Горгонопсия - Gorgonopsia
| Горгонопсийцы | |
|---|---|
 | |
| Скелет Иностранец виа александри на Museo delle Scienze, Тренто, Италия | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Терапсида |
| Clade: | Neotherapsida |
| Clade: | Териодонтия Сили, 1895 |
| Clade: | †Горгонопсия Lydekker, 1890 |
| Роды | |
Горгонопсия (от Древнегреческий γοργών, смысл "горгона ", и ὄψ, что означает "аспект") является вымершим клады ранних саблезубых терапсиды от Середина к Верхняя пермь. Для них характерен длинный и узкий череп, а также набухший верхний, а иногда и нижний. клыки и резцы которые, вероятно, использовались как рубящее и колющее оружие. Постклыковые зубы, как правило, уменьшены или отсутствуют. Для охоты на крупную добычу они, возможно, использовали тактику укуса и отступления, устроили засаду и нанесли изнуряющий укус цели, и следовали за ней на безопасном расстоянии, пока ее раны не исчерпали ее, после чего горгонопсий схватил животное и нанес смертельный укус. У них был бы невероятный зев, возможно, более 90 °, без необходимости расшатывать челюсть.
Со временем они заметно увеличились в размерах, от небольших черепов длиной 10-15 см (4-6 дюймов) в средней перми до нести -подобные пропорции до 60 см (2 фута) в верхней перми. Последние горгонопсийцы, Rubidgeinae, были самыми крепкий группы и могли производить особенно сильные укусы. Считается, что они полностью земной и мог ходить полустоячей походкой, с таким же наземным диапазоном движений, как и современные крокодилы. Возможно, они были более проворными, чем их жертвы, но, вероятно, были инерционными. теплокровные скорее, чем эндотермы как современный тероцефалы и цинодонты, и, следовательно, были, вероятно, сравнительно менее активными. Хотя горгонопсийцы могли поддерживать довольно высокую температуру тела, неясно, был ли у них также мех, потовые железы, или же усы. У них было больше рептилоид чем млекопитающее мозг. Большинство видов могли быть преимущественно дневной, активны в течение дня, хотя некоторые из них могли быть сумеречный или же ночной образ жизни. Считается, что они имели бинокулярное зрение, а теменный глаз, острое обоняние, функциональное вомероназальный орган («Орган Якобсона») и, возможно, рудиментарный барабанная перепонка.
Все основные группы терапевсидов произошли 265 миллионов лет назад изпеликозавр 'предок (плохо определенная группа, включающая всех синапсиды которые не являются терапсидами). Захват терапевпсида у пеликозавров произошел в средней перми, когда мир постепенно становился суше. Горгонопсии выросли, чтобы стать высшими хищниками своей среды после исчезновения диноцефалы и крупные тероцефалы после средней перми. Несмотря на существование единого континента в пермский период, Пангея, горгонопсии обнаружены только в Кару Супергруппа (в первую очередь в Южной Африке, но также в Танзании, Замбии и Малави) Формация Моради Нигера и западной части России с вероятными останками, известными из Индии. Эти места были полузасушливыми районами с сильным сезонным выпадением осадков. Таксоны горгонопсий очень мало различаются, и, следовательно, многие виды были названы на основе неубедительных и весьма изменчивых различий с момента их открытия в конце 19 века, и клады подвергались нескольким таксономическим пересмотрам.
Они вымерли во время фазы Пермско-триасовое вымирание происходящих в самом конце перми, когда большая вулканическая активность (которая привела к Сибирские ловушки ) и, как следствие, массивный всплеск парниковые газы вызвало быстрое иссушение из-за скачка температуры, кислотный дождь, частый пожары, и потенциальная поломка озоновый слой. Единственная группа терапсидов, которая выжила в этом событии, - цинодонты, которые в конечном итоге эволюционировали в млекопитающих. Большой хищный ниши будет передан архозавры (а именно крокодилов и динозавры ) в Мезозойский.
Описание

Анатомия горгонозов очень мало различается.[1] Многие виды отличаются нечеткими пропорциональными различиями, и, следовательно, более мелкие виды могут фактически представлять молодь более крупных таксонов. Примечательно, что сошник кончик морды варьируется у разных видов с точки зрения степени его расширения, а также положения, степени раскрытия и формы трех гребней.[2] У них обычно длинный и узкий череп.[3] Несовершеннолетний Rubidgea кажется, у них были морды шире, чем длинные.[4]

Мозг горгонопсии, как и другие не-млекопитающее терапсиды, отсутствует расширение неокортекс, имеет относительно большой задний мозг по сравнению с передним мозгом, большой эпифизарный нерв (встречается у существ с теменный глаз на макушке) увеличенный гипофиз и в целом удлиненной формы; в целом напоминающий рептилоид мозг.[5] В черепная коробка также был скорее рептильным, будучи сравнительно меньше и не таким толстым.[6]:280 В хлопья, часть мозжечок, пропорционально велика и связана с вестибулоокулярный рефлекс (неподвижный взгляд, двигая головой). Судя по ориентации полукруглые каналы в ухе (которое должно быть ориентировано параллельно земле), голова экземпляра горгонопсии GPIT / RE / 7124 наклонилась бы вперед примерно на 41 °, увеличивая перекрытие полей зрения двух глаз и улучшая бинокулярное зрение полезно хищнику. В отличие от рептилий или млекопитающие, полукруглые каналы плоские, вероятно, потому, что он вклинился между опистотический (ан внутреннее ухо кость) и надзатылочный кости.[5]
Как и другие пермские терапсиды, горгонопсии развили несколько характеристик млекопитающих, в том числе парасагиттальную походку (конечности были ориентированы вертикально и двигались параллельно позвоночнику) в отличие от походки распростертых животных. амфибии и более ранние синапсиды, что привело к уменьшению размера хвоста и формулы фаланги[7] (количество суставов на палец, которое было 2.3.4.5.3, как у рептилий[3]), волокнисто-пластинчатый кортикальная кость, а височное окно (дыра на височная кость ), глубоко посаженные зубы и вторичное небо (который отделяет рот от носовой полости, но у горгонопсий этого, возможно, не было).[7]

Как и многие другие млекопитающие, горгонопсии были гетеродонты, с четко определенными резцы, клыки, и постклыковые зубы гомологичный с премоляры и коренные зубы.[2] У них было 5 резцов на верхней челюсти (у большинства первые 3 были одного размера, а последние 2 были короче) и 4 на нижней.[6]:17–18 У большинства горгонопсий резцы были большими, а верхние клыки были удлинены в сабли, как и у более поздних саблезубые кошки. Некоторые горгонопсии имели исключительно длинные верхние клыки, такие как Иностранность, и у некоторых из них был фланец на нижней челюсти, чтобы защитить кончик клыка, когда рот был закрыт. Считается, что сабли использовались как колющее или рубящее оружие, которое потребовало бы чрезвычайно широкого раскрытия. И верхние, и нижние клыки Rubidgea были налиты, и животному нужно было бы еще больше разглядеть.[8] С другой стороны, постклыковые зубы уменьшились как в размере, так и в количестве; много Rubidgeines (у новейших горгонопсийцев) постклыков нижней челюсти не было,[9] и Clelandia их совсем не хватало.[4]
