Фенилаланин аммиак-лиаза - Phenylalanine ammonia-lyase
| фенилаланин аммиак-лиаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Рендеринг PDB на основе 1T6J. | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 4.3.1.24 | ||||||||
| Количество CAS | 9024-28-6 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Фенилаланин аммиаклиаза (EC 4.3.1.24 ) является фермент который катализирует реакция превращения L-фенилаланин к аммиак и транс-коричная кислота.[1] Фенилаланинаммиаклиаза (PAL) - первая и совершенный шаг в фенилпропаноидный путь и поэтому участвует в биосинтез из полифенол такие соединения, как флавоноиды, фенилпропаноиды, и лигнин в растения.[2][3] Фенилаланинаммиаклиаза широко встречается в растениях, а также в некоторых бактерии, дрожжи, и грибы, с изоферменты существуют у многих различных видов. Оно имеет молекулярная масса в диапазоне 270–330кДа.[1][4] Активность PAL резко индуцируется в ответ на различные раздражители, такие как повреждение тканей, патогенный атака, свет, низкие температуры и гормоны.[1][5] PAL недавно был изучен на предмет возможных терапевтических преимуществ у людей, страдающих фенилкетонурия.[6] Он также использовался при создании L-фенилаланин как предшественник подсластитель аспартам.[7]
Фермент входит в состав аммиака. лиазе семейство, которое разрывает связи углерод – азот. Как и другие лиазы, для PAL требуется только один субстрат для прямой реакции, но две для обратной. Считается, что механистически он похож на родственный фермент гистидин-аммиак-лиаза (EC: 4.3.1.3, HAL).[8] В систематическое название этого класса ферментов L-фенилаланин-аммиак-лиаза (образующая транс-циннамат). Ранее он обозначался EC 4.3.1.5, но этот класс был переименован в EC 4.3.1.24 (фенилаланинаммиаклиазы), EC 4.3.1.25 (тирозин-аммиак-лиазы) и EC 4.3.1.26 (фенилаланин / тирозин аммиак-лиазы). Другие широко используемые имена включают тираза, фенилаланин дезаминаза, тирозин аммиак-лиаза, L-тирозин аммиак-лиаза, фенилаланин-аммоний-лиаза, PAL, и L-фенилаланин аммиак-лиаза.
Механизм
Фенилаланин-аммиаклиаза специфична для L-phe, и в меньшей степени, L-тирозин.[9][10] Реакция, катализируемая PAL, носит спонтанный характер. реакция элиминации а не окислительное дезаминирование.[11]
- L-фенилаланин транс-коричная кислота + NH3

В кофактор 3,5-дигидро-5-метилдиен-4ЧАС-имидазол-4-он (MIO) участвует в реакции и располагается на положительном полюсе трех полярных спиралей в активном центре, что помогает увеличить его электрофильность.[12] MIO атакован ароматическое кольцо из L-phe, который активирует связь C-H на β-углероде для депротонирования посредством базовый остаток.[13][14] В карбанион средний этого E1cB-реакция элиминирования, который стабилизируется частичными положительными областями в активном центре, затем удаляет аммиак с образованием алкена коричной кислоты. Считается, что механизм реакции PAL аналогичен механизму родственного фермента гистидинаммиаклиазы.[13]

Долгое время считалось, что остаток дегидроаланина является ключевым электрофильным каталитическим остатком в PAL и HAL, но позже было обнаружено, что активный остаток представляет собой MIO, который является еще более электрофильным.[16][17] Он образуется в результате циклизации и дегидратации консервативного трипептидного сегмента Ala-Ser-Gly. Первым этапом образования MIO является циклизация-элиминация посредством внутримолекулярной нуклеофильной атаки азота Gly204 на карбонильную группу Ala202. Последующее отщепление воды из боковой цепи Ser203 завершает систему перекрестно сопряженных двойных связей.[15] Цифры даны для фенилаланинаммиаклиазы из Петроселин хрустящий (PDB 1W27). Хотя MIO представляет собой модификацию полипептида, было предложено назвать ее простетической группой, поскольку она имеет качество добавленного органического соединения.[8]
PAL ингибируется транс-коричной кислотой, а у некоторых видов может ингибироваться транс-производные коричной кислоты.[1][18] В неприродные аминокислоты D-Phe и D-Тырь, энантиомерный формы нормального субстрата, являются конкурентные ингибиторы.[9]
