Незначительный антиген гистосовместимости - Minor histocompatibility antigen
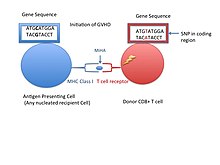
Незначительный антиген гистосовместимости (также известный как MiHA) являются рецепторами на клеточной поверхности донорских органов, которые, как известно, дают иммунологический ответ при трансплантации некоторых органов.[1] Они реже вызывают проблемы отторжения, чем проблемы главный комплекс гистосовместимости (MHC). Незначительные антигены гистосовместимости (MiHA) представляют собой разнообразные короткие сегменты белки и называются пептидами. Эти пептиды обычно имеют длину около 9-12 аминокислот и связаны как с основным комплексом гистосовместимости (MHC), так и с ним. учебный класс я и класс II белки.[2] Последовательности пептидов могут различаться у разных людей, и эти различия возникают из-за SNP в кодирующей области генов, делеции генов, мутации сдвига рамки считывания, или же вставки.[3] Около трети охарактеризованных MiHA происходят из Y-хромосомы.[4] Белки состоят из одного иммуногенного аллеля HLA.[2] Прежде чем стать короткой пептидной последовательностью, белки, экспрессируемые этими полиморфными или различными генами, должны быть переварены в протеасома на более короткие пептиды. Эти эндогенные или собственные пептиды затем транспортируются в эндоплазматический ретикулум с насосом-переносчиком пептидов, называемым КРАН где они встречаются и связываются с молекулой MHC класса I. Это контрастирует с Молекулы MHC класса II антигены, которые представляют собой пептиды, полученные в результате фагоцитоза / эндоцитоза и молекулярной деградации белков чужеродных сущностей, обычно антигенпрезентирующими клетками. Антигены MiHA либо повсеместно экспрессируются в большинстве тканей, таких как кожа и кишечник, либо ограниченно экспрессируются в иммунных клетках.[5]
Незначительные антигены гистосовместимости происходят из-за нормальных белков, которые сами по себе полиморфны в данной популяции. Даже если донор и реципиент трансплантата идентичны в отношении их главный комплекс гистосовместимости гены, аминокислотные различия в незначительный Белки могут вызывать медленное отторжение трансплантированной ткани. Некоторые из идентифицированных MiHA, кодируемых аутосомно и Y-хромосомой[4]
Известные минорные антигены гистосовместимости
В следующей таблице перечислены известные MiHA, варианты генов, кодирующие пептиды MiHA и их ограниченные аллели HLA.
| MiHA ID | Пептид MiHA | Ограниченный HLA | Хромосома | Координаты | ID SNP | Ген | Идентификатор гена Ensembl |
|---|---|---|---|---|---|---|---|
| HA-1 / A2 | VL [H / R] DDLLEA | А * 02: 01 | chr19 | 1068739 | rs1801284 | HMHA1 | ENSG00000180448 |
| HA-2 | YIGEVLVS [В / М] | А * 02: 01 | chr7 | 44977022 | rs61739531 | MYO1G | ENSG00000136286 |
| HA-8 | [R / P] TLDKVLEV | А * 02: 01 | chr9 | 2828765 | rs2173904 | KIAA0020 | ENSG00000080608 |
| HA-3 | V [T / M] EPGTAQY | А * 01: 01 | chr15 | 85579423 | rs2061821 | AKAP13 | ENSG00000170776 |
| C19ORF48 | CIPPD [S / T] LLFPA | А * 02: 01 | chr19 | 50798945 | rs3745526 | C19ORF48 | ENSG00000167747 |
| LB-ADIR-1F | СВАПАЛАЛ [Ф / С] ПА | А * 02: 01 | chr1 | 179082165 | rs2296377 | TOR3A | ENSG00000186283 |
| LB-HIVEP1-1S | СЛПХ [С / Н] ВТИ | А * 02: 01 | chr6 | 12123016 | rs2228220 | HIVEP1 | ENSG00000095951 |