Горгонопсийцы были полифиодонты, и зубы постоянно росли на протяжении всей жизни человека.[10] Как и в случае с некоторыми терапсидами, когда была одна функциональная собака, другая собака росла, чтобы заменить ее, когда она неизбежно сломалась. Левая и правая стороны челюстей не обязательно должны быть синхронными, поэтому, например, 1-й клык с левой стороны может функционировать, в то время как 1-й клык с правой стороны все еще растет.[10] Такой метод, возможно, использовался для того, чтобы всегда иметь набор функциональных клыков, поскольку наличие одного клыка или отсутствие клыка могло серьезно затруднить охоту, а выращивание таких больших зубов могло занять много времени. С другой стороны, поскольку функциональный клык обычно находится в самой передней лунке зуба (а не в равной степени в любой лунке), вполне возможно, что замена клыка происходила конечное количество раз, и в конечном итоге животное оставалось с единый постоянный набор функциональных клыков на этих гнездах. В 1984 году британские палеонтологи Дорис и Кеннет Кермак предположили, что клыки росли до размеров черепа и постоянно отламывались, пока животное не переставало расти, и что горгонопсии отличались ранней версией конечного замещения зубов, наблюдаемой у многих млекопитающих. Схемы замены других зубов неясны.[8] Постклыковые зубы заменялись медленнее, чем другие зубы, вероятно, из-за отсутствия у них функциональной значимости.[2]

Ранние горгонопсиды в средней перми были довольно маленькими, с длиной черепа 10–15 см (4–6 дюймов), тогда как некоторые более поздние роды достигли массивных медвежьих размеров с черепами до 60 см (2 фута) в длину. Тем не менее, мелкие горгонопсии оставались многочисленными до исчезновения (хотя мелкие виды могут фактически представлять ювенильные особи других таксонов).[2]
В отличие от эвтериодонтов, затылочная кость (в задней части черепа) прямоугольный (шире, чем высокий) и вогнутый, в отличие от треугольного.[6]:279 7 шейные позвонки (в шее) все одного размера, кроме последнего, который короче и ниже; есть 1 атлас и 1 ось. Как и у саблезубых кошек, у них длинная шея с хорошо развитыми мускулами, что было бы особенно полезно, когда клыки были погружены в животное. Как и другие ранние синапсиды, у горгонопсийцев один затылочный мыщелок, а сочленение (суставы) шейных позвонков в целом рептилоидное, разрешающее движение головы из стороны в сторону, но ограничивающее движение вверх и вниз. Последний шейный отдел имеет форму спинные позвонки. Спинные кости имеют форму катушки и выглядят примерно одинаково. Остистые отростки круто выступают из центральной части и имеют острый киль на передней и задней сторонах. В отличие от эутериодонтов, горгонопсии не выделяют поясничных позвонков. Тем не менее, спинные мышцы, относящиеся к этой серии, похожи на поясничные части саблезубых кошек с круто ориентированными зигопофазами, полезными для стабилизации нижней части спины, особенно при удержании борющейся добычи. Есть 3 крестцовые позвонки, а серия, прикрепленная к таз по 1-му позвонку. Таз рептильный, с отделенным подвздошная кость, седалищная кость, и лобок. В бедренная кость немного s-образный, короткий, но длиннее и тоньше, чем плечевая кость. Для большинства большеберцовая кость и малоберцовая кость сильно изогнуты друг в друга, а большеберцовая кость более крепкая, чем малоберцовая.[6]:291–299 Сустав между лодыжкой и пяточной костью мог быть несколько подвижным. Пятая цифра для рук и ног не была прикреплена к запястье /предплюсна, а вместо этого подключался напрямую к локтевая кость / пяточная кость.[11]
Таксономия
Классификация

Первые останки горгонопсий были обнаружены в Beaufort Group из Кару Супергруппа в южноафриканском обнажении и были классифицированы как Горгонопс торвус в 1876 г. английский биолог Ричард Оуэн. Название Горгонопс происходит из Древнегреческий γοργών, смысл "горгона ", и ὄψ, что означает «аспект». Оуэн предположил, что этот и несколько других описанных им таксонов супергруппы были хладнокровный рептилий, но у них были зубы, напоминающие зубы хищных млекопитающих, поэтому было предложено отнести их всех к недавно придуманным порядок Териодонтия (который он поместил в учебный класс Рептилии). Он решил разделить Theriodontia на семьи основаны на анатомии ноздрей (костные ноздри) - «Mononarialia» для тех, у кого в черепе 1 отверстие для носа, как у млекопитающих, «Binarialia» для тех, у кого есть 2 отверстия, как у рептилий, и «Tectinarialia» для Горгонопс потому что его отверстие было затенено толстой костяной крышей[12] (тектус является латинский для «крытый, крытый, палубный»).[13] В 1890 году английский натуралист Ричард Лидеккер сделали Горгонопс то типовой вид семейства Gorgonopsidae.[14] В 1895 году британский палеонтолог Гарри Сили верил Горгонопс не имел отверстия в височной кости (височных отверстий), что является диагностическим признаком Theriodontia, и таким образом поднял Gorgonopsidae до Gorgonopsia, в отличие от Theriodontia. Он классифицировал все южноафриканские материалы, несущие черты рептилий и млекопитающих, в отряд «Theriosuchia», и рассмотрел Gorgonopsia и Theriodontia. подзаказы этого.[15] В 1903 году американский палеонтолог Генри Фэрфилд Осборн полностью переработана классификация рептилий и выделены 2 основные группы: Диапсида и Synapsida,[16] а в 1905 г. южноафриканский палеонтолог Роберт Брум создал 3-ю группу, Терапсида, для размещения «млекопитающих рептилий», включая Theriodontia. Он также оспорил требование Сили и понизил Горгонопс обратно в Theriodontia, но он поместил его в свою недавно построенную подгруппу Тероцефалия, растворяя горгонопсию.[17] В 1913 г., особенно в свете почти полного Г. торвус череп, обнаруженный преподобным Джоном Х. Уайтсом, Метла восстановила горгонопсию.[18]
Впервые горгонопсии были обнаружены в России в 1890-х годах в урочище Соколки на р. Река Северная Двина в Сибири под наблюдением российского палеонтолога Владимир Прохорович Амалицкий. В посмертной публикации это было описано как Иностранец виа александри, и это один из самых известных и крупных горгонопсийцев. С тех пор описано всего несколько русских родов: Православлевия, Вяткогоргон, Сухогоргон, Леогоргон, и Ночница.[19] В Африке горгонопсии также были обнаружены в обнажениях Кару в Долина Рухуху, Танзания; то Верхняя долина Луангва, Замбия; и Chiweta, Малави.[6]:7 Горгонопсии заметно отсутствуют за пределами этих двух областей.[19] В 1979 году китайский палеонтолог Ян Чжунцзянь описал китайского горгонопса "Wangwusaurus tayuensis "на основе зубов поздней перми Формация Цзиюань,[20] но в 1981 году палеонтологи Дениз Сигоньо-Рассел и Ай-Лин Сун обнаружила, что назначенный материал представляет собой случайную совокупность, из которых только 2 имеют хотя бы отдаленное сходство с горгонопсией.[21] В 2003 году индийские палеонтологи Сангхамитра Рей и Сасвати Бандьопадхьяи отнесли некоторые фрагменты черепа из поздней перми. Формация Кундарам горгонопсийцу среднего размера,[22] хотя характеристики горгонопсий также были задокументированы у некоторых тероцефалов.[19] В 2008 г. крупный и, вероятно, рубиджевый фрагмент верхней челюсти и клыка был обнаружен в поздней перми. Формация Моради в Нигере (одна из немногих низкоширотных позднепермских формаций, несущих четвероногих), и является первым свидетельством низкоширотного горгонопсиана.[23]
