Сердечная физиология - Cardiac physiology
Сердечная физиология или же функция сердца исследование здоровой, неповрежденной функции сердце: вовлечение кровотока; миокард структура; система электропроводности сердца; сердечный цикл и сердечный выброс, а также то, как они взаимодействуют и зависят друг от друга.
Кровоток
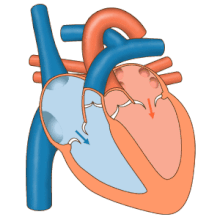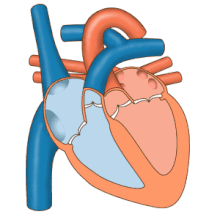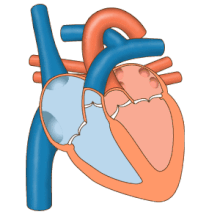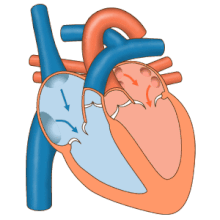


Сердце работает как насос и действует как двойной насос в сердечно-сосудистая система для обеспечения непрерывной циркуляции крови по всему телу. Этот тираж включает Систематическая циркуляция и легочное кровообращение. Оба контура транспортируют кровь, но их также можно увидеть с точки зрения переносимых ими газов. Легочная циркуляция собирает кислород из легких и доставляет углекислый газ для выдоха. Системный контур транспортирует кислород к телу и возвращает относительно обескислороженную кровь и углекислый газ в легочный контур.[1]
Кровь течет через сердце в одном направлении, от предсердий к желудочкам, и выходит через легочная артерия в малый круг кровообращения, а аорта в системный кровоток. Легочная артерия (также ствол) разветвляется на левую и правую. легочные артерии для снабжения каждого легкого. Кровь не может течь назад (регургитация ) трикуспидальным, двустворчатым, аортальным и легочным клапанами.
Функция правое сердце, заключается в сборе обескислороженной крови в правом предсердии из тела через верхнюю полую вену, нижнюю полую вену и из коронарного синуса и перекачивание ее через трикуспидальный клапан через Правый желудочек, через полулунный легочной клапан и в легочную артерию в малом круге кровообращения, где углекислый газ может быть обменялся для кислорода в легких. Это происходит через пассивный процесс распространение. в левое сердце насыщенная кислородом кровь возвращается в левое предсердие через легочную вену. Затем он закачивается в левый желудочек через двустворчатый клапан и в аорту для системного кровообращения. Со временем в системных капиллярах происходит обмен с тканевой жидкостью и клетками тела; кислород и питательные вещества поставляются клеткам для их метаболизма и обмениваются на углекислый газ и отходы[1] В этом случае кислород и питательные вещества выходят из системных капилляров для использования клетками в их метаболических процессах, а углекислый газ и продукты жизнедеятельности попадают в кровь.[1]
Желудочки сильнее и толще, чем предсердия, а мышечная стенка, окружающая левый желудочек, толще, чем стенка, окружающая правый желудочек, из-за большей силы, необходимой для перекачивания крови через Систематическая циркуляция. Атрия облегчить кровообращение, прежде всего, обеспечивая непрерывный венозный поток к сердцу, предотвращая инерцию прерывистого венозного кровотока, которая в противном случае возникла бы при каждой систоле желудочков.[2]
Сердечная мышца
Ткань сердечной мышцы имеет авторитмичность, уникальная способность инициировать потенциал сердечного действия с фиксированной скоростью - быстрое распространение импульса от ячейки к ячейке для запуска сокращение всего сердца. Эта авторитмичность все еще модулируется эндокринный и нервная система.[1]
Есть два типа клеток сердечной мышцы: кардиомиоциты которые обладают способностью легко сокращаться, и модифицированные кардиомиоциты клетки кардиостимулятора проводящей системы. Кардиомиоциты составляют основную часть (99%) клеток предсердий и желудочков. Эти сократительные клетки реагируют на импульсы потенциал действия от клеток кардиостимулятора и отвечают за сокращения, которые прокачивают кровь по телу. Клетки водителя ритма составляют всего (1% клеток) и образуют проводящую систему сердца. Как правило, они намного меньше сократительных клеток и имеют мало миофибриллы или же миофиламенты что означает, что у них ограниченная сократимость. Их функция во многом схожа с нейроны.[1] В связка Его и Волокна Пуркинье представляют собой специализированные кардиомиоциты, которые функционируют в проводящей системе.
