Кривая диссоциации кислород-гемоглобин - Oxygen–hemoglobin dissociation curve
Эта статья нужны дополнительные цитаты для проверка. (Март 2009 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
В кривая диссоциации кислород-гемоглобин, также называемый кривая диссоциации оксигемоглобина или же кривая диссоциации кислорода (ODC), это изгиб который отображает долю гемоглобин в его насыщенном (кислород -загрузка) на вертикальной оси против преобладающего напряжение кислорода по горизонтальной оси. Эта кривая является важным инструментом для понимания того, как наша кровь переносит и выделяет кислород. В частности, кривая диссоциации оксигемоглобина соотносит насыщение кислородом (SО2) и частичное давление кислорода в крови (PО2), и определяется так называемым «сродством гемоглобина к кислороду»; то есть насколько легко гемоглобин поглощает и выделяет кислород молекулы в жидкость, которая его окружает.

Фон
Гемоглобин (Hb) является основным транспортным средством для перевозки кислород в кровь. Каждая молекула гемоглобина может нести четыре молекулы кислорода. Эти молекулы кислорода связываются с утюг из гем протезная группа.[1]
Когда гемоглобин не имеет связанного кислорода и не связан углекислый газ, имеет несвязанную конформацию (форму). Связывание первой молекулы кислорода вызывает изменение формы гемоглобина, что увеличивает его способность связываться с тремя другими молекулами кислорода.
В присутствии растворенного диоксида углерода pH изменений крови; это вызывает другое изменение формы гемоглобина, которое увеличивает его способность связывать углекислый газ и снижает его способность связывать кислород. С потерей первой молекулы кислорода и связыванием первой молекулы диоксида углерода происходит еще одно изменение формы, которое дополнительно снижает способность связывать кислород и увеличивает способность связывать диоксид углерода. Кислород, связанный с гемоглобином, попадает в плазму крови и всасывается в ткани, а углекислый газ в тканях связан с гемоглобином.
в легкие имеет место обратный процесс. С потерей первой молекулы углекислого газа форма снова изменяется, что облегчает высвобождение трех других углекислых газов.
Кислород также переносится растворенным в крови. плазма, но в гораздо меньшей степени. Гемоглобин содержится в красные кровяные тельца. Гемоглобин высвобождает связанный кислород, когда угольная кислота присутствует, как и в тканях. в капилляры, куда углекислый газ Кислород, связанный с гемоглобином, выделяется в плазму крови и всасывается тканями.
Сколько из этой емкости заполнено кислородом в любой момент, называется насыщение кислородом. Выраженное в процентах, насыщение кислородом - это отношение количества кислорода, связанного с гемоглобином, к кислородной способности гемоглобина. Переносимость кислорода гемоглобином определяется типом гемоглобина, присутствующего в крови. Количество кислорода, связанного с гемоглобином в любое время, в значительной степени связано с частичное давление кислорода, которому подвергается гемоглобин. В легких на альвеолярно-капиллярный интерфейс парциальное давление кислорода обычно высокое, и поэтому кислород легко связывается с присутствующим гемоглобином. Поскольку кровь циркулирует в других тканях тела, в которых парциальное давление кислорода меньше, гемоглобин выделяет кислород в ткань, потому что гемоглобин не может поддерживать свою полную способность связывать кислород в присутствии более низких парциальных давлений кислорода.
Сигмовидная форма

Кривая обычно лучше всего описывается сигмовидный сюжет, используя формулу вида:

Молекула гемоглобина может обратимым способом связывать до четырех молекул кислорода.
Форма кривой является результатом взаимодействия связанных молекул кислорода с входящими молекулами. Связывание первой молекулы затруднено. Однако это облегчает связывание второго, третьего и четвертого, это связано с индуцированным конформационным изменением структуры молекулы гемоглобина, вызванным связыванием молекулы кислорода.