Структура
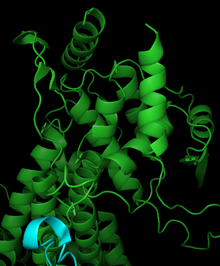
Фенилаланинаммиаклиаза состоит из четырех идентичных субъединиц, состоящих в основном из альфа-спирали, с парами мономеры формирование единого активный сайт.[17] Катализ в PAL может регулироваться дипольные моменты семи различных альфа-спиралей, связанных с активным центром.[19] Активный центр содержит электрофильную группу MIO, нековалентно связанную с тремя спиралями. Leu266, Asn270, Val269, Leu215, Lys486 и Ile472 расположены на спиралях активного сайта, а Phe413, Glu496 и Gln500 способствуют стабилизации кофактора MIO. Ориентация дипольных моментов, создаваемых спиралями в активном центре, создает электроположительную область для идеальной реакционной способности с MIO. Частично положительные области в активном центре также могут помочь стабилизировать заряд карбанионного интермедиата. PAL структурно похож на механически родственную гистидин-аммиаклиазу, хотя PAL имеет примерно 215 дополнительных остатков.[17]
Функция
Фенилаланин-аммиаклиаза может выполнять разные функции у разных видов. Он содержится в основном в некоторых растениях и грибах (например, в дрожжах). В клетках грибов и дрожжей PAL играет важную роль. катаболический роль, порождающая углерод и азот.[2] У растений это ключевой фермент биосинтеза, который катализирует первую стадию синтеза различных полифенильных соединений. [2][3] и в основном участвует в защитных механизмах. PAL участвует в 5 метаболические пути: метаболизм тирозина, метаболизм фенилаланина, метаболизм азота, биосинтез фенилпропаноидов, и биосинтез алкалоидов.
Актуальность болезни
Ферментативная заместительная терапия с использованием PAL для лечения фенилкетонурии (PKU), аутосомно-рецессивного генетического заболевания у людей, при котором мутации в фенилаланингидроксилаза (PAH, EC 1.14.16.1) ген, инактивирующий фермент, изучается.[6] Это приводит к неспособности пациента метаболизировать фенилаланин, что приводит к повышению уровня Phe в кровотоке (гиперфенилаланинемия ) и умственной отсталостью, если терапию не начали при рождении.[6]
В мае 2018 года FDA одобрило Pegvaliase, а рекомбинантный ПЭГилированный фенилаланинаммиаклиаза для лечения ФКУ, разработанная Биомарин.[20][21]
Исследование
Искусственные подсластители
Обратная реакция, катализируемая PAL, была исследована на предмет использования для преобразования транс-коричная кислота L-фенилаланин, который является предшественником подсластителя аспартама. Этот процесс был разработан Genex Corporation, но так и не получил коммерческого применения.[22]
Синтез неприродных аминокислот
Аналогично тому, как синтезируется аспартам, PAL также используется для синтеза неприродные аминокислоты из различных заменен коричная кислота в исследовательских целях.[23] Стерическое препятствие из замена арена однако ограничивает возможности PAL для этой цели.[24] Например, когда Rhodotorula glutinis был использован, чтобы повлиять на это биотрансформация было обнаружено, что фермент не переносит все параграф заместители кроме F, предположительно из-за небольшой радиус атома. Мета и орто позиции оказались более толерантными, но все же ограничены более крупными заместителями. Например, фермент активный сайт разрешенный орто метокси замена но запретил мета этокси. Другие организмы с другими версиями фермента могут быть менее ограничены таким образом.[25][26]
Структурные исследования
На конец 2007 г. 5 структуры были решены для этого класса ферментов, с PDB коды доступа 1T6J, 1T6P, 1W27, 1Y2M, и 2NYF.
Рекомендации
- ^ а б c d Camm EL, Towers G (1 мая 1973 г.). «Фенилаланинаммиаклиаза». Фитохимия. 12 (5): 961–973. Дои:10.1016/0031-9422(73)85001-0.
- ^ а б c Фриц Р.Р., Ходгинс Д.С., Абель К.В. (август 1976 г.). «Фенилаланин аммиак-лиаза. Индукция и очистка от дрожжей и клиренс у млекопитающих». Журнал биологической химии. 251 (15): 4646–50. PMID 985816.