Строение сердечной мышцы

Кардиомиоциты, значительно короче и имеют меньший диаметр, чем скелетные миоциты. Сердечная мышца (как и скелетная мышца) характеризуется: полосы - полосы темных и светлых полос в результате организованного расположения миофиламентов и миофибриллы в саркомер по длине клетки. Т (поперечные) канальцы глубокие инвагинации от сарколемма (клеточная мембрана), которые проникают в клетку, позволяя электрическим импульсам проникать внутрь. В сердечной мышце Т-канальцы находятся только в Z-линии.[1] Когда потенциал действия заставляет клетки сокращаться, кальций освобожден от саркоплазматический ретикулум клеток, а также Т-канальцев. Высвобождение кальция вызывает скольжение актин и миозин фибриллы, приводящие к сокращению.[3] Обильный запас митохондрии обеспечить энергию сокращений. Обычно кардиомиоциты имеют одно центральное ядро, но могут также иметь два или более.[1]
Клетки сердечной мышцы свободно разветвляются и связаны соединениями, известными как вставные диски которые помогают синхронизированному сокращению мышцы.[4] В сарколемма (мембрана) из соседних клеток связываются вместе на вставленных дисках. Они состоят из десмосомы, специализированные ссылки протеогликаны, узкие стыки, и большое количество щелевые соединения которые позволяют ионам проходить между клетками и помогают синхронизировать сокращение. Межклеточный соединительная ткань также помогает прочно связывать клетки вместе, чтобы противостоять силам сжатия.[1]
Сердечная мышца подвергается аэробного дыхания паттерны, в первую очередь метаболизм липидов и углеводов. Кислород из легких прикрепляется к гемоглобин и также хранится в миоглобин, так что имеется обильный запас кислорода. Липиды, и гликоген также хранятся в саркоплазма и они расщепляются митохондриями, чтобы высвободить АТФ. Клетки подвергаются схватки подергивающего типа с длинным рефрактерные периоды с последующими короткими периодами расслабления, когда сердце наполняется кровью для следующего цикла.[1]
Электрическая проводимость

Не очень хорошо известно, как электрический сигнал движется в предсердиях. Вроде бы радиально движется, но Связка Бахмана и коронарный синус мышцы играют роль в проводимости между двумя предсердиями, которые имеют почти одновременное систола.[5][6][7] Находясь в желудочках, сигнал передается специальной тканью, называемой Волокна Пуркинье которые затем передают электрический заряд миокард.[8]
Если эмбриональные клетки сердца разделить в чашку Петри и сохранить в живых, каждая из них будет способна генерировать собственный электрический импульс, за которым следует сокращение. Когда две независимо бьющиеся клетки эмбриональной сердечной мышцы помещаются вместе, клетка с более высокой собственной частотой задает темп, и импульс распространяется от более быстрой клетки к более медленной, вызывая сокращение. Чем больше ячеек соединяется вместе, тем скорость передачи берет на себя самая быстрая ячейка. Полностью развитое сердце взрослого человека сохраняет способность генерировать собственный электрический импульс, инициируемый самыми быстрыми клетками, как часть системы сердечной проводимости. Компоненты системы сердечной проводимости включают предсердный и желудочковый синцитий, синоатриальный узел, атриовентрикулярный узел, связка Его (атриовентрикулярный пучок), связка ветвей, и клетки Пуркинье.[1]
Синоатриальный (SA) узел

Нормальный синусовый ритм устанавливается синоатриальный (SA) узел, сердце кардиостимулятор. Узел SA - это специализированная группа кардиомиоцитов в верхней и задней стенках правого предсердия, очень близко к отверстию верхняя полая вена. Узел SA имеет самую высокую скорость деполяризация.[1]
Этот импульс распространяется от своего инициирования в узле SA через предсердия через специализированные межузловые пути к сократительным клеткам миокарда предсердий и атриовентрикулярному узлу. Межузловые пути состоят из трех полос (передней, средней и задней), которые ведут непосредственно от узла SA к следующему узлу проводящей системы, атриовентрикулярному узлу. Импульс проходит между этими двумя узлами примерно за 50 мс (миллисекунд). Относительная важность этого пути обсуждалась, поскольку импульс достигнет атриовентрикулярного узла, просто следуя клеточному пути через сократительные клетки миокарда в предсердиях. Кроме того, существует специальный путь, называемый пучком Бахмана или межпредсердной связкой, который проводит импульс непосредственно из правого предсердия в левое. Независимо от пути, когда импульс достигает атриовентрикулярной перегородки, соединительная ткань сердечного скелета препятствует распространению импульса в клетки миокарда желудочков, за исключением атриовентрикулярного узла.[1] Электрическое событие - волна деполяризации - запускает мышечное сокращение. Волна деполяризации начинается в правом предсердии, и импульс распространяется через верхние части обоих предсердий, а затем вниз через сократительные клетки. Затем сократительные клетки начинают сокращение от верхней части предсердия к нижней, эффективно перекачивая кровь в желудочки.[1]
Атриовентрикулярный (АВ) узел
Атриовентрикулярный (АВ) узел - это второй кластер специализированных миокардиальных проводящих клеток, расположенный в нижней части правого предсердия в пределах атриовентрикулярной перегородки. Перегородка препятствует распространению импульса непосредственно на желудочки, не проходя через АВ-узел. Существует критическая пауза перед тем, как АВ-узел деполяризуется и передает импульс атриовентрикулярному пучку. Эта задержка передачи частично объясняется небольшим диаметром ячеек узла, которые замедляют импульс. Кроме того, проводимость между узловыми клетками менее эффективна, чем между проводящими клетками. Эти факторы означают, что импульс проходит через узел примерно за 100 мс. Эта пауза имеет решающее значение для работы сердца, так как позволяет кардиомиоцитам предсердий завершить свое сокращение, которое перекачивает кровь в желудочки до того, как импульс будет передан клеткам самого желудочка. При экстремальной стимуляции узлом SA, узел AV может передавать импульсы с максимальной скоростью 220 в минуту. Таким образом устанавливается типичная максимальная частота пульса у здорового молодого человека. Поврежденные сердца или сердца, стимулированные лекарствами, могут сокращаться с большей скоростью, но с такой скоростью сердце больше не может эффективно перекачивать кровь.[1]
