Мотив распознавания РНК - RNA recognition motif
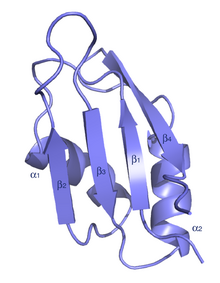
| Мотив узнавания РНК. (он же домен RRM, RBD или RNP) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||
| Символ | RRM_1 | ||||||||||
| Pfam | PF00076 | ||||||||||
| Pfam клан | RRM CL0221 RRM | ||||||||||
| ИнтерПро | IPR000504 | ||||||||||
| PROSITE | PDOC00030 | ||||||||||
| SCOP2 | 1sxl / Объем / СУПФАМ | ||||||||||
| |||||||||||
Мотив распознавания РНК, РНП-1 это предполагаемый РНК переплет домен из примерно 90 аминокислот, которые, как известно, связывают одноцепочечные РНК. Он был найден во многих эукариотический белки.[1][2][3]
Самая большая группа однониточных РНК-связывающий белок представляет собой семейство мотивов узнавания эукариотической РНК (RRM), которое содержит консенсусную последовательность из восьми аминокислот RNP-1.[4][5]
Белки RRM обладают разнообразными предпочтениями и функциями связывания РНК и включают гетерогенные ядерные рибонуклеопротеины (hnRNPs ), белки, участвующие в регуляции альтернативное сращивание (SR, U2AF2, Sxl ), белковые компоненты малых ядерных рибонуклеопротеидов (U1 и U2 snRNPs ) и белки, регулирующие стабильность и трансляцию РНК (PABP, Ла, Ху).[2][3][5] RRM в гетеродимерном факторе сплайсинга Вспомогательный фактор U2 snRNP по-видимому, имеет два RRM-подобных домена со специализированными функциями для распознавания белков.[6] Мотив также присутствует в нескольких белках, связывающих одноцепочечную ДНК.
Типичный RRM состоит из четырех антипараллельных бета-нити и два альфа-спирали расположены в складке бета-альфа-бета-бета-альфа-бета с боковыми цепями, которые складываются с основаниями РНК. В некоторых случаях при связывании РНК присутствует третья спираль.[7] RRM рассматривается в ряде публикаций.[8][9][10]
Белки человека, содержащие этот домен
A2BP1; АКФ; БОЛЛ; БРУНОЛ4; БРУНОЛ5; БРУНОЛ6; CCBL2; CGI-96; CIRBP; CNOT4; CPEB2; CPEB3; CPEB4; CPSF7; CSTF2; CSTF2T; CUGBP1; CUGBP2; D10S102; DAZ1; DAZ2; DAZ3; DAZ4; DAZAP1; DAZL; ДНКJC17; DND1; EIF3S4; EIF3S9; EIF4B; EIF4H; ELAVL1; ELAVL2; ELAVL3; ELAVL4; ENOX1; ENOX2; EWSR1; FUS; FUSIP1; G3BP; G3BP1; G3BP2; GRSF1; HNRNPL; HNRPA0; HNRPA1; HNRPA2B1; HNRPA3; HNRPAB; HNRPC; HNRPCL1; HNRPD; HNRPDL; HNRPF; HNRPH1; HNRPH2; HNRPH3; HNRPL; HNRPLL; HNRPM; HNRPR; HRNBP1; HSU53209; HTATSF1; IGF2BP1; IGF2BP2; IGF2BP3; LARP7; MKI67IP; MSI1; MSI2; МССП-2; MTHFSD; MYEF2; NCBP2; NCL; NOL8; НЕТ НЕТ; P14; PABPC1; PABPC1L; PABPC3; PABPC4; PABPC5; PABPN1; POLDIP3; PPARGC1; PPARGC1A; PPARGC1B; PPIE; PPIL4; PPRC1; PSPC1; PTBP1; PTBP2; PUF60; РАЛИ; РАЛИЛ; RAVER1; RAVER2; RBM10; RBM11; RBM12; РБМ12Б; RBM14; RBM15; РБМ15Б; RBM16; RBM17; RBM18; RBM19; RBM22; RBM23; RBM24; RBM25; RBM26; RBM27; RBM28; RBM3; RBM32B; RBM33; RBM34; RBM35A; РБМ35Б; RBM38; RBM39; RBM4; RBM41; RBM42; RBM44; RBM45; RBM46; RBM47; RBM4B; RBM5; RBM7; RBM8A; RBM9; RBMS1; RBMS2; RBMS3; RBMX; RBMX2; RBMXL2; RBMY1A1; RBMY1B; RBMY1E; RBMY1F; RBMY2FP; RBPMS; RBPMS2; RDBP; RNPC3; RNPC4; RNPS1; ROD1; SAFB; SAFB2; SART3; SETD1A; SF3B14; SF3B4; SFPQ; SFRS1; SFRS10; SFRS11; SFRS12; SFRS15; SFRS2; SFRS2B; SFRS3; SFRS4; SFRS5; SFRS6; SFRS7; SFRS9; SLIRP; SLTM; SNRP70; SNRPA; SNRPB2; SPEN; SR140; SRRP35; SSB; SYNCRIP; TAF15; TARDBP; THOC4; TIA1; TIAL1; TNRC4; TNRC6C; TRA2A; TRSPAP1; TUT1; U1SNRNPBP; U2AF1; U2AF2; УХМК1; ZCRB1; ZNF638; ЗРСР1; ЗРСР2;
Рекомендации
- ^ Суонсон М.С., Дрейфус Г., Пинол-Рома С. (1988). «Гетерогенные ядерные рибонуклеопротеидные частицы и путь образования мРНК». Trends Biochem. Наука. 13 (3): 86–91. Дои:10.1016/0968-0004(88)90046-1. PMID 3072706.
