Дендритный шип - Dendritic spine
| Дендритный шип | |
|---|---|
 Колючий дендрит полосатый средний шиповатый нейрон. | |
 Распространенные типы дендритных шипов. | |
| Подробности | |
| Идентификаторы | |
| латинский | gemmula dendritica |
| MeSH | D049229 |
| TH | H2.00.06.1.00036 |
| Анатомические термины микроанатомии | |
А дендритный позвоночник (или позвоночник) - это небольшой перепончатый выступ нейрона дендрит который обычно получает ввод от одного аксон на синапс. Дендритные шипы служат местом хранения синаптической силы и помогают передавать электрические сигналы телу клетки нейрона. У большинства колючек есть выпуклая головка (головка позвоночника) и тонкая шейка, которая соединяет головку позвоночника с стержнем дендрита. Дендриты одного нейрона могут содержать от сотен до тысяч шипов. Помимо шипов, обеспечивающих анатомический субстрат для хранения памяти и синаптической передачи, они также могут служить для увеличения количества возможных контактов между нейронами.[1] Также было высказано предположение, что изменения в активности нейронов положительно влияют на морфологию позвоночника.[2]
Структура
Дендритные шипы небольшие с объемом головки шипа 0,01 мкм.3 до 0,8 мкм3. Шипы с сильными синаптическими контактами обычно имеют большую головку шипа, которая соединяется с дендритом через перепончатую шейку. Наиболее заметными классами формы позвоночника являются «тонкий», «короткий», «грибовидный» и «разветвленный». Электронная микроскопия исследования показали, что между этими категориями существует континуум форм. Считается, что переменная форма и объем позвоночника коррелируют с силой и зрелостью каждого синапса позвоночника.
Распределение
Дендритные шипы обычно получают возбуждающий вклад от аксонов, хотя иногда и тормозные, и возбуждающие связи связаны с одной и той же головкой позвоночника. Близость возбуждающего аксона к дендритным шипам не достаточна для прогнозирования наличия синапса, что продемонстрировано Лихтман lab в 2015 году.[3]
Колючки встречаются на дендриты из самых главных нейроны в мозгу, включая пирамидные нейроны из неокортекс, то средние шиповатые нейроны из полосатое тело, а Клетки Пуркинье из мозжечок. Дендритные шипы встречаются с плотностью до 5 шипов / 1. мкм участок дендрита. Гиппокамп и корковый пирамидные нейроны могут получать десятки тысяч в основном возбуждающих сигналов от других нейроны на их одинаково многочисленные шипы, тогда как количество шипов на дендритах нейронов Пуркинье на порядок больше.
Цитоскелет и органеллы
Цитоскелет дендритных шипов особенно важен с точки зрения их синаптической пластичности; без динамического цитоскелета шипы не смогли бы быстро менять свой объем или форму в ответ на стимулы. Эти изменения формы могут повлиять на электрические свойства позвоночника. Цитоскелет дендритных шипов в основном состоит из нитчатого актина (F-актин ). тубулин Мономеры и белки, связанные с микротрубочками (MAP) присутствуют и организованы микротрубочки присутствуют.[4] Поскольку шипы имеют цитоскелет, состоящий в основном из актина, это позволяет им быть очень динамичными по форме и размеру. Актиновый цитоскелет напрямую определяет морфологию позвоночника, а регуляторы актина - мелкие. GTPases Такие как Rac, RhoA, и CDC42, быстро модифицируют этот цитоскелет. Сверхактивный Rac1 приводит к постоянно меньшему размеру дендритных шипов.
В дополнение к своей электрофизиологической активности и активности, опосредованной рецепторами, шипы, по-видимому, являются везикулярно активными и могут даже передавать белки. Сложенные диски гладкая эндоплазматическая сеть (SER) были идентифицированы в дендритных шипах. Формирование этого "аппарат позвоночника "зависит от белка синаптоподин и считается, что он играет важную роль в обработке кальция. "Гладкий" пузырьки также были обнаружены в шипах, поддерживающих везикулярную активность в дендритных шипах. Наличие полирибосомы in spines также предполагает трансляционную активность белков в самом позвоночнике, а не только в дендрите.