Число южноафриканских родов быстро росло в 20-м веке во главе с Брумом, обширная работа которого по терапсидам Кару - с начала его карьеры в стране в 1897 году до его смерти в 1951 году - привела к его описанию 57 горгонопсидов. образцы голотипа и 29 родов. Многие таксоны Брума позже будут признаны недействительными.[24] Многие другие современные исследователи создали совершенно новые виды или роды на основе единичных экземпляров.[6]:57 Следовательно, Gorgonopsia была предметом многих таксономических беспорядков и является одной из самых проблемных групп синапсидов. Поскольку анатомия черепа очень мало различается для разных таксонов, многие из них определяются на основе нечетких пропорциональных различий, включая даже хорошо известных представителей. Номинальный вид отличаются преимущественно чертами, которые, как известно, весьма изменчивы в зависимости от возраста человека, в том числе глазная орбита размер, длина морды и количество постклыковых зубов. Таким образом, возможно, что некоторые таксоны синоним друг с другом, и представляют разные стадии развития.[25]
Среди первых попыток организовать кладу предпринял британский зоолог. Дэвид Мередит Сирс Уотсон и американский палеонтолог Альфред Ромер в 1956 г., которые разделили его на 20 семейств, из которых члены 3 (Burnetiidae, Hipposauridae и Phthinosuchidae) больше не считаются горгонопсами.[4] В 1970 и 1989 годах, преимущественно рассматривая африканские таксоны, Сигоньо-Рассел опубликовал всеобъемлющий монография на горгонопсии (определяя ее как инфраотряд ), и признали только 2 семейства: Watongiidae и Gorgonopidae. Watongia был перемещен в Varanopidae в 2004 г. Она разделила Gorgonopidae на 3 подсемейства - Gorgonopsinae, Rubidgeinae и Inostranceviinae - и сократила количество родов до 23.[26] В 2002 году российский палеонтолог Михаил Феодосевич Ивахненко, рассматривая российские таксоны, вместо этого считал горгонопсию подотрядом и сгруппировал ее вместе с Диноцефалия в заказ "Горгодонтия". Он разделил Горгонопсию на надсемейства «Gorgonopioidea» (семейства Gorgonopidae, Cyonosauridae и Galesuchidae) и «Rubidgeoidea» (Rubidgeidae, Phtinosuchidae и Inostranceviidae). В 2007 году биолог Ева В. И. Гебауэр в своем всеобъемлющем обзоре горгонопсии (ее докторская диссертация) отвергла модель Ивахненко в пользу модели Сигоньо-Рассела,[6]:57 и дополнительно уменьшили количество родов до 14 помимо русских родов: Алопозавр, Ционозавр, Аэлурозавр, Sauroctonus, Сцилакогнат, Eoarctops, Горгонопс, Нджалила, Lycaenops, Арктогнат, Аэлурогнатус, Сикозавр, Clelandina, и Rubidgea.[6]:244 В целом модель Сигоньо-Рассела поддерживается, но нет единого мнения о том, какие роды могут быть отнесены к каким подсемействам.[4] В 2015 году американский палеонтолог Кристиан Ф. Каммерер и его коллеги заново описали Эрипостома (который был обозначен как неопределенный териодонт) как горгонопсий,[27] и затонул Сцилакогнат и в следующем году Eoarctops внутрь.[4]
Первый филогения (генеалогическое древо) членов Gorgonopsia было опубликовано в 2016 году американским палеонтологом Кристианом Ф. Каммерером, который специально исследовал Rubidgeinae и повторно описал как подсемейство, так и 9 видов, которые он к нему причислял (уменьшив количество с 36 видов). Каммерер также воскрес Диногоргон, Леонтозавр, Ruhuhucerberus, и Smilesaurus. Каммерер не был уверен, что Леонтозавр, Clelandina, Диногоргон, и Rubidgea все представляют один и тот же таксон или нет (для которых Диногоргон имеет приоритет ), но он решил классифицировать их всех в племя Rubidgeini ожидает дальнейшего изучения.[4] В 2018 году Каммерер и российский палеонтолог Владимир Масютин выделили новый род Ночница как самые основные известные горгонопсии, и обнаружил, что все русские таксоны (кроме Вяткогоргон, который находится в ауткладе) образуют совершенно отдельную кладу от африканских таксонов.[19] Также в 2018 году воскресили палеобиолог Ева-Мария Бендель, Каммерер и коллеги. Цинариопс.[2]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Филогения горгонопсии по Bendel et al. 2018 г.[2] |
Эволюция
Синапсиды традиционно делятся на базальные "Пеликозаврия "и производное Therapsida. Первые включают в себя хладнокровных существ с расплывчатой походкой и предположительно более низким метаболизмом, которые развились в Верхний карбон. С середины до конца 20 века американский палеонтолог Эверетт К. Олсон исследовали разнообразие синапсидов в средней перми. Сан-Анджело, Цветочный горшок, и Чикаша Образований в Северной Америке, и отметил, что разнообразие пеликозавров уменьшилось с 6 до 3 в этих формациях и что они сосуществовали с несколькими фрагментарными образцами, которые он интерпретировал как терапсиды. Затем он предположил, что сдвиг произошел во время средней перми (Вымирание Олсона ); однако классификация этих "терапсидов" и возраст образований с тех пор подверглись сомнению. Таким образом, точное время адаптивного перехода от уровня пеликозавра к уровню терапии неясно, но 7 основных групп терапии (Биармозухия, Диноцефалия, Аномодонтия, Gorgonopsia, Therocephalia и Cynodontia) существовали 265 миллионов лет назад.[28]
| Synapsida[28] | |||||||||||||||||||||||||||||||||||||||
|

Пермь постепенно становилась все суше и суше. В верхнем карбоне и нижней перми пеликозавры, кажется, цеплялись за вечно влажный угольное болото местообитания вблизи экватора (окаменелости известны в пределах 10 ° по обе стороны от палеоэкватора); дальше, примерно до 30 °, была обширная пустыня, которая простиралась до побережья, отделяя болота от регионов с умеренным климатом. К средней перми экваториальные леса перешли на сезонную влажную / засушливую систему, но болота были связаны с зонами умеренного климата через прибрежные проходы вдоль Восточной Пангеи, что позволило межконтинентальной миграции из нынешней Южной Африки на территорию современной России. . Терапсиды, по-видимому, эволюционировали в этом сезонно влажном / засушливом ландшафте, распространившись даже на зоны с умеренным климатом. На тот момент синапсиды были единственными крупными наземными животными в их среде обитания; и пеликозавры, возможно, не смогли адаптироваться к аридификации. Примерно во время вымирания пеликозавров терапсиды испытали серьезный адаптивный радиация (все хищники) продолжаются в Верхнюю Пермь.[28]
