Евчамберсия - Euchambersia
| Евчамберсия | |
|---|---|
 | |
| КТ черепов типового (справа) и второго (слева) образцов | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Терапсида |
| Подотряд: | †Тероцефалия |
| Семья: | †Akidnognathidae |
| Род: | †Евчамберсия Веник, 1931 г. |
| Типовой вид | |
| †Euchambersia mirabilis Веник, 1931 г. | |
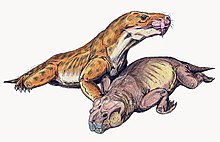
Евчамберсия это род тероцефал терапсид которые жили во время Поздняя пермь, примерно 255 миллионов лет назад, в том, что сейчас Южная Африка. Род состоит из одного вида, Euchambersia mirabilis, названный палеонтологом Роберт Брум в 1931 г. из черепа отсутствует нижняя челюсть; второй череп, принадлежавший незрелой особи, был описан позже. Это член семьи Akidnognathidae, который исторически также упоминается как синоним Euchambersiidae (названный в честь Евчамберсия).
Евчамберсия был маленьким тероцефалом с короткой мордой и большим клыки что типично для группы. Однако среди тероцефалов он примечателен наличием гребней на клыках и большой выемкой на боковой стороне черепа. Исходя из ошибочного предположения, что клыки имеют бороздки, а не гребни, было высказано предположение, что эти структуры поддерживают механизм доставки яда. Совсем недавно внутреннее строение черепа Евчамберсия был использован как более сильное доказательство в пользу гипотезы о ядовитости; другие возможности, такие как углубление, поддерживающее какой-то орган чувств, все еще остаются правдоподобными.
Открытие и наименование

Типовой образец Евчамберсия был найден Роберт Брум на Южноафриканский ферма Ванвиксфонтейн, принадлежащая мистеру Грейтхеду, недалеко от города Норвалспонт.[1] Он состоит из единственного искаженного черепа, занесенного в каталог как NHMUK R5696, который был описан Брумом в 1931 году. Второй череп меньшего размера с номером экземпляра BP / 1/4009 был найден в 1966 году.[2] и описан Джеймсом Китчингом в 1977 году.[3] У обоих экземпляров отсутствует нижняя челюсть. Они произошли из того же самого пласта породы, в верхнем Цистецефал Зона сборки из Бофорт Групп в пределах Кару Супергруппа.[3] В Цистецефал AZ был приурочен к Wuchiapingian этап Поздняя пермь,[4] от 256,2 до 255,2 млн лет назад.[5]
Веник назвал род Евчамберсия, которую он считал "самым выдающимся тероцефалом, когда-либо обнаруженным" после выдающегося Шотландский издатель и эволюционный мыслитель Роберт Чемберс, чей Остатки естественной истории творения Брум считал «очень замечательным произведением», хотя «многие насмехались над ним».[1]
Описание
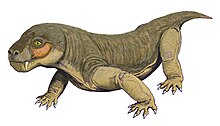
Евчамберсия был маленьким и короткомордым для тероцефал реконструированная длина черепа составляет приблизительно 116 миллиметров (4,6 дюйма), что объясняет раздавливание и деформацию окаменелости. Второй известный череп принадлежал более мелкому человеку, его длина составляла 80 миллиметров (3,1 дюйма); Судя по отсутствию сращения черепа, вероятно, он был незрелым.[2]
Согласно первоначальному описанию, глазница Евчамберсия был довольно маленьким. Филиалы заглазничный и скуловой которые обычно окружают заднюю и нижнюю часть глазницы у тероцефалов, оказываются либо очень уменьшенными, либо полностью отсутствуют. Между тем, верхняя часть глазницы образована префронтальный, а лобной тоже маленький. Череп не несет шишковидное отверстие. Нравиться Whaitsia, то крыловидный и небный из нёбо не отделены от транспалатина дальше сбоку челюсти никаким отверстием.[1]
Зубы
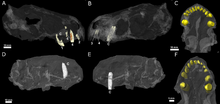