Физиология
Рецепторная активность
Дендритные шипы экспрессируют рецепторы глутамата (например. Рецептор AMPA и Рецептор NMDA ) на их поверхности. В TrkB рецептор для BDNF также экспрессируется на поверхности позвоночника и, как полагают, играет роль в выживании позвоночника. На кончике позвоночника находится электронно-плотная область, называемая "постсинаптическая плотность "(PSD). PSD непосредственно назначает активная зона его синапсирующего аксона и составляет ~ 10% площади поверхности мембраны позвоночника; нейротрансмиттеры, высвобождаемые из активной зоны, связывают рецепторы в постсинаптической плотности позвоночника. Половина синапсирующих аксонов и дендритных шипов физически связана кальций -зависимый кадгерин, который формирует межклеточные соединения между двумя нейронами.
Рецепторы глутамата (GluR) локализованы в постсинаптической плотности и прикреплены элементами цитоскелета к мембране. Они расположены непосредственно над своим сигнальным механизмом, который обычно привязан к нижней стороне плазматической мембраны, позволяя передавать сигналы GluR в цитозоль для дальнейшего распространения ближайшими сигнальными элементами для активации каскады передачи сигналов. Локализация сигнальных элементов в их GluR особенно важна для обеспечения активации сигнального каскада, поскольку GluRs неспособны влиять на определенные нижестоящие эффекты без соседних сигнализаторов.
Передача сигналов от GluR опосредуется наличием большого количества белков, особенно киназ, которые локализованы в постсинаптической плотности. К ним относятся кальций -зависимый кальмодулин, CaMKII (кальмодулин-зависимая протеинкиназа II), PKC (Протеинкиназа C), PKA (Протеинкиназа А), Протеиновая фосфатаза-1 (ПП-1) и Fyn тирозинкиназа. Некоторые сигнализаторы, такие как CaMKII, активируются в ответ на активность.
Шипы особенно полезны для нейронов, разделяя биохимические сигналы на части. Это может помочь кодировать изменения в состоянии отдельного синапса, не обязательно влияя на состояние других синапсов того же нейрона. Длина и ширина шейки позвоночника имеют большое влияние на степень разделения, при этом тонкие иглы являются наиболее биохимически изолированными иглами.
Пластичность
Дендритные шипы очень «пластичны», то есть шипы значительно меняют форму, объем и количество за небольшие промежутки времени. Поскольку шипы имеют в первую очередь актин цитоскелет, они динамичны, и большинство шипов меняют свою форму за секунды или минуты из-за динамичности ремоделирование актина. Кроме того, количество шипов очень изменчиво, и шипы приходят и уходят; в течение нескольких часов на пирамидных клетках коры головного мозга могут спонтанно появляться или исчезать 10-20% шипов, хотя шипы более крупной «грибовидной» формы являются наиболее устойчивыми.
Поддержание и пластичность позвоночника зависят от активности[5] и независимый от деятельности. BDNF частично определяет уровни позвоночника,[6] и низкий уровень Рецептор AMPA активность необходима для поддержания выживания позвоночника, а синаптическая активность, связанная с Рецепторы NMDA способствует росту позвоночника. Более того, двухфотонная лазерная сканирующая микроскопия и конфокальная микроскопия показали, что объем позвоночника изменяется в зависимости от типов стимулов, которые предъявляются к синапсу.