На протяжении средней перми часто гигантские диноцефалы были доминирующими животными в своих экосистемах. Они исчезают из летописи окаменелостей во время Мирян, прямо перед Капитанское массовое вымирание вызвано вулканической активностью, которая сформировала китайский Эмейшанские ловушки. Точная причина их исчезновения неясна, но их заменили горгонопсии и дицинодонты (которые начали сильно увеличиваться в размерах) и более мелких тероцефалов.[29] Рубиджи были последней и, следовательно, самой массивной и крепко сложенной группой горгонопсий.[4]
Палеобиология
Кусать

Горгонопсии, вероятно, были активными хищниками. У рубиджинов особенно сложен череп среди горгонопсийцев, сравнимый с черепами огромных хищников, которые используют свои черепа в качестве основного оружия, таких как мозазавры или несколько теропод динозавры. Менее крепкие горгонопсии с более длинными клыками и гораздо более слабым прикусом, например Smilesaurus или же Иностранность, вместо этого, вероятно, использовали свои клыки для рубящих ударов, гораздо больше похожих на саблезубых кошек. Пост собачьи Clelandina были заменены гладким гребнем в отличие от дицинодонтов, у которых ороговевший гребень, и, возможно, он преимущественно преследовал добычу, которую мог проглотить целиком. Таксоны горгонопсий действительно сосуществовали друг с другом - целых 7 одновременно - и тот факт, что некоторые рубиджины обладают постклыками, в то время как другие современные не предполагают, что они практиковали перегородка ниши и преследовали разные предметы добычи.[4]
Считалось, что удлиненные клыки играют важную роль в их тактике охоты. Горгонопсий шарнир челюсти был двойным сочленением и состоял из несколько подвижных и вращающихся костей, что позволяло им открывать рот невероятно широко - возможно, более чем на 90 ° - без необходимости расшатывать челюсть.[30] В качестве альтернативы было высказано предположение, что сабли среди любой саблезубой группы синапсидов эволюционировали в основном из-за половой отбор (как форма вязки отображать ), как у некоторых современных видов оленей, но это трудно проверить из-за отсутствия живых саблезубых хищников-синапсидов. Считается, что у саблезубых кошек таксоны длиннозубых («кортикозубых») были преследователи, в то время как таксоны с короткими зубьями ("ятаганозубые"), как полагают, были устраивать засады на хищников.[31] Среди кортикозубых кошек предполагается, что эти хищники убивали удачно нанесенным ударом по горлу после схватывания добычи, но горгонопсии, возможно, были менее точны в размещении укусов, вооруженных челюстями рептилии и расположением зубов. Вместо этого горгонопсийцы, возможно, использовали тактику укуса и отступления: горгонопсийец устраивал засаду на свою добычу и делал из нее значительный и изнурительный укус, а затем следовал за ним, когда он пытался убежать, прежде чем поддаться его ранению, после чего горгонопсийец доставил смертельный укус. Поскольку постклыки были уменьшены или полностью отсутствовали, мясо было бы силой оторвано от туши и проглочено целиком.[30]

Горгонопсии, наряду с другими ранними хищниками, а также крокодилами, в основном полагались на «кинетико-инерциальную систему» (KI) укуса добычи, в которой крыловидные кости и височные мышцы быстро зажал челюсти, используя импульс и кинетическая энергия челюстей и зубов, чтобы схватить жертву. Плотоядные млекопитающие, включая саблезубых кошек, вместо этого полагаются в основном на «систему статического давления» (SP), в которой височные и жевательные мышцы производить сильную силу укуса, чтобы убить добычу.[9] Височная мышца и жевательная мышца разделились только у млекопитающих, а у горгонопсий вместо этого была мышца, тянущаяся с нижней стороны крыша черепа, назад к чешуйчатая кость (в задней части черепа) и по скулам; часть, закрепленная щеками, стабилизировала челюстную кость и позволяла ей двигаться из стороны в сторону при смыкании, и, возможно, это было очень важно при кусании, поскольку скулы становятся сильнее в тандеме с удлинением клыков.[6]:278
Меньшие горгонопсии, такие как Ционозавр (который на самом деле может представлять молодь другого вида), имел изящные черепа и сабли и, возможно, действовал во многом как шакалы и лисы. Более крупные горгонопсии, такие как Горгонопс, имел длинные крепкие морды с сильно расклешенными щеками, которые поддерживали бы сильные крыловидные кости и мощный укус KI. Среднего размера Арктогнат имел коробчатый череп и, как следствие, мощную морду, которая позволяла бы сильные изгибающие и скручивающие движения, а также комбинацию элементов укуса KI и SP. Еще более крупные горгонопсии, такие как Арктопс, имел более короткую и выпуклую морду, как у более раннего сфенекодонта. Диметродон, и мог бы быстро зажать челюсти из-за широкой щели (что было бы необходимо, учитывая длинные клыки). Еще более крупные Rubidgeinae имели чрезвычайно мощные, крепко сложенные, укрепленные черепа с широкой мордой, щеками с сильными выступами и чрезвычайно длинными зубами; сабли Rubidgea atrox длиннее зубов Тиранозавр.[9]
В отличие от плотоядных млекопитающих, горгонопсии (и тероцефалы) имели уменьшенные или полностью отсутствующие постклыки, и челюсть, вероятно, не могла оказывать сдвигающее давление, необходимое для раздавливания кости, открытой для доступа к Костный мозг. Им это не пригодилось, поскольку синапсиды в то время не имели костного мозга.[32]
Передвижение
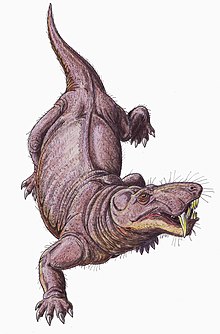
Считается, что горгонопсийцы были строго земной.[33] Считается, что горгонопсийцы могли передвигаться прямоходящей походкой, подобной той, что используют крокодилы, конечности располагаются почти вертикально, а не горизонтально, как у ящериц. В суставная полость на лопатке сильно наклонена к хвосту, поэтому конечности имели ограниченное движение вперед и, возможно, имели короткую длину шага. Ящерицы часто перемещают свои шипы из стороны в сторону, чтобы увеличить длину шага, но более вертикально ориентированные фасеточные суставы соединение позвонков у горгонопсий сделало бы позвоночник более жестким и устойчивым, затруднив такое движение.[6]:259–260
Плечевой сустав горгонопсии имеет весьма необычную конфигурацию. Головка плечевой кости, которая соединяется с плечом, длиннее суставной впадины, поэтому она не могла войти в полость. Следовательно, они могли быть прикреплены с помощью большой массы хряща, при этом плечевая кость совершала перекатывающее движение по суставному суставу. Теоретически это могло бы сделать угол между плечевой костью и гленоидом где-нибудь в диапазоне 80–145 °, если смотреть лицом к животному. Если бы угол находился на нижнем конце (то есть немного под углом к голове), это было бы довольно прочное соединение, позволяющее дельтовидные мышцы для приложения большой силы через переднюю конечность, например, когда прижимают сопротивляющуюся добычу или удерживают тушу при отрыве от плоти. Если бы плечевая кость была расположена под большим углом (под углом вниз), это могло бы позволить увеличенное разгибание вперед и назад (вдоль длинной оси) и, следовательно, большую длину шага, полезную в атаке или короткой погоне. Лопатка расширяется к бокам животного (выступает в стороны), также обеспечивая большое прикрепление дельтовидных мышц. Разработана насадка для всех лопаточно-плечевые мышцы, особенно дельтовидные мышцы. При разгибании передних конечностей дельтовидные мышцы могли приподнять переднюю сторону (передний край) плечевой кости, и коракобрахиальная мышца опущена тыльная сторона (задний край). При втягивании передней конечности грудная мышца могли сдвинуть передний край вниз, и подлопаточная мышца подтянул задний край вверх.[6]:260–264