Хотя черепа сохранились не полностью, КТ сканирование предполагает, что каждый предчелюстная кость провел пять резцы, с Розетки становится все больше от первого до пятого резца. Как и другие териодонты коронки резцов конические; у них также нет зубцов, в отличие от горгонопсийцы и сцилакозавр тероцефалы. Внутренний край резцов кажется слегка вогнутым, а задний край имеет выступ. У меньшего экземпляра смещенный резец сохранился в носовой полости; он более сильно изогнут и имеет следы износа на его верхнем крае, что позволяет предположить, что это, вероятно, нижний резец. За его четвертым резцом также растет замещающий зуб, сопровождаемый рассасывание корня.[2]
Типовой образец сохраняет право собачий.[2] Как и другие тероцефалы, собаки Евчамберсия был очень большим, что привело к особому образу жизни хищников, который включает укус саблезуба при убийстве добычи.[6] Он круглый в поперечном сечении,[3] и имеет выступающий гребень сбоку на передней поверхности. Непосредственно рядом с этим гребнем находится неглубокое углубление, которое становится шире у вершины зуба, вероятно, имеющее ту же структуру, что и канавка, интерпретируемая некоторыми авторами.[2][7] Териодонты обычно заменяют зубы чередующимися зубами.[8] (или дистихиальный) узор,[9][10] такой, что клык всегда в рабочем состоянии; оба черепа Евчамберсия не обнаруживают никаких признаков развития замещающих зубов, что позволяет предположить, что Евчамберсия полагался на наличие и функциональности обоих клыков одновременно.[2]
Верхнечелюстная ямка и связанные с ней каналы

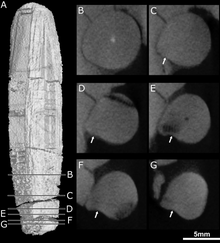
За резцами и клыками на челюсти дополнительных зубов не было. Там, где зубы располагаются у тероцефалов, у которых есть зубы позади клыков, вместо этого есть большое углубление или ямка на стороне верхняя челюсть, который также ограничен снизу частью слезный и, возможно, часть скуловой кости.[1] Эта ямка составляет 48% длины челюсти у типового образца и 38% во втором черепе. В обоих черепах эта ямка разделена на две части: более мелкий гребень наверху и более крупное и глубокое углубление внизу. Широкая борозда, начинающаяся позади клыка, контактирует со дном ямки, а затем переходит во внутреннюю часть рта. Нижняя часть ямки сильно изъедена и имеет небольшое отверстие, или отверстие, как на передней, так и на задней поверхности.[2]
КТ показывает, что эти отверстия ведут к каналам, которые соединяются с тройничный нерв, который контролирует чувствительность лица. Направленный вперед канал также разделяется на три основных ветви подглазничный нерв,[11] все они подключаются к розетке клыка; стык происходит примерно на 3–6 миллиметрах (0,12–0,24 дюйма) вдоль канала, это еще одна точка различия между двумя черепами. Верхняя ветвь, наружная ветвь носа, разделяется на четыре ветви в типичном черепе, но не разделяется во втором черепе. В другом терапсиды подобно Тринаксодон, Баурия, и Olivierosuchus, внешняя ветвь носа обычно разделяется на три или более ответвлений. Все эти каналы подводили нервы и богатые питательными веществами ткани к корню клыков и остальной части верхней челюсти.[2][11]
Классификация
В 1934 г. Евчамберсия был отнесен к новому семейству Euchambersiidae Льюве Дирк Бунстра.[12][13] Изначально Бунстра неправильно написал имя как Euchambersidae (что является неправильным латинским), и впоследствии было исправлено Фридрих фон Хюене в 1940 г. Euchambersiidae первоначально считалась отдельным от семейств Moschorhinidae и Annatherapsididae; в 1974 году Кристиана Мендес определила эти группы как тесно связанные подсемейства (переименованные в Annatherapsidinae, Moschorhininae и Euchambersiinae) в более широкой группе переопределенных ею Moschorhinidae (хотя она также назвала их Annatherapsididae).[14]