Важность обучения и памяти
Доказательства важности
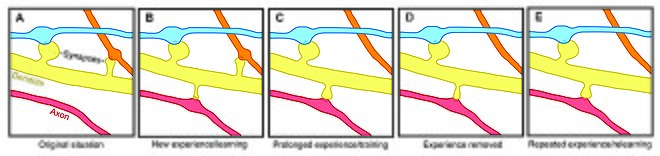
Пластичность позвоночника влияет на мотивация, учусь, и объем памяти.[7][8][9] Особенно, Долгосрочная память частично опосредуется ростом новых дендритных шипов (или увеличением ранее существовавших шипов) для усиления определенного нервного пути. Поскольку дендритные шипы представляют собой пластиковые структуры, на срок службы которых влияет входная активность,[10] динамика позвоночника может играть важную роль в поддержании памяти на протяжении всей жизни.
Возрастные изменения скорости обновления позвоночника позволяют предположить, что стабильность позвоночника влияет на обучение в процессе развития. В молодости обновление дендритных шипов относительно велико и приводит к чистой потере шипов.[1][11][12] Эта высокая скорость обновления позвоночника может характеризовать критические периоды развития и отражать способность к обучению в подростковом возрасте - разные области коры демонстрируют разные уровни синаптического обмена во время развития, возможно, отражая разные критические периоды для определенных областей мозга.[8][11] Однако в зрелом возрасте большинство шипов остаются устойчивыми, и период полураспада шипов увеличивается.[1] Эта стабилизация происходит из-за регулируемого в процессе развития замедления удаления позвоночника, процесса, который может лежать в основе стабилизации воспоминаний в зрелом возрасте.[1][11]
Вызванные опытом изменения стабильности дендритного позвоночника также указывают на оборот позвоночника как на механизм, участвующий в поддержании долговременных воспоминаний, хотя неясно, как сенсорный опыт влияет на нейронные цепи. Две общие модели могут описывать влияние опыта на структурную пластичность. С одной стороны, опыт и активность могут управлять дискретным формированием соответствующих синаптических связей, которые хранят значимую информацию для обучения. С другой стороны, синаптические связи могут быть сформированы в избытке, а опыт и активность могут привести к сокращению посторонних синаптических связей.[1]
У лабораторных животных всех возрастов обогащение окружающей среды связано с ветвлением дендритов, плотностью шипов и общим числом синапсов.[1] Кроме того, было показано, что тренировка навыков приводит к формированию и стабилизации новых шипов, дестабилизируя старые шипы,[7][13] предполагая, что обучение новому навыку включает в себя процесс перепрограммирования нейронных цепей. Поскольку степень ремоделирования позвоночника коррелирует с успехом обучения, это предполагает решающую роль синаптической структурной пластичности в формировании памяти.[13] Кроме того, изменения в стабильности и укреплении позвоночника происходят быстро и наблюдаются в течение нескольких часов после тренировки.[7][8]
И наоборот, хотя обогащение и тренировка связаны с улучшением формирования и стабильности позвоночника, в долгосрочной перспективе сенсорная депривация приводит к увеличению скорости удаления позвоночника[1][11] и, следовательно, влияет на долговременные нейронные схемы. После восстановления сенсорного восприятия после депривации в подростковом возрасте удаление позвоночника ускоряется, что позволяет предположить, что опыт играет важную роль в чистой потере позвоночника во время развития.[11] Кроме того, было показано, что другие парадигмы сенсорной депривации, такие как обрезка усов, увеличивают стабильность новых шипов.[14]