| ЛБ-НИЩ-1А | ALAPAP [A / V] EV | А * 02: 01 | chr3 | 52489389 | rs887515 | НИШ | ENSG00000010322 |
| LB-SSR1-1S | [S / L] LAVAQDLT | А * 02: 01 | chr6 | 7310026 | rs10004 | SSR1 | ENSG00000124783 |
| LB-WNK1-1I | RTLSPE [I / M] ITV | А * 02: 01 | chr12 | 889199 | rs12828016 | WNK1 | ENSG00000060237 |
| T4A | GLYTYWSAG [A / E] | А * 02: 01 | chr3 | 140688418 | rs9876490 | TRIM42 | ENSG00000155890 |
| UTA2-1 | QL [L / P] NSVLTL | А * 02: 01 | chr12 | 31981704 | rs2166807 | KIAA1551 | ENSG00000174718 |
| PANE1 | RVWDLPGVLK | А * 03: 01 | chr22 | 41940168 | rs5758511 | CENPM | ENSG00000100162 |
| SP110 | SLP [R / G] ГЦТПК | А * 03: 01 | chr2 | 230207994 | rs1365776 | SP110 | ENSG00000135899 |
| ACC-1C | DYLQ [Y / C] VLQI | А * 24: 02 | chr15 | 79971064 | rs1138357 | BCL2A1 | ENSG00000140379 |
| ACC-1Y | DYLQ [Y / C] VLQI | А * 24: 02 | chr15 | 79971064 | rs1138357 | BCL2A1 | ENSG00000140379 |
| P2RX7 | WFHHC [H / R] PKY | А * 29: 02 | chr12 | 121167552 | rs7958311 | P2RX7 | ENSG00000089041 |
| АКК-4 | ATLPLLCA [R / G] | А * 31: 01 | chr15 | 78944951 | rs2289702 | CTSH | ENSG00000103811 |
| АКК-5 | WATLPLLCA [R / G] | А * 33: 03 | chr15 | 78944951 | rs2289702 | CTSH | ENSG00000103811 |
| LB-APOBEC3B-1K | [K / E] PQYHAEMCF | В * 07: 02 | chr22 | 38985821 | rs2076109 | APOBEC3B | ENSG00000179750 |
| LB-ARHGDIB-1R | LPRACW [R / P] EA | В * 07: 02 | chr12 | 14942624 | rs4703 | АРХГДИБ | ENSG00000111348 |
| LB-BCAT2-1R | QP [R / T] RALLFVIL | В * 07: 02 | chr19 | 48799813 | rs11548193 | BCAT2 | ENSG00000105552 |
| LB-EBI3-1I | RPRARYY [I / V] QV | В * 07: 02 | chr19 | 4236999 | rs4740 | EBI3 | ENSG00000105246 |
| LB-ECGF-1H | RP [H / R] AIRRPLAL | В * 07: 02 | chr22 | 50525826 | rs112723255 | TYMP | ENSG00000025708 |
| LB-ERAP1-1R | HPRQEQIALLA | В * 07: 02 | chr5 | 96803547 | rs26653 | ERAP1 | ENSG00000164307 |
| LB-FUCA2-1V | RLRQ [V / M] GSWL | В * 07: 02 | chr6 | 143502020 | rs3762002 | FUCA2 | ENSG00000001036 |
| LB-GEMIN4-1V | FPALRFVE [V / E] | В * 07: 02 | chr17 | 746265 | rs4968104 | БЛИЗНЕЦ4 | ENSG00000179409 |
| LB-PDCD11-1F | GPDSSKT [F / L] LCL | В * 07: 02 | chr10 | 103434329 | rs2986014 | PDCD11 | ENSG00000148843 |
| ЛБ-ТЕП1-1С | АПДГАКВА [S / P] L | В * 07: 02 | chr14 | 20383870 | rs1760904 | TEP1 | ENSG00000129566 |
| LRH-1 | TPNQRQNVC | В * 07: 02 | chr17 | 3690983 | rs3215407 | P2X5 | ENSG00000083454 |
| ЗАФИР | IPRDSWWVEL | В * 07: 02 | chr19 | 57492212 | rs2074071 | ZNF419 | ENSG00000105136 |
| HEATR1 | ИСКЕРА [E / G] AL | В * 08: 01 | chr1 | 236554626 | rs2275687 | HEATR1 | ENSG00000119285 |
| HA-1 / B60 | KECVL [H / R] DDL | В * 40: 01 | chr19 | 1068739 | rs1801284 | HMHA1 | ENSG00000180448 |
| LB-SON-1R | SETKQ [R / C] TVL | В * 40: 01 | chr21 | 33553954 | rs13047599 | СЫН | ENSG00000159140 |
| LB-SWAP70-1Q | MEQLE [Q / E] LEL | В * 40: 01 | chr11 | 9748015 | rs415895 | SWAP70 | ENSG00000133789 |
| LB-TRIP10-1EPC | G [E / G] [P / S] QDL [C / G] TL | В * 40: 01 | chr19 | 6751268 | rs1049229 | TRIP10 | ENSG00000125733 |
| SLC1A5 | AE [A / P] TANGGLAL | В * 40: 02 | chr19 | 46787917 | rs3027956 | SLC1A5 | ENSG00000105281 |
| АКК-2 | KEFED [D / G] IINW | В * 44: 03 | chr15 | 79970875 | rs3826007 | BCL2A1 | ENSG00000140379 |