Филогенетический анализ 1986 г. Джеймс Хопсон и Херб Баргусен поддержали гипотезу Мендеса о трех подсемействах в пределах Moschorhinidae, но они решили использовать название Euchambersiidae. В 2009 году Адам Хаттенлокер и его коллеги утверждали, что названия Annatherapsididae, Moschorhinidae и Euchambersiidae являются младшими синонимами Akidnognathidae, поскольку Akidnognathus (который также принадлежит к той же семье) был назван первым перед любым другим членом семьи.[14] Это имя получило более широкое признание среди исследователей.[14][15][16] Huttenlocker и другие. также позже переопределил Moschorhininae как всех Akidnognathidae, за исключением Аннатерапсидус и Akidnognathus.[17]

В 2008 году Ивахненко включил Akidnognathidae (как Euchambersiidae) как сестринскую группу семейства. Whaitsiidae в надсемействе Whaitsioidea.[13] Однако другие исследователи не включают Akidnognathidae в Whaitsioidea. Филогении Хаттенлокера и другие. обнаружил, что Akidnognathidae были ближе всего к Chthonosauridae, причем два образуют сестринская группа в группу, содержащую Whaitsioidea и Baurioidea. Топология, восстановленная анализом Huttenlocker в 2016 г. и другие. показано ниже.[17]
| Тероцефалия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология

Яд
Большая челюстная ямка Евчамберсия были постоянными предметами споров относительно их функции. Однако большинство исследователей сходятся во мнении, что в них была какая-то секреторная железа. Хотя Брум первоначально утверждал, что ямки могли содержать околоушный слюнные железы,[1] это предложение было отвергнуто Boonstra и J.P. Lehman, которые отметили, что околоушные железы обычно располагаются за глазом; они соответственно предположили, что ямки были изменены слезные железы и Железы Хардера.[2] Однако последнее также маловероятно, потому что железы Хардера обычно располагаются внутри глазницы. Франц Нопца предположили, что в верхнечелюстных ямках располагались ядовитые железы (которые, возможно, произошли от слезных желез), с ребристыми клыками и выемками за клыками, позволяющими яду пассивно течь в кровоток жертвы.[18] Эта гипотеза была широко принята на протяжении ХХ века.[15][19][20][21] и характерная морфология Евчамберсия использовался для поддержки возможной адаптации к ядовитости среди различных других доисторических животных,[7][13][22][23] включая тероцефалов Ичибенгопс.[24]

Во многом это признание было основано на ошибочном предположении, что клыки имеют бороздки, а не гребни;[3] рифленые клыки в Евчамберсия будут параллельны клыкам различных ядовитых змей, а также доставляющим яд резцам живых соленодоны.[21] Эта интерпретация, которая постоянно появлялась в литературе, опубликованной после 1986 года, была определена Жюльеном Бенуа как результат распространения чрезмерно реконструированной схемы черепа Брума без контекста реальных образцов. Таким образом, он считал необходимым пересмотреть гипотезу о ядовитом укусе в Евчамберсия.[3] Кроме того, Бенуа утверждал, что клыки с бороздками и гребнями не обязательно связаны с ядовитыми животными, о чем свидетельствует их присутствие в гиппопотамы, мунтжаки, и бабуины, в котором они играют роль в уходе за зубами или их заточке;[3][21][25] в последних двух ребристые клыки также сопровождаются отчетливой ямкой перед глазом, которая совершенно не связана с ядом.[21][26] Кроме того, рифленые и гребенчатые зубы у неядовитых змей используются для уменьшения сопротивления всасыванию при поимке скользкой добычи, такой как рыба или беспозвоночные.[27]
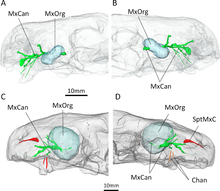