В самом простом виде кривая диссоциации оксигемоглобина описывает соотношение между парциальным давлением кислорода (ось x) и насыщением кислородом (ось y). Сродство гемоглобина к кислороду увеличивается по мере связывания последовательных молекул кислорода. По мере увеличения парциального давления кислорода связывается больше молекул, пока не будет достигнуто максимальное количество, которое может быть связано. По мере приближения к этому пределу происходит очень небольшое дополнительное связывание, и кривая выравнивается, поскольку гемоглобин насыщается кислородом. Следовательно, кривая имеет сигмоидальную или S-образную форму. При давлениях выше примерно 60 мм рт. Ст. Стандартная кривая диссоциации является относительно плоской, что означает, что содержание кислорода в крови существенно не изменяется даже при значительном повышении парциального давления кислорода. Чтобы получить больше кислорода к тканям, потребуется переливание крови для увеличения количества гемоглобина (и, следовательно, способность переносить кислород) или дополнительного кислорода, который увеличивает растворенный в плазме кислород, хотя связывание кислорода с гемоглобином продолжается до некоторой степени для давления около 50 мм рт.ст., когда парциальное давление кислорода уменьшается в этой крутой области кривой, кислород легко выгружается в периферические ткани, поскольку сродство гемоглобина уменьшается. Парциальное давление кислорода в крови, при котором гемоглобин насыщается на 50%, обычно около 26,6 мм рт. Ст. (3,5 кПа) для здорового человека известен как п50. P50 является стандартной мерой сродства гемоглобина к кислороду. При наличии заболевания или других состояний, которые изменяют сродство гемоглобина к кислороду и, следовательно, сдвигают кривую вправо или влево, P50 изменяется соответственно. Повышенный P50 указывает на сдвиг стандартной кривой вправо, что означает, что необходимо большее парциальное давление для поддержания 50% насыщения кислородом. Это указывает на пониженное сродство. И наоборот, более низкий P50 указывает на сдвиг влево и более высокое сродство.
Участок "плато" кривой диссоциации оксигемоглобина - это диапазон, который существует в легочных капиллярах (минимальное снижение переносимого кислорода до тех пор, пока p (O2) падает на 50 мм рт. ст.).
«Крутой» участок кривой диссоциации оксигемоглобина - это диапазон, который существует в системных капиллярах (небольшое падение в системных капиллярах p (O2) может привести к высвобождению большого количества кислорода для метаболически активных клеток).
Чтобы увидеть относительное сродство каждого последующего кислорода по мере удаления / добавления кислорода из / в гемоглобин из кривой, сравните относительное увеличение / уменьшение p (O2), необходимого для соответствующего увеличения / уменьшения s (O2).
Факторы, влияющие на стандартную кривую диссоциации
На силу связывания кислорода с гемоглобином влияет несколько факторов. Эти факторы сдвигают или изменяют форму кривой диссоциации оксигемоглобина. Сдвиг вправо указывает на то, что исследуемый гемоглобин имеет пониженное сродство к кислороду. Это затрудняет связывание гемоглобина с кислородом (требуется более высокое парциальное давление кислорода для достижения такого же насыщения кислородом), но облегчает гемоглобину высвобождение связанного с ним кислорода. Эффект этого сдвига кривой вправо увеличивает парциальное давление кислорода в тканях, когда это больше всего необходимо, например, во время физических упражнений или геморрагического шока. Напротив, кривая смещается влево из-за противоположных этим условиям. Этот сдвиг влево указывает на то, что исследуемый гемоглобин имеет повышенное сродство к кислороду, так что гемоглобин легче связывает кислород, но разгружает его более неохотно. Сдвиг кривой влево является признаком повышенного сродства гемоглобина к кислороду (например, в легких). Точно так же сдвиг вправо показывает снижение сродства, как это могло бы появиться при повышении температуры тела, ионов водорода, 2,3-бисфосфоглицерат (2,3-БПГ) концентрация или концентрация углекислого газа.