Тазовый сустав имеет обычный шаровое соединение конфигурация. Несколько сплющенный бедренная головка теоретически мог бы вписаться в бедра в широком диапазоне углов. В 1982 году палеонтолог Том С. Кемп предположил, что ранние териодонты, в том числе горгонопсии, могли располагать бедренную кость как под горизонтальным углом при раскинувшейся походке, так и под более вертикальным углом при прямой походке. Он сравнил двигательные привычки этих существ с таковыми крокодилов, которые используют раскидистую походку на короткие расстояния, но переключаются на прямую во время бега или перемещения на большие расстояния. Хотя бедро Sauroctonus кажется анатомически промежуточным между Диметродон и млекопитающие - с подвздошная кость расширяется больше в направлении головы, чем в направлении хвоста, и лобок несколько сокращена - puboischiofemoralis (большая мышца, переносимая только рептилиями и идущая от таза к бедренной кости), широко прикрепленная к нижней стороне лобка и седалищная кость, что позволило бы ему произвести сильную приводящий сила (подтягивание ног к телу), полезная при походке с распростертыми объятиями. Также возможно, что горгонопсии в первую очередь задействовали эту мышцу во время схватки с добычей.[6]:264–270 Голени относительно короткие по сравнению с бедренной костью, что говорит о том, что горгонопсии не были хорошо приспособлены для бега на большие расстояния.[6]:298–299

Что касается того, как ступни были поставлены на землю, горгонопсии - единственные ранние терапсиды, которые проявляют эктатаксонию (последняя цифра имеет наибольший вес), гомоподию (отпечатки пальцев и ладони выглядят одинаково) и полуплантиграды (до некоторой степени ступни были поставлены на землю).[34] Эти приспособления, возможно, сделали горгонопсий более быстрыми и проворными, чем их жертвы.[30] У горгонопсийцев были довольно подвижные пальцы, что свидетельствовало о способности хватать как руками, так и ногами, возможно, для схватывания борющейся добычи, чтобы предотвратить чрезмерную нагрузку на клыки и последующие переломы или поломки клыков, когда они были погружены в жертву.[11]
Чувства

В отличие от более производных териодонтов, но, как и некоторые современные эктотермические существа, все горгонопсии обладали шишковидным глазом на макушке головы, который используется для определения дневного света (и, следовательно, оптимальной температуры для активности). Возможно, что другие териодонты потеряли это из-за эволюции либо эндотермии, либо по своей природе светочувствительные ганглиозные клетки сетчатки в глазах - в тандеме с потерей цветового зрения и переходом к ночному образу жизни - или и тем, и другим.[35] Ночной образ жизни долгое время считалось, что поведение происходит у млекопитающих (ночное узкое место ), но большой размер орбиты и наличие склеротические кольца Во многих ранних синапсидах, уходящих еще в каменноугольный период, можно предположить, что способность выходить на улицу в условиях низкой освещенности возникла намного раньше. Исходя из этих аспектов, Sauroctonus parringtoni возможно, имел мезопическое зрение, и Ционозавр скотопический или же фотопическое зрение.[36] Диаметр склеротических колец для маленьких Вяткогоргон пропорционально большие, с внутренним диаметром 1,5 см (0,59 дюйма) и внешним диаметром 2,3 см (0,91 дюйма), по сравнению с диаметром 2,8 см (1,1 дюйма) для самой орбиты, что предполагает, что они совершали преимущественно ночные экскурсии.[37] Среди горгонопсийцев рубиджин Clelandina имеет необычно маленькие склеротические кольца, что указывает на фотопическое зрение и строгое дневной; Каммерер предположил, что разделение ниш между рубиджинами (поскольку на территории сосуществуют до 7 различных таксонов) отчасти выражалось в том, что разные виды активны в разное время суток, но склеротические кольца только Clelandina среди этого подсемейства были идентифицированы, что делает эту гипотезу весьма умозрительной.[4]
Горгонопсийцы имеют довольно короткую носовую полость, как у пеликозавров, но с обильными продольными гребнями позади них. внутренние ноздри (которые соединяют носовую полость с горлом); поскольку вдыхаемый воздух не проходил бы через них, они обычно интерпретируются как обонятельные носовые раковины и дали горгонопсийцам довольно развитое обоняние.[38] Горгонопсийцы обладали вомероназальный орган («Орган Якобсона») - часть вспомогательная обонятельная система –Который был бы размещен в основании носовая перегородка; В отличие от дицинодонтов и тероцефалов, по всей видимости, существовал канал, соединяющий орган со ртом, что указывает на его функциональность у горгонопсий.[39]

Было высказано предположение, что ранние териодонты (включая горгонопсийцев) обладали барабанная перепонка, в отличие от более ранних пеликозавров, потому что связь между квадратная кость (на шарнире челюсти) и крыловидная кость (на нёбо ) начал уменьшаться, позволяя квадрату до некоторой степени независимо вибрировать. Это могло позволить обнаруживать воздушные звуки с низким амплитуда менее 1 мм (0,039 дюйма), но барабанная перепонка будет поддерживаться хрящ или же связки вместо кости. Если правильно, то задние кости (которые у ранних млекопитающих образуют среднее ухо кости), чтобы отделиться от зубной кости (челюстной кости); летопись окаменелостей горгонопсий, кажется, указывает на сокращение постдентарной и дентарной связи. Что касается горгонопсийцев, альтернативно предполагалось, что изоляция квадрата должна была вызвать стрептостилия (вращающийся квадрат) для увеличения зевки, а не для облегчения слуха.[7]
Терморегуляция
Между более ранними пеликозаврами и терапсидами произошел значительный анатомический сдвиг, который, как предполагается, был связан с увеличением метаболизм и происхождение гомеотермия (поддержание высокой температуры тела). Развитие вторичного неба и отделение рта от носовой полости может повысить эффективность вентиляции, связанную с высоким уровнем аэробный Мероприятия; Горгонопсийцы не имели костного вторичного неба, но, возможно, имели одно из мягких тканей, но даже тогда, возможно, это было в основном из-за пищевого поведения. Постулируется, что реорганизация скелета (от растягивающейся до парасагиттальной походки) свидетельствует о наличии диафрагма и, таким образом, улучшенная вентиляция для аэробной активности; но вместо этого это могло быть увеличение ускорения или ловкости, что не обязательно означает интенсивную аэробную активность, как у крокодилов. Фиброзная пластинчатая кортикальная кость, которая была у всех ранних терапсидов, может указывать на повышенную скорость роста, но это может не быть связано со скоростью метаболизма.[40]