КТ-сканирование известных образцов Евчамберсия Бенуа и его коллеги впоследствии использовали его для более конкретной поддержки гипотезы о яде. Каналы, ведущие в верхнечелюстные ямки и из них, как показали сканирование, в первую очередь поддерживали тройничный нерв, а также кровеносные сосуды.[28][29][30] Однако тот факт, что каналы также напрямую ведут к корню клыков, предполагает, что они играли второстепенную роль в доставке яда. В целом, Евчамберсия по-видимому, имел ядовитую железу (расположенную в верхнечелюстных ямках), механизм доставки яда (верхнечелюстные каналы) и инструмент, с помощью которого можно было нанести рану для доставки яда (ребристые клыки), которые удовлетворяли критериям ядовитого животного по определению Вольфганга Бюхерля.[31] Бенуа и другие. отметил, что это не окончательно демонстрирует, что Евчамберсия был действительно ядовитым, особенно с учетом ранее заявленных возражений. Кроме того, нет живых животных с системой доставки, аналогичной предложенной системе для Евчамберсия (большинство доставляют яд через нижнюю челюсть,[32][33] в то время как у змей есть специализированные протоки.[2][34]
Альтернативная гипотеза, предложенная Бенуа и другие. включает в себя какой-то сенсорный орган, занимающий верхнечелюстную ямку. Уникально среди терапсидов,[11] канал в верхней челюсти обнажен на тыльной стороне верхнечелюстной ямки, что означает, что канал, по которому проходит тройничный нерв, вероятно, простирался бы через ямку за пределы контура черепа. Бенуа и другие. предположили, что ямка могла поддерживать специализированный орган чувств, аналогичный ямочному органу яма гадюк и некоторые другие змеи,[35] или альтернативно ганглий нервных клеток. Также возможно, что этот орган функционировал как замена теменный глаз в Евчамберсия, как ямочный орган у ямочной гадюки.[36] Однако такой расширенный орган чувств был бы беспрецедентным среди четвероногие, и несколько других тероцефалов, у которых также отсутствует теменный глаз, также не имеют челюстной ямки.[37] Таким образом, Бенуа и другие. считал гипотезу о яде более правдоподобной.[2]
Палеоэкология
В Цистецефал Зона сборки, откуда Евчамберсия известно, представляет собой пойма это было покрыто множеством небольших, относительно прямых ручьев. Уровень воды в этих ручьях, вероятно, зависел от сезона.[4] Судя по пыльце, сохранившейся в Цистецефал AZ, таксон пыльцы Питиоспориты (который, вероятно, произошел от растения, похожего на Глоссоптерис ) было очень распространено, составляя от 80% до 90% обнаруженной пыльцы (хотя преобладающие отложения не были идеальными для сохранения пыльцы).[38]
в Цистецефал AZ, другие сопутствующие тероцефалы включены Хофмейрия, Гомодонтозавр, Иктидостома, Ictidosuchoides, Иктидозухопс, Macroscelesaurus, Полицинодон, и Проалопекопсис. Однако более многочисленными были горгонопсии, в том числе Аэлурогнатус, Аэлурозавр, Алопозавр, Арктогнат, Арктопс, Cerdorhinus, Clelandina, Ционозавр, Диногоргон, Горгонопс, Lycaenops, Леонтоцефал, Пардоцефал, Прорубиджея, Rubidgea, Сцилакопы, Скимногнат, и Сикозавр.[4]
Безусловно, самым многочисленным травоядным животным был дицинодонт Дииктодон, с более чем 1900 известными экземплярами из Цистецефал AZ. Включены другие дицинодонты Аулацефалодон, Цистецефал, Дицинодон, Dicynodontoides, Дигалодон, Dinanomodon, Emydops, Эндотиодон, Kingoria, Китчинганомодон, Oudenodon, Palemydops, Пеланомодон, Пристеродон, и Рахиоцефал. В биармозухи Лемурозавр, Lycaenodon, Парабурнетия, и Рубидгина также присутствовали вместе с цинодонты Цинозавр и Procynosuchus. Несинапсиды включают архозавроморф Янгиния; то парарептилии Anthodon, Milleretta, Нанопария, Оуэнетта, и Парейазавр; и темноспондил Rhinesuchus.[4]
Смотрите также
Рекомендации
- ^ а б c d е Веник, Р. (1931). «Уведомления о некоторых новых родах и видах ископаемых рептилий Карру». Записи музея Олбани. 4 (1): 161–166.