| Факторы контроля | Изменять | Сдвиг кривой |
|---|---|---|
| Температура | ↑ | → |
| ↓ | ← | |
| 2,3-БПГ | ↑ | → |
| ↓ | ← | |
| pCO2 | ↑ | → |
| ↓ | ← | |
| Кислотность [H+] | ↑ | → |
| ↓ | ← |
Примечание:
- Левый сдвиг: выше O2 близость
- Сдвиг вправо: нижний O2 близость
- гемоглобин плода имеет более высокое содержание O2 сродство, чем у взрослого гемоглобина; в первую очередь из-за значительно сниженного сродства к 2,3-бисфосфоглицерат .
Причины смещения вправо можно запомнить, используя мнемонический, "КАДЕТ, лицом вправо! "для CО2, АСид, 2,3-DPG,[Примечание 1] Eупражнения и Ттемпература.[2] Факторы, которые сдвигают кривую диссоциации кислорода вправо, - это те физиологические состояния, при которых тканям требуется больше кислорода. Например, во время упражнений у мышц более высокая скорость метаболизма, и, следовательно, им нужно больше кислорода, они производят больше углекислого газа и молочной кислоты, и их температура повышается.
pH
Снижение pH (увеличение ЧАС+ концентрация ионов) сдвигает стандартную кривую вправо, а увеличение - влево. Это происходит потому, что при большем ЧАС+ Концентрация ионов, различные аминокислотные остатки, такие как гистидин 146, существуют преимущественно в протонированной форме, что позволяет им образовывать ионные пары, которые стабилизируют дезоксигемоглобин в состоянии T.[3] Состояние T имеет более низкое сродство к кислороду, чем состояние R, поэтому с повышенной кислотностью гемоглобин связывает меньше O2 для данного PO2 (и более H+). Это известно как Эффект Бора.[4] Снижение общей связывающей способности гемоглобина по отношению к кислороду (т. Е. Смещение кривой вниз, а не только вправо) из-за снижения pH называется корневой эффект. Это наблюдается у костистой рыбы. Сродство связывания гемоглобина с O2 является наибольшим при относительно высоком pH.
Углекислый газ
Углекислый газ влияет на кривую двумя способами. Во-первых, CO2 причины накопления карбамино соединения, которые образуются в результате химических взаимодействий, которые связываются с образованием гемоглобина карбаминогемоглобин . CO2 считается Аллостерическая регуляция поскольку ингибирование происходит не в месте связывания гемоглобина.[5] Во-вторых, это влияет внутриклеточный pH из-за образования бикарбонат-иона. Образование карбаминогемоглобина стабилизирует гемоглобин в Т-состоянии за счет образования ионных пар.[3] Только около 5–10% от общего количества CO2 содержимое крови транспортируется в виде карбаминовых соединений, тогда как (80–90%) транспортируется в виде ионов бикарбоната, а небольшое количество растворяется в плазме. Образование бикарбонат-иона будет высвобождать протон в плазму, снижая pH (повышенную кислотность), что также сдвигает кривую вправо, как обсуждалось выше; низкий CO2 уровни в кровотоке приводят к высокому pH и, таким образом, обеспечивают более оптимальные условия связывания гемоглобина и O2. Это физиологически предпочтительный механизм, поскольку гемоглобин будет терять больше кислорода, поскольку концентрация углекислого газа резко возрастает, когда тканевое дыхание происходит быстро и кислород нуждается.[6][7]
2,3-БПГ
2,3-Бисфосфоглицерат или 2,3-BPG (ранее назывался 2,3-дифосфоглицерат или 2,3-DPG - ссылка?) - это органофосфат, образующийся в красные кровяные тельца в течение гликолиз и это сопряженное основание из 2,3-бисфосфоглицериновая кислота. Продукция 2,3-BPG, вероятно, является важным адаптивным механизмом, потому что продукция увеличивается в нескольких условиях в присутствии уменьшенной периферической ткани O2 доступность, например гипоксемия, хроническое заболевание легких, анемия, и хроническая сердечная недостаточность, среди прочего. Высокие уровни 2,3-BPG смещают кривую вправо (как в детстве), в то время как низкие уровни 2,3-BPG вызывают сдвиг влево, наблюдаемый в таких состояниях, как септический шок, и гипофосфатемия.[4] В отсутствие 2,3-БПГ повышается сродство гемоглобина к кислороду. 2,3-БПГ действует как гетероаллостерический эффектор гемоглобина, снижая сродство гемоглобина к кислороду за счет связывания преимущественно с дезоксигемоглобином. Повышенная концентрация БПГ в красных кровяных тельцах способствует формированию Т (напряженного или напряженного) состояния гемоглобина с низким сродством, поэтому кривая связывания кислорода смещается вправо.