| АКК-6 | МЕЙФИЕВФШФ | В * 44: 03 | chr18 | 63953532 | rs9945924 | HMSD | ENSG00000221887 |
| HB-1H | EEKRGSL [H / Y] VW | В * 44: 03 | chr5 | 143820488 | rs161557 | HMHB1 | ENSG00000158497 |
| HB-1Y | EEKRGSL [H / Y] VW | В * 44: 03 | chr5 | 143820488 | rs161557 | HMHB1 | ENSG00000158497 |
| DPH1 | S [V / L] LPEVDVW | В * 57: 01 | chr17 | 2040586 | rs35394823 | DPH1 | ENSG00000108963 |
| UTDP4-1 | R [I / N] LAHFFCGW | DPB1 * 04 | chr9 | 128721272 | rs11539209 | ZDHHC12 | ENSG00000160446 |
| CD19 | WEGEPPC [L / V] P | DQB1 * 02: 01 | chr16 | 28933075 | rs2904880 | CD19 | ENSG00000177455 |
| LB-PI4K2B-1S | SRSS [S / P] AELDRSR | DQB1 * 06: 03 | chr4 | 25234395 | rs313549 | PI4K2B | ENSG00000038210 |
| LB-MTHFD1-1Q | SSIIAD [Q / R] IALKL | DRB1 * 03: 01 | chr14 | 64442127 | rs2236225 | MTHFD1 | ENSG00000100714 |
| LB-LY75-1K | LGITYR [N / K] KSLMWF | DRB1 * 13: 01 | chr2 | 159819916 | rs12692566 | LY75 | ENSG00000054219 |
| SLC19A1 | [R / H] LVCYLCFY | DRB1 * 15: 01 | chr21 | 45537880 | rs1051266 | SLC19A1 | ENSG00000173638 |
| LB-PTK2B-1T | ВЫМНД [Т / К] СПЛТПЭК | DRB3 * 01: 01 | chr8 | 27451068 | rs751019 | PTK2B | ENSG00000120899 |
| LB-MR1-1R | YFRLGVSDPI [R / H] G | DRB3 * 02: 02 | chr1 | 181049100 | rs2236410 | MR1 | ENSG00000153029 |
Ответ Т-клеток на MiHA
MiHA, связанные с MHC, представленным на поверхности клетки, могут распознаваться как собственный пептид или не распознаваться ни одним из них. CD8 + или CD4 + Т-клетки. Отсутствие распознавания Т-клеткой этого аутоантигена является причиной того, что аллогенные стволовые клетки трансплантация для HLA-согласованного гена или MiHA развивающегося плода во время беременности могут не распознаваться Т-клетками и помечаться как чужеродные, что приводит к иммунному ответу. Хотя рецепторы B-клеток также могут распознавать MHC, иммунные ответы, по-видимому, вызываются только T-клетками.[6] Последствия иммунного ответа видны при трансплантации аллогенных гемопоэтических стволовых клеток (HCT), когда пептиды, кодируемые полиморфными генами, различаются между реципиентом и донорскими Т-клетками. В результате донорские Т-клетки могут нацеливаться на клетки-реципиенты, что называется болезнью трансплантат против хозяина (РТПХ ).[5] Хотя отторжение трансплантата или костного мозга может иметь пагубные последствия, существуют преимущества иммунотерапии, когда цитотоксические Т-лимфоциты специфичны для аутоантигена и могут нацеливаться на антигены, избирательно экспрессируемые на лейкозных клетках, чтобы разрушить эти опухолевые клетки, называемые эффект трансплантата против лейкемии (ГВЛ).[3]
Узнавание зрелой Т-клетки этого аутоантигена не должно вызывать иммунного ответа. Во время селекции тимуса в вилочковой железе только тимоцит TCR, который распознает молекулу MHC класса I или класса II плюс пептид, должен выжить положительный отбор. Однако есть смерть от апоптоз тимоцитов, которые не взаимодействуют с молекулами MHC или имеют высокоаффинные рецепторы для собственного MHC плюс аутоантиген, процесс, называемый отрицательный выбор. Следовательно, процесс положительного и отрицательного отбора означает, что меньшее количество самореактивных зрелых Т-клеток покидает тимус и приводит к аутоиммунным проблемам.