Пучок Гиса, ветви пучка и волокна Пуркинье
Возникающий из АВ-узла пучок Гиса проходит через межжелудочковую перегородку, прежде чем разделиться на две части. связка ветвей, обычно называемые левой и правой ветвями пучка. Левая ножка пучка Гиса имеет два пучка. Левая ножка пучка Гиса снабжает левый желудочек, а правая ножка пучка Гиса - правый желудочек. Поскольку левый желудочек намного больше правого, левая ножка пучка Гиса также значительно больше правой. Части правой ножки пучка Гиса находятся в зоне модератора и снабжают правые сосочковые мышцы. Из-за этой связи каждая сосочковая мышца получает импульс примерно в одно и то же время, поэтому они начинают сокращаться одновременно, незадолго до остальной части сократительных клеток миокарда желудочков. Считается, что это способствует развитию напряжения на сухожильных хордах до сокращения правого желудочка. Слева нет соответствующей группы модераторов. Обе ветви пучка опускаются вниз и достигают вершины сердца, где соединяются с волокнами Пуркинье. Этот проход занимает примерно 25 мс.[1]
Волокна Пуркинье - это дополнительные проводящие волокна миокарда, которые передают импульс сократительным клеткам миокарда в желудочках. Они проходят через миокард от верхушки сердца к атриовентрикулярной перегородке и основанию сердца. Волокна Пуркинье обладают высокой скоростью проведения, и электрический импульс достигает всех мышечных клеток желудочков примерно за 75 мс. Поскольку электрический стимул начинается с верхушки, сокращение также начинается с верхушки и распространяется к основанию сердца, подобно сдавливанию тюбика с зубной пастой снизу. Это позволяет перекачивать кровь из желудочков в аорту и легочный ствол. Общее время, прошедшее от инициирования импульса в узле SA до деполяризации желудочков, составляет примерно 225 мс.[1]
Мембранные потенциалы и движение ионов в сердечных проводящих клетках
Потенциалы действия проводящих и сокращающихся кардиомиоцитов значительно различаются. Пока натрий Na+ и калий K+ ионы играют важную роль, ионы кальция Ca2+ также важны для обоих типов клеток. В отличие от скелетных мышц и нейронов, проводящие клетки сердца не обладают стабильным потенциалом покоя. Проводящие клетки содержат ряд натрия ионные каналы которые обеспечивают нормальный и медленный приток ионов натрия, который вызывает медленное повышение мембранного потенциала от начального значения -60 мВ до примерно -40 мВ. В результате движение ионов натрия создает спонтанное деполяризация (или предпотенциальная деполяризация).[1]
На этой точке, кальциевые каналы открыть и Ca2+ попадает в клетку, деполяризуя ее с большей скоростью, пока не достигнет значения приблизительно +5 мВ. В этот момент каналы ионов кальция закрываются и калиевые каналы открытый, позволяющий отток K+ и приводит к реполяризации. Когда мембранный потенциал достигает примерно -60 мВ, K+ каналы закрываются и Na+ каналы открываются, и снова начинается препотенциальная фаза. Этот процесс наделяет сердечную мышцу авторитмичностью.[1]

Мембранные потенциалы и движение ионов в сократительных клетках сердца
Существует совершенно иной электрический паттерн, затрагивающий сократительные клетки. В этом случае происходит быстрая деполяризация, за которой следует фаза плато, а затем реполяризация. Это явление объясняет долгую рефрактерные периоды необходим для того, чтобы клетки сердечной мышцы могли эффективно перекачивать кровь, прежде чем они смогут выстрелить во второй раз. Эти сердечные миоциты обычно не инициируют свой собственный электрический потенциал, хотя они способны на это, а скорее ожидают импульса, который до них дойдет.[1]
Сократительные клетки демонстрируют гораздо более стабильную фазу покоя, чем проводящие клетки, при примерно -80 мВ для клеток в предсердиях и -90 мВ для клеток в желудочках. Несмотря на это первоначальное различие, другие компоненты их потенциалов действия практически идентичны. В обоих случаях, когда они стимулируются потенциалом действия, потенциал-управляемые каналы быстро открываются, запуская механизм деполяризации с положительной обратной связью. Этот быстрый приток положительно заряженных ионов повышает мембранный потенциал примерно до +30 мВ, после чего натриевые каналы закрываются. Период быстрой деполяризации обычно длится 3–5 мс. За деполяризацией следует фаза плато, в которой мембранный потенциал снижается относительно медленно. Это во многом связано с открытием медленного Ca2+ каналы, позволяющие Ca2+ войти в камеру, пока несколько K+ каналы открыты, что позволяет K+ выйти из камеры. Относительно длинная фаза плато длится примерно 175 мс. Как только мембранный потенциал достигает примерно нуля, Ca2+ каналы закрываются и K+ каналы открыты, позволяя K+ выйти из камеры. Реполяризация длится примерно 75 мс. В этот момент мембранный потенциал падает, пока снова не достигнет уровня покоя, и цикл повторяется. Все событие длится от 250 до 300 мс.[1]
Абсолютный рефрактерный период для сердечной сократительной мышцы длится примерно 200 мс, а относительный рефрактерный период длится примерно 50 мс, всего 250 мс. Этот продолжительный период имеет решающее значение, поскольку сердечная мышца должна сокращаться, чтобы эффективно перекачивать кровь, и сокращение должно следовать за электрическими событиями. Без продолжительных рефрактерных периодов в сердце возникли бы преждевременные сокращения, которые были бы несовместимы с жизнью.[1]

(а) Имеется длительная фаза плато из-за притока ионов кальция. Увеличенный рефрактерный период позволяет клетке полностью сжаться до того, как может произойти другое электрическое событие.