Современные крупные рептилии естественным образом отдают тепло телу медленнее, чем более мелкие, и считаются «инерционными гомеотермами», но они поддерживают низкую температуру тела - 25–30 ° C (77–86 ° F). Если бы терапсидам требовалась более высокая температура тела - 35–40 ° C (95–104 ° F), они должны были быть либо эндотермы (генерируют тепло собственного тела) или имеют больший контроль над потерей тепла (то есть лучше гомеотермия). Парасагиттальная походка могла помочь последнему, так как она удерживала большую часть тела от земли, а также позволяла крови оставаться в брюшной полости вместо того, чтобы циркулировать через придатки, что уменьшило бы передачу тепла земле. и стабилизировать температура ядра. Уменьшенный хвост также уменьшил бы общую площадь поверхности животного, дополнительно минимизируя потерю тепла.[41] Среди терапсидов только эутериодонты (не горгонопсии) имеют респираторные заболевания. носовые раковины, которые помогают удерживать влагу при вдыхании большого количества воздуха, и их эволюция обычно связана с началом уровня потребления кислорода у «млекопитающих» и возникновением эндотермии.[38]
Если горгонопсии были инерционными гомеотермами, не исключено, что у них были волосы. Морда обычно пронизана отверстие (небольшие отверстия, которые связаны с кровеносными сосудами), которые потенциально могут указывать на наличие дряблой кожи (в отличие от чешуек), волос, различных кожные железы (Такие как потовые железы ), и усы; тем не менее, у некоторых рептилий наблюдается похожий рисунок отверстий, которые, скорее, связаны с развитием зубов, а не с кожей.[42]
Палеопатология
Переднелатеральный аспект слева радиус (кость предплечья) образца горгонопсии NHCC LB396 представляет собой округлое костное поражение с неравномерным расположением костей.радиальный шипы из кортикальной кости, окруженные тонким слоем поднадкостничная кость, который быстро рос за один вегетационный период. Это соответствует периостит скорее всего, происходит из поднадкостничного гематома. Это специфическое состояние, а также высокая скорость роста больше напоминают млекопитающих и динозавров, чем крокодилов или крокодилов. варан. Среди ранних синапсидов единственная отмеченная патология - это остеомиелит в нескольких группах пеликозавров.[43]
На губной (языковой) стороне корня зуба функционального клыка RB382 представлено до 8 поражений, сгруппированных по средней линии зуба, которые напоминают миниатюрные зубы с выступом. мякоть, дентин, и тонкий эмаль покрытие. Они примерно круглые - с диаметром, варьирующимся от 0,3–3,9 мм (0,012–0,154 дюйма), - хотя они становятся менее круглыми примерно в средней точке корня, пока не пройдут через шейка зуба. Это примерно соответствует человеческому недугу. одонтома, наиболее частый тип одонтогенная опухоль, который раньше в летописи окаменелостей простирался всего на несколько миллионов лет назад. RB382, возраст которого 255 миллионов лет, представляет собой самый старый из известных случаев одонтомы.[44]
Палеоэкология

После исчезновения диноцефалов и (в Южной Африке) базальных тероцефалов Scylacosauridae и Lycosuchidae, горгонопсии эволюционировали из мелких и необычных форм в большие высшие хищники. В средней и верхней перми в Южной Африке дицинодонты были наиболее распространенными животными, тогда как парейазавры Deltavjatia и Скутозавр были самыми многочисленными в русских формациях, содержащих горгонопсий.[45] В период верхней перми южноафриканская группа Бофорта была полузасушливой. холодная степь с большим, эфемерный (временные) реки и поймы, истощающие источники воды намного дальше на север в море Кару, с некоторыми проявлениями паводки после внезапного обильного дождя;[46][47] распределение карбонаты согласуется с современными Caliche отложения, которые образуются в климате со средней температурой 16–20 ° C (61–68 ° F) и 100–500 мм (3,9–19,7 дюйма) сезонных осадков.[47] Саларевская свита на западе России, вероятно, также образовалась в полузасушливых условиях с сильно сезонными осадками и характеризовалась гигрофит и галофит растения в прибрежных районах, а также более засухоустойчивые хвойные породы на возвышенностях.[48] Формация Моради была засушливой пустыней, в которой преобладали рептилии. Морадизавр и парейазавр Буностегос, который, вероятно, питался хвойными деревьями.[23]
Горгонопсии вымерли в конце верхней перми во время Пермско-триасовое вымирание, что было в первую очередь вызвано несколькими сериями крупной вулканической активности; исчезновение на границе перми и триаса было вызвано в первую очередь вулканизмом, который с тех пор сформировал Сибирские ловушки. В результате массивный всплеск парниковые газы вызвало быструю аридификацию из-за: скачка температуры (до 8–10 ° C на экваторе со средней экваториальной температурой 32–35 ° C (90–95 ° F) в начале триаса), кислотный дождь (с pH всего 2 или 3 во время извержения и 4 во всем мире, и последующая нехватка лесов в течение первых 10 миллионов лет триаса), частые пожары (хотя они уже были довольно распространены на всей территории Перми), и потенциальная поломка озоновый слой (возможно кратковременное увеличение УФ-излучение бомбардировка на 400% на экваторе и 5000% на полюсах).[49] Среди терапсидов крупные травоядные аномодонты сумели пересечь границу перми и триаса и выжили до Карнийский в Верхний триас, но выжили только мелкотелые виды цинодонтов. В ниши они оставили открытыми были в конечном итоге заполнены архозавры (включая крокодилов и динозавры ) на ранних стадиях Триасовый.[50]
Смотрите также
Примечания
- ^ ce, мозжечок; cnI, обонятельный нерв; cnV + vcm—тройничный нерв и медиальная вена; cnVI, отводящий нерв; cnVII, лицевой нерв; cnIX-XI, языкоглоточный и вагоакцессорные нервы; cnXII, подъязычный нерв; en, эпифизарный нерв; фб, передний мозг; fcl, хлопья; ibic, внутреннее отделение внутренняя сонная артерия; лоб, слева обонятельная луковица; ob - обонятельная луковица; от, обонятельный тракт; pg, гипофиз; pgll, боковые доли гипофиза; ПФ, мостовой изгиб; грабить, правая обонятельная луковица; vc, Видиан канал; vc = спа, видианский канал, где клиновидно-небная артерия проходит; vcd, спинная вена головного мозга
Рекомендации
- ^ Angielczyk, Kenneth D .; Каммерер, Кристиан Ф. (2018). «Синапсиды не млекопитающих: глубокие корни генеалогического древа млекопитающих». В Zachos, Frank E .; Ашер, Роберт Дж. (Ред.). Эволюция, разнообразие и систематика млекопитающих. Берлин: Де Грюйтер. ISBN 9783110275902.