Открытие МИХА
Значение MiHA в иммунном ответе было признано после трансплантации. У реципиента развилась РТПХ, несмотря на наличие HLA-совпадающих генов в главном локусе гистосовместимости. Эксперимент поднял вопросы о возможности существования MiHA. В частности, первая MiHA была обнаружена, когда трансплантация костного мозга произошла между представителями противоположного пола. Женщина-реципиент получила MHC-подходящие клетки костного мозга, но все еще имела активные цитотоксические Т-клетки (CD8 +).[3] CD8 + Т-клетки были активными и нацелены на клетки мужского костного мозга. Было обнаружено, что мужские клетки костного мозга представляют пептид в бороздке MHC, кодируемый геном на Y-хромосоме. Пептид был чужеродным для женских Т-клеток, а самки лишены Y-хромосомы и, следовательно, этого MiHA. MiHA, кодируемые Y-хромосомой, известны как антигены HY.[3]
H-Y антиген
Антигены H-Y кодируются генами на Y-хромосоме. Было обнаружено, что оба аллеля HLA класса I и II представляют эти антигены. Некоторые из этих антигенов повсеместно экспрессируются в ядросодержащих мужских клетках, и присутствие этих антигенов было связано с повышенным риском развития трансплантации аллогенных стволовых клеток GVHD для гена, согласованного с HLA, когда есть реципиент-мужчина и донор-женщина.[7] H-Y MiHA играют роль при беременности плодом мужского пола, потому что фетальные клетки могут переходить из плаценты в кровоток матери, где материнские Т-клетки отвечают на чужеродный антиген, представленный как на MHC класса I, так и на II. Следовательно, H-Y-специфические CD8 + Т-клетки развиваются в материнской крови и могут нацеливаться на клетки плода с ядром, экспрессирующим антиген на молекуле MHC класса I. Ответ на эти фетальные антигены H-Y проявляется у женщин, перенесших вторичный повторный выкидыш, которые ранее были беременны плодом мужского пола.[3] Женщины с более ранней мужской беременностью имеют Т-клетки, которые ранее подвергались воздействию этих антигенов H-Y, и, следовательно, быстрее их распознают. Было обнаружено, что женщины с рецидивирующим невынашиванием также содержат MHC II со способностью представлять эти антигены Т-хелперам (CD4 +), что важно для активации CD8 +.[8]
Антиген гистосовместимости 1 (HA1)
HA1 - результат SNP преобразование неиммуногенного аллеля (KECVLрDDLLEA) к иммуногенному аллелю (KECVLЧАСDDLLEA). Этот SNP приводит к лучшей способности связывания пептида с бороздкой определенных молекул MHC класса I, обнаруженных на антигенпрезентирующих клетках.[5] Значение превращения пептида в иммуногенную форму заключается в том, что теперь специфические Т-клетки, ограниченные HLA-A 0201, могут распознавать пептид, представленный молекулами HLA-A0201 класса I MHC. Это распознавание приводит к иммунному ответу, если Т-клетки распознают пептид как чужеродный. Это распознавание происходит, когда у человека отсутствует иммуногенная версия пептида, но он подвергается воздействию пептида HA-1 во время беременности или трансплантации аллогенных стволовых клеток. Было обнаружено, что во время беременности фетальный HA-1 происходит из плаценты, и были идентифицированы специфические материнские CD8 + Т-клетки, распознающие этот MiHA.[5]
Иммунотерапия Эффект трансплантата против лейкемии