Исследования неврологических заболеваний и травм пролили дополнительный свет на природу и важность обновления позвоночника. После Инсульт, заметное увеличение структурной пластичности происходит около места травмы, и наблюдается пяти-восьмикратное увеличение скорости вращения позвоночника по сравнению с контролем.[15] Дендриты быстро распадаются и собираются заново во время ишемия - как и при инсульте, у выживших наблюдалось увеличение оборота дендритных позвонков.[16] В то время как чистая потеря шипов наблюдается у Болезнь Альцгеймера и случаи Интеллектуальная недееспособность, употребление кокаина и амфетамина было связано с увеличением ветвления дендритов и плотности шипов в префронтальная кора и прилежащее ядро.[17] Поскольку значительные изменения плотности позвоночника происходят при различных заболеваниях головного мозга, это предполагает сбалансированное состояние динамики позвоночника в нормальных обстоятельствах, которое может быть восприимчивым к нарушению равновесия при различных патологических состояниях.[17]
Есть также некоторые свидетельства потери дендритных шипов в результате старения. Одно исследование с использованием мышей отметило корреляцию между возрастным снижением плотности позвоночника в гиппокампе и возрастным снижением обучения и памяти гиппокампа.[18]
Важность оспаривается
Несмотря на экспериментальные данные которые указуют на роль дендритных динамики позвоночника в опосредовании обучения и памяти, степень важности остается структурной Пластичностью спорно. Например, исследования показывают, что только небольшая часть позвоночника, сформированная во время тренировки, действительно способствует обучению на протяжении всей жизни.[13] Кроме того, образование новых шипов может не вносить значительного вклада в связность мозга, а образование шипов может не оказывать такого большого влияния на сохранение памяти, как другие свойства структурной пластичности, такие как увеличение размеров головок шипов.[19]
Моделирование
Теоретики на протяжении десятилетий выдвигали гипотезы о потенциальной электрической функции шипов, но наша неспособность исследовать их электрические свойства до недавнего времени не позволяла теоретической работе продвинуться слишком далеко. Последние достижения в области методов визуализации наряду с более широким использованием двухфотонного извлечения глутамата привели к множеству новых открытий; теперь мы подозреваем, что есть напряжение-зависимый натрий,[20] калий,[21] и кальций[22] каналы в головках позвоночника.[23]
Теория кабеля обеспечивает теоретическую основу наиболее "простого" метода моделирования потока электрических токов по пассивным нервным волокнам. Каждый позвоночник можно рассматривать как два отдела, один из которых представляет шею, а другой - голову. Отделение, представляющее только головку позвоночника, должно нести активные свойства.
Модель континуума Бэра и Ринзеля
Чтобы облегчить анализ взаимодействий между многими шипами, Баер и Ринзель сформулировали новую теорию кабеля, согласно которой распределение шипов рассматривается как континуум.[24] В этом представлении напряжение на головке позвоночника - это среднее локальное пространственное значение мембранного потенциала в соседних отростках. Формулировка сохраняет свойство, заключающееся в отсутствии прямой электрической связи между соседними шипами; Распространение напряжения по дендритам - единственный способ взаимодействия шипов.
Модель шип-диффуз-шип
Модель SDS была задумана как упрощенная в вычислительном отношении версия полной модели Бэра и Ринзеля.[25] Он был разработан так, чтобы быть аналитически управляемым и иметь как можно меньше свободных параметров, сохраняя при этом наиболее важные, такие как сопротивление шеи позвоночника. Модель отказывается от приближения континуума и вместо этого использует пассивный дендрит, связанный с возбудимыми шипами в дискретных точках. Мембранная динамика позвоночника моделируется с помощью интегральных и огненных процессов. Спайковые события моделируются дискретным образом с формой волны, традиционно представляемой в виде прямоугольной функции.