Температура
Повышение температуры сдвигает ODC вправо. Если температура повышается, то же самое, то насыщение кислородом уменьшается, потому что связь между железом в и денатурируется. Точно так же с повышением температуры также увеличивается парциальное давление кислорода. Таким образом, процент насыщения гемоглобина будет меньше для тех же или более высокое парциальное давление кислорода. Таким образом, любая точка кривой сместится вправо (из-за повышенного парциального давления кислорода) и вниз (из-за ослабления связь). Следовательно, сдвиг кривой вправо.[8]
![{ displaystyle { ce {[O2]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8f8f06466530fdc7eb5967cec6cbfed6a59582aa "Концентрация кислорода")



Монооксид углерода
Гемоглобин связывается с монооксид углерода В 210 раз легче, чем с кислородом.[4] Из-за более высокого сродства гемоглобина к окиси углерода, чем к кислороду, окись углерода является очень успешным конкурентом, который вытесняет кислород даже при крохотных парциальных давлениях. Реакция HbO2 + CO → HbCO + O2 практически необратимо вытесняет молекулы кислорода, образующие карбоксигемоглобин; Связывание оксида углерода с железным центром гемоглобина намного сильнее, чем у кислорода, и сайт связывания остается заблокированным до конца жизненного цикла затронутого эритроцита.[9] При повышенном уровне окиси углерода человек может сильно пострадать в тканях гипоксия при сохранении нормального рО2 потому что карбоксигемоглобин не переносит кислород к тканям.
Последствия метгемоглобинемии
Метгемоглобинемия представляет собой форму аномального гемоглобина, при которой центр железа был окислен из железо +2 степень окисления (нормальная форма) к железо +3 государственный. Это вызывает сдвиг влево кривой диссоциации кислородного гемоглобина, поскольку любой остаточный гем с оксигенированным двухвалентным железом (состояние +2) не может выгрузить связанный кислород в ткани (поскольку 3+ железо нарушает кооперативность гемоглобина), тем самым увеличивая его сродство с кислородом. . Однако метгемоглобин имеет повышенное сродство к цианид, и поэтому полезен при лечении отравление цианидом. В случае случайного проглатывания введение нитрит (Такие как амилнитрит ) может использоваться для преднамеренного окисления гемоглобина и повышения уровня метгемоглобина, восстанавливая функционирование цитохромоксидаза. Нитрит также действует как вазодилататор, способствуя доставке кислорода клетками, а добавление соли железа обеспечивает конкурентное связывание свободного цианида как биохимически инертного гексацианоферрат (III) ион, [Fe (CN)6]3−. Альтернативный подход предполагает введение тиосульфат, тем самым превращая цианид в тиоцианат, SCN−, который выводится через почки. Метгемоглобин также образуется в небольших количествах, когда диссоциация оксигемоглобина приводит к образованию метгемоглобина и супероксид, O2−, вместо обычных продуктов. Супероксид - это свободный радикал и вызывает биохимические повреждения, но нейтрализуется действием фермента супероксиддисмутаза.