- ^ а б c d е ж Бендель, Ева-Мария; Каммерер, Кристиан Ф .; Карджилов Николай; Фернандес, Винсент; Фребиш, Йорг (2018). «Черепная анатомия горгонопсии. Цинариопс робустус на основе КТ-реконструкции ». PLOS ONE. 13 (11): e0207367. Дои:10.1371 / journal.pone.0207367. ЧВК 6261584. PMID 30485338.
- ^ а б Суинтон, В. Э. (1954). Ископаемые амфибии и рептилии. Британский музей (естественная история). п. 33.
- ^ а б c d е ж грамм час я j Каммерер, Кристиан Ф. (2016). «Систематика Rubidgeinae (Therapsida: Gorgonopsia)». PeerJ. 4: e1608. Дои:10.7717 / peerj.1608. ЧВК 4730894. PMID 26823998.
- ^ а б Араухо, Рикардо; Фернандес, Винсент; Polcyn, Майкл Дж .; Fröbisch, Jörg; Мартинс, Руи М. С. (2017). «Аспекты палеобиологии и эволюции горгонопсий: выводы из базикраниума, затылка, костного лабиринта, сосудистой сети и нейроанатомии». PeerJ. 5: e3119. Дои:10.7717 / peerj.3119. ЧВК 5390774. PMID 28413721.
- ^ а б c d е ж грамм час я j k л м Гебауэр, Э. В. И. (2007). Филогения и эволюция горгонопсии с особым упоминанием черепа и скелета в GPIT / RE / 7113 (Кандидат наук). Тюбингенский университет Эберхарда-Карла.
- ^ а б c Лаурин, М. (1998). «Новые данные о черепной анатомии Lycaenops (Synapsida, Gorgonopsidae) и размышления о возможном наличии стрептостилии у горгонопсий ». Журнал палеонтологии позвоночных. 18 (4): 765–776. Дои:10.1080/02724634.1998.10011105.
- ^ а б Кермак, Д. М.; Кермак, К.А. (1984). «Зубные протезы, замена зубов и сустав челюсти». Эволюция характеров млекопитающих. С. 71–79. Дои:10.1007/978-1-4684-7817-4_5. ISBN 978-1-4684-7817-4.
- ^ а б c Van Valkenburgh, B .; Дженкинс, И. (2002). «Эволюционные закономерности в истории пермотриасовых и кайнозойских синапсидных хищников». Документы Палеонтологического общества. 8: 273–275. Дои:10.1017 / S1089332600001121.
- ^ а б Кермак, К. А. (1956). «Замена зубов у млекопитающих рептилий подотряда Gorgonopsia и Therocephalia». Философские труды Лондонского королевского общества B: Биологические науки. 240 (670): 95–133. Дои:10.1098 / рстб.1956.0013.
- ^ а б Kümmell, S. B .; Фрей, Э. (2014). «Диапазон движения в луче I мануса и песа и цепкость автоподий в ранней перми и позднемеловой неаномодонтовой синапсиде». PLOS ONE. 9 (12): e113911. Дои:10.1371 / journal.pone.0113911. ЧВК 4269487. PMID 25517726.
- ^ Оуэн, Р. (1986). Описательный и иллюстрированный каталог ископаемых рептилий Южной Африки в коллекции Британского музея. Британский музей (естественная история). С. 27–29.
- ^ тектус в Чарльтоне Т. Льюисе (1891) Элементарный латинский словарь, Харпер и братья
- ^ Лидеккер, Р. (1890). Каталог ископаемых рептилий и амфибий в Британском музее (естественная история) Часть IV.. Британский музей (естественная история). п. 111.
- ^ Сили, Х. (1895). «Исследования структуры, организации и классификации ископаемых рептилий. - Часть IX. Раздел 1. О Therosuchia». Летопись и журнал естественной истории. 13 (6): 375. Дои:10.1080/00222939408677718.
- ^ Осборн, Х.Ф. (1903). "Подклассы рептилий Diapsida и Synapsida и ранняя история диаптозаврии. Том I. Часть VIII" (PDF): 451–466. Цитировать журнал требует
| журнал =(помощь) - ^ Веник, Р. (1905). «Об употреблении термина аномодонт». Записи музея Олбани. 1: 266–269.
- ^ Веник, Р. (1913). "О горгонопсии, подотряде млекопитающих рептилий". Труды Лондонского зоологического общества. 1913 (2): 225–230. Дои:10.1111 / j.1469-7998.1913.tb07574.x.
- ^ а б c d Каммерер, К. Ф .; Масютин, В. (2018). «Терапсиды горгонопсийские (Ночница ген. ноя и Вяткогоргон) из Пермского урочища Котельнич России ». PeerJ. 6: e4954. Дои:10.7717 / peerj.4954. ЧВК 5995105. PMID 29900078.
- ^ Янг, К. С. (1979). «Поздняя пермская фауна из Цзиюаня, Хэнань». Позвоночные PalAsiatica. 17 (2): 99–113.
- ^ Сигоньо-Рассел, Д.; Вс, А.-Л. (1981). «Краткий обзор китайских синапсидов». Geobios. 14 (2): 276. Дои:10.1016 / S0016-6995 (81) 80012-5.
- ^ «Позднепермское сообщество позвоночных в долине Пранхита-Годавари, Индия». Цитировать журнал требует
| журнал =(помощь) - ^ а б Смайли, Т. М .; Sidor, C.A .; Мага, А .; Иде, О. (2008). «Фауна позвоночных верхней перми Нигера. VI. Первое свидетельство горгонопсийного терапсида». Журнал палеонтологии позвоночных. 28 (2): 543–547. Дои:10.1671 / 0272-4634 (2008) 28 [543: TVFOTU] 2.0.CO; 2.
- ^ Уилли, А. (2003). "Обзор терапевтических голотипов Роберта Брума: выдержали ли они испытание временем?" (PDF). Palaeontologia Africana. 39 (39): 2.
- ^ Каммерер, К. Ф. (2013). «Териодонтия: Введение». Ранняя эволюционная история синапсиды. Палеобиология и палеоантропология позвоночных. Springer. С. 165–169. Дои:10.1007/978-94-007-6841-3_10. ISBN 978-94-007-6841-3.
- ^ Сигоньо-Рассел, Дениз (1989). Веллнхофер, Питер (ред.). Theriodontia I: Phthinosuchia, Biarmosuchia, Eotitanosuchia, Gorgonopsia.. Энциклопедия палеогерпетологии. 17 B / I. Штутгарт: Густав Фишер Верлаг. ISBN 978-3437304873.
- ^ Каммерер, К. Ф .; Smith, R.MH .; Day, M.O .; Рубидж, Б. С. (2015). «Новые сведения о морфологии и стратиграфическом диапазоне среднепермского горгонопса Eriphostoma microdon Broom, 1911». Статьи по палеонтологии. 1 (2): 201–221. Дои:10.1002 / spp2.1012.
- ^ а б c Кемп, Т. С. (2006). «Происхождение и раннее излучение терапевпсидных рептилий, подобных млекопитающим: палеобиологическая гипотеза». Журнал эволюционной биологии. 19 (4): 1231–1247. Дои:10.1111 / j.1420-9101.2005.01076.x. PMID 16780524. S2CID 3184629.