Моделирование переходных процессов кальция в позвоночнике
Переходные процессы кальция в шипах являются ключевым триггером синаптической пластичности.[26] Рецепторы NMDA, которые обладают высокой проницаемостью для кальция, проводят ионы только в том случае, если мембранный потенциал достаточно деполяризован. Количество кальция, поступающего в позвоночник во время синаптической активности, поэтому зависит от деполяризации головки позвоночника. Данные экспериментов по визуализации кальция (двухфотонная микроскопия ) и из компартментное моделирование указывает на то, что шипы с высоким сопротивлением шеи испытывают большие переходные процессы кальция во время синаптической активности.[23][27]
Разработка
Дендритные шипы могут развиваться непосредственно из дендритных стержней или из дендритные филоподии.[28] В течение синаптогенез дендриты быстро прорастают и втягивают филоподии, небольшие мембранные органеллы без мембранных выступов. Недавно было обнаружено, что белок MIM I-BAR вносит вклад в процесс инициации.[29] В течение первой недели рождения в мозгу преобладают филоподии, которые со временем развивают синапсы. Однако после этой первой недели филоподии заменяются колючими дендритами, но также небольшими, короткими шипами, которые выступают из колючих дендритов. При развитии определенных филоподий в шипы филоподии рекрутируют пресинаптический контакт с дендритом, что стимулирует образование шипов для управления специализированным постсинаптическим контактом с пресинаптическими выступами.
Однако после образования колючки требуют созревания. Незрелые шипы обладают нарушенными способностями к передаче сигналов и обычно лишены «голов» (или имеют очень маленькие головы), только шеи, тогда как зрелые шипы поддерживают и головы, и шеи.
Клиническое значение
Когнитивные расстройства, такие как СДВГ, Болезнь Альцгеймера, аутизм, Интеллектуальная недееспособность, и синдром ломкой Х-хромосомы, может быть результатом аномалии дендритных шипов, особенно количества шипов и их зрелости.[30][31] Соотношение зрелых и незрелых шипов важно в их передаче сигналов, поскольку незрелые шипы имеют нарушенную синаптическую передачу сигналов. Синдром ломкой Х-хромосомы характеризуется избытком незрелых шипов, которые имеют несколько филоподий в корковых дендритах.
История
Дендритные шипы были впервые описаны в конце XIX века. Сантьяго Рамон-и-Кахаль на нейроны мозжечка.[32] Затем Рамон-и-Кахаль предположил, что дендритные шипы могут служить сайтами контакта между нейронами. Это было продемонстрировано более 50 лет спустя благодаря появлению электронной микроскопии.[33] До появления конфокальной микроскопии живых тканей обычно считалось, что шипы формируются во время эмбрионального развития, а затем остаются стабильными после рождения. В этой парадигме вариации синаптического веса считались достаточными для объяснения процессов памяти на клеточном уровне. Но примерно десять лет назад новые методы конфокальной микроскопии продемонстрировали, что дендритные шипы действительно являются подвижными и динамическими структурами, которые постоянно обновляются даже после рождения.[34][35][28]
Рекомендации
- ^ а б c d е ж грамм Альварес, В .; Сабатини, Б. (2007). «Анатомо-физиологическая пластичность дендритных шипов». Ежегодный обзор нейробиологии. 30: 79–97. Дои:10.1146 / annurev.neuro.30.051606.094222. PMID 17280523.
- ^ Такенберг, К., Гори, А., и Брандт, Р. (2009). Тонкий, короткий или грибовидный: патология позвоночника при болезни Альцгеймера. Текущее исследование болезни Альцгеймера, 6(3), 261-268. DOI: http: //dx.doi.org.libaccess.lib.mcmaster.ca/10.2174/156720509788486554
- ^ Kasthuri N, Hayworth KJ, Berger DR, Schalek RL, Conchello JA, Knowles-Barley S, Lee D, Vázquez-Reina A, Kaynig V, Jones TR, Roberts M, Morgan JL, Tapia JC, Seung HS, Roncal WG, Vogelstein JT, Бернс Р., Сассман Д.Л., Прибе CE, Пфистер Х., Лихтман Дж. В. (2015). «Насыщенная реконструкция объема неокортекса». Клетка. 162 (3): 648–661. Дои:10.1016 / j.cell.2015.06.054. ISSN 0092-8674. PMID 26232230.
- ^ Капитеин, 2010 г.