(b) Потенциал действия сердечной мышцы сравнивается с потенциалом действия скелетных мышц.[1]
Ионы кальция
Ионы кальция играют две важнейшие роли в физиологии сердечной мышцы. Их приток через медленные кальциевые каналы объясняет длительную фазу плато и период абсолютной рефрактерности. Ионы кальция также сочетаются с регуляторным белком. тропонин в тропониновый комплекс. Обе роли позволяют миокарду функционировать должным образом.[1]
Примерно 20 процентов кальция, необходимого для сокращения, поступает за счет притока кальция.2+ во время фазы плато. Оставшийся Ca2+ для сокращения высвобождается из хранилища в саркоплазматическом ретикулуме.[1]
Сравнительные показатели стрельбы проводящей системы
Паттерн предпотенциальной или спонтанной деполяризации, за которой следует только что описанная быстрая деполяризация и реполяризация, виден в узле SA и некоторых других проводящих клетках сердца. Поскольку узел SA является кардиостимулятором, он достигает порога быстрее, чем любой другой компонент проводящей системы. Это инициирует распространение импульсов на другие проводящие клетки. Узел SA без нервного или эндокринного контроля будет инициировать сердечный импульс примерно 80–100 раз в минуту. Хотя каждый компонент проводящей системы способен генерировать свой собственный импульс, скорость постепенно снижается от узла SA к волокнам Пуркинье. Без узла SA, узел AV генерировал бы частоту сердечных сокращений 40-60 ударов в минуту. Если атриовентрикулярный узел был заблокирован, атриовентрикулярный пучок запускался бы со скоростью примерно 30-40 импульсов в минуту. Пучковые ветви будут иметь внутреннюю частоту 20–30 импульсов в минуту, а волокна Пуркинье будут срабатывать со скоростью 15–20 импульсов в минуту. В то время как несколько исключительно тренированных спортсменов-аэробистов демонстрируют частоту пульса в состоянии покоя в диапазоне 30–40 ударов в минуту (самый низкий зарегистрированный показатель составляет 28 ударов в минуту для Мигель Индурайн, велосипедист) - для большинства людей частота ниже 50 ударов в минуту указывает на состояние, называемое брадикардией. В зависимости от конкретного человека, когда показатели упадут намного ниже этого уровня, сердце будет неспособно поддерживать адекватный кровоток к жизненно важным тканям, что поначалу приведет к уменьшению потери функций систем, потере сознания и, в конечном итоге, к смерти.[1]
Сердечный цикл

Период времени, который начинается с сокращения предсердий и заканчивается расслаблением желудочков, известен как сердечный цикл. Период сокращения, который испытывает сердце, перекачивая кровь в кровоток, называется систолой. Период расслабления, который наступает при наполнении камер кровью, называется диастолой. И предсердия, и желудочки подвергаются систоле и диастоле, и важно, чтобы эти компоненты тщательно регулировались и координировались, чтобы обеспечить эффективную перекачку крови в организм.[1]

Давление и расход
Жидкости перемещаются из областей с высоким давлением в области с более низким давлением. Соответственно, когда камеры сердца расслаблены (диастола), кровь будет течь в предсердия из-за более высокого давления в венах. Когда кровь течет в предсердия, давление будет расти, поэтому кровь сначала будет пассивно перемещаться из предсердий в желудочки. Когда потенциал действия вызывает сокращение мышц предсердий (систола предсердий), давление внутри предсердий повышается еще больше, перекачивая кровь в желудочки. Во время систолы желудочков в желудочках повышается давление, перекачивая кровь в легочный ствол из правого желудочка и в аорту из левого желудочка.[1]
Фазы сердечного цикла
В начале сердечного цикла и предсердия, и желудочки расслаблены (диастола). Кровь течет в правое предсердие из верхней и нижней полых вен и коронарного синуса. Кровь течет в левое предсердие из четырех легочных вен. Два атриовентрикулярных клапана, трикуспидальный и митральный клапаны, открыты, поэтому кровь беспрепятственно течет из предсердий в желудочки. Таким методом происходит примерно 70–80% наполнения желудочков. Два полулунных клапана, легочный и аортальный клапаны, закрыты, что предотвращает обратный ток крови в правый и левый желудочки из легочного ствола справа и аорты слева.[1]
Систола и диастола предсердий
