 26163368.
- ^ Томас К. Дж. Тан; Руман Рахман; Фарах Джабер-Хиджази; Даниэль А. Феликс; Чен Чен; Эдвард Дж. Луи и Азиз Абубейкер (февраль 2012 г.). «Поддержание теломер и активность теломеразы по-разному регулируются у бесполых и половых червей». PNAS. 109 (9): 4209–14. Bibcode:2012ПНАС..109.4209Т. Дои:10.1073 / pnas.1118885109. ЧВК 3306686. PMID 22371573.
- ^ Лисенкова, А. А .; Григоренко, А.П .; Тяжелова, Т. В .; Андреева, Т. В .; Гусев, Ф. Э .; Манахов, А.Д .; Гольцов, А.Ю .; Piraino, S .; Miglietta, M.P .; Рогаев Э.И. (1 февраля 2017 г.). «Полный митохондриальный геном и эволюционный анализ Turritopsis dohrnii,« бессмертной »медузы с обратимым жизненным циклом». Молекулярная филогенетика и эволюция. 107: 232–238. Дои:10.1016 / j.ympev.2016.11.007. ISSN 1055-7903. PMID 27845203.
- ^ Piraino, S .; Boero, F .; Aeschbach, B .; Шмид В. (1 июня 1996 г.). «Обращение вспять жизненного цикла: превращение медуз в полипы и трансдифференцировка клеток в Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический бюллетень. 190 (3): 302–312. Дои:10.2307/1543022. ISSN 0006-3185. JSTOR 1543022. PMID 29227703.
- ^ В. Ваупель, Джеймс; Баудиш, Аннетт; Дёллинг, Мартин; А. Роуч, Дебора; Гамп, Ютта (июнь 2004 г.). «Дело об отрицательном старении». Теоретическая популяционная биология. 65 (4): 339–51. Дои:10.1016 / j.tpb.2003.12.003. PMID 15136009.
- ^ Фабиан, Даниэль; Флатт, Томас (2011). «Эволюция старения». Scitable. Издательская группа Nature.
- ^ Медавар ПБ (1946). «Старость и естественная смерть». Modern Quarterly. 1: 30–56.
- ^ Медавар, Питер Б. (1952). Нерешенная проблема биологии. Лондон: Х. К. Льюис.[страница нужна ]
- ^ Уильямс, Джордж К. (декабрь 1957 г.). «Плейотропия, естественный отбор и эволюция старения». Эволюция. 11 (4): 398–411. Дои:10.2307/2406060. JSTOR 2406060.
- ^ Bowen RL; Этвуд CS (2011). "Теория старения репродуктивно-клеточного цикла: обновление". Экспериментальная геронтология. 46 (2): 100–07. Дои:10.1016 / j.exger.2010.09.007. PMID 20851172. S2CID 20998909.
- ^ Молотки, М .; Ричардсон, Д. С .; Burke, T .; Комдеур, Дж. (2013). «Влияние репродуктивных инвестиций и условий окружающей среды в раннем возрасте на старение: поддержка гипотезы одноразовой сомы» (PDF). Журнал эволюционной биологии. 26 (9): 1999–2007. Дои:10.1111 / jeb.12204. HDL:11370 / 9cc6749c-f67d-40ab-a253-a06650c32102. ISSN 1420-9101. PMID 23961923.
- ^ Кирквуд, Т. Б. Л .; Rose, M. R .; Харви, Пол Х .; Куропатка, Линда; Саутвуд, сэр Ричард (29 апреля 1991 г.). «Эволюция старения: поздняя выживаемость в жертву воспроизводству». Философские труды Лондонского королевского общества. Серия B: Биологические науки. 332 (1262): 15–24. Дои:10.1098 / рстб.1991.0028. PMID 1677205.
- ^ а б Bernstein, H; Пейн, СМ; Бернштейн, К; Garewal, H; Дворжак, К. (2008). «Рак и старение как последствия неремонтированного повреждения ДНК».. В Кимуре, Хонока; Судзуки, Аой (ред.). Новое исследование повреждений ДНК. Издательство Nova Science. С. 1–47. ISBN 978-1604565812.
- ^ Baker, D .; Wijshake, T .; Tchkonia, T .; LeBrasseur, N .; Чайлдс, B .; van de Sluis, B .; Kirkland, J .; ван Дерсен, Дж. (10 ноября 2011 г.). «Удаление p16Ink4a-позитивных стареющих клеток задерживает связанные со старением расстройства». Природа. 479 (7372): 232–36. Bibcode:2011Натура.479..232Б. Дои:10.1038 / природа10600. ЧВК 3468323. PMID 22048312.