- ^ а б c d е ж грамм час я j k Benoit, J .; Norton, L.A .; Manger, P.R .; Рубидж, Б.С. (2017). "Переоценка отравляющей способности Euchambersia mirabilis (Therapsida, Therocephalia) с использованием методов μCT-сканирования ". PLoS ONE. 12 (2): e0172047. Bibcode:2017PLoSO..1272047B. Дои:10.1371 / journal.pone.0172047. ЧВК 5302418. PMID 28187210.
- ^ а б c d е ж Бенуа, Дж. (2016). «Обзор гипотезы« ядовитого тероцефала »и то, как многочисленные репрезентации Евчамберсия повлияли на его успех и наоборот ". Bulletin de la Société Géologique de France. 187 (4): 217–224. Дои:10.2113 / gssgfbull.187.4-5.217.
- ^ а б c d Smith, R .; Rubidge, B .; ван дер Вальт, Меррилл (2012). «Образцы биоразнообразия терапсид и палеоокружение в бассейне Кару, Южная Африка». В Чинсамы-Туран, А. (ред.). Предшественники млекопитающих: радиация, гистология, биология. Блумингтон: Издательство Индианского университета. С. 31–64. ISBN 978-0-253-00533-5.
- ^ Rubidge, B.S .; Erwin, D.H .; Ramezani, J .; Bowring, S.A .; де Клерк, W.J. (2013). «Высокоточная временная калибровка биостратиграфии поздних пермских позвоночных: ограничения U-Pb циркона из супергруппы Karoo, Южная Африка». Геология. 41 (3): 363–366. Bibcode:2013Гео .... 41..363р. Дои:10.1130 / G33622.1.
- ^ Андерссон, К .; Norman, D .; Верделин, Л. (2011). "Саблезубые хищники и убийство крупной добычи". PLoS ONE. 6 (10): e24971. Bibcode:2011PLoSO ... 624971A. Дои:10.1371 / journal.pone.0024971. ЧВК 3198467. PMID 22039403.
- ^ а б Сьюз, Х.-Д. (1991). «Зубы, проводящие яд у рептилий триасового периода». Природа. 351 (6322): 141–143. Bibcode:1991Натура.351..141С. Дои:10.1038 / 351141a0.
- ^ Kermack, D.W .; Кермак, К. (1984). «Зубные протезы, замена зубов и сустав челюсти». Эволюция характеров млекопитающих. Springer США. С. 66–68. Дои:10.1007/978-1-4684-7817-4. ISBN 978-1-4684-7819-8.
- ^ Кермак, К. (1956). «Замена зубов у млекопитающих рептилий подотряда Gorgonopsia и Therocephalia». Философские труды Королевского общества B. 240 (670): 95–133. Bibcode:1956РСПТБ.240 ... 95К. Дои:10.1098 / рстб.1956.0013.
- ^ Хопсон, Дж. (1964). «Замена зубов цинодонтов, дицинодонтов и тероцефальных рептилий». Журнал зоологии. 142 (4): 625–654. Дои:10.1111 / j.1469-7998.1964.tb04632.x.
- ^ а б c Benoit, J .; Manger, P.R .; Рубидж, Б. (2016). «Палеоневрологические ключи к эволюции определения характеристик мягких тканей млекопитающих». Научные отчеты. 6: 25604. Bibcode:2016НатСР ... 625604Б. Дои:10.1038 / srep25604. ЧВК 4860582. PMID 27157809.
- ^ Бунстра Л.Д. 1934. "Вклад в морфологию млекопитающих рептилий подотряда Therocephalia". Летопись Южноафриканского музея, 31: 215–267
- ^ а б c Ивахненко, М.Ф. (2008). «Первый вайциид (Therocephalia, Theromorpha)». Палеонтологический журнал. 42 (4): 409–413. Дои:10.1134 / S0031030108040102.