- ^ Day, M.O .; Ramezani, J .; Bowring, S.A .; и другие. (2015). «Когда и как произошло массовое вымирание наземных животных в середине перми? Свидетельства из записи четвероногих в бассейне Кару, Южная Африка». Труды Королевского общества B. 282 (1811): 20150834. Дои:10.1098 / rspb.2015.0834. ЧВК 4528552. PMID 26156768.
- ^ а б c Антон, Маурисио (2013). Саблезубый. Блумингтон, Индиана: Университет Индианы Press. С. 204–207. ISBN 9780253010421.
- ^ Randau, M .; Carbone, C .; Турвей, С. Т. (2013). "Эволюция собак у саблезубых хищников: естественный отбор или половой отбор?". PLOS ONE. 8 (8): e72868. Дои:10.1371 / journal.pone.0072868. ЧВК 3738559. PMID 23951334.
- ^ Крукшанк, А. Р. И. (1973). «Образ жизни горгонопсийцев» (PDF). Palaeontologica Africana. 15 (1–2): 65–67.
- ^ Kriloff, A .; Germain, D .; Canoville, A .; Vincent, P .; Sache, M .; Лаурин, М. (2008). «Эволюция костной микроанатомии большеберцовой кости четвероногих и ее использование в палеобиологических выводах». Журнал эволюционной биологии. 21 (3): 807–826. Дои:10.1111 / j.1420-9101.2008.01512.x. PMID 18312321.
- ^ Marchetti, L .; Klein, H .; Buchwitz, M .; и другие. (2019). «Следы пермско-триасовых позвоночных из Южной Африки: ихнотаксономия, продуценты и биостратиграфия через два основных фаунистических кризиса». Исследования Гондваны. 72: 159. Дои:10.1016 / j.gr.2019.03.009.
- ^ Benoit, J .; Abdala, F .; Manger, P. R .; Рубидж, Б. С. (2016). «Шестое чувство у предшественников млекопитающих: изменчивость теменного отверстия и эволюция шишковидной железы у южноафриканских пермотриасовых эутериодонтовых терапсид». Acta Palaeontologica Polonica. 61 (4): 777–789. Дои:10.4202 / app.00219.2015. S2CID 59143925.
- ^ Angielczyk, K. D .; Шмитц, Л. (2014). «Ночной образ жизни в синапсидах предшествует возникновению млекопитающих более чем на 100 миллионов лет». Труды Королевского общества B. 281 (1793). Дои:10.1098 / rspb.2014.1642. ЧВК 4173690. PMID 25186003.
- ^ Каммерер, Кристиан Ф .; Масютин, Владимир (2018). «Терапсиды горгонопсийские (Ночница ген. ноя и Вяткогоргон) из Пермского урочища Котельнич России ». PeerJ. 6: e4954. Дои:10.7717 / peerj.4954. ЧВК 5995105. PMID 29900078.
- ^ а б Хиллениус, В. Дж. (1992). Носовые раковины и эволюция эндотермии млекопитающих (Кандидат наук). Государственный университет Орегона. С. 74–76. HDL:1957/35986.
- ^ Хиллениус, В. Дж. (2000). «Септомаксилла синапсидов не млекопитающих: корреляты мягких тканей и новая функциональная интерпретация». Журнал морфологии. 245 (1): 29–50. Дои:10.1002 / 1097-4687 (200007) 245: 1 <29 :: AID-JMOR3> 3.0.CO; 2-B.
- ^ Кемп, Т. С. (2006). «Происхождение эндотермии млекопитающих: парадигма эволюции сложной биологической структуры». Зоологический журнал Линнеевского общества. 147 (4): 473–488. Дои:10.1111 / j.1096-3642.2006.00226.x.
- ^ Тернер, Дж. С .; Трейси, К. Р. (1986). «Размер тела, гомеотермия и контроль температуры тела у млекопитающих рептилий». Экология и биология млекопитающих рептилий (PDF). Пресса Смитсоновского института.
- ^ Ван Велан, Л. (1960). «Терапсиды как млекопитающие». Эволюция. 14 (3): 304–313. Дои:10.2307/2405973. JSTOR 2405973.
- ^ Като, К. М .; Rega, E. A .; Sidor, C.A .; Хаттенлокер, А. К. (2020). «Исследование поражения костей у горгонопсиса (Synapsida) из пермского периода в Замбии и периостальных реакций у ископаемых четвероногих не млекопитающих». Философские труды Королевского общества B. 375 (1793). Дои:10.1098 / rstb.2019.0144. ЧВК 7017433. PMID 31928188.
- ^ Whitney, M. R .; Mose, L .; Сидор, К. А. (2017). «Одонтома у млекопитающего, которому 255 миллионов лет». JAMA Онкология. 3 (7): 998–1000. Дои:10.1001 / jamaoncol.2016.5417. ЧВК 5824274. PMID 27930769.
- ^ Каммерер, Кристиан Ф .; Масютин, Владимир (2018). "Новый тероцефал (Gorynychus masyutinae ген. et sp. nov.) из п. Пермь Котельнич, Кировская область, Россия ". PeerJ. 6: e4933. Дои:10.7717 / peerj.4933. ЧВК 5995100. PMID 29900076.
- ^ Wilson, A .; Flint, S .; Payenberg, T .; Тохвер, Э. (2014). «Архитектурные стили и седиментология речной группы Нижнего Бофорта, бассейн Кару, Южная Африка». Журнал осадочных исследований. 84 (4): 326–348. Дои:10.2110 / jsr.2014.28.
- ^ а б Смит, Р. М. Х. (1987). «Морфология и история осадконакопления эксгумированных пермских отмелей на юго-западе Кару, Южная Африка». Журнал осадочных исследований. 57 (1): 19–20. Дои:10.1306 / 212F8A8F-2B24-11D7-8648000102C1865D.
- ^ Якименко, Э .; Иноземцев, С. А .; Наугольных, С.В. «Палеопочвы верхней перми (саларевская свита) центральной части Русской платформы: палеоэкология и палеосреда». 21 (1): 110–119. Цитировать журнал требует
| журнал =(помощь) - ^ Бентон, М. Дж. (2018). «Гипертермические массовые вымирания: модели убийства во время пермско-триасового массового вымирания». Философские труды Королевского общества A. 376 (2130). Дои:10.1098 / rsta.2017.0076. ЧВК 6127390. PMID 30177561.
- ^ Sookias, R. B .; Батлер, Р. Дж .; Бенсон, Р. Б. Дж. (2012). «Рост динозавров показывает, что основные изменения размеров тела вызваны пассивными процессами эволюции черт». Труды Королевского общества B. 279 (1736): 2180–2187. Дои:10.1098 / rspb.2011.2441. PMID 22298850. S2CID 1231897.
дальнейшее чтение
- Баккер, Р. (1986), Ереси динозавров, Kensington Publishing Corp.
- Кокс, Б. и Сэвидж, Р.Дж.Г. и Гардинер, Б., Харрисон, К., Палмер, Д. (1988) Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 2-е издание, Marshall Publishing.
- Фентон, К.Л. и Фентон, М.А. (1958) Книга окаменелостей, Doubleday Publishing.
- Хор, П. (2005), Эпоха динозавров, Выпуск № 18. Национальный музей динозавров.
- Уорд, П. (2004), Горгона, Пингвин викинг.
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||