- ^ Сюй, М; Палмер, АК; Дин, Н; Вейвода, ММ; Пирцхалава, Т; Белый, ТА; Сепе, А; Джонсон, нокаут; Стаут, МБ; Гиоргадзе, Н; Дженсен, доктор медицины; LeBrasseur, NK; Чкония, Т; Киркланд, JL (2015). «Нацеливание на стареющие клетки улучшает адипогенез и метаболическую функцию в пожилом возрасте». eLife. 4: e12997. Дои:10.7554 / eLife.12997. ЧВК 4758946. PMID 26687007.
- ^ Быстро, Даррен (3 февраля 2016 г.). «Очистка поврежденных клеток у мышей увеличивает продолжительность жизни до 35 процентов». www.gizmag.com.
- ^ Регаладо, Антонио (3 февраля 2016 г.). «В новой стратегии борьбы со старением очистка старых клеток увеличивает продолжительность жизни мышей на 25 процентов». Обзор технологий MIT.
- ^ Шахини А., Чоудхури Д., Асмани М., Чжао Р., Лей П., Андредис С. (январь 2018 г.). «NANOG восстанавливает нарушенный потенциал миогенной дифференцировки скелетных миобластов после многократного удвоения популяции». Исследования стволовых клеток. 26: 55–66. Дои:10.1016 / j.scr.2017.11.018. PMID 29245050.
- ^ Шахини А., Мистриотис П., Асмани М., Чжао Р., Андредис С. (июнь 2017 г.). «NANOG восстанавливает сократимость стареющих микротканей на основе мезенхимальных стволовых клеток». Ткань Eng Часть A. 23 (11–12): 535–45. Дои:10.1089 / ten.TEA.2016.0494. ЧВК 5467120. PMID 28125933.
- ^ Mistriotis P, Bajpai V, Wang X, Rong N, Shahini A, Asmani M, Liang M, Wang J, Lei P, Liu S, Zhao R, Andreadis S (январь 2017 г.). «NANOG обращает вспять потенциал миогенной дифференцировки стареющих стволовых клеток, восстанавливая нитевидную организацию ACTIN и экспрессию генов, зависимую от SRF». Стволовые клетки. 35 (1): 207–21. Дои:10.1002 / шток.2452. PMID 27350449.
- ^ Хан Дж., Мистриотис П., Лей П., Ван Д., Лю С., Чжао Р., Андредис С. (декабрь 2012 г.). «Nanog обращает вспять эффекты старения организма на пролиферацию мезенхимальных стволовых клеток и потенциал миогенной дифференцировки». Стволовые клетки. 30 (12): 2746–59. Дои:10.1002 / шток.1223. ЧВК 3508087. PMID 22949105.
- ^ Munst B, Thier M, Winnemoller D, Helfen M, Thummer R, Edenhofer F (январь 2016 г.). «Nanog вызывает подавление старения за счет подавления экспрессии p27KIP1». Журнал клеточной науки. 129 (5): 912–20. Дои:10.1242 / jcs.167932. ЧВК 4813312. PMID 26795560.
- ^ Стивенс С. (апрель 2005 г.). «Старение: стареют даже бактерии». Curr. Биол. 15 (8): R308–10. Дои:10.1016 / j.cub.2005.04.006. PMID 15854899. S2CID 15499468.
- ^ Hanahan D; Вайнберг Р.А. (январь 2000 г.). «Признаки рака». Клетка. 100 (1): 57–70. Дои:10.1016 / S0092-8674 (00) 81683-9. PMID 10647931. S2CID 1478778.
- ^ Hayflick L; Moorhead PS (декабрь 1961 г.). «Серийное культивирование штаммов диплоидных клеток человека». Exp. Cell Res. 25 (3): 585–621. Дои:10.1016/0014-4827(61)90192-6. PMID 13905658.
- ^ Хён, Анника; Вебер, Даниэла; Юнг, Тобиас; Отт, Кристиана; Хьюго, Мартин; Кочлик, Бастиан; Кем, Ричард; Кениг, Жаннетт; Грун, Тилман; Кастро, Хосе Педро (7 декабря 2016 г.). «Счастливо (н) навсегда: старение в контексте окислительного стресса, потери протеостаза и клеточного старения». Редокс Биология. 11: 482–501. Дои:10.1016 / j.redox.2016.12.001. ISSN 2213-2317. ЧВК 5228102. PMID 28086196.
- ^ Перл, Раймонд (1928). Уровень жизни, являющийся отчетом некоторых экспериментальных исследований биологии продолжительности жизни. Нью-Йорк: Альфред А. Кнопф.[страница нужна ]
- ^ Брюне-Россинни АК; Austad SN (2004). «Исследования старения летучих мышей: обзор». Биогеронтология. 5 (4): 211–22. Дои:10.1023 / B: BGEN.0000038022.65024.d8. PMID 15314271. S2CID 22755811.
- ^ de Magalhães JP; Коста Дж; Церковь GM (1 февраля 2007 г.). «Анализ взаимосвязи между метаболизмом, графиком развития и долголетием с использованием филогенетических независимых контрастов». Журналы геронтологии, серия A: биологические и медицинские науки. 62 (2): 149–60. CiteSeerX 10.1.1.596.2815. Дои:10.1093 / gerona / 62.2.149. ЧВК 2288695. PMID 17339640.