- ^ а б c Хаттенлокер, А. (2009). «Исследование кладистических взаимоотношений и монофилии тероцефальных терапсид (Amniota: Synapsida)». Зоологический журнал Линнеевского общества. 157 (4): 865–891. Дои:10.1111 / j.1096-3642.2009.00538.x.
- ^ а б Rubidge, B.S .; Сидор, К.А. (2001). «Эволюционные закономерности среди пермо-триасовых терапсид». Ежегодный обзор экологии и систематики. 32: 449–480. Дои:10.1146 / annurev.ecolsys.32.081501.114113.
- ^ Сигурдсен, Т. (2006). «Новые особенности морды и орбиты тероцефального терапсида из Южной Африки». Acta Palaeontologica Polonica. 51 (1): 63–75.
- ^ а б Huttenlocker, A.K .; Сидор, К.А. (2016). «Первые каренитиды (Therapsida, Therocephalia) из верхней перми Гондваны и биогеография пермо-триасовых тероцефалов». Журнал палеонтологии позвоночных. 36 (4): e1111897. Дои:10.1080/02724634.2016.1111897.
- ^ Нопса, Ф. (1933). «О биологии тероморфных рептилий. Евчамберсия". Летопись и журнал естественной истории. 10. 12 (67): 125–126. Дои:10.1080/00222933308673757.
- ^ Watson, D.M .; Ромер, А. (1956). «Классификация терапевсидных рептилий». Вестник Музея сравнительной зоологии. 114: 35–89.
- ^ Ван Вален, Л. (1960). «Терапсиды как млекопитающие». Эволюция. 14 (3): 304–313. Дои:10.2307/2405973. JSTOR 2405973.
- ^ а б c d Folinsbee, K.E .; Muller, J .; Reisz, R.R. (2007). «Собачьи борозды: морфология, функция и отношение к яду». Журнал палеонтологии позвоночных. 27 (2): 547–551. Дои:10.1671 / 0272-4634 (2007) 27 [547: cgmfar] 2.0.co; 2. JSTOR 30126324.
- ^ Сьюз, Х.-Д. (1996). «Зуб рептилии с очевидными ядовитыми каналами из группы Чинле (верхний триас) в Аризоне». Журнал палеонтологии позвоночных. 16 (3): 571–572. Дои:10.1080/02724634.1996.10011340.
- ^ Gong, E .; Martin, L.D .; Burnham, D.A .; Фальк, А. (2009). "Птичий хищник Синорнитозавр был ядовитым ". Труды Национальной академии наук Соединенных Штатов Америки. 107 (2): 766–768. Bibcode:2010ПНАС..107..766Г. Дои:10.1073 / pnas.0912360107. ЧВК 2818910. PMID 20080749.
- ^ Huttenlocker, A.K .; Sidor, CA; Ангельчик, К. (2015). «Новый эвтероцефал (Therapsida, Therocephalia) из верхней пермской формации аргиллита Мадумабиса (бассейн Луангвы) в Замбии». Журнал палеонтологии позвоночных. 35 (5): e969400. Дои:10.1080/02724634.2015.969400.
- ^ Mitchell, J.S .; Heckert, A.B .; Сьюз, Х.-Д. (2010). «Канавки для трубок: эволюция системы доставки яда у рептилий позднего триаса»"". Naturwissenschaften. 97 (12): 1117–1121. Bibcode:2010NW ..... 97.1117M. Дои:10.1007 / s00114-010-0729-0. PMID 21060984.
- ^ Orr, C.M .; Делезене; Scott, J.E .; Tocheri, M.W .; Шварц, Г. (2007). «Сравнительный метод и вывод систем доставки яда у ископаемых млекопитающих». Журнал палеонтологии позвоночных. 27 (2): 541–546. Дои:10.1671 / 0272-4634 (2007) 27 [541: TCMATI] 2.0.CO; 2.