CD8 + Т-клетки, специфичные для MiHA, могут нацеливаться на эти антигены, когда они специфически экспрессируются на опухолевых клетках, что позволяет уничтожать вредоносные опухолевые клетки. Было показано, что у мышей CD8 + Т-клетки донора аллогенной трансплантации стволовых клеток, специфичные для MiHA, обнаруженные у реципиента, ингибируют деление лейкозный клетки. Однако существует риск развития GVHD, если Т-клетки специфичны для MiHA, повсеместно экспрессируемых на эпителиальный клетки. Более конкретно, HA-8, UGT2B17 и SMCY MiHA, которые экспрессируются повсеместно, представляют более высокий риск развития GVHD. Следовательно, для предотвращения неблагоприятных эффектов GVHD ограниченные иммунными клетками MiHA являются идеальными мишенями для трансплантат-против-лейкемии (GVL), поскольку не все ядросодержащие клетки нацелены на реагирующие Т-клетки. Примером идеальной мишени является MiHA HB-1, который высоко экспрессируется во вредных В-клетках, но имеет низкую экспрессию в клетках других тканей.[9]
Клинические последствия
Иммунизация матерей против специфичных для самцов минорных антигенов гистосовместимости (H-Y) играет патогенную роль во многих случаях вторичный повторный выкидыш, то есть повторный выкидыш при беременности, наступившей после предыдущего живорождения. Примером этого эффекта является то, что соотношение детей, рожденных до и после вторичного повторного выкидыша, составляет 1,49 и 0,76 соответственно.[10]
Смотрите также
Рекомендации
- ^ Робертсон Н.Дж., Чай Дж. Г., Миллрейн М., Скотт Д., Хашим Ф., Манктелоу Э., Лемонье Ф., Симпсон Э., Дайсон Дж. (Март 2007 г.). «Естественная регуляция иммунитета к минорным антигенам гистосовместимости». Журнал иммунологии. 178 (6): 3558–65. Дои:10.4049 / jimmunol.178.6.3558. PMID 17339452.
- ^ а б Дзежак-Митла М., Маркевич М., Сиекьера Ю., Мизия С., Кочлега А., Зелинска П., Собчик-Крушельницка М., Кирч-Кшемиен С. (2012). «Возникновение и влияние несоответствий незначительных антигенов гистосовместимости на результаты трансплантации гемопоэтических стволовых клеток от HLA-подходящих родственных доноров». Исследование костного мозга. 2012: 257086. Дои:10.1155/2012/257086. ЧВК 3502767. PMID 23193478.
- ^ а б c d е Линшайд С, Петров М.Г. (апрель 2013 г.). «Незначительные антигены гистосовместимости и иммунный ответ матери на плод во время беременности». Американский журнал репродуктивной иммунологии. 69 (4): 304–14. Дои:10.1111 / aji.12075. ЧВК 4048750. PMID 23398025.
- ^ а б Хираяма М, Адзума Э, Комада Y (2012). Основные и второстепенные антигены гистосовместимости с ненаследственными материнскими антигенами (NIMA), гистосовместимость. ИНТЕК. п. 146. ISBN 978-953- 51-0589-3.
- ^ а б c d Бликли М., Ридделл С.Р. (март 2011 г.). «Использование Т-клеток, специфичных для минорных антигенов гистосовместимости человека, для терапии лейкемии». Иммунология и клеточная биология. 89 (3): 396–407. Дои:10.1038 / icb.2010.124. ЧВК 3061548. PMID 21301477.
- ^ Perreault C, Décary F, Brochu S, Gyger M, Bélanger R, Roy D (1990). «Незначительные антигены гистосовместимости» (PDF). Кровь. 76 (7): 1269–80. PMID 2207305.
- ^ Nielsen HS (01.07.2011). «Вторичный невынашивание беременности и иммунитет к HY». Обновление репродукции человека. 17 (4): 558–74. Дои:10.1093 / humupd / dmr005. PMID 21482560.
- ^ Лиссауэр Д., Пайпер К., Гудиер О, Килби, доктор медицины, Мосс, Пенсильвания (июль 2012 г.). «Плодоспецифические CD8 + цитотоксические Т-клеточные ответы развиваются во время нормальной беременности человека и демонстрируют широкие функциональные возможности». Журнал иммунологии. 189 (2): 1072–80. Дои:10.4049 / jimmunol.1200544. PMID 22685312.
- ^ Бликли М, Ридделл С.Р. (2004). «Молекулы и механизмы эффекта трансплантат против лейкемии». Обзоры природы. Рак. 4 (5): 371–80. Дои:10.1038 / nrc1365. PMID 15122208.
- ^ Нильсен HS (2011). «Вторичный невынашивание беременности и иммунитет к HY». Обновление репродукции человека. 17 (4): 558–74. Дои:10.1093 / humupd / dmr005. PMID 21482560.
внешняя ссылка
- Незначительный + гистосовместимость + антигены в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)