- ^ а б Танака Ю., Мацуока М., Яманото Н., Охаши Ю., Кано-Мураками Ю., Озэки Ю. (август 1989 г.). «Структура и характеристика клона кДНК фенилаланинаммиак-лиазы из поврежденных корней сладкого картофеля». Физиология растений. 90 (4): 1403–7. Дои:10.1104 / pp.90.4.1403. ЧВК 1061903. PMID 16666943.
- ^ Appert C, Logemann E, Hahlbrock K, Schmid J, Amrhein N (октябрь 1994 г.). «Структурные и каталитические свойства четырех изоферментов фенилаланин-аммиак-лиазы из петрушки (Petroselinum crispum Nym.)». Европейский журнал биохимии. 225 (1): 491–9. Дои:10.1111 / j.1432-1033.1994.00491.x. PMID 7925471.
- ^ Hahlbrock K, Grisebach H (1 июня 1979 г.). «Ферментные средства контроля в биосинтезе лигнина и флавоноидов». Ежегодный обзор физиологии растений. 30 (1): 105–130. Дои:10.1146 / annurev.pp.30.060179.000541.
- ^ а б c Саркисян К.Н., Гамес А. (декабрь 2005 г.). «Фенилаланинаммиаклиаза, заместительная ферментная терапия фенилкетонурии, где мы сейчас?». Молекулярная генетика и метаболизм. 86 Приложение 1: S22-6. Дои:10.1016 / j.ymgme.2005.06.016. PMID 16165390.
- ^ Эванс К., Ханна К., Конрад Д., Петерсон В., Мисава М. (1 февраля 1987 г.). «Производство фенилаланинаммиаклиазы (PAL): выделение и оценка штаммов дрожжей, пригодных для коммерческого производства L-фенилаланина». Прикладная микробиология и биотехнология. 25 (5): 406–414. Дои:10.1007 / BF00253309. S2CID 40066810.
- ^ а б Шведе Т.Ф., Рети Дж., Шульц Г.Э. (апрель 1999 г.). «Кристаллическая структура гистидин-аммиак-лиазы, раскрывающая новую модификацию полипептида в качестве каталитического электрофила». Биохимия. 38 (17): 5355–61. Дои:10.1021 / bi982929q. PMID 10220322.
- ^ а б Ходгинс Д.С. (май 1971 г.). «Дрожжевой фенилаланин-аммиак-лиаза. Очистка, свойства и идентификация каталитически необходимого дегидроаланина». Журнал биологической химии. 246 (9): 2977–85. PMID 5102931.
- ^ Баррос Дж., Серрани-Ярс Дж. К., Чен Ф., Бакстер Д., Венейблс Б. Дж., Диксон Р. А. (май 2016 г.). «Роль бифункциональной аммиак-лиазы в биосинтезе клеточной стенки травы». Природа Растения. 2 (6): 16050. Дои:10.1038 / nplants.2016.50. PMID 27255834. S2CID 3462127.
- ^ Кукол Дж, Conn EE (Октябрь 1961 г.). «Метаболизм ароматических соединений у высших растений. IV. Очистка и свойства фенилаланиндезаминазы Hordeum vulgare». Журнал биологической химии. 236: 2692–8. PMID 14458851.
- ^ Алунни С., Чипичани А., Фиорони Г., Оттави Л. (апрель 2003 г.). «Механизмы ингибирования фенилаланинаммиаклиазы фенольными ингибиторами и синергическими ингибиторами фенол / глицин». Архивы биохимии и биофизики. 412 (2): 170–5. Дои:10.1016 / с0003-9861 (03) 00007-9. PMID 12667480.
- ^ а б Лангер Б, Лангер М, Рети Дж. (2001). «Метилиден-имидазолон (MIO) из гистидина и фенилаланин-аммиак-лиазы». Достижения в химии белков. 58: 175–214. Дои:10.1016 / s0065-3233 (01) 58005-5. ISBN 9780120342587. PMID 11665488.
- ^ Фрей П.А., Хегеман А.Д. (2007). «Метиленимидазолон-зависимое отщепление и присоединение: фенилаланин-аммиак-лиаза». Механизмы ферментативных реакций. Издательство Оксфордского университета. стр.460 –466. ISBN 9780195352740.