- ^ Vaeth, R.H .; Россман, Д.А.; Шуп, W. (1985). «Наблюдения за морфологией поверхности зубов у змей». Журнал герпетологии. 19 (1): 20–26. Дои:10.2307/1564416. JSTOR 1564416.
- ^ Беллэрс, A.D'A. (1949). "Наблюдения за мордой Варанус, и сравнение с другими ящерицами и змеями ». Журнал анатомии. 83 (2): 116–146. ЧВК 1273152. PMID 17105074.
- ^ Абдель-Кадер, Т.Г .; Ali, R.S .; Ибрагим, Н. М. (2011). "Черепные нервы Mabuya quinquetaeniata III: Нервус тройничного нерва " (PDF). Журнал наук о жизни. 8 (4): 650–669.
- ^ Leitch, D.B .; Катания, К. (2012). «Строение, иннервация и ответные свойства покровных органов чувств крокодилов». Журнал экспериментальной биологии. 215 (23): 4217–4230. Дои:10.1242 / jeb.076836. ЧВК 4074209. PMID 23136155.
- ^ Бюхерль, В. (1968). "Вступление". In Bücherl, W .; Бакли, E.E .; Деулофеу В. (ред.). Ядовитые животные и их яды. 1. Нью-Йорк: Academic Press. С. 9–12. Дои:10.1016 / B978-1-4832-2949-2.50006-0. ISBN 9781483229492.
- ^ Fry, B.G .; Wroe, S .; Teeuwisse, W .; van Osch, M.J.P .; Морено, К .; Ingle, J .; McHenry, C .; Феррара, Т .; Clausen, P .; Scheib, H .; Winter, K.L .; Greisman, L .; Roelants, K .; van der Weerd, L .; Clemente, C.J .; Гианнакис, Э. (2009). "Яд играет центральную роль в борьбе с хищниками. Varanus komodoensis (Дракон Комодо) и вымерший гигант Варан (мегалания) приск". Труды Национальной академии наук Соединенных Штатов Америки. 106 (22): 8969–8974. Bibcode:2009PNAS..106.8969F. Дои:10.1073 / pnas.0810883106. ЧВК 2690028. PMID 19451641.
- ^ Ligabue-Braun, R .; Verli, H .; Карлини, C.R. (2012). «Ядовитые млекопитающие: обзор». Токсикон. 59 (7): 680–695. Дои:10.1016 / j.toxicon.2012.02.012. PMID 22410495.
- ^ Weinstein, S.A .; Smith, T.L .; Кардонг, К. (2009). "Ядовитые железы рептилий: форма, функции и будущее" (PDF). В Mackessy, S.P. (ред.). Справочник по ядам и токсинам рептилий. Бока-Ратон: CRC Press. С. 65–91.
- ^ Горис, Р. (2011). «Инфракрасные органы змей: неотъемлемая часть зрения». Журнал герпетологии. 45 (1): 2–14. Дои:10.1670/10-238.1.
- ^ Krochmal, A.R .; Баккен, Г.С.; LaDuc, T.J. (2004). «Тепло на кухне эволюции: эволюционные взгляды на функции и происхождение лицевых ямок змеевиков (Viperidae: Crotalinae)». Журнал экспериментальной биологии. 207 (24): 4231–4238. Дои:10.1242 / jeb.01278. PMID 15531644.
- ^ Benoit, J .; Abdala, F .; Manger, P.R .; Рубидж, Б.С. (2016). «Шестое чувство у предшественников млекопитающих: изменчивость теменного отверстия и эволюция шишковидной железы у южноафриканских пермо-триасовых эутериодонтовых терапсидов». Acta Palaeontologica Polonica. 61 (4): 777–789. Дои:10.4202 / app.00219.2015.
- ^ Андерсон, Дж. М. (1977). «Преемственность микрофлоры: выводы и обсуждение». Обзор пермской палинологии Гондваны с особым упором на бассейн Северного Кару в Южной Африке. Воспоминания о ботаническом исследовании Южной Африки. 41. С. 42–58.
| |||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||