- ^ Де Ру, М .; Klauser, P .; Mendez, P .; Poglia, L .; Мюллер, Д. (2007). «Зависимое от активности формирование PSD и стабилизация вновь образованных шипов в культурах среза гиппокампа». Кора головного мозга. 18 (1): 151–161. Дои:10.1093 / cercor / bhm041. ISSN 1047-3211. PMID 17517683.
- ^ Канеко М .; Xie Y .; JJ .; Страйкер М.П .; Сюй Б. (2012). «Дендритный синтез BDNF необходим для поздней фазы созревания позвоночника и восстановления корковых реакций после сенсорной депривации». J. Neurosci. 32 (14): 4790–4802. Дои:10.1523 / JNEUROSCI.4462-11.2012. ЧВК 3356781. PMID 22492034.
- ^ а б c Xu, T .; Yu, X .; Perlik, A.J .; Tobin, W. F .; Zweig, J. A .; Tennant, K .; Jones, T .; Цзо, Ю. (2009). «Быстрое формирование и избирательная стабилизация синапсов для устойчивых моторных воспоминаний». Природа. 462 (7275): 915–919. Дои:10.1038 / природа08389. ЧВК 2844762. PMID 19946267.
- ^ а б c Робертс, Т .; Tschida, K .; Klein, M .; Муни, Р. (2010). «Быстрая стабилизация позвоночника и усиление синапсов в начале поведенческого обучения». Природа. 463 (7283): 948–952. Дои:10.1038 / природа08759. ЧВК 2918377. PMID 20164928.
- ^ Tschida, K. A .; Муни, Р. (2012). «Оглушение вызывает специфические для клеточного типа изменения в дендритных шипах в сенсомоторном ядре, важном для выученной вокализации». Нейрон. 73 (5): 1028–1039. Дои:10.1016 / j.neuron.2011.12.038. ЧВК 3299981. PMID 22405211.
- ^ Де Ру, М .; Klauser, P .; Мюллер, Д. (2008). «LTP способствует селективной долгосрочной стабилизации и кластеризации дендритных шипов». PLoS Biol. 6 (9): e219. Дои:10.1371 / journal.pbio.0060219. ЧВК 2531136. PMID 18788894.
- ^ а б c d е Zuo, Y .; Lin, A .; Chang, P .; Ган, В. Б. (2005). «Развитие долговременной стабильности дендритного позвоночника в различных областях коры головного мозга». Нейрон. 46 (2): 181–189. Дои:10.1016 / j.neuron.2005.04.001. PMID 15848798.
- ^ Holtmaat, A.J .; Trachtenberg, J. T .; Wilbrecht, L .; Шеперд, Г. М .; Чжан, X .; и другие. (2005). «Временные и стойкие дендритные шипы в неокортексе in vivo». Нейрон. 45 (2): 279–291. Дои:10.1016 / j.neuron.2005.01.003. PMID 15664179.
- ^ а б c Ян, G .; Pan, F .; Ган, В. Б. (2009). «Стабильно поддерживаемые дендритные шипы связаны с воспоминаниями на всю жизнь». Природа. 462 (7275): 920–924. Дои:10.1038 / природа08577. ЧВК 4724802. PMID 19946265.
- ^ Holtmaat, A .; Wilbrecht, L .; Knott, G.W .; Welker, E .; Свобода, К. (2006). «Зависящий от опыта и специфичный для клеточного типа рост позвоночника в неокортексе». Природа. 441 (7096): 979–983. Дои:10.1038 / природа04783. PMID 16791195.
- ^ Brown, C .; Li, P .; Boyd, J .; Delaney, K .; Мерфи, Т. (2007). «Обширный оборот дендритных шипов и ремоделирование сосудов в корковых тканях, восстанавливающихся после инсульта». Журнал неврологии. 27 (15): 4101–4109. Дои:10.1523 / JNEUROSCI.4295-06.2007. ЧВК 6672555. PMID 17428988.