Сокращение предсердий следует за деполяризацией, представленной Зубец P ЭКГ. Когда мышцы предсердий сокращаются от верхней части предсердий к атриовентрикулярной перегородке, внутри предсердий повышается давление, и кровь перекачивается в желудочки через открытые атриовентрикулярные (трехстворчатые, митральные или двустворчатые) клапаны. В начале систолы предсердий желудочки обычно заполняются примерно на 70–80% своей емкости из-за притока во время диастолы. Сокращение предсердий, также называемое «предсердным толчком», составляет оставшиеся 20–30 процентов наполнения. Систола предсердий длится примерно 100 мс и заканчивается до систолы желудочков, когда мышца предсердия возвращается в диастолу.[1]
Систола желудочков
Систола желудочков следует за деполяризацией желудочков и представлена QRS комплекс в ЭКГ. Его удобно разделить на две фазы общей продолжительностью 270 мс. В конце систолы предсердий и непосредственно перед сокращением желудочков желудочки содержат примерно 130 мл крови у отдыхающего взрослого человека в положении стоя. Этот объем известен как конечный диастолический объем (EDV) или предварительная нагрузка.[1]
Первоначально, когда мышцы желудочка сокращаются, давление крови внутри камеры повышается, но оно еще недостаточно высоко, чтобы открыть полулунные (легочные и аортальные) клапаны и выбрасывать ее из сердца. Однако артериальное давление быстро поднимается выше, чем в предсердиях, которые теперь расслаблены и находятся в диастоле. Это повышение давления заставляет кровь течь обратно к предсердиям, закрывая трикуспидальный и митральный клапаны. Поскольку кровь не выбрасывается из желудочков на этой ранней стадии, объем крови в камере остается постоянным. Следовательно, эта начальная фаза систолы желудочков известна как изоволюмическое сокращение, также называемое изоволюметрическим сокращением.[1]
Во второй фазе систолы желудочков, фазе выброса желудочков, сокращение желудочковой мышцы увеличивает давление в желудочке до такой степени, что оно превышает давление в легочном стволе и аорте. Кровь перекачивается из сердца, открывая полулунные клапаны легочной артерии и аорты. Давление, создаваемое левым желудочком, будет значительно выше, чем давление, создаваемое правым желудочком, поскольку существующее давление в аорте будет намного выше. Тем не менее, оба желудочка перекачивают одинаковое количество крови. Эта величина обозначается как ударный объем. Ударный объем обычно находится в диапазоне 70–80 мл. Поскольку систола желудочков началась с КДО, равной примерно 130 мл крови, это означает, что после сокращения в желудочке остается 50–60 мл крови. Этот объем крови известен как конечный систолический объем (КСО).[1]
Желудочковая диастола
Желудочковая релаксация или диастола следует за реполяризацией желудочков и представлена Зубец Т ЭКГ. Он также разделен на две отдельные фазы и длится примерно 430 мс.[1]
Во время ранней фазы желудочковой диастолы, когда мышца желудочка расслабляется, давление на оставшуюся кровь в желудочке начинает падать. Когда давление в желудочках падает ниже давления как в легочном стволе, так и в аорте, кровь течет обратно к сердцу, образуя дикротическую выемку (небольшой провал), наблюдаемую при измерении артериального давления. Полулунные клапаны закрываются, чтобы предотвратить обратный ток в сердце. Поскольку в этот момент атриовентрикулярные клапаны остаются закрытыми, объем крови в желудочке не изменяется, поэтому ранняя фаза желудочковой диастолы называется фазой изоволюмической желудочковой релаксации, также называемой фазой изоволюметрической желудочковой релаксации.[1]
Во второй фазе желудочковой диастолы, называемой поздней желудочковой диастолой, когда мышца желудочка расслабляется, давление на кровь внутри желудочков падает еще больше. В конце концов, давление в предсердиях падает ниже. Когда это происходит, кровь течет из предсердий в желудочки, открывая трикуспидальный и митральный клапаны. Когда давление в желудочках падает, кровь течет из основных вен в расслабленные предсердия, а оттуда - в желудочки.Обе камеры находятся в диастоле, атриовентрикулярные клапаны открыты, а полулунные клапаны остаются закрытыми. Сердечный цикл завершен.[1]
Тоны сердца
Один из простейших методов оценки состояния сердца - это Слушать к нему с помощью стетоскоп.[1] В здоровом сердце слышны только два звуки сердца, называемые S1 и S2. Первый тон сердца S1 - это звук, создаваемый закрытием атриовентрикулярных клапанов во время сокращения желудочков, и обычно его называют «lub». Второй тон сердца, S2, представляет собой звук закрытия полулунных клапанов во время желудочковой диастолы и описывается как «даб».[1] Каждый звук состоит из двух компонентов, отражающих небольшую разницу во времени, когда два клапана закрываются.[9] S2 май расколоть на два разных звука, возникающих в результате вдоха или различных клапанных или сердечных проблем.[9] Также могут присутствовать дополнительные сердечные тоны, которые вызывают ритмы галопа. А третий звук сердца, S3 обычно указывает на увеличение объема желудочковой крови. А четвертый звук сердца S4 называется предсердным галопом и вызывается звуком крови, выталкиваемой в жесткий желудочек. Совместное присутствие S3 и S4 дает четырехкратный галоп.[1]