- ^ Speakman JR; Селман С; McLaren JS; Харпер EJ (1 июня 2002 г.). «Быстро жить, когда умереть? Связь между старением и энергией». Журнал питания. 132 (6 Прил. 2): 1583S – 97S. Дои:10.1093 / jn / 132.6.1583S. PMID 12042467.
- ^ Фридман Н.Д .; Парк Y; Abnet CC; Hollenbeck AR; Sinha R (май 2012 г.). «Связь употребления кофе с общей и конкретной причиной смертности». N. Engl. J. Med. 366 (20): 1891–904. Дои:10.1056 / NEJMoa1112010. ЧВК 3439152. PMID 22591295.
- ^ Ян Y; Chan SW; Hu M; Walden R; Томлинсон Б (2011). «Влияние некоторых общих пищевых компонентов на сердечно-сосудистые заболевания». ISRN Кардиол. 2011: 1–16. Дои:10.5402/2011/397136. ЧВК 3262529. PMID 22347642.
- ^ Пан М.Р., Ли К., Линь С.И., Хунг В.К. (май 2016 г.). «Соединяя точки: от повреждения и восстановления ДНК до старения». Int J Mol Sci. 17 (5): 685. Дои:10.3390 / ijms17050685. ЧВК 4881511. PMID 27164092.
- ^ Спенсер Р.Л.; Хатчисон К.Э. (1999). «Алкоголь, старение и реакция на стресс» (PDF). Исследования алкоголя и здоровье. 23 (4): 272–83. ЧВК 6760387. PMID 10890824.
- ^ Джордж Т. Бейкер, III и Ричард Л. Спротт (1988). «Биомаркеры старения». Экспериментальная геронтология. 23 (4–5): 223–39. Дои:10.1016/0531-5565(88)90025-3. PMID 3058488. S2CID 31039588.
- ^ Ван Несте Д., Тобин Д. Д. (2004). «Цикл волос и пигментация волос: динамические взаимодействия и изменения, связанные со старением». МИКРОН. 35 (3): 193–200. Дои:10.1016 / j.micron.2003.11.006. PMID 15036274.
- ^ Миллер Р.А. (2001). «Биомаркеры старения: прогнозирование долголетия с использованием определения подгруппы Т-клеток с учетом возраста в генетически гетерогенной популяции мышей среднего возраста». Журналы геронтологии. 56 (4): B180–86. Дои:10.1093 / gerona / 56.4.b180. PMID 11283189.
- ^ Хорват С. (2013). «Возраст метилирования ДНК человеческих тканей и типов клеток». Геномная биология. 14 (10): R115. Дои:10.1186 / gb-2013-14-10-r115. ЧВК 4015143. PMID 24138928.
- ^ Жаворонков А (2016). «Глубинные биомаркеры старения человека: применение глубоких нейронных сетей для разработки биомаркеров». Старение. 8 (5): 1021–33. Дои:10.18632 / старение.100968. ЧВК 4931851. PMID 27191382.
- ^ Петерс М (2015). «Транскрипционный ландшафт периферической крови человека». Nature Communications. 6: 8570. Bibcode:2015НатКо ... 6.8570.. Дои:10.1038 / ncomms9570. ЧВК 4639797. PMID 26490707.
- ^ Райли Дж; Перейра-Смит О.М. (2006). «Микрофлюидное устройство для анализа экспрессии генов отдельных клеток в Saccharomyces cerevisiae». Дрожжи. 23 (14–15): 1065–73. Дои:10.1002 / год.1412. PMID 17083143.
- ^ Вайдья А., Мао З., Тиан Х, Спенсер Б., Селуанов А., Горбунова В. (июль 2014 г.). «Мыши-репортеры с поврежденными генами демонстрируют, что восстановление ДНК с помощью негомологичных концевых соединений снижается с возрастом». PLOS Genet. 10 (7): e1004511. Дои:10.1371 / journal.pgen.1004511. ЧВК 4102425. PMID 25033455.
- ^ Ли З, Чжан В., Чен Й, Го В., Чжан Дж., Тан Х, Сюй З, Чжан Х, Тао Й, Ван Ф, Цзян Й, Сунь Флорида, Мао З (ноябрь 2016 г.). «Нарушение репарации двухцепочечных разрывов ДНК способствует возрастанию геномной нестабильности у людей». Разница в гибели клеток. 23 (11): 1765–77. Дои:10.1038 / cdd.2016.65. ЧВК 5071568. PMID 27391797.
- ^ Mounkes LC; Козлов С (2003). «Прогероидный синдром у мышей вызван дефектами ламинов А-типа» (PDF). Природа. 423 (6937): 298–301. Bibcode:2003Натура.423..298М. Дои:10.1038 / природа01631. PMID 12748643. S2CID 4360055.