- ^ Brown, C .; Мерфи, Т. Х. (2008). «Livin 'on the edge: визуализация дендритного оборота позвоночника в периинфарктной зоне во время ишемического инсульта и восстановления». Нейробиолог. 14 (2): 139–146. Дои:10.1177/1073858407309854. PMID 18039977.
- ^ а б Bhatt, D .; Zhang, S .; Ган, В. Б. (2009). «Динамика дендритного позвоночника». Ежегодный обзор физиологии. 71: 261–282. Дои:10.1146 / annurev.physiol.010908.163140. PMID 19575680.
- ^ фон Болен и Хальбах О., Захер С., Гасс П., Анзикер К. (2006). «Возрастные изменения в шипах гиппокампа и нарушения пространственной памяти у мышей». J Neurosci Res. 83 (4): 525–531. Дои:10.1002 / jnr.20759. PMID 16447268.
- ^ Harris, K .; Fiala, J .; Острофф, Л. (2003). «Структурные изменения в дендритных синапсах позвоночника во время долговременной потенциации». Философские труды: биологические науки. 358 (1432): 745–748. Дои:10.1098 / rstb.2002.1254. ЧВК 1693146. PMID 12740121.
- ^ Araya, R .; Николенко, В .; Eisenthal, K. B .; Юсте, Р. (2007). «Натриевые каналы усиливают потенциалы позвоночника». PNAS. 104 (30): 12347–12352. Дои:10.1073 / pnas.0705282104. ЧВК 1924793. PMID 17640908.
- ^ Ngo-Anh, T. J .; Bloodgood, B.L .; Lin, M .; Sabatini, B.L .; Maylie, J .; Адельман, Дж. П. (2005). «SK каналы и рецепторы NMDA образуют опосредованную Ca2 + петлю обратной связи в дендритных шипах». Природа Неврология. 8 (5): 642–649. Дои:10.1038 / nn1449. PMID 15852011.
- ^ Юсте, Р .; Денк, В. (1995). «Дендритные шипы как основные функциональные единицы нейрональной интеграции». Природа. 375 (6533): 682–684. Дои:10.1038 / 375682a0. PMID 7791901.
- ^ а б Bywalez, W. G .; Патирниче, Д .; Rupprecht, V .; Stemmler, M .; Herz, A. V .; Pálfi, D .; Balázs, R .; Эггер, В. (2015). «Локальная постсинаптическая активация потенциалзависимых натриевых каналов в дендритных шипах гранулярных клеток обонятельной луковицы». Нейрон. 85 (3): 590–601. Дои:10.1016 / j.neuron.2014.12.051. PMID 25619656.
- ^ Baer, S.M .; Ринзель, Дж. (1991). «Распространение дендритных шипов через возбудимые шипы: теория континуума». Журнал нейрофизиологии. 65 (4): 874–890. Дои:10.1152 / ян.1991.65.4.874. PMID 2051208.
- ^ Coombes, S .; Бресслофф, П. С. (2000). «Уединенные волны в модели дендритного кабеля с активными шипами». Журнал SIAM по прикладной математике. 61 (2): 432–453. CiteSeerX 10.1.1.104.1307. Дои:10.1137 / s0036139999356600. JSTOR 3061734.
- ^ Невиан, Т .; Сакманн, Б. (2006). «Передача сигналов Ca2 + в позвоночнике при пластичности, зависящей от времени спайка». Журнал неврологии. 26 (43): 11001–11013. Дои:10.1523 / JNEUROSCI.1749-06.2006. ЧВК 6674669. PMID 17065442.
- ^ Grunditz, A .; Holbro, N .; Tian, L .; Zuo, Y .; Эртнер, Т. Г. (2008). «Пластичность шейки позвоночника контролирует постсинаптические кальциевые сигналы посредством электрической компартментализации». Журнал неврологии. 28 (50): 13457–13466. Дои:10.1523 / JNEUROSCI.2702-08.2008. ЧВК 6671740. PMID 19074019.