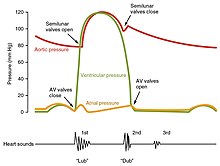
Шумы в сердце это аномальные сердечные тоны, которые могут быть патологическими или доброкачественными и бывают многих видов.[10] Шумы классифицируются по громкости от 1) самого тихого до 6) самого громкого и оцениваются по их соотношению с тоном сердца и положением в сердечном цикле.[9] Фонокардиограммы можно записывать эти звуки.[1] Шумы могут быть результатом сужения (стеноза), регургитации или недостаточности любого из основных сердечных клапанов, но они также могут быть результатом ряда других заболеваний, включая предсердный и дефекты межжелудочковой перегородки.[9] Один из примеров ропота: Все еще бормотание, который представляет собой музыкальный звук у детей, не имеет симптомов и исчезает в подростковом возрасте.[11]
Другой тип звука, трение перикарда можно услышать в случаях перикардита, когда воспаленные оболочки могут тереться.[12]
Частота сердцебиения
Частота сердечных сокращений новорожденного в состоянии покоя может составлять 120 ударов в минуту, и она постепенно снижается до зрелости, а затем постепенно снова увеличивается с возрастом. Частота сердечных сокращений взрослого в состоянии покоя колеблется в пределах 60–100 ударов в минуту. Уровни физических упражнений и физической подготовки, возраст и базальная скорость метаболизма все может повлиять на частоту сердечных сокращений. Частота сердечных сокращений спортсмена может быть ниже 60 ударов в минуту. Во время тренировки частота может составлять 150 ударов в минуту, а максимальная - от 200 до 220 ударов в минуту.[1]
Сердечно-сосудистые центры


Нормальный синусовый ритм пульса генерируется Узел SA. На это также влияет центральный факторы через сочувствующий и парасимпатический нервы[3]:116–22 из двух парных сердечно-сосудистые центры из продолговатый мозг. Активность увеличивается за счет симпатической стимуляции кардиоускорительных нервов и подавляется парасимпатической стимуляцией со стороны блуждающий нерв. Во время покоя обычно преобладает стимуляция блуждающего нерва, поскольку, если он не регулируется, узел SA инициирует синусовый ритм примерно 100 ударов в минуту.[1]
И симпатические, и парасимпатические стимулы проходят через парные сердечное сплетение у основания сердца. Без какой-либо нервной стимуляции узел SA установил бы синусовый ритм приблизительно 100 ударов в минуту. Поскольку частота отдыха значительно меньше, становится очевидным, что парасимпатическая стимуляция обычно замедляет ЧСС.[1] Кардиоускорительный центр также отправляет дополнительные волокна, формирующие сердечные нервы через симпатические ганглии (шейные ганглии плюс верхние грудные ганглии T1 – T4) к SA и AV узлам, а также дополнительные волокна к предсердиям и желудочкам. Желудочки больше иннервируются симпатическими волокнами, чем парасимпатическими волокнами. Симпатическая стимуляция вызывает высвобождение нейромедиатора. норэпинефрин (также известный как норадреналин ) на нервномышечное соединение сердечных нервов. Это сокращает период реполяризации, тем самым ускоряя скорость деполяризации и сокращения, что приводит к увеличению частоты сердечных сокращений. Он открывает химические или управляемые лигандами каналы для ионов натрия и кальция, обеспечивая приток положительно заряженных ионов.[1] Норэпинефрин связывается с рецептором бета – 1. Высокое кровяное давление лекарства используются, чтобы заблокировать эти рецепторы и таким образом уменьшить частоту сердечных сокращений.[1]
Сердечно-сосудистые центры получают входные данные от ряда висцеральных рецепторов с импульсами, проходящими через висцеральные сенсорные волокна внутри блуждающего нерва и симпатические нервы через сердечное сплетение. Среди этих рецепторов есть различные проприорецепторы, барорецепторы, и хеморецепторы, плюс стимулы от лимбическая система которые обычно позволяют точно регулировать сердечную функцию посредством сердечных рефлексов. Повышенная физическая активность приводит к увеличению скорости возбуждения различных проприорецепторов, расположенных в мышцах, суставных капсулах и сухожилиях. Сердечно-сосудистые центры контролируют эту повышенную частоту возбуждения, подавляя парасимпатическую стимуляцию или увеличивая симпатическую стимуляцию по мере необходимости, чтобы увеличить кровоток.[1]