- ^ а б Риттер Х., Шульц Г.Е. (декабрь 2004 г.). «Структурная основа вступления в метаболизм фенилпропаноидов, катализируемый фенилаланинаммиаклиазой». Растительная клетка. 16 (12): 3426–36. Дои:10.1105 / tpc.104.025288. ЧВК 535883. PMID 15548745.
- ^ Рети, Янош (2003). «Открытие и роль метилиден имидазолона, высоко электрофильной простетической группы». Biochimica et Biophysica Acta (BBA) - Белки и протеомика. 1647 (1–2): 179–184. Дои:10.1016 / S1570-9639 (03) 00091-8. PMID 12686130.
- ^ а б c Calabrese JC, Jordan DB, Boodhoo A, Sariaslani S, Vannelli T (сентябрь 2004 г.). «Кристаллическая структура фенилаланин аммиаклиазы: множественные спиральные диполи, участвующие в катализе». Биохимия. 43 (36): 11403–16. Дои:10.1021 / bi049053 +. PMID 15350127.
- ^ Сато Т., Киучи Ф., Санкава У. (1 января 1982 г.). «Ингибирование фенилаланин-аммиак-лиазы производными коричной кислоты и родственными соединениями». Фитохимия. 21 (4): 845–850. Дои:10.1016/0031-9422(82)80077-0.
- ^ Пилбак С., Томин А., Рети Дж., Поппе Л. (март 2006 г.). «Существенная конформация тирозин-содержащей петли и роль C-концевой мультиспиральной области в эукариотических фенилаланин-аммиак-лиазах». Журнал FEBS. 273 (5): 1004–19. Дои:10.1111 / j.1742-4658.2006.05127.x. PMID 16478474. S2CID 33002042.
- ^ Powers M (29 мая 2018 г.). «Заключительный экзамен по биомарину: Palynziq получил одобрение FDA для лечения фенилкетонурии у взрослых». BioWorld.
- ^ Леви Х.Л., Саркисян С.Н., Стивенс Р.С., Скривер С.Р. (июнь 2018 г.). «Фенилаланин-аммиаклиаза (PAL): от открытия до ферментативной заместительной терапии фенилкетонурии». Молекулярная генетика и метаболизм. 124 (4): 223–229. Дои:10.1016 / j.ymgme.2018.06.002. PMID 29941359.
- ^ Straathof AJ, Adlercreutz P (2014). Прикладной биокатализ. CRC Press. п. 146. ISBN 9781482298420.
- ^ Хьюз А. (2009). Аминокислоты, пептиды и белки в органической химии, том 1. Weinheim Германия: Wiley VCH. п. 94. ISBN 9783527320967.
- ^ Ренар Дж., Гийе Дж., Бур С, Мальта-Валетт V, Лернер Д. (1992). «Синтез аналогов L-фенилаланина с помощью Rhodotorula glutinis. Биоконверсия производных коричной кислоты». Письма о биотехнологии. 14 (8): 673–678. Дои:10.1007 / BF01021641. S2CID 46423586.
- ^ Лавлок С.Л., Тернер Нью-Джерси (октябрь 2014 г.). «Бактериальная фенилаланин-аммиаклиаза Anabaena variabilis: биокатализатор с широкой субстратной специфичностью». Биоорганическая и медицинская химия. 22 (20): 5555–7. Дои:10.1016 / j.bmc.2014.06.035. PMID 25037641.
- ^ Сева Д., Хиробуми А. «Процесс производства производных L-фенилаланина микроорганизмами». Патенты Google. Центральная исследовательская лаборатория Хиробуми. Получено 20 июля 2014.
дальнейшее чтение
- Koukol J, Conn EE (октябрь 1961 г.). «Метаболизм ароматических соединений у высших растений. IV. Очистка и свойства фенилаланиндезаминазы Hordeum vulgare» (PDF). Журнал биологической химии. 236 (10): 2692–8. PMID 14458851.
- Молодой MR, Neish AC (1966). «Свойства аммиачных лиаз, дезаминирующих фенилаланин и родственные соединения в Triticum sestivum и Pteridium aquilinum». Фитохимия. 5 (6): 1121–1132. Дои:10.1016 / S0031-9422 (00) 86105-1.