- ^ а б Кин Дж. Д., Чемберс Дж. К., Кенан Д., Мартин Б. Дж. (1988). «Домены геномной структуры и аминокислотной последовательности аутоантигена La человека». J. Biol. Chem. 263 (34): 18043–51. PMID 3192525.
- ^ а б Дэвис Р. В., Сакс А. Б., Корнберг Р. Д. (1987). «Единственный домен дрожжевого поли (A) -связывающего белка необходим и достаточен для связывания РНК и жизнеспособности клеток». Мол. Клетка. Биол. 7 (9): 3268–76. Дои:10.1128 / mcb.7.9.3268. ЧВК 367964. PMID 3313012.
- ^ Bandziulis RJ, Swanson MS, Dreyfuss G (1989). «РНК-связывающие белки как регуляторы развития». Genes Dev. 3 (4): 431–437. Дои:10.1101 / gad.3.4.431. PMID 2470643.
- ^ а б Кин Дж. Д., Query CC, Bentley RC (1989). «Общий мотив распознавания РНК, идентифицированный в пределах определенного связывающего домена РНК U1 белка snRNP U1 70K». Клетка. 57 (1): 89–101. Дои:10.1016 / 0092-8674 (89) 90175-Х. PMID 2467746. S2CID 22127152.
- ^ Зеленый MR, Kielkopf CL, Lucke S (2004). «Мотивы гомологии U2AF: узнавание белка в мире RRM». Genes Dev. 18 (13): 1513–1526. Дои:10.1101 / gad.1206204. ЧВК 2043112. PMID 15231733.
- ^ Кумар С., Бирни Э, Крайнер А.Р. (1993). «Анализ мотива распознавания РНК и доменов RS и RGG: консервация в факторах сплайсинга пре-мРНК многоклеточных животных». Нуклеиновые кислоты Res. 21 (25): 5803–5816. Дои:10.1093 / nar / 21.25.5803. ЧВК 310458. PMID 8290338.
- ^ Кин Дж. Д., Кенан Д. Д., Query CC (1991). «Распознавание РНК: к определению детерминант специфичности». Trends Biochem. Наука. 16 (6): 214–20. Дои:10.1016 / 0968-0004 (91) 90088-д. PMID 1716386.
- ^ Аллен Ф.Х., Домингес С., Марис С. (2005). «Мотив распознавания РНК, пластиковая платформа для связывания РНК, регулирующая посттранскрипционную экспрессию генов». FEBS J. 272 (9): 2118–31. Дои:10.1111 / j.1742-4658.2005.04653.x. PMID 15853797.
- ^ Теплова М, Юань Ю.Р., Патель Д.Д., Малинина Л, Теплов А, Фан АТ, Ильин С (2006). «Структурная основа для распознавания и секвестрации UUU (OH) 3 'temini растущих транскриптов РНК-полимеразы III La, аутоантигеном ревматического заболевания». Мол. Клетка. 21 (1): 75–85. Дои:10.1016 / j.molcel.2005.10.027. ЧВК 4689297. PMID 16387655.
внешняя ссылка
- Ресурс Eukaryotic Linear Motif класс мотива LIG_ULM_U2AF65_1