Точно так же барорецепторы - это рецепторы растяжения, расположенные в синусе аорты, каротидных телах, полых венах и других местах, включая легочные сосуды и правую сторону самого сердца. Скорость срабатывания барорецепторов отражает артериальное давление, уровень физической активности и относительное распределение крови. Сердечные центры контролируют срабатывание барорецепторов для поддержания сердечного гомеостаза - механизма, называемого барорецепторным рефлексом. При увеличении давления и растяжения увеличивается скорость активации барорецепторов, а сердечные центры уменьшают симпатическую стимуляцию и увеличивают парасимпатическую стимуляцию. По мере уменьшения давления и растяжения скорость активации барорецепторов уменьшается, а сердечные центры усиливают симпатическую стимуляцию и уменьшают парасимпатическую стимуляцию.[1]
Есть аналогичный рефлекс, называемый предсердным рефлексом или Рефлекс Бейнбриджа, связанный с различной скоростью кровотока в предсердиях. Повышенный венозный возврат растягивает стенки предсердий, в которых расположены специализированные барорецепторы. Однако по мере того, как предсердные барорецепторы увеличивают частоту возбуждения и растягиваются из-за повышенного кровяного давления, сердечный центр реагирует усилением симпатической стимуляции и подавлением парасимпатической стимуляции для увеличения ЧСС. Обратное тоже верно.[1]
Факторы, влияющие на частоту сердечных сокращений
В добавок к автономная нервная система, на это могут повлиять другие факторы. К ним относятся адреналин, норэпинефрин и гормоны щитовидной железы; уровни различных ионов, включая кальций, калий и натрий; температура тела; гипоксия; и баланс pH.[1]
| ||||||||||||||||||||||||||
|
Факторы, увеличивающие частоту сердечных сокращений, также вызывают увеличение ударного объема. Как и в случае со скелетными мышцами, при упражнениях сердце может увеличиваться в размерах и увеличиваться в объеме.[1] Таким образом, спортсмены на выносливость, такие как марафонцы может иметь сердце, у которого есть гипертрофированный до 40%.[3]:1063–64 Разница между максимальным и минимальным сердечным выбросами называется сердечным резервом и измеряет остаточную способность перекачивать кровь.[1] Частота сердечных сокращений может достигать 185–195 во время упражнений, в зависимости от того, насколько человек физически подготовлен.[3]
Сердечный выброс

Сердечный выброс (СО) - это измерение количества крови, перекачиваемой каждым желудочком (ударный объем, УЗ) за одну минуту. Чтобы рассчитать это, умножьте ударный объем (SV) на частоту сердечных сокращений (HR) в Ударов в минуту.[1] Это может быть представлено уравнением: CO = HR x SV[1]
SV обычно измеряется с помощью эхокардиограмма для записи конечного диастолического объема (EDV) и конечного систолического объема (ESV) и вычисления разницы: SV = EDV - ESV. SV также можно измерить с помощью специализированного катетера, но это инвазивная процедура и гораздо более опасна для пациента. Средняя SV для отдыхающего человека весом 70 кг (150 фунтов) будет примерно 70 мл. Существует несколько важных переменных, включая размер сердца, физическое и психическое состояние человека, пол, сократимость, продолжительность сокращения, предварительную нагрузку или EDV, а также постнагрузку или сопротивление. Нормальный диапазон для SV составляет 55–100 мл. Средняя ЧСС в состоянии покоя составляет примерно 75 ударов в минуту, но у некоторых людей может колебаться в пределах 60–100.[1] Используя эти числа (которые относятся к каждому желудочку, а не к обоим), среднее значение CO составляет 5,25 л / мин с диапазоном 4,0–8,0 л / мин.[1]

SV также используются для расчета фракции выброса, которая представляет собой часть крови, которая перекачивается или выбрасывается из сердца при каждом сокращении. Для расчета фракции выброса SV делится на EDV. Несмотря на название, фракция выброса обычно выражается в процентах. Фракция выброса составляет примерно 55–70 процентов, в среднем 58 процентов.[1]
Ударный объем


Многие факторы, регулирующие частоту сердечных сокращений, также влияют на сердечную функцию, изменяя ударный объем. Хотя здесь задействован ряд переменных, ударный объем зависит от разницы между конечным диастолическим объемом и конечным систолическим объемом. Три основных фактора: предварительная нагрузка, последующая нагрузка и сократимость.[1]
Предварительная загрузка