Эффекты ITPP
Мио-инозитол триспирофосфат (ITPP), также известный как OXY111A, является инозитолфосфат что вызывает сдвиг вправо в кривой диссоциации кислородного гемоглобина через аллостерическая модуляция гемоглобина в красных кровяных тельцах. Это экспериментальный препарат, предназначенный для сокращения тканей гипоксия. Эффект длится примерно до тех пор, пока пораженные эритроциты остаются в обращении.
Гемоглобин плода
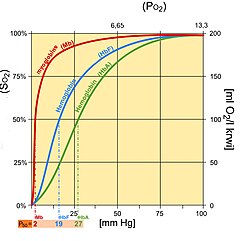
Гемоглобин плода (HbF) структурно отличается от нормального гемоглобин взрослых (HbA), что дает HbF более высокое сродство к кислороду, чем HbA. HbF состоит из двух альфа и двух гамма цепей, тогда как HbA состоит из двух альфа и двух бета цепей. Кривая диссоциации плода смещена влево относительно кривой для нормального взрослого человека из-за этих структурных различий.
Обычно артериальное давление кислорода у плода ниже, чем у взрослых. Следовательно, более высокое сродство к связыванию кислорода требуется при более низких уровнях парциального давления у плода, чтобы обеспечить диффузию кислорода через плацента. В плаценте наблюдается более высокая концентрация 2,3-БПГ 2,3-BPG легко связывается с бета-цепями, а не с альфа-цепями. В результате 2,3-BPG более прочно связывается с гемоглобином взрослого человека, заставляя HbA выделять больше кислорода для поглощения плодом, чей HbF не зависит от 2,3-BPG.[10] Затем HbF доставляет связанный кислород к тканям с еще более низким парциальным давлением, откуда он может высвобождаться.
Смотрите также
Примечания
- ^ 2,3-DPG - это сокращение от 2,3-Dяпхофограммлицериновая кислота, устаревшее название 2,3-BPG
Рекомендации
- ^ Ахерн, Кевин; Раджагопал, Индира; Тан, Таралин (2017). Биохимия - бесплатно для всех (PDF) (1.2 изд.). NC: Creative Commons.
- ^ «Медицинская мнемоника». LifeHugger. Получено 2009-12-19.
- ^ а б Ленингер. Принципы биохимии (6-е изд.). п. 169.
- ^ а б c Жакес, Джон (1979). Респираторная физиология. Макгроу-Хилл. С. 156–175.
- ^ Ахерн, Кевин; Раджагопал, Индира; Тан, Таралин (5 августа 2017 г.). Биохимия - бесплатно для всех (1.2 изд.). NC-Creative Commons. п. 370.
- ^ Ахерн, Кевин; Раджагопал, Индира; Тан, Таралин (5 августа 2017 г.). Биохимия - бесплатно для всех (1.2 изд.). NC-Creative Commons. п. 134.
- ^ Донна, Ларсон (2017). Клиническая химия: основы и лабораторные методы. Сент-Луис, Миссури: Эльзевир. п. 226. ISBN 978-1-4557-4214-1.
- ^ Шмидт-Нильсен (1997). Физиология животных: адаптация и окружающая среда. Издательство Кембриджского университета. ISBN 0521570980.
- ^ Коц, Джон (август 2012). Химия и химическая реакционная способность (8-е изд.). Cengage Learning. п. 1032. ISBN 978-1133420071. Получено 2015-07-01.
- ^ Иллюстрированный обзор Липпинкотта: биохимия, 4-е издание. Северная Америка: Липпинкотт, Уильямс и Уилкинс. 2007. С. 24–35. ISBN 978-0-7817-6960-0.
внешняя ссылка
- Носек, Томас М. "Раздел 4 / 4ч5 / с4ч5_18". Основы физиологии человека. Архивировано из оригинал на 24.03.2016.
- Интерактивная кривая диссоциации оксигемоглобина
- Моделирование параметров CO2, pH и температура на кривой диссоциации кислород – гемоглобин (сдвиг влево или вправо)