- ^ а б Йошихара, Ю., Де Ру, М., Мюллер, Д. «Формирование и стабилизация дендритных шипов. Curr Opin Neurobiol (2009).
- ^ Саарикангас, Джуха и др. «MIM-индуцированное изгибание мембраны способствует инициации дендритного шипа». Клетка развития 33.6 (2015): 644-659.
- ^ Пелуччи, Сильвия; Стрингхи, Рамона; Марчелло, Елена (2020). «Дендритные шипы при болезни Альцгеймера: как актиновый цитоскелет способствует синаптической недостаточности». Международный журнал молекулярных наук. 21 (3): 908. Дои:10.3390 / ijms21030908. ISSN 1422-0067.
- ^ Penzes, P .; Cahill, M.E .; Джонс, К. А .; Vanleeuwen, J. E .; Вулфри, К. М. (2011). «Патология дендритного отдела позвоночника при нервно-психических расстройствах». Nat Neurosci. 14 (3): 285–293. Дои:10.1038 / № 2741. ЧВК 3530413. PMID 21346746.
- ^ Рамон-и-Кахал, S. Estructura de los centros nerviososde las aves. Rev. Trim. Histol. Норма. Пат. 1, 1-10 (1888).
- ^ Грей, Э. Г. (1959). «Электронная микроскопия синаптических контактов на дендритных шипах коры головного мозга». Природа. 183 (4675): 1592–1593. Дои:10.1038 / 1831592a0. PMID 13666826.
- ^ Dailey, M.E .; Смит, С. Дж. (1996). «Динамика дендритной структуры в развивающихся срезах гиппокампа». J Neurosci. 16 (9): 2983–2994. Дои:10.1523 / JNEUROSCI.16-09-02983.1996.
- ^ Bonhoeffer, T .; Юсте, Р. (2002). «Подвижность позвоночника. Феноменология, механизмы и функции». Нейрон. 35 (6): 1019–1027. Дои:10.1016 / s0896-6273 (02) 00906-6. PMID 12354393.
дальнейшее чтение
- Sudhof, T. C .; Стивенс, К. Ф .; Коуэн, В. М. (2001). Синапсы. Балтимор: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-6498-8.
- Левитан, И. Б .; Качмарек, Л. К. (2002). Нейрон: клеточная и молекулярная биология (Третье изд.). Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-514522-9.
- Нимчинский Э, Сабатини Б, Свобода К (2002). «Строение и функция дендритных шипов». Анну Рев физиол. 64: 313–53. Дои:10.1146 / annurev.physiol.64.081501.160008. PMID 11826272.
- Мацузаки М., Хонкура Н., Эллис-Дэвис Г., Касаи Н. (2004). «Структурные основы долговременной потенциации одиночных дендритных шипов». Природа. 429 (6993): 761–6. Дои:10.1038 / природа02617. ЧВК 4158816. PMID 15190253.
- Юсте Р., Маевская А., Холтхофф К. (2000). «От формы к функции: компартментализация кальция в дендритных шипах». Nat Neurosci. 3 (7): 653–9. Дои:10.1038/76609. PMID 10862697.
- Lieshoff C, Bischof H (2003). «Динамика изменения плотности позвоночника». Behav Brain Res. 140 (1–2): 87–95. Дои:10.1016 / S0166-4328 (02) 00271-1. PMID 12644282.
- Касаи Х., Мацузаки М, Ногучи Дж., Ясумацу Н. (2002). «Дендритные структуры и функции позвоночника». Нихон Синкей Сейшин Якуригаку Засши. 22 (5): 159–64. PMID 12451686.
- Линч Г., Рекс К.С., Галл С.М. (2007). «Консолидация LTP: основы, объяснительная сила и функциональное значение». Нейрофармакология. 52 (1): 12–23. Дои:10.1016 / j.neuropharm.2006.07.027. PMID 16949110.
внешняя ссылка
| Авторитетный контроль |
|---|