Предварительная загрузка это еще один способ выражения EDV. Следовательно, чем больше EDV, тем больше предварительная нагрузка. Главный фактор - время наполнения желудочков. Чем быстрее происходит сокращение, тем короче время наполнения, а также ниже EDV и предварительная нагрузка.[1]
Взаимосвязь между растяжением и сокращением желудочков была указана в Механизм Франка-Старлинга который говорит, что сила сокращения прямо пропорциональна исходной длине мышечного волокна. Так что чем больше растяжение желудочка, тем сильнее сокращение. Любая симпатическая стимуляция венозной системы увеличивает венозный возврат к сердцу и наполнение желудочков.[1]
Afterload
Желудочки должны развивать определенное напряжение, чтобы перекачивать кровь, преодолевая сопротивление сосудистой системы. Это напряжение называется последующая нагрузка. Когда сопротивление увеличивается, особенно из-за стенозирующий При повреждении клапана постнагрузка обязательно должна возрасти. Также может наблюдаться снижение нормального сопротивления сосудов. Различные сердечные реакции восстанавливают гомеостаз давления и кровотока.[1]
Сократимость
Способность миокарда сокращаться (его сократимость ), управляет ударным объемом, который определяет конечный систолический объем. Чем больше сокращение, тем больше ударный объем и меньше конечный систолический объем. Положительным или отрицательным инотропные факторы посредством симпатической и парасимпатической стимуляции соответственно может увеличивать или уменьшать силу сокращений. Симпатическая стимуляция вызывает высвобождение норэпинефрина из сердечных нервов, а также стимулирует кора надпочечников секретировать и адреналин, и норадреналин. Эти выделения увеличивают частоту сердечных сокращений, скорость последующего метаболизма и сократительную способность. Парасимпатическая стимуляция стимулирует высвобождение ацетилхолин (АЧ) из блуждающий нерв который снижает сократительную способность, и ударный объем, который увеличивает конечный систолический объем.
Было разработано несколько синтетических препаратов, которые могут действовать как инотропный стимулятор или ингибитор. Стимуляторы инотропы, такие как Дигоксин, вызывают более высокие концентрации ионов кальция, которые увеличивают сократимость. Избыток кальция (гиперкальциемия ) также является положительным инотропом. К препаратам, которые являются отрицательными инотропами, относятся: бета-блокаторы и блокаторы кальциевых каналов. Гипоксия, ацидоз, гиперкалиемия также являются отрицательными инотропными агентами.
| |||||||||||||||
|
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в au средний ау топор ай az ба bb до н.э bd быть парень bg бх би Ъ bk бл бм млрд бо бп бк br bs bt бу bv чб bx к bz ок cb cc CD ce ср cg Беттс, Дж. Гордон (2013). Анатомия и физиология. С. 787–846. ISBN 1938168135. Получено 11 августа 2014.[1]
- ^ Андерсон, РМ. Общая физиология сердечно-сосудистой системы (2-е изд., 2012). См. «Глава 1: Нормальная физиология».
- ^ а б c d Холл, Артур С. Гайтон, Джон Э. (2005). Учебник медицинской физиологии (11-е изд.). Филадельфия: W.B. Сондерс. п. 106. ISBN 978-0-7216-0240-0.
- ^ Покок, Джиллиан (2006). Физиология человека (Третье изд.). Издательство Оксфордского университета. п. 85. ISBN 978-0-19-856878-0.
- ^ Анц, Матиас; и другие. (1998). «Электрическая проводимость между правым и левым предсердиями через мускулатуру коронарного синуса». Тираж. 98 (17): 1790–95. Дои:10.1161 / 01.CIR.98.17.1790. PMID 9788835.
- ^ Де Понти, Роберто; и другие. (2002). «Электроанатомический анализ распространения синусового импульса в нормальных предсердиях человека». Журнал сердечно-сосудистой электрофизиологии. 13 (1): 1–10. Дои:10.1046 / j.1540-8167.2002.00001.x. PMID 11843475.
- ^ «Определение узла SA - определения в медицинском словаре популярных медицинских терминов, легко определяемых на MedTerms». Medterms.com. 27 апреля 2011 г.. Получено 7 июн 2012.
- ^ «Волокна Пуркинье». Biology.about.com. 9 апреля 2012 г.. Получено 7 июн 2012.
- ^ а б c d Талли, Николас Дж .; О'Коннор, Саймон. Клиническое обследование. Черчилль Ливингстон. С. 76–82. ISBN 9780729541985.
- ^ Дорланда (2012). Иллюстрированный медицинский словарь Дорланда (32-е изд.). Elsevier Saunders. п. 1189. ISBN 978-1-4160-6257-8.
- ^ Ньюбургер, Джейн (2006). Детская кардиология Надас 2-е издание. Филадельфия: Эльзевьер. п. 358. ISBN 978-1-4160-2390-6.
- ^ Cantarini L, Lopalco G; и другие. (Октябрь 2014 г.). «Аутоиммунитет и аутовоспаление как инь и ян идиопатического рецидивирующего острого перикардита». Аутоиммунный Rev. 14: 90–97. Дои:10.1016 / j.autrev.2014.10.005. PMID 25308531.